
Ctp1 and Yhm2: Two Mitochondrial Citrate Transporters to Support Metabolic Flexibility of Saccharomyces cerevisiae


Abstract
:1. Introduction
2. Results
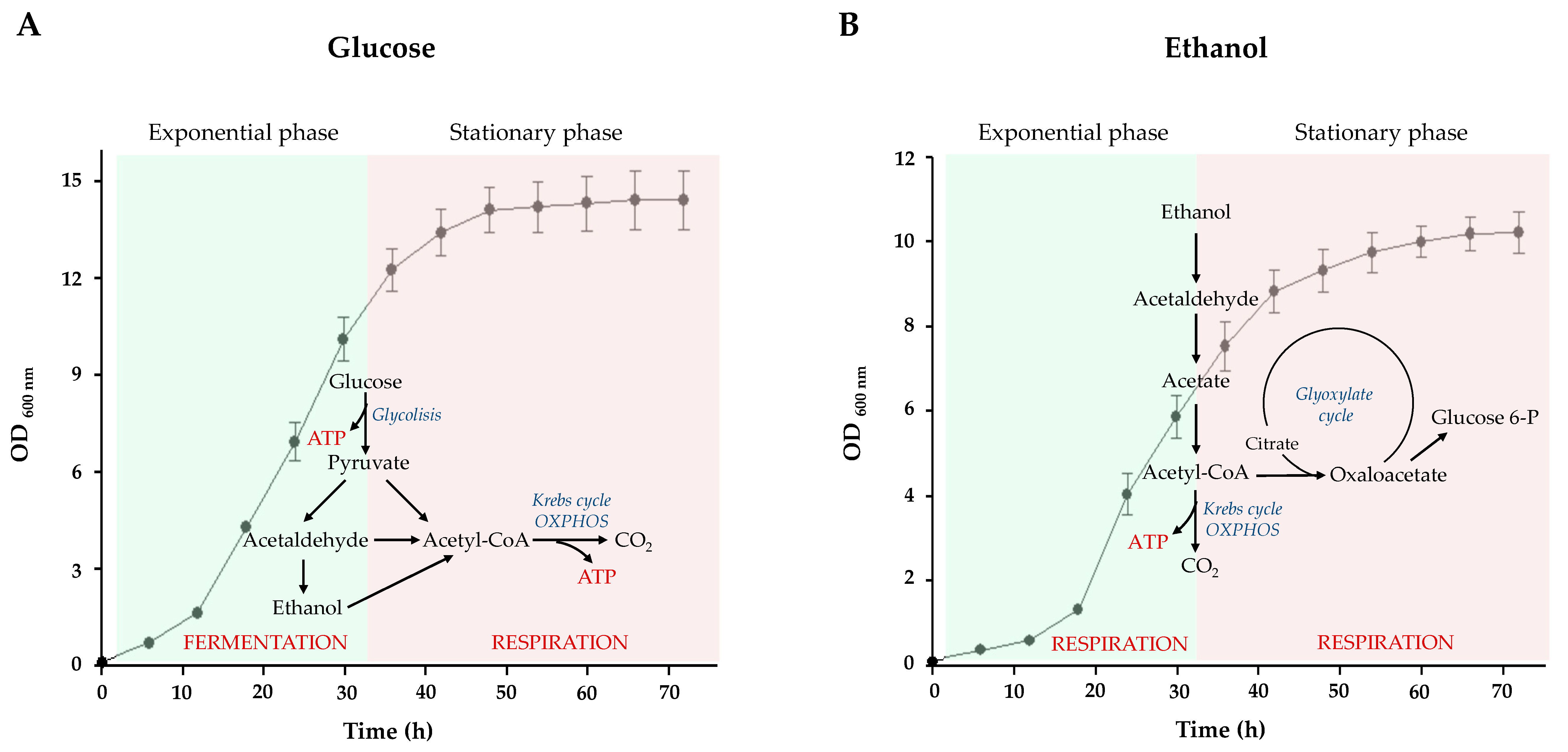
2.1. Glucose and Ethanol Metabolism during S. cerevisiae Growth
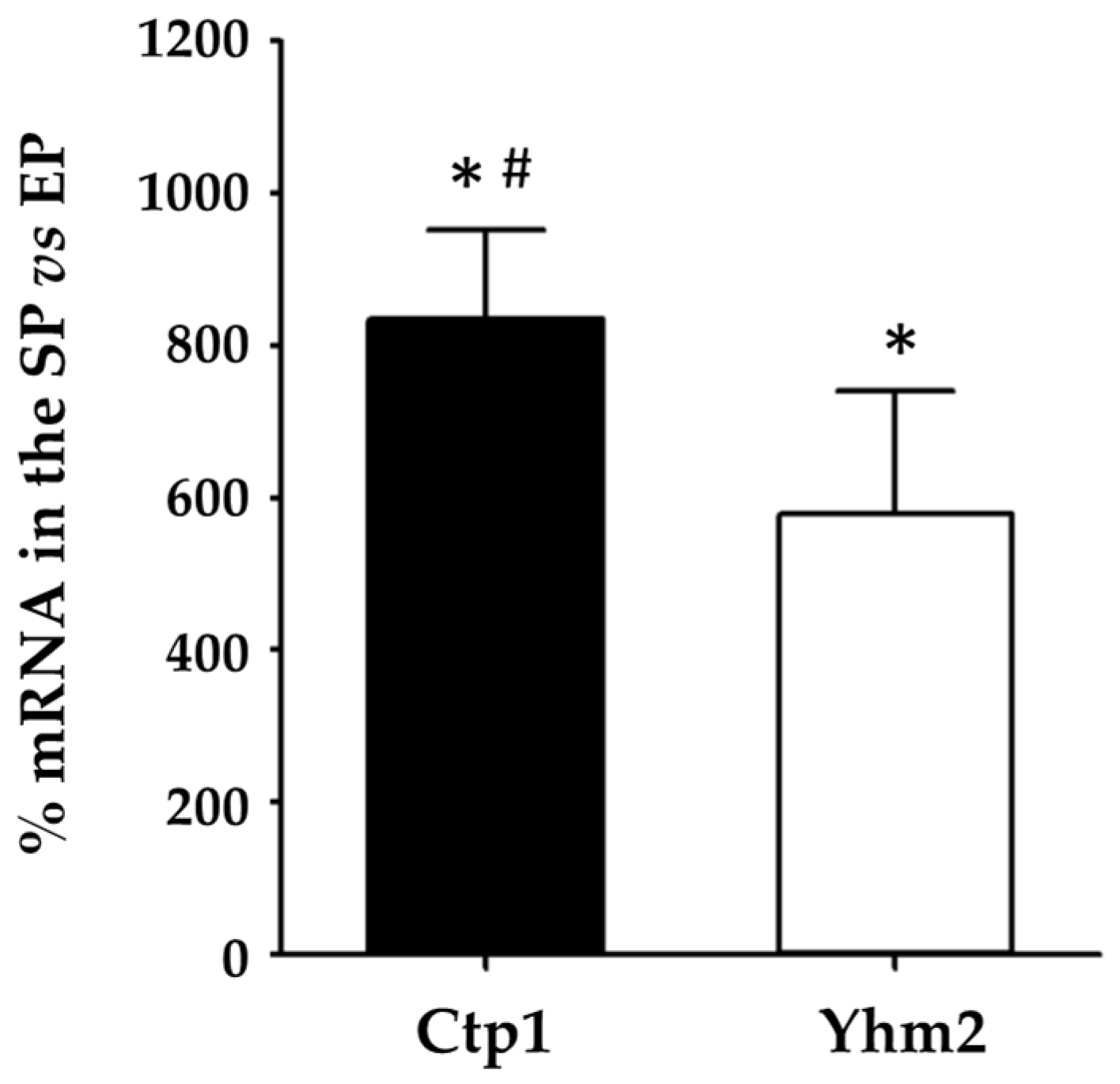
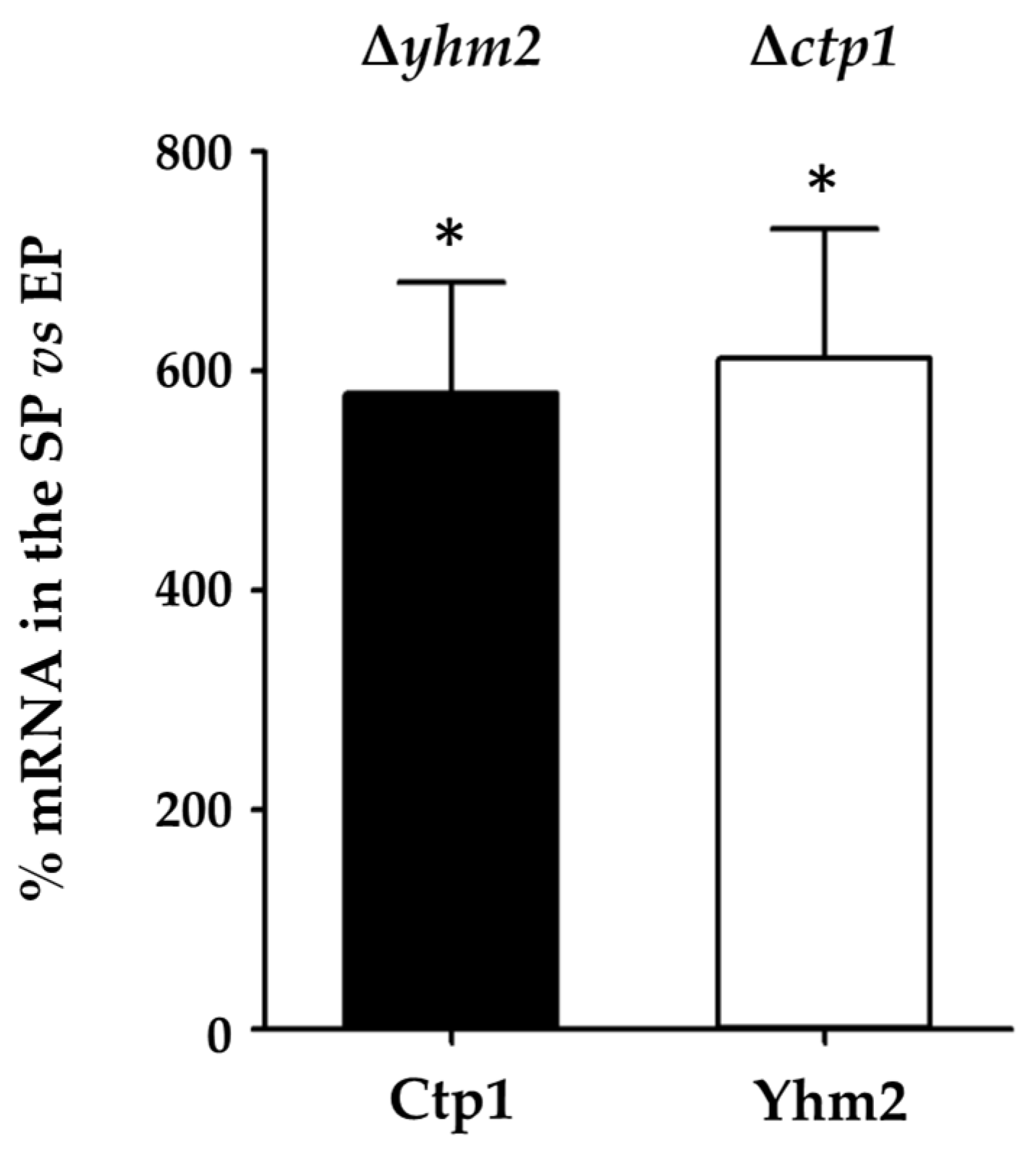
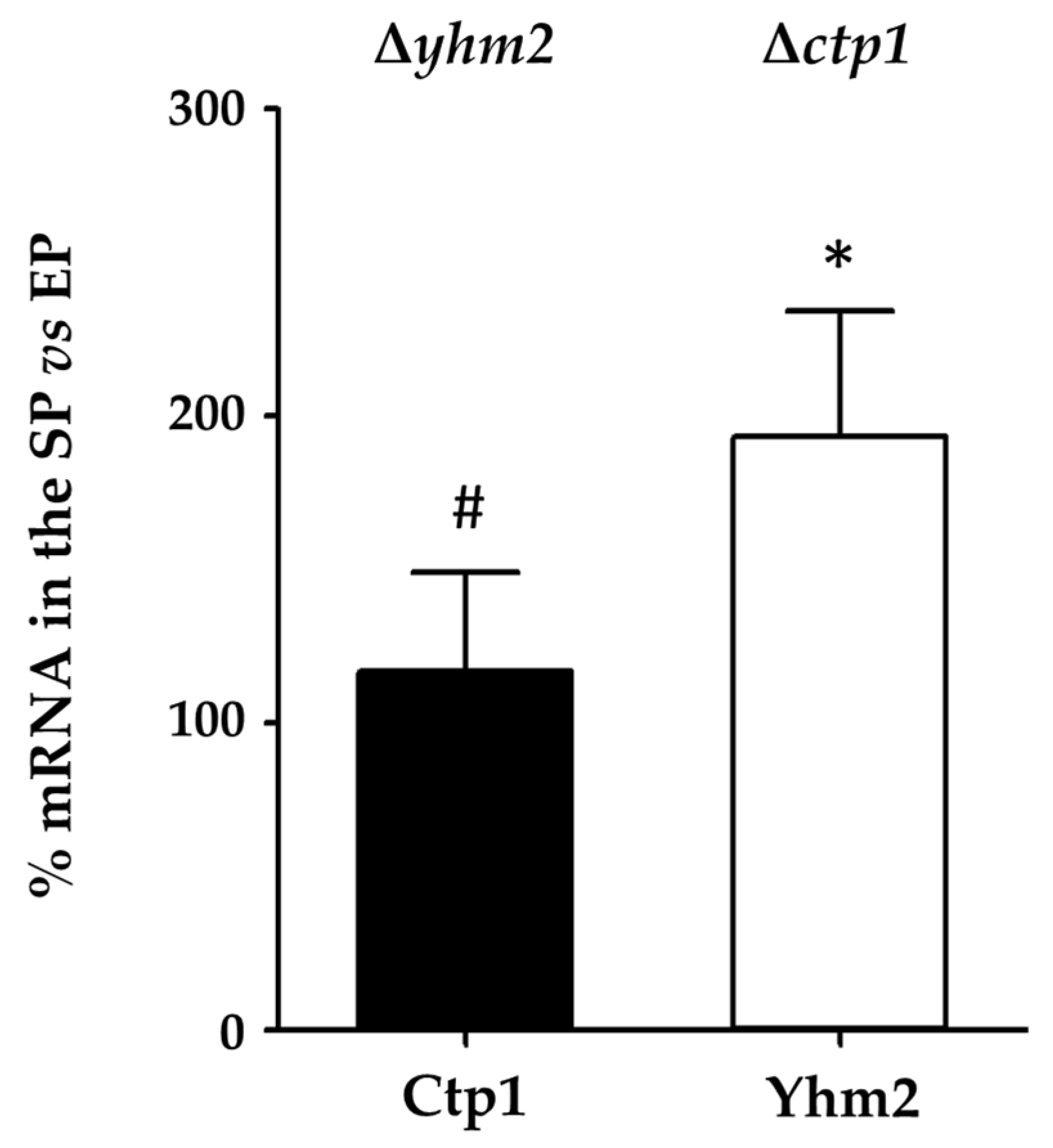
2.2. CTP1 and YHM2 Expression Levels in Wild-Type S. cerevisiae Cells Grown in Culture Media Containing Glucose or Ethanol
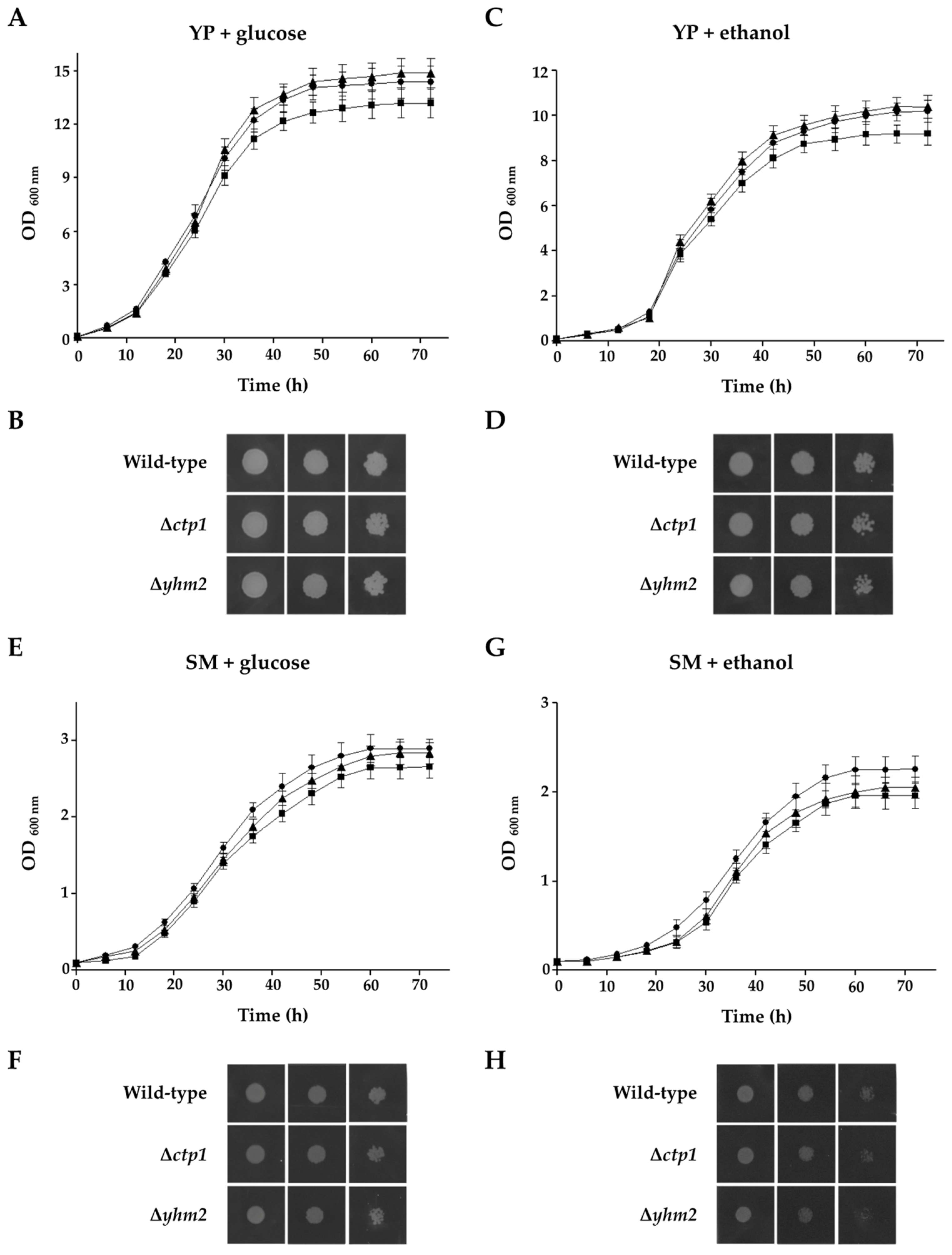
2.3. Effects of Deletion of CTP1 or YHM2 on S. cerevisiae Growth Behaviour
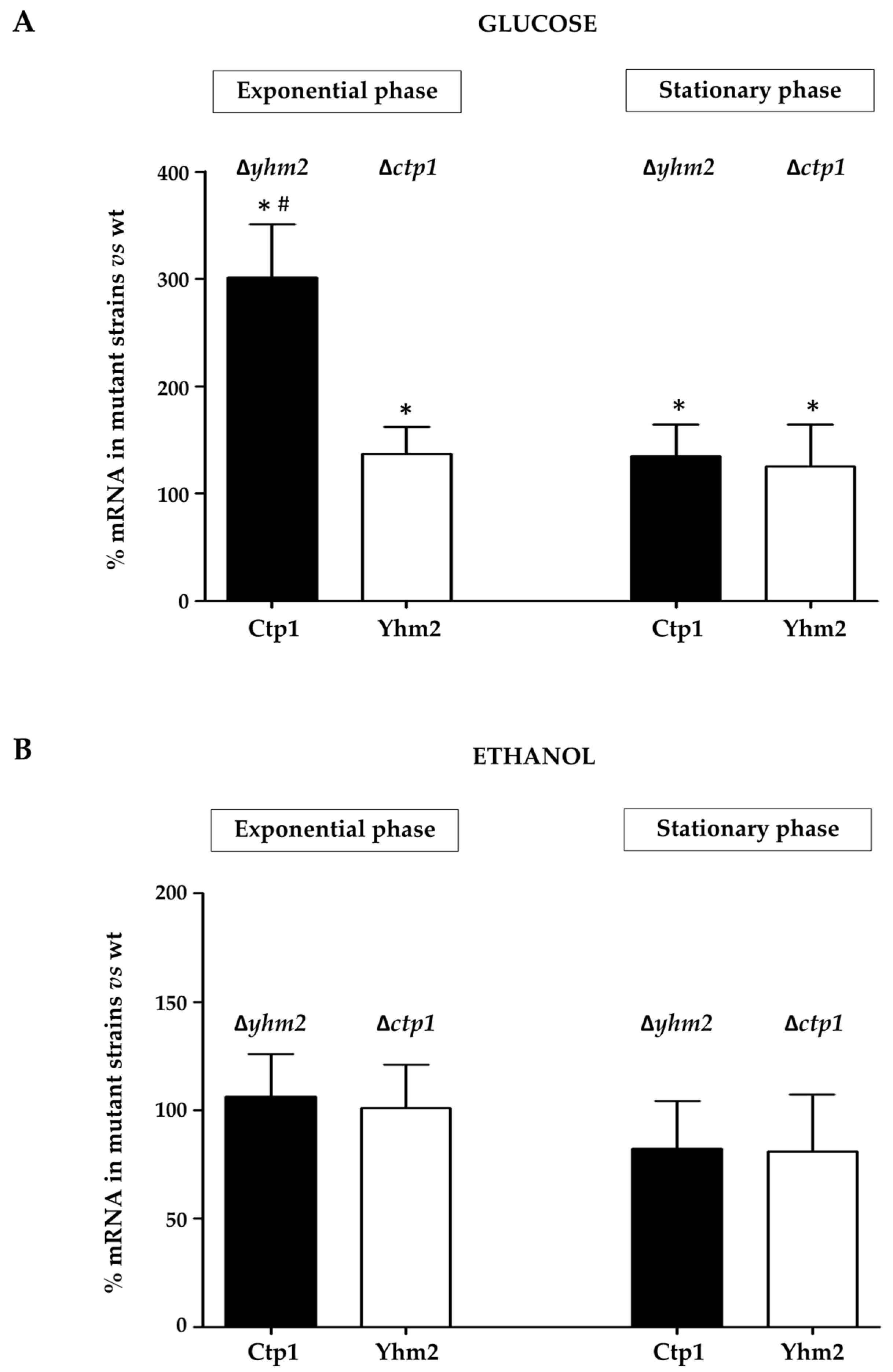
2.4. Expression Levels of Mitochondrial Citrate Transporters in Mutant Yeast Strains Grown in Culture Media Containing Glucose or Ethanol
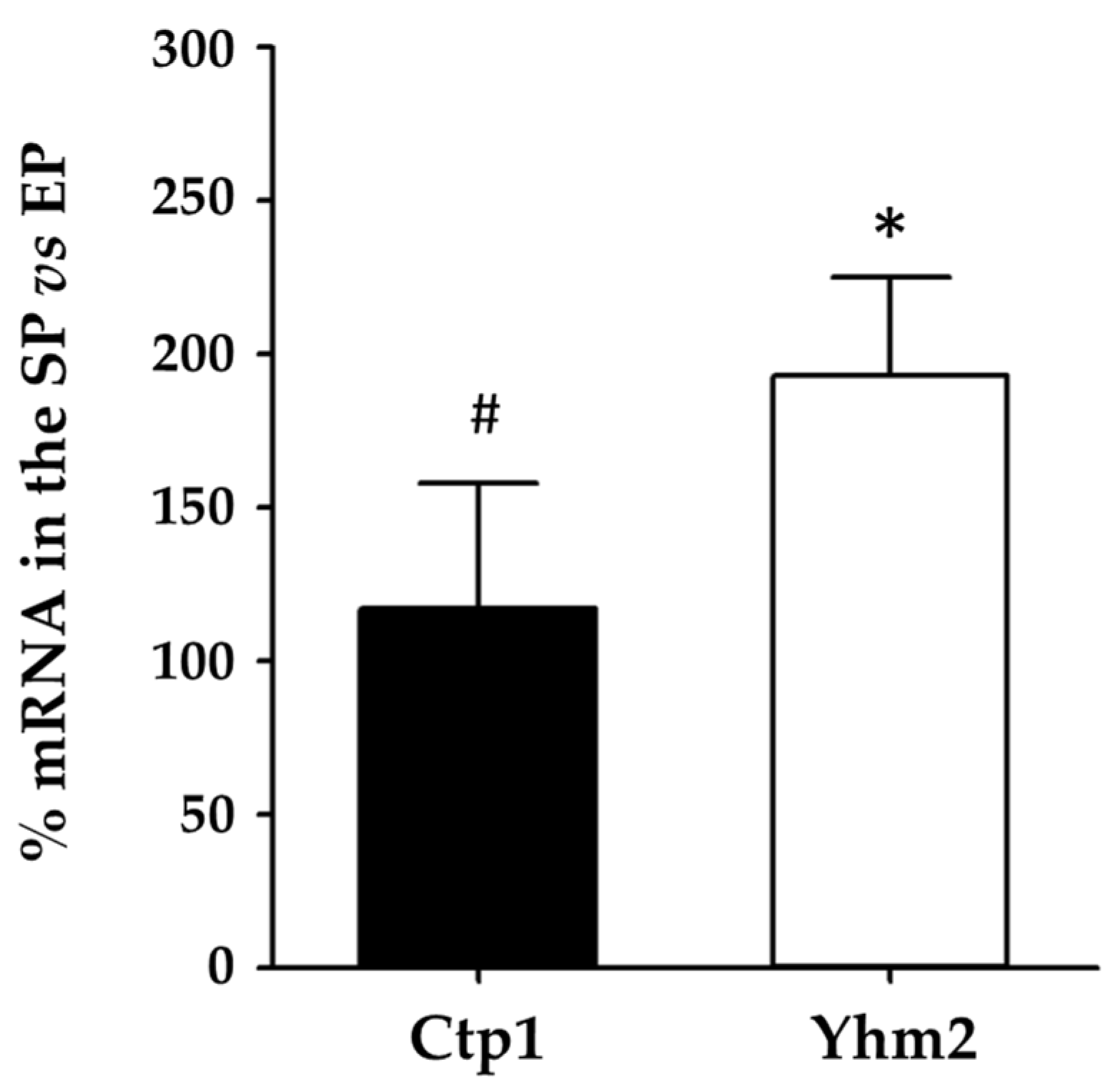
2.5. Differences in Mitochondrial Citrate Transporter Expression between Mutant and Wild-Type Yeast Strains Growing in Glucose- or Ethanol-Supplemented Culture Media
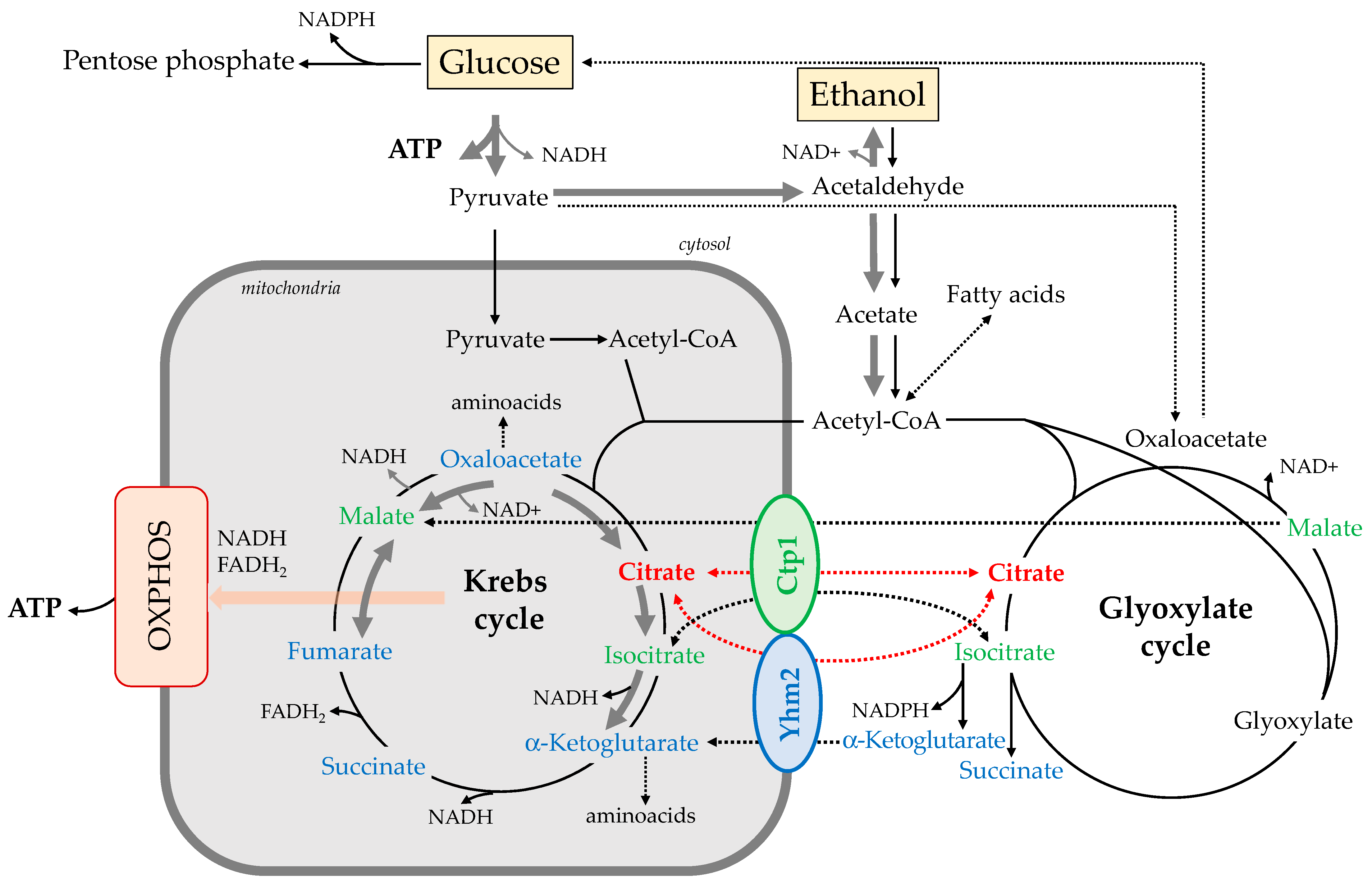
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Growth Conditions
4.2. RNA Extraction and Real-Time Quantitative PCR Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, T.; Bu, P.; Zeng, J.; Vancura, A. Increased heme synthesis in yeast induces a metabolic switch from fermentation to respiration even under conditions of glucose repression. J. Biol. Chem. 2017, 292, 16942–16954. [Google Scholar] [CrossRef]
- Yu, T.; Zhou, Y.J.; Huang, M.; Liu, Q.; Pereira, R.; David, F.; Nielsen, J. Reprogramming Yeast Metabolism from Alcoholic Fermentation to Lipogenesis. Cell 2018, 174, 1549–1558.e14. [Google Scholar] [CrossRef]
- Nielsen, J. Yeast cells handle stress by reprogramming their metabolism. Nature 2019, 572, 84–185. [Google Scholar] [CrossRef]
- Swanson, W.H.; Clifton, C.E. Growth and Assimilation in Cultures of Saccharomyces cerevisiae. J. Bacteriol. 1948, 56, 115–124. [Google Scholar] [CrossRef]
- Carlson, M. Glucose repression in yeast. Curr. Opin. Microbiol. 1999, 2, 202–207. [Google Scholar] [CrossRef]
- Johnston, M. Feasting, fasting and fermenting. Glucose sensing in yeast and other cells. Trends Genet. 1999, 15, 29–33. [Google Scholar] [CrossRef]
- Snowdon, C.; Schierholtz, R.; Poliszczuk, P.; Hughes, S.; van der Merwe, G. ETP1/YHL010c is a novel gene needed for the adaptation of Saccharomyces cerevisiae to ethanol. FEMS Yeast Res. 2009, 9, 372–380. [Google Scholar] [CrossRef]
- Turcotte, B.; Liang, X.B.; Robert, F.; Soontorngun, N. Transcriptional regulation of nonfermentable carbon utilization in budding yeast. FEMS Yeast Res. 2010, 10, 2–13. [Google Scholar] [CrossRef]
- Schüller, H.J. Transcriptional control of nonfermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 2003, 43, 139–160. [Google Scholar] [CrossRef]
- Palmieri, F.; Agrimi, G.; Blanco, E.; Castegna, A.; Di Noia, M.A.; Iacobazzi, V.; Lasorsa, F.M.; Marobbio, C.M.; Palmieri, L.; Scarcia, P.; et al. Identification of mitochondrial carriers in Saccharomyces cerevisiae by transport assay of reconstituted recombinant proteins. Biochim. Biophys. Acta 2006, 1757, 1249–1262. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Mitochondrial Carriers and Substrates Transport Network: A Lesson from Saccharomyces cerevisiae. Int. J. Mol. Sci. 2021, 22, 8496. [Google Scholar] [CrossRef]
- Mentel, M.; Chovančíková, P.; Zeman, I.; Polčic, P. Learning from Yeast about Mitochondrial Carriers. Microorganisms 2021, 9, 2044. [Google Scholar] [CrossRef]
- Kim, K.S.; Rosenkrantz, M.S.; Guarente, L. Saccharomyces cerevisiae contains two functional citrate synthase genes. Mol. Cell. Biol. 1986, 6, 1936–1942. [Google Scholar]
- Lewin, A.S.; Hines, V.; Small, G.M. Citrate synthase encoded by the CIT2 gene of Saccharomyces cerevisiae is peroxisomal. Mol. Cell. Biol. 1990, 10, 1399–1405. [Google Scholar]
- Palmieri, L.; Runswick, M.J.; Fiermonte, G.; Walker, J.E.; Palmieri, F. Yeast mitochondrial carriers: Bacterial expression, biochemical identification and metabolic significance. J. Bioenerg. Biomembr. 2000, 32, 67–77. [Google Scholar] [CrossRef]
- Kaplan, R.S.; Mayor, J.A.; Gremse, D.A.; Wood, D.O. High level expression and characterization of the mitochondrial citrate transport protein from the yeast Saccharomyces cerevisiae. J. Biol. Chem. 1995, 270, 4108–4114. [Google Scholar] [CrossRef]
- Castegna, A.; Scarcia, P.; Agrimi, G.; Palmieri, L.; Rottensteiner, H.; Spera, I.; Germinario, L.; Palmieri, F. Identification and functional characterization of a novel mitochondrial carrier for citrate and oxoglutarate in Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 17359–17370. [Google Scholar] [CrossRef]
- Di Noia, M.A.; Scarcia, P.; Agrimi, G.; Ocheja, O.B.; Wahid, E.; Pisano, I.; Paradies, E.; Palmieri, L.; Guaragnella, C.; Guaragnella, N. Inactivation of HAP4 Accelerates RTG-Dependent Osmoadaptation in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2023, 24, 5320. [Google Scholar] [CrossRef]
- Zara, V.; Assalve, G.; Ferramosca, A. Multiple roles played by the mitochondrial citrate carrier in cellular metabolism and physiology. Cell. Mol. Life Sci. 2022, 79, 428. [Google Scholar] [CrossRef]
- Evans, C.T.; Scragg, A.H.; Ratledge, C. A comparative study of citrate efflux from mitochondria of oleaginous and non-oleaginous yeasts. Eur. J. Biochem. 1983, 130, 195–204. [Google Scholar] [CrossRef]
- Cho, J.H.; Ha, S.J.; Kao, L.R.; Megraw, T.L.; Chae, C.B. A novel DNA-binding protein bound to the mitochondrial inner membrane restores the null mutation of mitochondrial histone Abf2p in Saccharomyces cerevisiae. Mol. Cell. Biol. 1998, 18, 5712–5723. [Google Scholar] [CrossRef]
- Janssens, G.E.; Meinema, A.C.; González, J.; Wolters, J.C.; Schmidt, A.; Guryev, V.; Bischoff, R.; Wit, E.C.; Veenhoff, L.M.; Heinemann, M. Protein biogenesis machinery is a driver of replicative aging in yeast. Elife 2015, 4, e08527. [Google Scholar] [CrossRef]
- Huberts, D.H.; Niebel, B.; Heinemann, M. A flux-sensing mechanism could regulate the switch between respiration and fermentation. FEMS Yeast Res. 2012, 12, 118–128. [Google Scholar] [CrossRef]
- Olivares-Marin, I.K.; González-Hernández, J.C.; Regalado-Gonzalez, C.; Madrigal-Perez, L.A. Saccharomyces cerevisiae Exponential Growth Kinetics in Batch Culture to Analyze Respiratory and Fermentative Metabolism. J. Vis. Exp. 2018, 139, 58192. [Google Scholar]
- Werner-Washburne, M.; Braun, E.; Johnston, G.C.; Singer, R.A. Stationary phase in the yeast Saccharomyces cerevisiae. Microbiol. Rev. 1993, 57, 383–401. [Google Scholar] [CrossRef]
- Werner-Washburne, M.; Braun, E.L.; Crawford, M.E.; Peck, V.M. Stationary phase in Saccharomyces cerevisiae. Mol. Microbiol. 1996, 19, 1159–1166. [Google Scholar] [CrossRef]
- Bitterman, K.J.; Medvedik, O.; Sinclair, D.A. Longevity regulation in Saccharomyces cerevisiae: Linking metabolism, genome stability, and heterochromatin. Microbiol. Mol. Biol. Rev. 2003, 67, 376–399. [Google Scholar] [CrossRef]
- Watson, J.A.; Lowenstein, J.M. Citrate and the conversion of carbohydrate into fat. Fatty acid synthesis by a combination of cytoplasm and mitochondria. J. Biol. Chem. 1970, 245, 5993–6002. [Google Scholar] [CrossRef]
- Endemann, G.; Goetz, P.G.; Edmond, J.; Brunengraber, H. Lipogenesis from ketone bodies in the isolated perfused rat liver. Evidence for the cytosolic activation of acetoacetate. J. Biol. Chem. 1982, 257, 3434–3440. [Google Scholar] [CrossRef]
- Zara, V.; Assalve, G.; Ferramosca, A. Insights into the malfunctioning of the mitochondrial citrate carrier: Implications for cell pathology. Biochim. Biophys. Acta Mol. Basis Dis. 2023, 1869, 166758. [Google Scholar] [CrossRef]
- Kadooka, C.; Izumitsu, K.; Onoue, M.; Okutsu, K.; Yoshizaki, Y.; Takamine, K.; Goto, M.; Tamaki, H.; Futagami, T. Mitochondrial Citrate Transporters CtpA and YhmA Are Required for Extracellular Citric Acid Accumulation and Contribute to Cytosolic Acetyl Coenzyme A Generation in Aspergillus luchuensis mut. kawachii. Appl. Environ. Microbiol. 2019, 85, e03136-18. [Google Scholar] [CrossRef]
- Xiberras, J.; Klein, M.; Nevoigt, E. Glycerol as a substrate for Saccharomyces cerevisiae based bioprocesses—Knowledge gaps regarding the central carbon catabolism of this ‘non-fermentable’ carbon source. Biotechnol. Adv. 2019, 37, 107378. [Google Scholar] [CrossRef]
- Boulton, C.A.; Ratledge, C. Correlation of lipid accumulation in yeasts with possession of ATP: Citrate lyase. J. Gen. Microbiol. 1981, 127, 169–176. [Google Scholar] [CrossRef]
- Leupold, S.; Hubmann, G.; Litsios, A.; Meinema, A.C.; Takhaveev, V.; Papagiannakis, A.; Niebel, B.; Janssens, G.; Siegel, D.; Heinemann, M. Saccharomyces cerevisiae goes through distinct metabolic phases during its replicative lifespan. Elife 2019, 8, e41046. [Google Scholar] [CrossRef]
- Rodrigues, F.; Ludovico, P.; Leão, C. Sugar Metabolism in Yeasts: An Overview of Aerobic and Anaerobic Glucose Catabolism. In Biodiversity and Ecophysiology of Yeasts. The Yeast Handbook; Péter, G., Rosa, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2006. [Google Scholar]
- Liao, X.S.; Small, W.C.; Srere, P.A.; Butow, R.A. Intramitochondrial functions regulate nonmitochondrial citrate synthase (CIT2) expression in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 38–46. [Google Scholar]
- Chevtzoff, C.; Yoboue, E.D.; Galinier, A.; Casteilla, L.; Daignan-Fornier, B.; Rigoulet, M.; Devin, A. Reactive oxygen species-mediated regulation of mitochondrial biogenesis in the yeast Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 1733–1742. [Google Scholar] [CrossRef]
- Kaplan, R.S.; Mayor, J.A.; Kakhniashvili, D.; Gremse, D.A.; Wood, D.O.; Nelson, D.R. Deletion of the nuclear gene encoding the mitochondrial citrate transport protein from Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 1996, 226, 657–662. [Google Scholar] [CrossRef]
- Scarcia, P.; Palmieri, L.; Agrimi, G.; Palmieri, F.; Rottensteiner, H. Three mitochondrial transporters of Saccharomyces cerevisiae are essential for ammonium fixation and lysine biosynthesis in synthetic minimal medium. Mol. Genet. Metab. 2017, 122, 54–60. [Google Scholar] [CrossRef]
- Aluvila, S.; Kotaria, R.; Sun, J.; Mayor, J.A.; Walters, D.E.; Harrison, D.H.T.; Kaplan, R.S. The yeast mitochondrial citrate transport protein: Molecular determinants of its substrate specificity. J. Biol. Chem. 2010, 285, 27314–27326. [Google Scholar] [CrossRef]
- Postma, E.D.; Couwenberg, L.G.F.; van Roosmalen, R.N.; Geelhoed, J.; de Groot, P.A.; Daran-Lapujade, P. Top-Down, Knowledge-Based Genetic Reduction of Yeast Central Carbon Metabolism. mBio 2022, 13, e0297021. [Google Scholar] [CrossRef]
- Xie, T.; Chen, M.; Nielsen, J.; Xia, J. Multi-omics analyses of the transition to the Crabtree effect in S. cerevisiae reveals a key role for the citric acid shuttle. FEMS Yeast Res. 2022, 22, foac030. [Google Scholar] [CrossRef]
- Lee, J.E.; Lee, E. The Probiotic Effects of the Saccharomyces cerevisiae 28-7 Strain Isolated from Nuruk in a DSS-Induced Colitis Mouse Model. J. Microbiol. Biotechnol. 2022, 32, 877–884. [Google Scholar] [CrossRef]
- Muratori, P.; Muratori, L.; Guidi, M.; Maccariello, S.; Pappas, G.; Ferrari, R.; Gionchetti, P.; Campieri, M.; Bianchi, F.B. Anti-Saccharomyces cerevisiae antibodies (ASCA) and autoimmune liver diseases. Clin. Exp. Immunol. 2003, 132, 473–476. [Google Scholar] [CrossRef]
- Granito, A.; Zauli, D.; Muratori, P.; Muratori, L.; Grassi, A.; Bortolotti, R.; Petrolini, N.; Veronesi, L.; Gionchetti, P.; Bianchi, F.B.; et al. Anti-Saccharomyces cerevisiae and perinuclear anti-neutrophil cytoplasmic antibodies in coeliac disease before and after gluten-free diet. Aliment. Pharmacol. Ther. 2005, 21, 881–887. [Google Scholar] [CrossRef]
- Granito, A.; Muratori, P.; Muratori, L. Editorial: Gut microbiota profile in patients with autoimmune hepatitis-a clue for adjunctive probiotic therapy? Aliment. Pharmacol. Ther. 2020, 52, 392–394. [Google Scholar] [CrossRef]
- Narendranath, N.V.; Power, R. Relationship between pH and medium dissolved solids in terms of growth and metabolism of lactobacilli and Saccharomyces cerevisiae during ethanol production. Appl. Environ. Microbiol. 2005, 71, 2239–2243. [Google Scholar] [CrossRef]
- Lunetti, P.; Damiano, F.; De Benedetto, G.; Siculella, L.; Pennetta, A.; Muto, L.; Paradies, E.; Marobbio, C.M.; Dolce, V.; Capobianco, L. Characterization of Human and Yeast Mitochondrial Glycine Carriers with Implications for Heme Biosynthesis and Anemia. J. Biol. Chem. 2016, 291, 19746–19759. [Google Scholar] [CrossRef]
- Lunetti, P.; Gorgoglione, R.; Curcio, R.; Marra, F.; Pignataro, A.; Vozza, A.; Riley, C.L.; Capobianco, L.; Palmieri, L.; Dolce, V.; et al. Drosophila melanogaster Uncoupling Protein-4A (UCP4A) Catalyzes a Unidirectional Transport of Aspartate. Int. J. Mol. Sci. 2022, 23, 1020. [Google Scholar] [CrossRef]
- Lunetti, P.; Di Giacomo, M.; Vergara, D.; De Domenico, S.; Maffia, M.; Zara, V.; Capobianco, L.; Ferramosca, A. Metabolic reprogramming in breast cancer results in distinct mitochondrial bioenergetics between luminal and basal subtypes. FEBS J. 2019, 286, 688–709. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Rate (h−1) | Doubling Time (h) | |||||||
|---|---|---|---|---|---|---|---|---|
| YP + Glucose | YP + Ethanol | SM + Glucose | SM + Ethanol | YP + Glucose | YP + Ethanol | SM + Glucose | SM + Ethanol | |
| Wild-type | 0.408 ± 0.037 | 0.300 ± 0.026 | 0.089 ± 0.006 | 0.064 ± 0.004 | 1.69 | 2.33 | 7.77 | 10.84 |
| Δctp1 | 0.421 ± 0.036 | 0.306 ± 0.025 | 0.088 ± 0.007 | 0.060 ± 0.004 | 1.64 | 2.29 | 7.85 | 11.39 |
| Δyhm2 | 0.437 ± 0.039 | 0.316 ± 0.027 | 0.087 ± 0.007 | 0.058 ± 0.004 | 1.58 | 2.22 | 7.90 | 11.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assalve, G.; Lunetti, P.; Zara, V.; Ferramosca, A. Ctp1 and Yhm2: Two Mitochondrial Citrate Transporters to Support Metabolic Flexibility of Saccharomyces cerevisiae. Int. J. Mol. Sci. 2024, 25, 1870. https://doi.org/10.3390/ijms25031870
Assalve G, Lunetti P, Zara V, Ferramosca A. Ctp1 and Yhm2: Two Mitochondrial Citrate Transporters to Support Metabolic Flexibility of Saccharomyces cerevisiae. International Journal of Molecular Sciences. 2024; 25(3):1870. https://doi.org/10.3390/ijms25031870
Chicago/Turabian StyleAssalve, Graziana, Paola Lunetti, Vincenzo Zara, and Alessandra Ferramosca. 2024. "Ctp1 and Yhm2: Two Mitochondrial Citrate Transporters to Support Metabolic Flexibility of Saccharomyces cerevisiae" International Journal of Molecular Sciences 25, no. 3: 1870. https://doi.org/10.3390/ijms25031870