
Genome-Wide Identification of the GhANN Gene Family and Functional Validation of GhANN11 and GhANN4 under Abiotic Stress
Abstract
:1. Introduction
2. Results
2.1. Identification and Physicochemical Properties of ANN Gene Family Members in Four Gossypium spp.
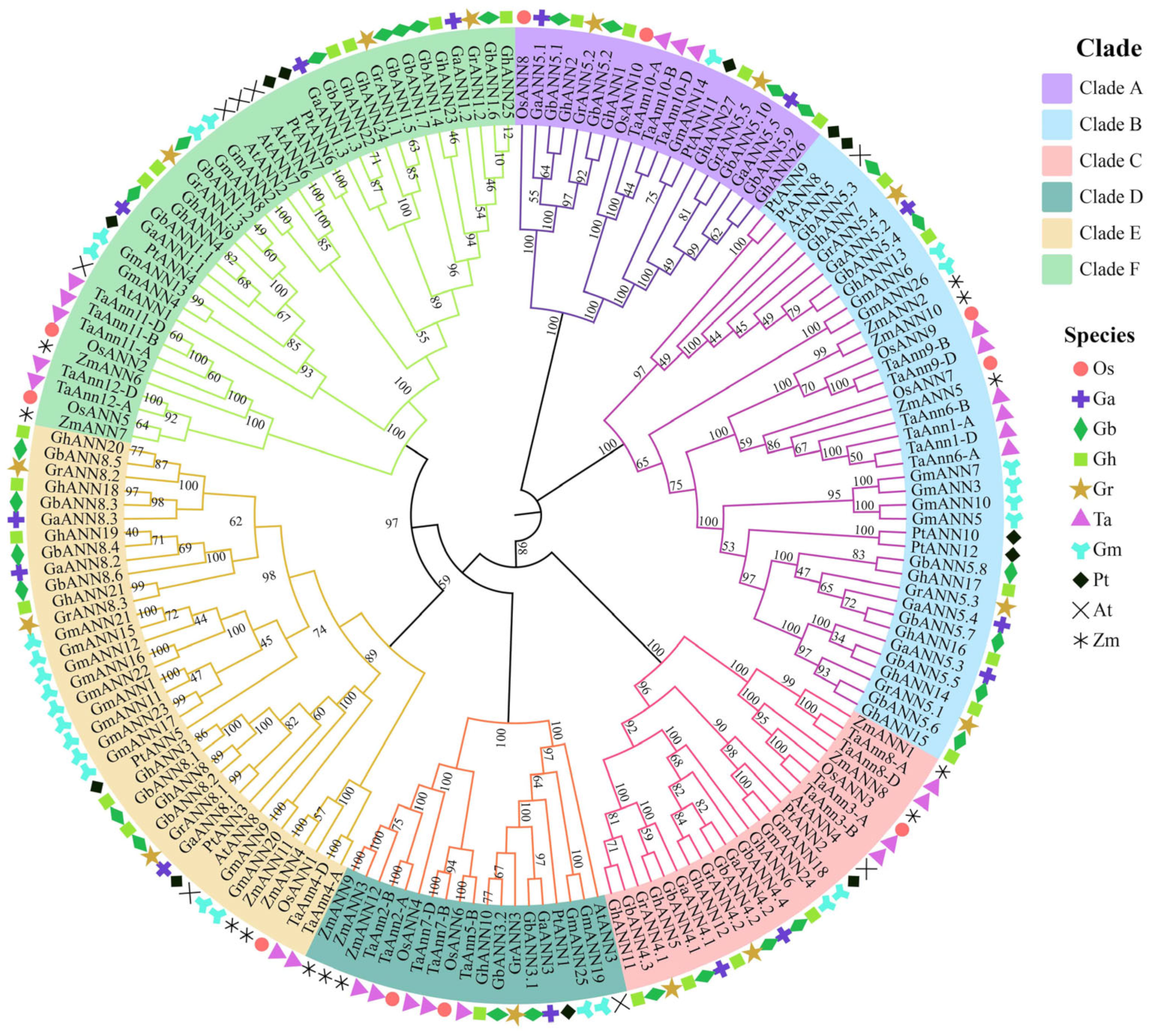
2.2. Phylogenetic Study of the ANN Gene Family Members
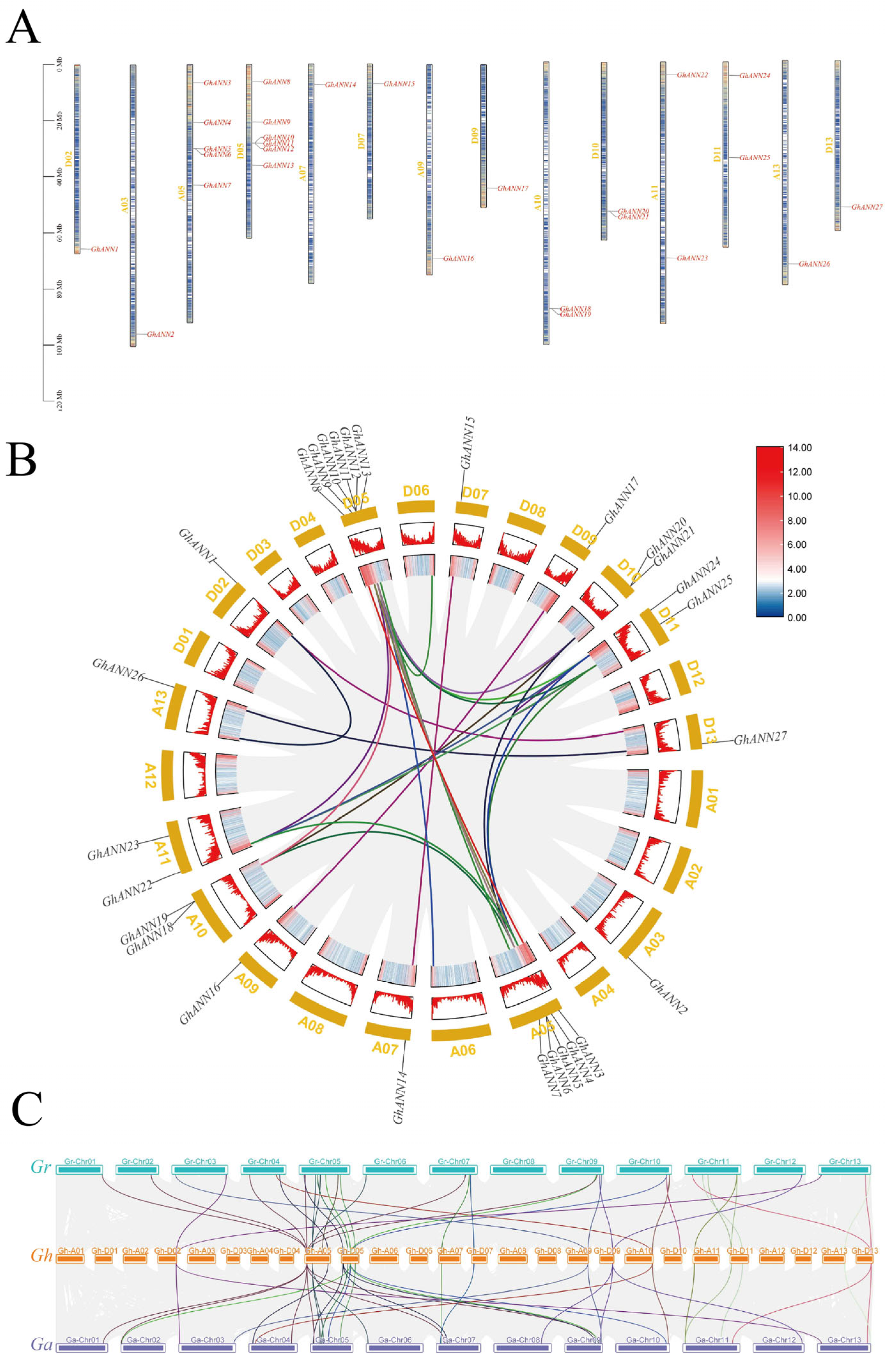
2.3. Distribution of Chromosomes, Intergenomic Interactions, and Collinear Relationships within the GhANN Family
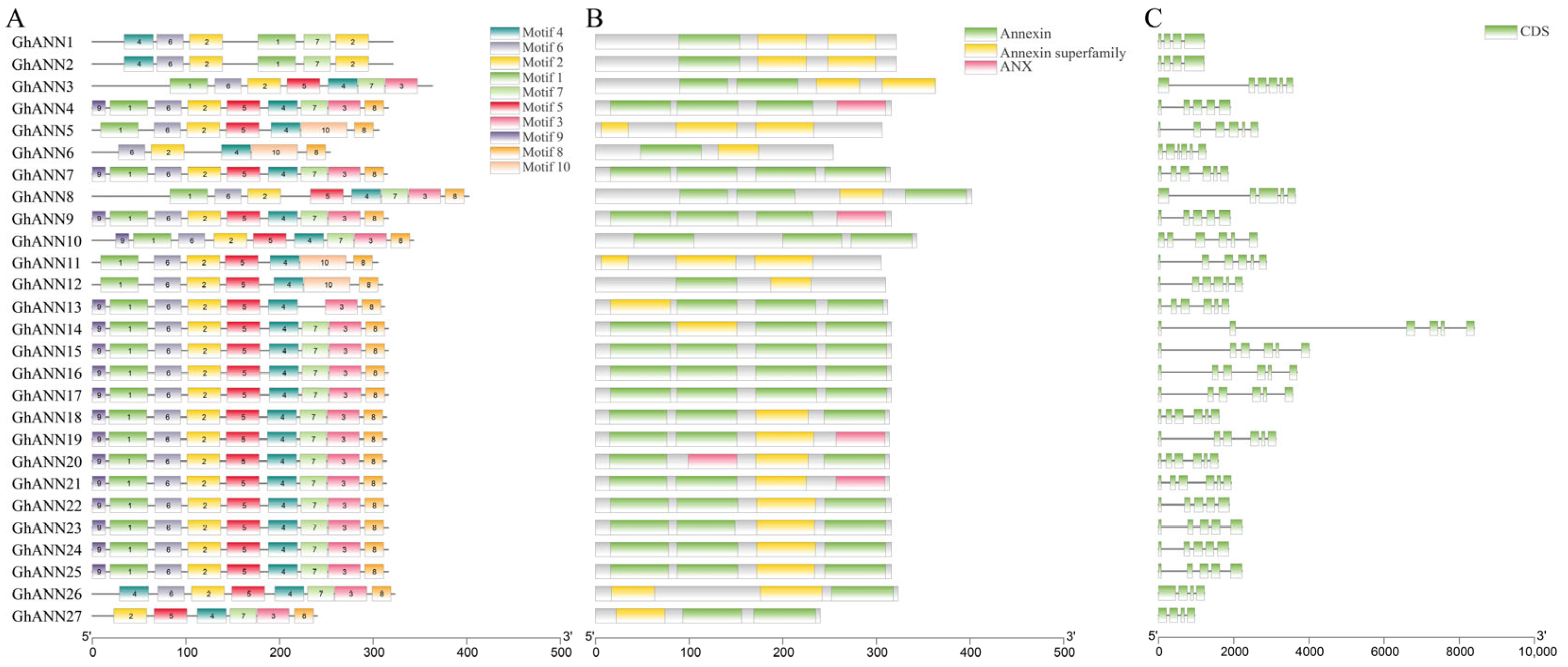
2.4. Conserved Polypeptide Motifs, Functional Domains, and Gene Structure of the ANN Family
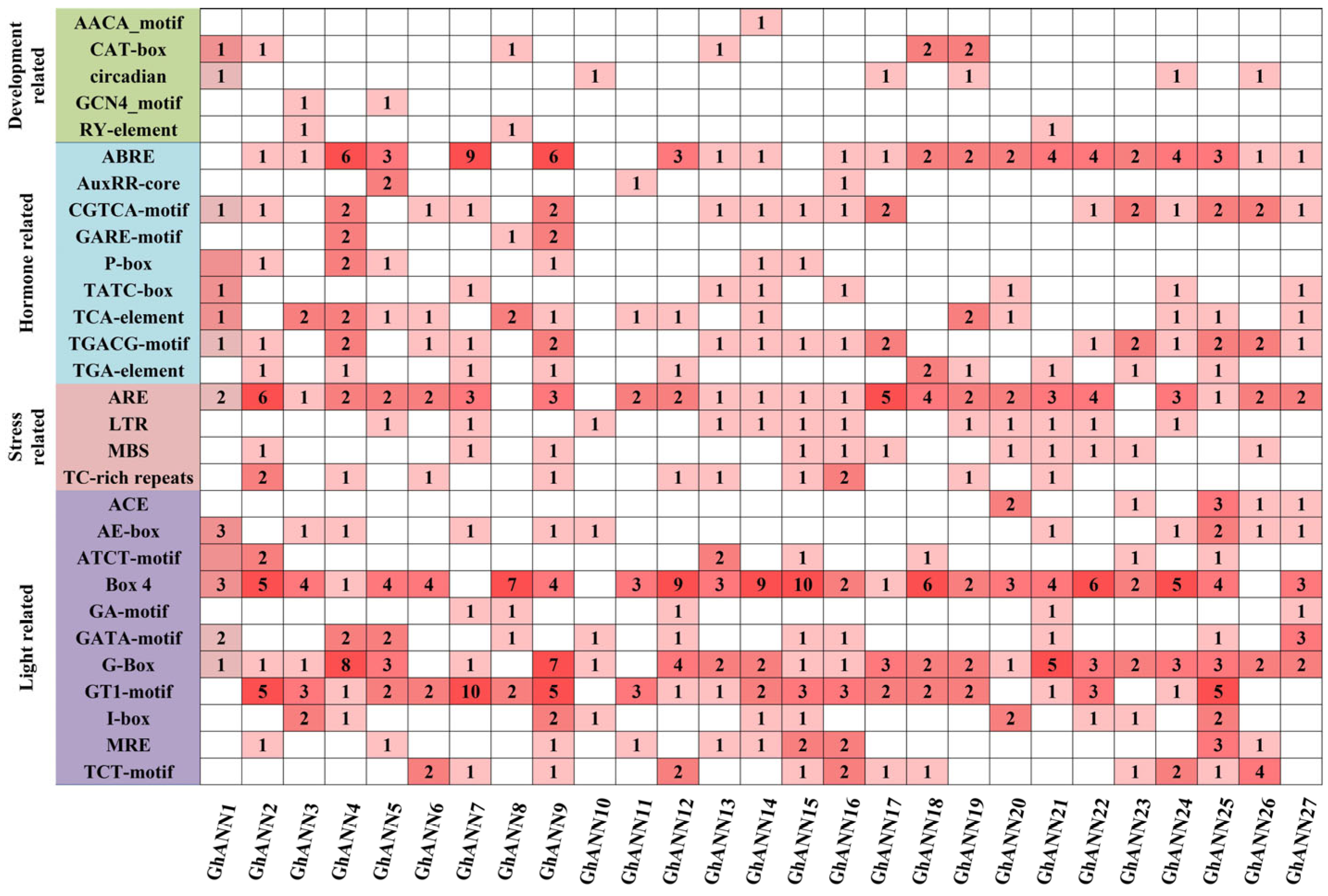
2.5. Examination of Cis-Acting Elements within the GhANN Gene Family
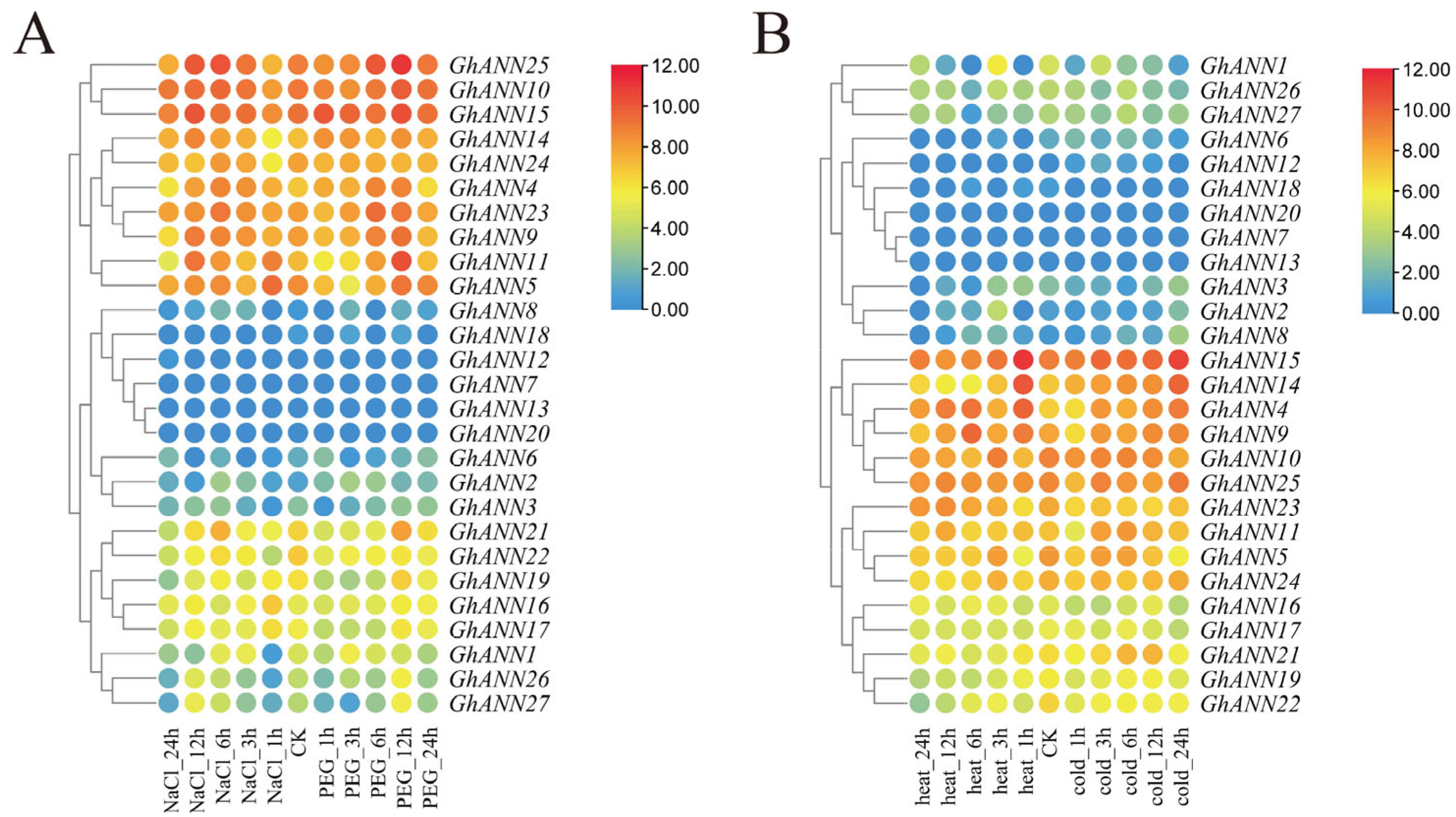
2.6. GhANN Gene Expression Patterns under Different Abiotic Stresses
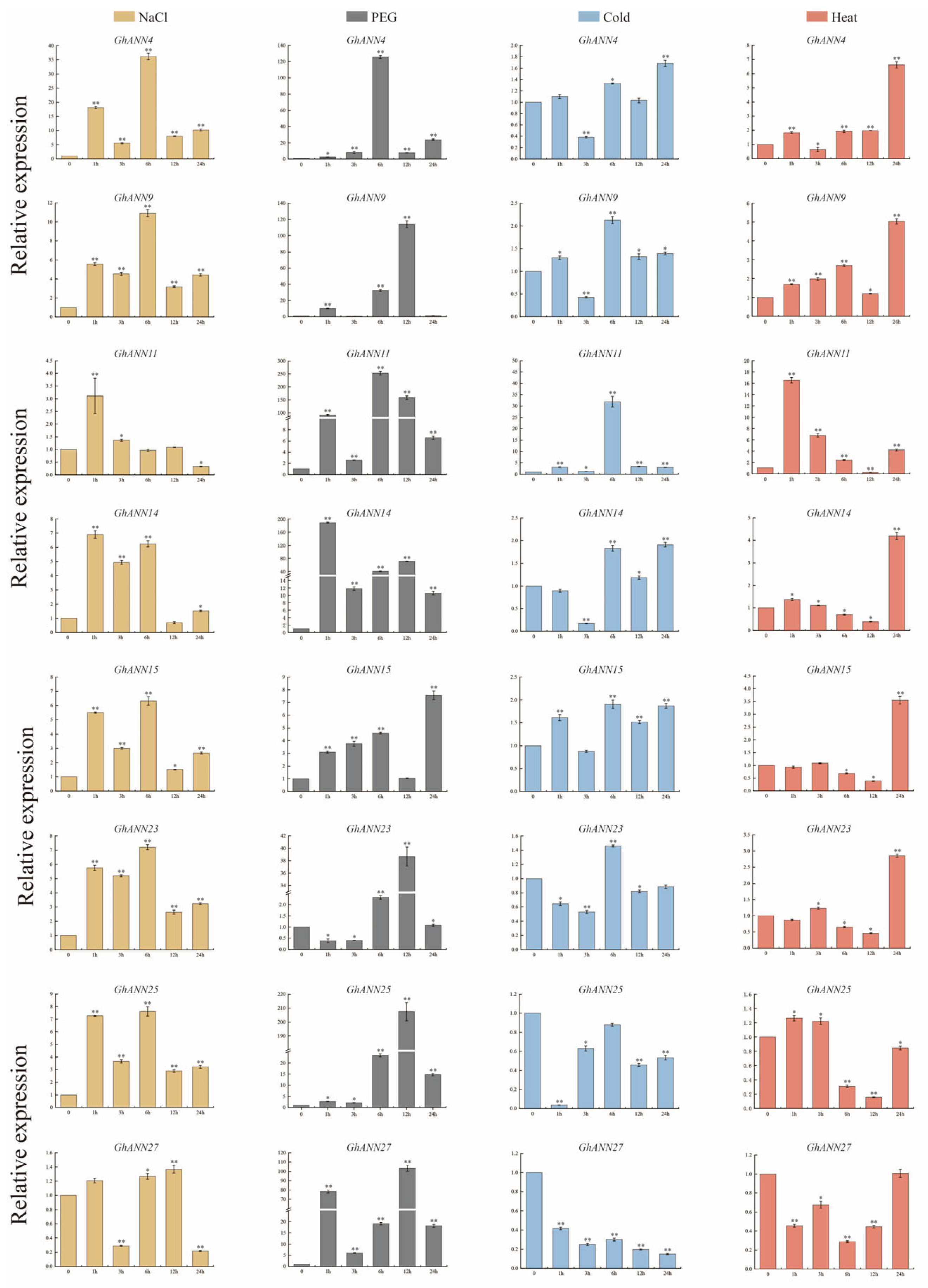
2.7. Validation of GhANN Gene Expression Patterns under Abiotic Stress
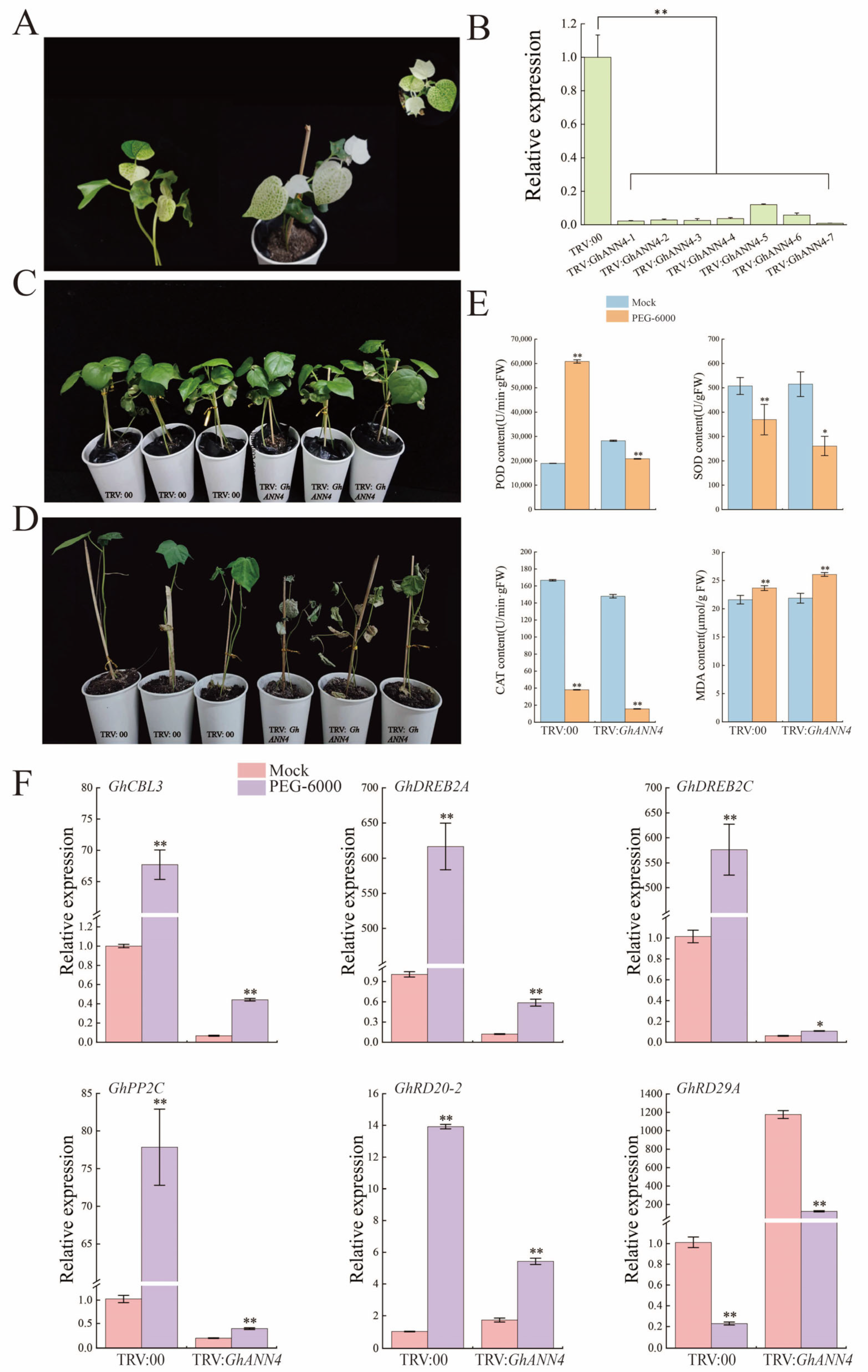
2.8. Silencing of GhANN4 Attenuated Drought and Salt Tolerance in Upland Cotton
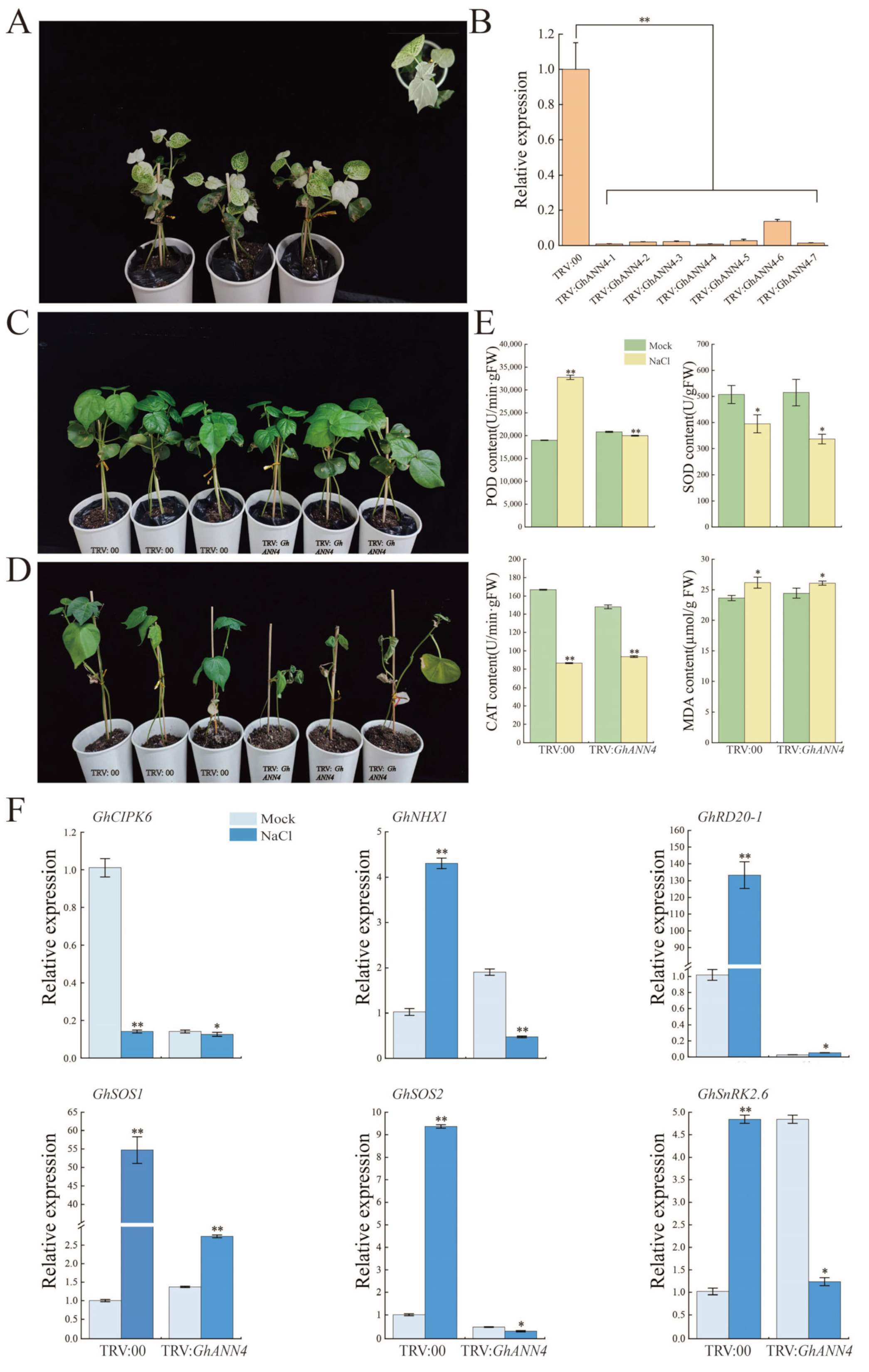
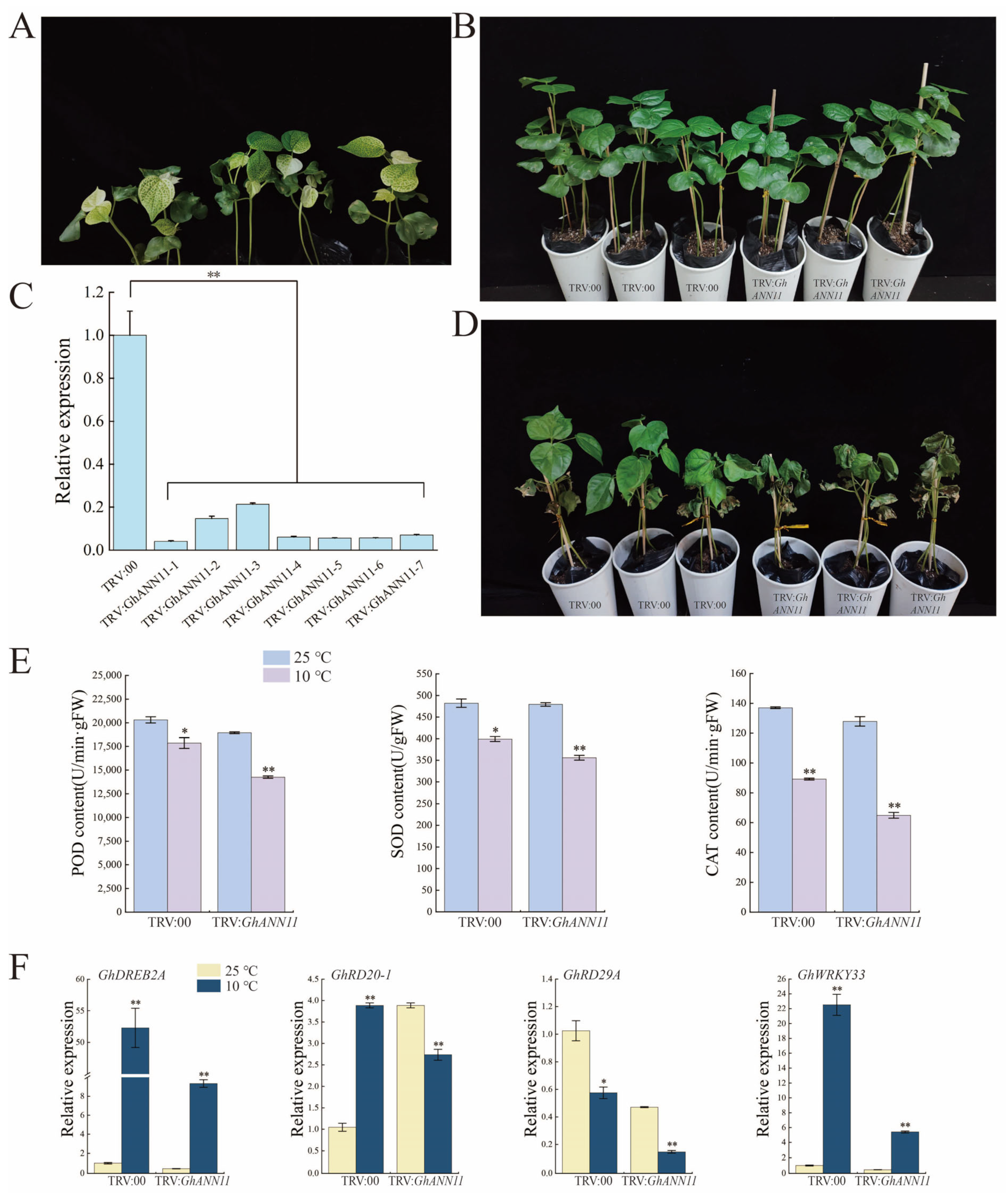
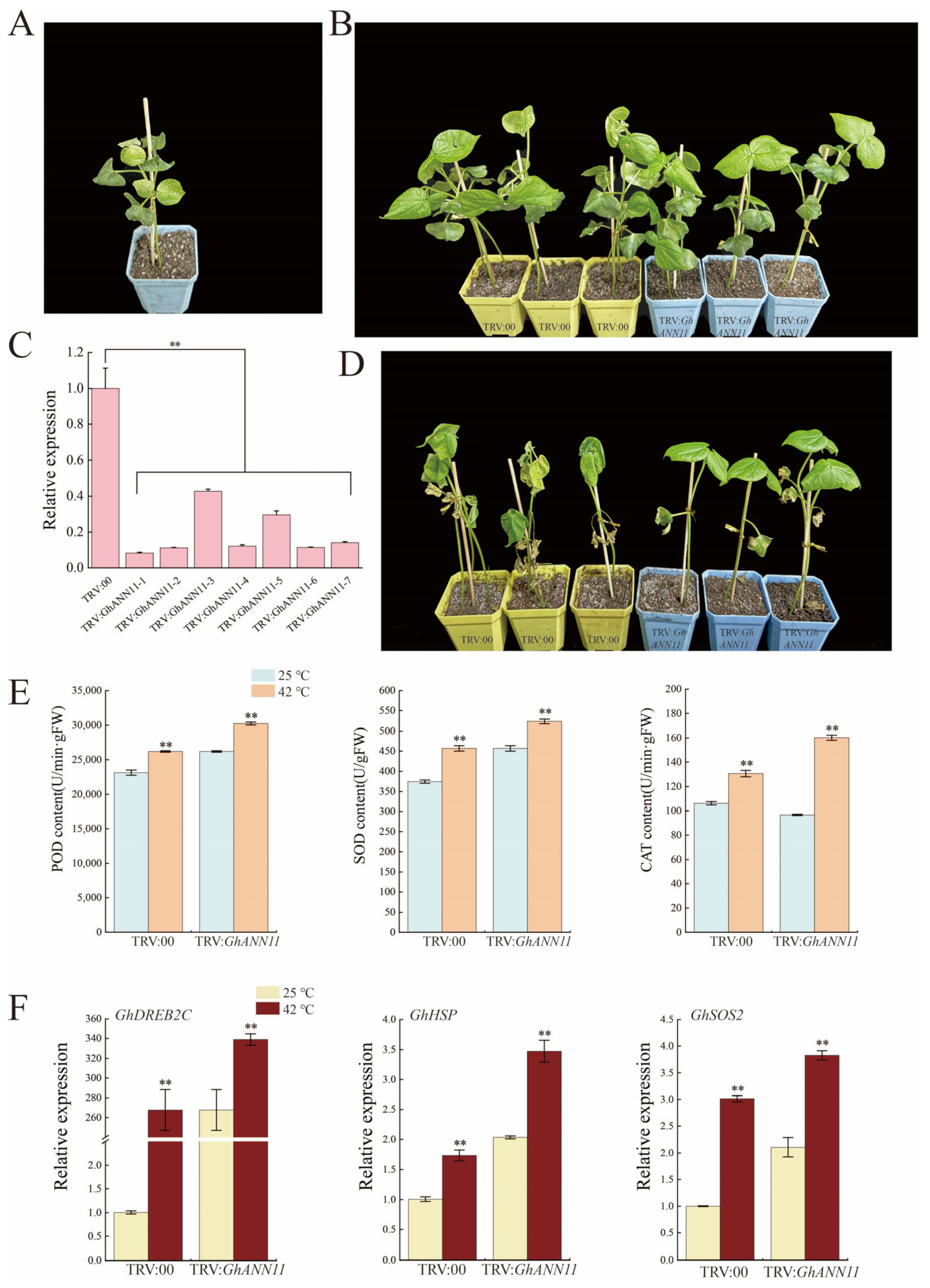
2.9. Effect of GhANN11 Gene Silencing on Cold Tolerance and Heat Resistance in Cotton
3. Discussion
4. Materials and Methods
4.1. Identification and Analysis of ANN Gene Family Members
4.2. Phylogenetic Tree Construction, Chromosome Mapping, and Collinearity Analysis of GhANN Family Genes
4.3. Examination of the GhANN Family Gene Structure and Conserved Motifs in Gossypium hirsutum
4.4. Prediction of Cis-Acting Regulatory Elements within GhANN Promoters
4.5. Analysis of the Gene Expression Characteristics of the GhANN Family in Gossypium hirsutum
4.6. Real-time Fluorescence-Based Quantitative PCR (qRT-PCR) of Selected GhANNs
4.7. VIGS of GhANN4 and GhANN11 in G. hirsutum
4.8. Salt, Drought, Cold, and Heat Resistance Stress Treatment
4.9. SOD, POD, and CAT Activity and MDA Content Determination
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, N.; Shang, Z.L. Annexins in Plant Cells. Plant. Physiol. Commun. 2010, 46, 277–283. [Google Scholar] [CrossRef]
- Creutz, C.E.; Pazoles, C.J.; Pollard, H.B. Identification and purification of an adrenal medullary protein (synexin) that causes calcium-dependent aggregation of isolated chromaffin granules. J. Biol. Chem. 1978, 253, 2858–2866. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.L.; Han, Z.Y.; Liu, R.Q.; Xue-Lei, Q.; Zhi-Ying, H.; Riu-Qi, L.; Chang-Long, Y.U.; Hai-Fang, Y.; Yu-Hua, L.I. Annexins:multi-functional complex in the process of plant growth. Guihaia 2013, 33, 703–709. [Google Scholar]
- Boustead, C.M.; Smallwood, M.; Small, H.; Bowles, D.J.; Walker, J.H. Identification of calcium-dependent phospholipid-binding proteins in higher plant cells. FEBS Lett. 1989, 244, 456–460. [Google Scholar] [CrossRef]
- Carrasco-Castilla, J.; Ortega-Ortega, Y.; Jauregui-Zuniga, D.; Juarez-Verdayes, M.A.; Arthikala. Down-regulation of a Phaseolus vulgaris annexin impairs rhizobial infection and nodulation. Environ. Exp. Bot. 2018, 153, 108–119. [Google Scholar] [CrossRef]
- Ijaz, R.; Ejaz, J.; Gao, S.; Liu, T.; Imtiaz, M.; Ye, Z.; Wang, T. Overexpression of annexin gene AnnSp2, enhances drought and salt tolerance through modulation of ABA synthesis and scavenging ROS in tomato. Sci. Rep. 2017, 7, 12087. [Google Scholar] [CrossRef]
- Clark, G.B.; Dauwalder, M.; Roux, S.J. Immunolocalization of an annexin-like protein in corn. Adv. Space Res. 1994, 14, 341–3466. [Google Scholar] [CrossRef]
- Clark, G.B.; Turnwald, S.; Tirlapur, U.K.; Haas, C.J.; von der Mark, K.; Roux, S.J.; Scheuerlein, R. Polar distribution of annexin-like proteins during phytochrome-mediated initiation and growth of rhizoids in the ferns Dryopteris and Anemia. Planta 1995, 197, 376–384. [Google Scholar] [CrossRef]
- Clark, G.B.; Lee, D.; Dauwalder, M.; Roux, S.J. Immunolocalization and histochemical evidence for the association of two different Arabidopsis annexins with secretion during early seedling growth and development. Planta 2005, 220, 621–631. [Google Scholar] [CrossRef]
- Xu, L.; Tang, Y.; Gao, S.; Su, S.; Hong, L.; Wang, W.; Fang, Z.; Li, X.; Ma, J.; Quan, W.; et al. Comprehensive analyses of the annexin gene family in wheat. BMC Genomics 2016, 17, 415. [Google Scholar] [CrossRef]
- Lichocka, M.; Rymaszewski, W.; Morgiewicz, K.; Barymow-Filoniuk, I.; Chlebowski, A.; Sobczak, M.; Samuel, M.A.; Schmelzer, E.; Krzymowska, M.; Hennig, J. Nucleus- and plastid-targeted annexin 5 promotes reproductive development in Arabidopsis and is essential for pollen and embryo formation. BMC Plant Biol. 2018, 18, 183–198. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mao, L.; Mi, H.; Lu, W.; Ying, T.; Luo, Z. Involvement of three annexin genes in the ripening of strawberry fruit regulated by phytohormone and calcium signal transduction. Plant Cell Rep. 2016, 35, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Gao, J.; Mu, J.; Duan, L.; Gu, Y.; Han, S.; Chen, L.; Li, Y.; Yan, Y.; Li, X. Interaction between serine carboxypeptidase-like protein TtGS5 and Annexin D1 in developing seeds of Triticum timopheevi. J. Appl. Genet. 2020, 61, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, D.; Hayashi, A.; Temmei, Y.; Kanzawa, N.; Tsuchiya, T. Biochemical and immuno-histochemical characterization of Mimosa annexin. Planta 2004, 219, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Gidrol, X.; Sabelli, P.A.; Fern, Y.S.; Kush, A.K. Annexin-like protein from Arabidopsis. thaliana rescues delta oxyR mutant of Escherichia coli from H2O2 stress. Proc. Natl. Acad. Sci. USA 1996, 93, 11268–11273. [Google Scholar] [CrossRef]
- He, B.; Cai, Q.; Qiao, L.; Huang, C.Y.; Wang, S.; Miao, W.; Ha, T.; Wang, Y.; Jin, H. RNA-binding proteins contribute to small RNA loading in plant extracellular vesicles. Nat. Plants 2021, 7, 342–352. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.X.; Xu, Y.J.; Lei, Y.; Li, Q.; Zhao, J.Q.; Li, Y.; Fan, J.; Xiao, S.; Wang, W.M. ANNEXIN 8 negatively regulates RPW8.1-mediated cell death and disease resistance in Arabidopsis. J. Integr. Plant. Biol. 2021, 63, 378–392. [Google Scholar] [CrossRef] [PubMed]
- Vandeputte, O.; Lowe, Y.O.; Burssens, S.; Van Raemdonck, D.; Hutin, D.; Boniver, D.; Geelen, D.; El Jaziri, M.; Baucher, M. The tobacco Ntann12 gene, encoding an annexin, is induced upon Rhodoccocus fascians infection and during leafy gall development. Mol. Plant Pathol. 2007, 8, 185–194. [Google Scholar] [CrossRef]
- Deka, S.; Barthakur, S. Impact of biorational pesticides and response of annexin gene expression in tomato (Solanum lycopersicum) and capsicum (Capsicum annum L.) in biotic stress management under greenhouse condition. Ann. Agric. Res. New Ser. 2016, 36, 233–242. [Google Scholar]
- Yadav, D.; Boyidi, P.; Ahmed, I.; Kirti, P.B. Plant annexins and their involvement in stress responses. Environ. Exp. Bot. 2018, 155, 293–306. [Google Scholar] [CrossRef]
- Jami, S.K.; Clark, G.B.; Turlapati, S.A.; Handley, C.; Roux, S.J.; Kirti, P.B. Ectopic expression of an annexin from Brassica. juncea confers tolerance to abiotic and biotic stress treatments in transgenic tobacco. Plant Physiol. Biochem. 2008, 46, 1019–1030. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, Q.; Zhang, X.; Liu, X.; Wang, P.; Hou, Y. Cloning and Characterization of an Annexin Gene from Cynanchum komarovii that Enhances Tolerance to Drought and Fusarium oxysporum in Transgenic Cotton. J. Plant Biol. 2011, 54, 303–313. [Google Scholar] [CrossRef]
- Zhao, J.; Li, L.; Liu, Q.; Liu, P.; Li, S.; Yang, D.; Chen, Y.; Pagnotta, S.; Favery, B.; Abad, P.; et al. A MIF-like effector suppresses plant immunity and facilitates nematode parasitism by interacting with plant annexins. J. Exp. Bot. 2019, 70, 5943–5958. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Lee, E.J.; Yang, E.J.; Lee, J.E.; Park, A.R.; Song, W.H.; Park, O.K. Proteomic identification of annexins, calcium-dependent membrane binding proteins that mediate osmotic stress and abscisic acid signal transduction in Arabidopsis. Plant Cell 2004, 16, 1378–1391. [Google Scholar] [PubMed]
- Huh, S.M.; Noh, E.K.; Kim, H.G.; Jeon, B.W.; Bae, K.; Hu, H.C.; Kwak, J.M.; Park, O.K. Arabidopsis annexins AnnAt1 and AnnAt4 interact with each other and regulate drought and salt stress responses. Plant Cell Physiol. 2010, 51, 1499–1514. [Google Scholar] [CrossRef]
- Wang, X.; Movahedi, A.; Wei, H.; Wu, X.; Zhuge, Q. Overexpression of PtAnnexin1 from Populus trichocarpa enhances salt and drought tolerance in transgenic poplars. Tree Genet. Genomes 2020, 16, 20–32. [Google Scholar] [CrossRef]
- Ahmed, I.; Yadav, D.; Shukla, P.; Vineeth, T.V.; Sharma, P.C.; Kirti, P.B. Constitutive expression of Brassica. juncea annexin, AnnBj2 confers salt tolerance and glucose and ABA insensitivity in mustard transgenic plants. Plant Sci. 2017, 265, 12–28. [Google Scholar] [CrossRef]
- Ahmed, I.; Yadav, D.; Shukla, P.; Kirti, P.B. Heterologous expression of Brassica juncea annexin, AnnBj2 confers salt tolerance and ABA insensitivity in transgenic tobacco seedlings. Funct. Integr. Genomics 2018, 18, 569–579. [Google Scholar] [CrossRef]
- Zhang, F.; Li, S.; Yang, S.; Wang, L.; Guo, W. Overexpression of a cotton annexin gene, GhAnn1, enhances drought and salt stress tolerance in transgenic cotton. Plant Mol. Biol. 2015, 87, 47–67. [Google Scholar] [CrossRef]
- Zhang, D.; Li, J.; Niu, X.; Deng, C.; Guo, W. GhANN1 modulates the salinity tolerance by regulating ABA biosynthesis, ion homeostasis and phenylpropanoid pathway in cotton. Environ. Exp. Bot. 2021, 185, 104427. [Google Scholar] [CrossRef]
- Mu, C.; Zhou, L.; Shan, L.; Li, F.; Li, Z. Phosphatase GhDsPTP3a interacts with annexin protein GhANN8b to reversely regulate salt tolerance in cotton (Gossypium spp.). New Phytol. 2019, 223, 1856–1872. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Q.; Yang, X.; Han, J.; Zhu, Z. OsANN3, a calcium-dependent lipid binding annexin is a positive regulator of ABA-dependent stress tolerance in rice. Plant Sci. 2019, 284, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Song, T.; Han, J.; He, M.; Zhang, Q.; Zhu, Y.; Zhu, Z. A calcium-dependent lipid binding protein, OsANN10, is a negative regulator of osmotic stress tolerance in rice. Plant Sci. 2020, 293, 110420. [Google Scholar] [CrossRef] [PubMed]
- Breton, G.; Vazquez-Tello, A.; Danyluk, J.; Sarhan, F. Two novel intrinsic annexins accumulate in wheat membranes in response to low temperature. Plant Cell Physiol. 2000, 41, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Ding, Y.; Shi, Y.; Ma, L.; Wang, Y.; Song, C.; Wilkins, K.A.; Davies, J.M.; Knight, H.; Knight, M.R.; et al. The calcium transporter ANNEXIN1 mediates cold-induced calcium signaling and freezing tolerance in plants. Embo J. 2021, 40, e104559. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Que, Z.; Xia, Y.; Tang, N.; Li, D.; He, R.; Cao, M. Knock out of the annexin gene OsAnn3 via CRISPR/Cas9-mediated genome editing decreased cold tolerance in rice. J. Plant Biol. 2017, 60, 539–547. [Google Scholar] [CrossRef]
- He, F.; Gao, C.; Guo, G.; Liu, J.; Gao, Y.; Pan, R.; Guan, Y.; Hu, J. Maize annexin genes ZmANN33 and ZmANN35 encode proteins that function in cell membrane recovery during seed germination. J. Exp. Bot. 2019, 70, 1183–1195. [Google Scholar] [CrossRef]
- Chu, P.; Chen, H.; Zhou, Y.; Li, Y.; Ding, Y.; Jiang, L.; Tsang, E.W.; Wu, K.; Huang, S. Proteomic and functional analyses of Nelumbo nucifera annexins involved in seed thermotolerance and germination vigor. Planta 2012, 235, 1271–1288. [Google Scholar] [CrossRef]
- Wang, X.; Ma, X.; Wang, H.; Li, B.; Clark, G.; Guo, Y.; Roux, S.; Sun, D.; Tang, W. Proteomic study of microsomal proteins reveals a key role for Arabidopsis annexin 1 in mediating heat stress-induced increase in intracellular calcium levels. Mol. Cell Proteomics 2015, 14, 686–694. [Google Scholar] [CrossRef]
- Liao, C.; Zheng, Y.; Guo, Y. MYB30 transcription factor regulates oxidative and heat stress responses through ANNEXIN-mediated cytosolic calcium signaling in Arabidopsis. New Phytol. 2017, 216, 163–177. [Google Scholar] [CrossRef]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; et al. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Brown, R.M., Jr. GTPase activity and biochemical characterization of a recombinant cotton fiber annexin. Plant Physiol. 1999, 119, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; He, Y.; Tu, L.; Wang, M.; Li, Y.; Ruan, Y.L.; Zhang, X. Down-regulating annexin gene GhAnn2 inhibits cotton fiber elongation and decreases Ca2+ influx at the cell apex. Plant Mol. Biol. 2014, 85, 613–625. [Google Scholar] [CrossRef]
- Guruprasad, K.; Reddy, B.V.; Pandit, M.W. Correlation between stability of a protein and its dipeptide composition: A novel approach for predicting in vivo stability of a protein from its primary sequence. Protein Eng. 1990, 4, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Clark, G.B.; Sessions, A.; Eastburn, D.J.; Roux, S.J. Differential expression of members of the annexin multigene family in Arabidopsis. Plant Physiol. 2001, 126, 1072–1084. [Google Scholar] [CrossRef]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Roux, S.J.; Kirti, P.B. Identification and characterization of annexin gene family in rice. Plant Cell Rep. 2012, 31, 813–825. [Google Scholar] [CrossRef]
- Feng, Y.M.; Wei, X.K.; Liao, W.X.; Huang, L.H.; Zhang, H.; Liang, S.C.; Peng, H. Molecular analysis of the annexin gene family in soybean. Biologia Plantarum 2013, 57, 655–662. [Google Scholar] [CrossRef]
- Chang, X.; He, X.; Li, J.; Liu, Z.; Pi, R.; Luo, X.; Wang, R.; Hu, X.; Lu, S.; Zhang, X.; et al. High-quality Gossypium hirsutum and Gossypium barbadense genome assemblies reveal the landscape and evolution of centromeres. Plant Commun. 2023, 22, 100722. [Google Scholar] [CrossRef]
- Harbaoui, M.; Ben Romdhane, W.; Ben Hsouna, A.; Brini, F.; Ben Saad, R. The durum wheat annexin, TdAnn6, improves salt and osmotic stress tolerance in Arabidopsis via modulation of antioxidant machinery. Protoplasma 2021, 258, 1047–1059. [Google Scholar] [CrossRef]
- Mushtaq, N.; Wang, Y.; Fan, J.; Li, Y.; Ding, J. Down-Regulation of Cytokinin Receptor Gene SlHK2 Improves Plant Tolerance to Drought, Heat, and Combined Stresses in Tomato. Plants 2022, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance [Review]. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide Dismutase and Stress Tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Kovács, I.; Ayaydin, F.; Oberschall, A.; Ipacs, I.; Bottka, S.; Pongor, S.; Dudits, D.; Tóth, E.C. Immunolocalization of a novel annexin-like protein encoded by a stress and abscisic acid responsive gene in alfalfa. Plant J. 1998, 15, 185–197. [Google Scholar] [CrossRef]
- Nakashima, K.; Shinwari, Z.K.; Sakuma, Y.; Seki, M.; Miura, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Organization and expression of two Arabidopsis DREB2 genes encoding DRE-binding proteins involved in dehydration- and high-salinity-responsive gene expression. Plant Mol. Biol. 2000, 42, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell 2006, 18, 1292–1309. [Google Scholar] [CrossRef] [PubMed]
- Xiu, Y.; Iqbal, A.; Zhu, C.; Wu, G.; Chang, Y.; Li, N.; Cao, Y.; Zhang, W.; Zeng, H.; Chen, S.; et al. Improvement and transcriptome analysis of root architecture by overexpression of Fraxinus pennsylvanica DREB2A transcription factor in Robinia pseudoacacia L. ‘Idaho’. Plant Biotechnol. J. 2016, 14, 1456–1469. [Google Scholar] [CrossRef]
- Je, J.; Chen, H.; Song, C.; Lim, C.O. Arabidopsis DREB2C modulates ABA biosynthesis during germination. Biochem. Biophys. Res. Commun. 2014, 452, 91–98. [Google Scholar] [CrossRef]
- Zhao, K.; Shen, X.; Yuan, H.; Liu, Y.; Liao, X.; Wang, Q.; Liu, L.; Li, F.; Li, T. Isolation and characterization of dehydration-responsive element-binding factor 2C (MsDREB2C) from Malus sieversii. Roem Plant Cell Physiol. 2013, 54, 1415–1430. [Google Scholar] [CrossRef]
- Aubert, Y.; Vile, D.; Pervent, M.; Aldon, D.; Ranty, B.; Simonneau, T.; Vavasseur, A.; Galaud, J.P. RD20, a stress-inducible caleosin, participates in stomatal control, transpiration and drought tolerance in Arabidopsis thaliana. Plant Cell Physiol. 2010, 51, 1975–1987. [Google Scholar] [CrossRef]
- Liu, J.; Ishitani, M.; Halfter, U.; Kim, C.S.; Zhu, J.K. The Arabidopsis thaliana SOS2 gene encodes a protein kinase that is required for salt tolerance. Proc. Natl. Acad. Sci. USA 2000, 97, 3730–3734. [Google Scholar] [CrossRef] [PubMed]
- Basso, M.F.; Costa, J.A.; Ribeiro, T.P.; Arraes, F.B.M.; Lourenço-Tessutti, I.T.; Macedo, A.F.; Neves, M.R.D.; Nardeli, S.M.; Arge, L.W.; Perez, C.E.A.; et al. Overexpression of the CaHB12 transcription factor in cotton (Gossypium hirsutum) improves drought tolerance. Plant Physiol. Biochem. 2021, 165, 80–93. [Google Scholar] [CrossRef]
- Shazadee, H.; Khan, N.; Wang, J.; Wang, C.; Zeng, J.; Huang, Z.; Wang, X. Identification and Expression Profiling of Protein Phosphatases (PP2C) Gene Family in Gossypium hirsutum L. Int. J. Mol. Sci. 2019, 20, 1395. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Zhang, G.; Sun, L.; Wang, J.; Hao, F. Genome-wide identification of CBL family and expression analysis of CBLs in response to potassium deficiency in cotton. PeerJ 2017, 5, e3653. [Google Scholar] [CrossRef] [PubMed]
- Long, L.; Zhao, J.R.; Guo, D.D.; Ma, X.N.; Xu, F.C.; Yang, W.W.; Gao, W. Identification of NHXs in Gossypium species and the positive role of GhNHX1 in salt tolerance. BMC Plant Biol. 2020, 20, 147. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.A.; Yang, G.D.; Meng, Q.W.; Zheng, C.C. The cotton GhNHX1 gene encoding a novel putative tonoplast Na(+)/H(+) antiporter plays an important role in salt stress. Plant Cell Physiol. 2004, 45, 600–607. [Google Scholar] [CrossRef]
- Chen, X.; Lu, X.; Shu, N.; Wang, D.; Wang, S.; Wang, J.; Guo, L.; Guo, X.; Fan, W.; Lin, Z.; et al. GhSOS1, a plasma membrane Na+/H+ antiporter gene from upland cotton, enhances salt tolerance in transgenic Arabidopsis thaliana. PLoS ONE 2017, 12, e0181450. [Google Scholar] [CrossRef]
- Che, B.; Cheng, C.; Fang, J.; Liu, Y.; Jiang, L.; Yu, B. The Recretohalophyte Tamarix TrSOS1 Gene Confers Enhanced Salt Tolerance to Transgenic Hairy Root Composite Cotton Seedlings Exhibiting Virus-Induced Gene Silencing of GhSOS1. Int. J. Mol. Sci. 2019, 20, 2930. [Google Scholar] [CrossRef]
- Lu, S.; Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Geer, R.C.; Gonzales, N.R.; Gwadz, M.; Hurwitz, D.I.; Marchler, G.H.; Song, J.S.; et al. Marchler-Bauer, A., CDD/SPARCLE: The conserved domain database in 2020. Nucleic Acids Res. 2020, 48, D265–D268. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wang, H.; Wei, H.; Gu, L.; Hao, P.; Sun, H.; Wu, A.; Cheng, S.; Yu, S. The MADS transcription factor GhAP1.7 coordinates the flowering regulatory pathway in upland cotton (Gossypium hirsutum L.). Gene 2021, 769, 145235. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Sun, H.; Hakim; Yang, X.; Zhang, X. A novel cotton WRKY gene, GhWRKY6-like, improves salt tolerance by activating the ABA signaling pathway and scavenging of reactive oxygen species. Physiol. Plant 2018, 162, 439–454. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Bharwana, S.A.; Ali, S.; Farooq, M.A.; Ali, B.; Iqbal, N.; Abbas, F.; Ahmad, M.S. Hydrogen sulfide ameliorates lead-induced morphological, photosynthetic, oxidative damages and biochemical changes in cotton. Environ. Sci. Pollut. Res. Int. 2014, 21, 717–731. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes Name | Genes ID | Number of Amino Acids (aa) | Molecular Weight (kDa) | Theoretical pI | The Instability Index | Grand Average of Hydropathicity (GRAVY) | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| GhANN1 | GH_D02G2140.1 | 321 | 36.53 | 9.71 | 42.78 | −0.307 | nucleus |
| GhANN2 | GH_A03G1973.1 | 321 | 36.37 | 9.75 | 42.37 | −0.312 | nucleus |
| GhANN3 | GH_A05G0751.1 | 363 | 41.14 | 6.65 | 42.58 | −0.425 | Golgi apparatus |
| GhANN4 | GH_A05G2275.1 | 316 | 35.97 | 6.39 | 35.10 | −0.474 | cytoplasm |
| GhANN5 | GH_A05G2824.1 | 306 | 34.70 | 6.88 | 45.71 | −0.395 | cytoskeleton |
| GhANN6 | GH_A05G2825.1 | 254 | 29.37 | 8.66 | 31.04 | −0.558 | cytoplasm |
| GhANN7 | GH_A05G3169.1 | 315 | 35.56 | 9.23 | 36.60 | −0.422 | cytoplasm |
| GhANN8 | GH_D05G0748.1 | 402 | 45.62 | 6.47 | 41.72 | −0.31 | plasma membrane |
| GhANN9 | GH_D05G2295.1 | 316 | 36.00 | 6.34 | 33.42 | −0.487 | cytoplasm |
| GhANN10 | GH_D05G2841.1 | 343 | 38.47 | 6.28 | 36.16 | −0.33 | peroxisome |
| GhANN11 | GH_D05G2842.1 | 305 | 34.60 | 6.33 | 44.93 | −0.372 | cytoskeleton |
| GhANN12 | GH_D05G2843.1 | 310 | 35.24 | 8.82 | 32.27 | −0.496 | cytoplasm |
| GhANN13 | GH_D05G3187.1 | 312 | 35.22 | 9.1 | 38.53 | −0.416 | cytoplasm |
| GhANN14 | GH_A07G0681.1 | 316 | 35.57 | 9.08 | 37.04 | −0.324 | cytoplasm |
| GhANN15 | GH_D07G0672.1 | 316 | 35.45 | 9.1 | 36.77 | −0.273 | cytoplasm |
| GhANN16 | GH_A09G1894.1 | 316 | 35.49 | 9.11 | 37.07 | −0.304 | chloroplast |
| GhANN17 | GH_D09G1846.1 | 316 | 35.57 | 9.12 | 37.39 | −0.327 | chloroplast |
| GhANN18 | GH_A10G2000.1 | 314 | 36.02 | 8.45 | 50.23 | −0.624 | cytoplasm |
| GhANN19 | GH_A10G2002.1 | 314 | 35.83 | 6.78 | 38.79 | −0.557 | cytoplasm |
| GhANN20 | GH_D10G2102.1 | 314 | 36.08 | 8.79 | 51.66 | −0.615 | cytoplasm |
| GhANN21 | GH_D10G2103.1 | 314 | 35.84 | 6.74 | 40.63 | −0.525 | cytoplasm |
| GhANN22 | GH_A11G0554.1 | 316 | 35.77 | 6.23 | 29.41 | −0.429 | cytoplasm |
| GhANN23 | GH_A11G2122.1 | 316 | 36.01 | 6.19 | 36.18 | −0.467 | cytoplasm |
| GhANN24 | GH_D11G0581.1 | 316 | 35.85 | 6.39 | 31.13 | −0.451 | cytoplasm |
| GhANN25 | GH_D11G2320.1 | 316 | 36.06 | 6.19 | 36.66 | −0.453 | cytoplasm |
| GhANN26 | GH_A13G1903.1 | 323 | 36.62 | 8.05 | 47.01 | −0.373 | cytoplasm |
| GhANN27 | GH_D13G1859.1 | 240 | 27.19 | 7.17 | 35.31 | −0.387 | cytoskeleton |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Li, M.; Ju, J.; Hai, H.; Wei, W.; Ling, P.; Li, D.; Su, J.; Zhang, X.; Wang, C. Genome-Wide Identification of the GhANN Gene Family and Functional Validation of GhANN11 and GhANN4 under Abiotic Stress. Int. J. Mol. Sci. 2024, 25, 1877. https://doi.org/10.3390/ijms25031877
Luo J, Li M, Ju J, Hai H, Wei W, Ling P, Li D, Su J, Zhang X, Wang C. Genome-Wide Identification of the GhANN Gene Family and Functional Validation of GhANN11 and GhANN4 under Abiotic Stress. International Journal of Molecular Sciences. 2024; 25(3):1877. https://doi.org/10.3390/ijms25031877
Chicago/Turabian StyleLuo, Jin, Meili Li, Jisheng Ju, Han Hai, Wei Wei, Pingjie Ling, Dandan Li, Junji Su, Xianliang Zhang, and Caixiang Wang. 2024. "Genome-Wide Identification of the GhANN Gene Family and Functional Validation of GhANN11 and GhANN4 under Abiotic Stress" International Journal of Molecular Sciences 25, no. 3: 1877. https://doi.org/10.3390/ijms25031877