
miRNAs Related to Immune Checkpoint Inhibitor Response: A Systematic Review
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
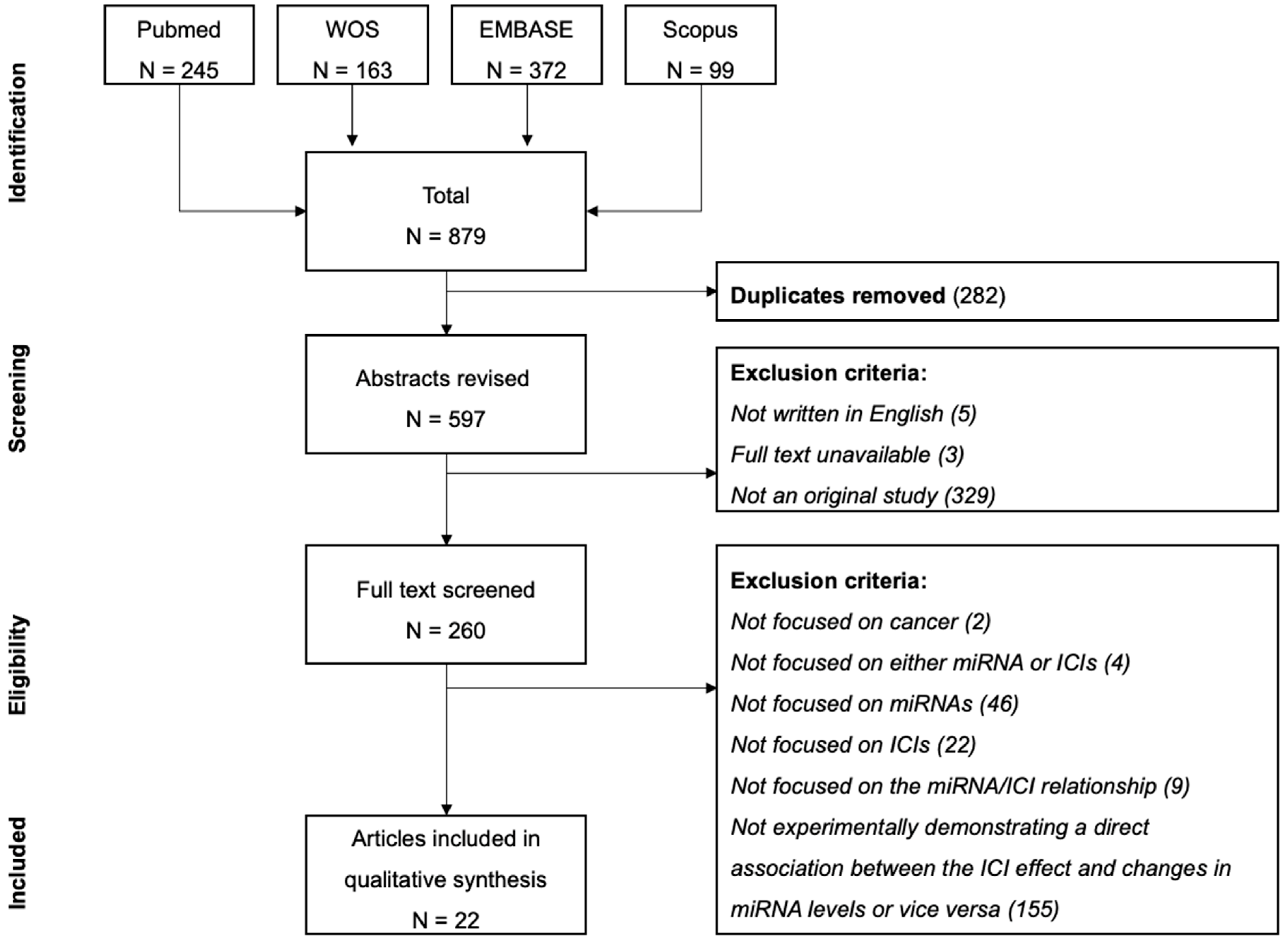
2. Materials and Methods
2.1. Search Strategy
2.2. Screening and Eligibility Criteria
2.3. Data Extraction
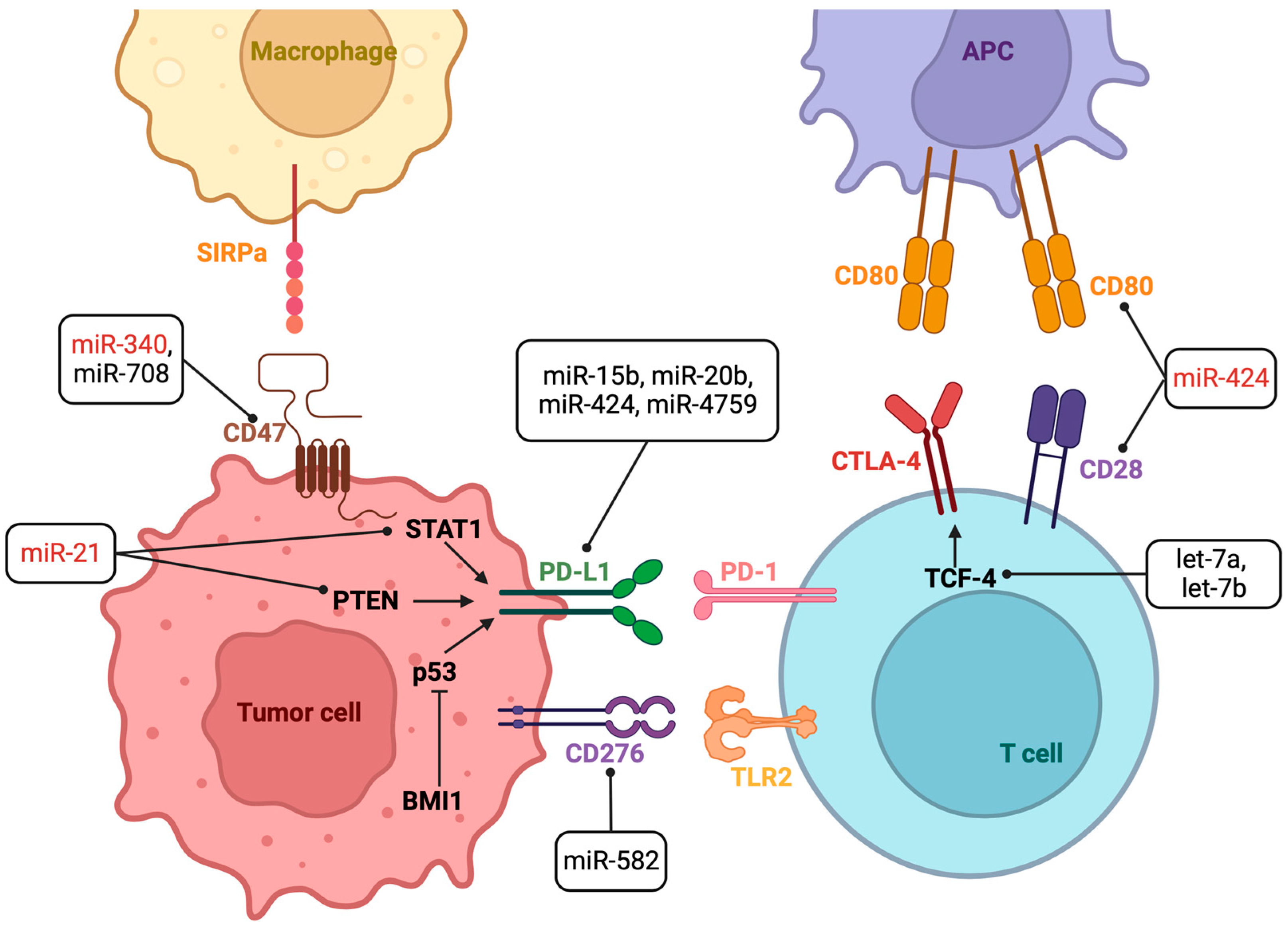
3. Results

{kind=link}
{kind=link}
{kind=link}
| miRNA | Cancer | miRNA Target Gene | ICI | Experimental Model | Effect on ICI Response | Refs. |
|---|---|---|---|---|---|---|
| let-7a and let-7b | Head and neck squamous cell carcinoma | TCF-4 * | anti CTLA-4 | Overexpressing let-7a/b tumor cells inoculated into mice + anti CTLA-4 | H | [25] |
| miR-15b-5p | Colorectal cancer | PD-L1 * | anti PD-1 | Overexpressing miR-15b-5p tumor cells inoculated into mice + anti PD-1. | H | [26] |
| miR-16-5p | Lung cancer | anti PD-L1 | Tumor cell + overexpressing miR-16-5p exosomes + anti PD-L1 | H | [27] | |
| miR-20b-5p | Lung cancer | PD-L1 * | anti PD-1 | Tumor cells transfected with miRNA mimic + Pembrolizumab | H | [28] |
| Breast cancer | Tumor cells transfected with miRNA mimic + Pembrolizumab | H | ||||
| miR-21 | Oral squamous cell carcinoma | PTEN | anti PD-L1 | Tumor cells inoculated into mice + miR-21 knockdown tumor-derived exosomes + anti-PD-L1 | L | [29] |
| Melanoma | STAT1 * | anti PD-1 | Tumor cells and knocked down miR-21 tumor-associated macrophages (TAM) subcutaneously injected in mice + anti PD-1 | L | [30] | |
| miR-128a | Laryngeal squamous cell carcinoma | BMI1 * | anti PD-1 | Overexpressing miR-128a tumor cells + Pembrolizumab | H | [31] |
| miR-155 | Metastatic melanoma | anti PD-1 + anti PD-L1 + anti CTLA-4 | Tumor cells inoculated into modified mice for knockout of miR-155 in CD4/8 T cells + anti PD-1, anti PD-L1 and anti CTLA-4 | L | [32] | |
| Diffuse large B-cell lymphoma | PD-L1 * | anti PD-L1 | Overexpressing miR-155 tumor cells inoculated into mice + anti PD-L1 | L | [33] | |
| Breast cancer | SOCS1 | anti PD-L1 | Overexpressing miR-155 tumor cells inoculated into mice + anti PD-L1 | H | [34] | |
| Melanoma | PD-L1 * | anti PD-L1 | Overexpressing miR-155 tumor cells co-cultured with peripheral blood mononuclear cells + anti PD-L1 | H | [22] | |
| miR-340 | Pancreatic carcinoma | CD47 * | anti CD47 | Overexpressing miR-340 tumor cells inoculated into mice + anti CD47 | L | [35] |
| miR-424 | Colorectal cancer | CD28 and CD80 * | anti PD-1 + anti CTLA-4 | Tumor cells inoculated into miR-424 knocked mice + anti PD-1 and anti CTLA-4 | L | [36] |
| Mouse cecum orthotopic colorectal cancer + miR-424 knocked tumor cell-derived extracellular vesicles + anti PD-1 and anti CTLA-4 | L | |||||
| Hepatocellular carcinoma | PD-L1 | anti PD-L1 | Tumor cells inoculated into mice + nanobubbles carrying miR-424 mimic and anti PD-L1 | H | [37] | |
| miR-582 | B-cell precursor acute lymphoblastic leukemia | CD276 * | anti CD276 | Overexpressing miR-582 tumor cells co-cultured with NK cells + anti CD276 | H | [38] |
| miR-708 | T-acute lymphoblastic leukemia | CD47 * | anti CD47 | Overexpressing miR-708 tumor cells + anti CD47. | H | [39] |
| miR-4759 | Breast cancer | PD-L1 * | anti PD-L1 | Overexpressing miR-4759 tumor cells co-cultured with peripheral blood mononuclear cells + anti PD-L1 | H | [40] |
| ICI | Experimental Model | miRNA | Experimental Effect on miRNA | Refs. |
|---|---|---|---|---|
| anti PD-1 | miRNA analysis in peripheral lymphocytes from 21 good responders (complete response, partial response, or stable disease) with metastatic renal cell carcinoma before and after a 4-weeks period (2 cycles) of nivolumab administration | miR-99a, miR-708, miR-655, miR-582-3p, miR-492, miR-487a, miR-485-3p, miR-449a, miR-433, miR-431, miR-429, miR-376c, miR-342-5p, miR-340, miR-339-5p, miR-335, miR-324-5p, miR-25, miR-24, miR-22, miR-221, miR-214, miR-194, miR-18b, miR-152, miR-143, miR-100, miR-let-7ev | High levels of expression in peripheral lymphocytes after treatment compared to before treatment in good responders. | [41] |
| miRNA analysis in peripheral lymphocytes from 17 good long-responders (complete response, partial response or stable disease and progression-free survival (PFS) > 18 months) with metastatic renal cell carcinoma before and after a 4-weeks period (2 cycles) of nivolumab administration | miR-22, miR-24, miR-99a, miR-194, miR-214, miR-335, miR-339, miR-708 | High expression levels in peripheral lymphocytes after treatment compared to before treatment in good responders. | ||
| anti CTLA-4 + anti PD-1 | Plasma from stage IV melanoma non-responders (13 patients), partial response (4 patients) and complete response (5 patients) before and after Ipilimumab and Nivolumab/Pembrolizumab treatment | miR-4649-3p and miR-615-3p | Increased levels in post- vs. pre-treatment in non-responders. No changes post- vs. pre-treatment in patients with partial response. Decreased levels post- vs. pre-treatment in patients with complete response. | [42] |
| ICI | miRNA | Immune-Related Adverse Events (irAEs) | Refs. |
|---|---|---|---|
| anti PD-1 | miR-34a | Anti PD-1 induced miR-34a-5p upregulation, modulating the miR-34a/Krüppel-like factor 4 (KLF4)-and miR-34a-5p/serine/threonine-protein phosphatase 1 regulatory subunit 10 (PNUTS) signaling pathway and inducing cardiac injury | [43,44] |
| anti PD-1 + anti CTLA-4 | miR-146a | Downregulation of miR-146 increased the risk of developing irAE after ICI treatment. Toxicity was accompanied by increased infiltration of neutrophil and lymphocyte into the damaged organs. | [45] |
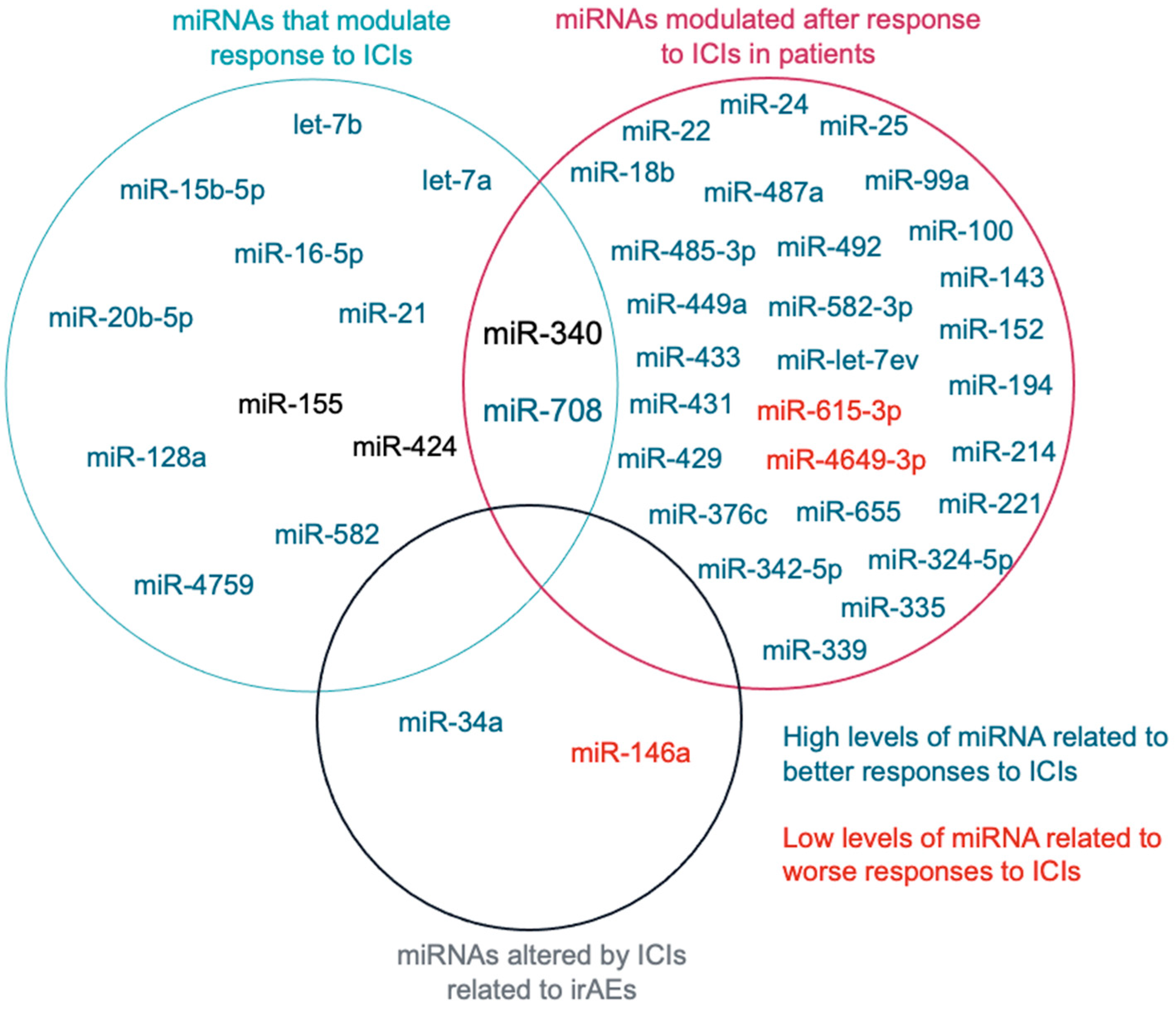
3.1. miRNAs That Modulate Response to ICIs
3.2. miRNAs Modulated after Response to ICIs
3.3. miRNAs That Regulate Immune-Related Adverse Events (irAEs)
3.4. miRNAs Related to ICIs Response
4. Discussion
4.1. miRNAs That Modulate Response to ICIs
4.2. miRNAs Modulated after Response to ICIs
4.3. miRNAs That Regulate Immune-Related Adverse Events (irAEs)
4.4. miRNAs Related to ICIs Response
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pandya, P.H.; Murray, M.E.; Pollok, K.E.; Renbarger, J.L. The Immune System in Cancer Pathogenesis: Potential Therapeutic Approaches. J. Immunol. Res. 2016, 2016, 4273943. [Google Scholar] [CrossRef]
- Kanchan, R.K.; Doss, D.; Khan, P.; Nasser, M.W.; Mahapatra, S. To Kill a Cancer: Targeting the Immune Inhibitory Checkpoint Molecule, B7-H3. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2022, 1877, 188783. [Google Scholar] [CrossRef]
- Khan, M.; Arooj, S.; Wang, H. Soluble B7-CD28 Family Inhibitory Immune Checkpoint Proteins and Anti-Cancer Immunotherapy. Front. Immunol. 2021, 12, 651634. [Google Scholar] [CrossRef] [PubMed]
- Esensten, J.H.; Helou, Y.A.; Chopra, G.; Weiss, A.; Bluestone, J.A. CD28 Costimulation: From Mechanism to Therapy. Immunity 2016, 44, 973–988. [Google Scholar] [CrossRef]
- Saadi, W.; Fatmi, A.; Pallardó, F.V.; García-Giménez, J.L.; Mena-Molla, S. Long Non-Coding RNAs as Epigenetic Regulators of Immune Checkpoints in Cancer Immunity. Cancers 2022, 15, 184. [Google Scholar] [CrossRef]
- Lee, Y.; Martin-Orozco, N.; Zheng, P.; Li, J.; Zhang, P.; Tan, H.; Park, H.J.; Jeong, M.; Chang, S.H.; Kim, B.-S.; et al. Inhibition of the B7-H3 Immune Checkpoint Limits Tumor Growth by Enhancing Cytotoxic Lymphocyte Function. Cell Res. 2017, 27, 1034–1045. [Google Scholar] [CrossRef]
- Yang, H.; Xun, Y.; You, H. The Landscape Overview of CD47-Based Immunotherapy for Hematological Malignancies. Biomark. Res. 2023, 11, 15. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Shay, C.; Abousaud, M.; Tang, C.; Li, Y.; Qin, Z.; Saba, N.F.; Teng, Y. Patterns of Toxicity Burden for FDA-Approved Immune Checkpoint Inhibitors in the United States. J. Exp. Clin. Cancer Res. 2023, 42, 4. [Google Scholar] [CrossRef]
- De Risi, I.; Sciacovelli, A.M.; Guida, M. Checkpoint Inhibitors Immunotherapy in Metastatic Melanoma: When to Stop Treatment? Biomedicines 2022, 10, 2424. [Google Scholar] [CrossRef] [PubMed]
- Ratner, L.; Waldmann, T.A.; Janakiram, M.; Brammer, J.E. Rapid Progression of Adult T-Cell Leukemia–Lymphoma after PD-1 Inhibitor Therapy. N. Engl. J. Med. 2018, 378, 1947–1948. [Google Scholar] [CrossRef]
- Nelson, B.E.; Ejezie, C.L.; Stephen, B.A.; Nardo, M.; Campbell, E.; Gong, J.; Hong, D.S.; Fu, S.; Yap, T.A.; Murphy, M.B.; et al. Spectrum of Immune Checkpoint Inhibitor Anemias: Results from a Single Center, Early-Phase Clinical Trials Case Series Experience. J. Hematol. 2022, 11, 113–120. [Google Scholar] [CrossRef]
- Ulas, E.B.; Hashemi, S.M.S.; Houda, I.; Kaynak, A.; Veltman, J.D.; Fransen, M.F.; Radonic, T.; Bahce, I. Predictive Value of Combined Positive Score and Tumor Proportion Score for Immunotherapy Response in Advanced NSCLC. JTO Clin. Res. Rep. 2023, 4, 100532. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Gu, S.; Pan, D.; Fu, J.; Sahu, A.; Hu, X.; Li, Z.; Traugh, N.; Bu, X.; Li, B.; et al. Signatures of T Cell Dysfunction and Exclusion Predict Cancer Immunotherapy Response. Nat. Med. 2018, 24, 1550–1558. [Google Scholar] [CrossRef]
- Grenda, A.; Nicoś, M.; Szczyrek, M.; Krawczyk, P.; Kucharczyk, T.; Jarosz, B.; Pankowski, J.; Sawicki, M.; Szumiło, J.; Bukała, P.; et al. MicroRNAs Aid the Assessment of Programmed Death Ligand 1 Expression in Patients with Non-Small Cell Lung Cancer. Oncol. Lett. 2019, 17, 5193–5200. [Google Scholar] [CrossRef]
- Lawlor, R.T.; Mattiolo, P.; Mafficini, A.; Hong, S.-M.; Piredda, M.L.; Taormina, S.V.; Malleo, G.; Marchegiani, G.; Pea, A.; Salvia, R.; et al. Tumor Mutational Burden as a Potential Biomarker for Immunotherapy in Pancreatic Cancer: Systematic Review and Still-Open Questions. Cancers 2021, 13, 3119. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Li, X.; Huang, Q.; Zheng, X.; Liu, M. Progress and Challenges of Predictive Biomarkers for Immune Checkpoint Blockade. Front. Oncol. 2021, 11, 617335. [Google Scholar] [CrossRef] [PubMed]
- McKean, W.B.; Moser, J.C.; Rimm, D.; Hu-Lieskovan, S. Biomarkers in Precision Cancer Immunotherapy: Promise and Challenges. Am. Soc. Clin. Oncol. Educ. Book 2020, 40, e275–e291. [Google Scholar] [CrossRef]
- Qin, S.; Jiang, J.; Lu, Y.; Nice, E.C.; Huang, C.; Zhang, J.; He, W. Emerging Role of Tumor Cell Plasticity in Modifying Therapeutic Response. Signal Transduct. Target. Ther. 2020, 5, 228. [Google Scholar] [CrossRef]
- Gong, W.-J.; Zhou, T.; Wu, S.-L.; Huang, Y.-F.; Xiang, L.-P.; Xu, J.-Q.; Han, Y.; Lv, Y.-N.; Zeng, F.; Zhang, Y. A Novel Immune-Related ceRNA Network That Predicts Prognosis and Immunotherapy Response in Lung Adenocarcinoma. Ann. Transl. Med. 2021, 9, 1484. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Q.; Qiu, Z.; Kang, Y.; Liu, J.; Ning, S.; Yin, Y.; Pang, D.; Xu, S. Noncoding RNAs: The Shot Callers in Tumor Immune Escape. Signal Transduct. Target. Ther. 2020, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dai, J.; Huang, W.; Chen, Q.; Chen, W.; He, Q.; Chen, F.; Zhang, P. Identification of a Competing Endogenous RNA Network Related to Immune Signature in Clear Cell Renal Cell Carcinoma. Aging 2021, 13, 25980–26002. [Google Scholar] [CrossRef]
- Vaxevanis, C.K.; Friedrich, M.; Tretbar, S.U.; Handke, D.; Wang, Y.; Blümke, J.; Dummer, R.; Massa, C.; Seliger, B. Identification and Characterization of Novel CD274 (PD-L1) Regulating microRNAs and Their Functional Relevance in Melanoma. Clin. Transl. Med. 2022, 12, e934. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, M.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust Enumeration of Cell Subsets from Tissue Expression Profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Liu, X.; Han, G.; Liu, Y.; Zhao, X.; Wang, D.; Bian, X.; Gu, T.; Wen, L. The Let-7 Family of microRNAs Suppresses Immune Evasion in Head and Neck Squamous Cell Carcinoma by Promoting PD-L1 Degradation. Cell Commun. Signal. 2019, 17, 173. [Google Scholar] [CrossRef]
- Liu, C.; Liu, R.; Wang, B.; Lian, J.; Yao, Y.; Sun, H.; Zhang, C.; Fang, L.; Guan, X.; Shi, J.; et al. Blocking IL-17A Enhances Tumor Response to Anti-PD-1 Immunotherapy in Microsatellite Stable Colorectal Cancer. J. Immunother. Cancer 2021, 9, e001895. [Google Scholar] [CrossRef]
- Chen, H.-L.; Luo, Y.-P.; Lin, M.-W.; Peng, X.-X.; Liu, M.-L.; Wang, Y.-C.; Li, S.-J.; Yang, D.-H.; Yang, Z.-X. Serum Exosomal miR-16-5p Functions as a Tumor Inhibitor and a New Biomarker for PD-L1 Inhibitor-Dependent Immunotherapy in Lung Adenocarcinoma by Regulating PD-L1 Expression. Cancer Med. 2022, 11, 2627–2643. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zou, H. microRNA-20b-5p Overexpression Combing Pembrolizumab Potentiates Cancer Cells to Radiation Therapy via Repressing Programmed Death-Ligand 1. Bioengineered 2022, 13, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cao, B.; Liang, X.; Lu, S.; Luo, H.; Wang, Z.; Wang, S.; Jiang, J.; Lang, J.; Zhu, G. Microenvironmental Oxygen Pressure Orchestrates an Anti- and pro-Tumoral Γδ T Cell Equilibrium via Tumor-Derived Exosomes. Oncogene 2019, 38, 2830–2843. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; Huang, Q.; Wang, L.; Ma, X.; Deng, Q.; Kumar, M.; Zhou, Z.; Li, L.; Zeng, C.; Young, K.H.; et al. miR-21 Depletion in Macrophages Promotes Tumoricidal Polarization and Enhances PD-1 Immunotherapy. Oncogene 2018, 37, 3151–3165. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Guo, Y.; Huang, J.; Zhou, L. Upregulating Hsa-miR-128a Increased the Effects of Pembrolizumab on Laryngeal Cancer Cells via the P53 Pathway. Biomed. Res. Int. 2021, 2021, 2342784. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, T.B.; Lee, S.-H.; Tang, W.W.; Wallace, J.A.; Alexander, M.; Runtsch, M.C.; Larsen, D.K.; Thompson, J.; Ramstead, A.G.; Voth, W.P.; et al. Antitumor Immunity Is Defective in T Cell-Specific microRNA-155-Deficient Mice and Is Rescued by Immune Checkpoint Blockade. J. Biol. Chem. 2017, 292, 18530–18541. [Google Scholar] [CrossRef]
- Zheng, Z.; Sun, R.; Zhao, H.-J.; Fu, D.; Zhong, H.-J.; Weng, X.-Q.; Qu, B.; Zhao, Y.; Wang, L.; Zhao, W.-L. MiR155 Sensitized B-Lymphoma Cells to Anti-PD-L1 Antibody via PD-1/PD-L1-Mediated Lymphoma Cell Interaction with CD8+T Cells. Mol. Cancer 2019, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Q.; Guan, Y.; Sun, Y.; Wang, X.; Lively, K.; Wang, Y.; Luo, M.; Kim, J.A.; Murphy, E.A.; et al. Breast Cancer Cell-Derived microRNA-155 Suppresses Tumor Progression via Enhancing Immune Cell Recruitment and Antitumor Function. J. Clin. Investig. 2022, 132, e157248. [Google Scholar] [CrossRef] [PubMed]
- Xi, Q.; Zhang, J.; Yang, G.; Zhang, L.; Chen, Y.; Wang, C.; Zhang, Z.; Guo, X.; Zhao, J.; Xue, Z.; et al. Restoration of miR-340 Controls Pancreatic Cancer Cell CD47 Expression to Promote Macrophage Phagocytosis and Enhance Antitumor Immunity. J. Immunother. Cancer 2020, 8, e000253. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yuan, C.; Wangmo, D.; Subramanian, S. Tumor-Secreted Extracellular Vesicles Regulate T-Cell Costimulation and Can Be Manipulated to Induce Tumor-Specific T-Cell Responses. Gastroenterology 2021, 161, 560–574.e11. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xie, Q.; Ma, Y.; Lin, C.; Li, J.; Hu, B.; Liu, C.; Zhao, Y. Nanobubbles Containing PD-L1 Ab and miR-424 Mediated PD-L1 Blockade, and Its Expression Inhibition to Enable and Potentiate Hepatocellular Carcinoma Immunotherapy in Mice. Int. J. Pharm. 2022, 629, 122352. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; He, F.; Gao, D.; Che, B.; Cao, X.; Huang, S.; Zheng, M.; Han, H. miR-582 Suppresses the Proliferation of B-Cell Precursor Acute Lymphoblastic Leukemia (BCP-ALL) Cells and Protects Them from Natural Killer Cell-Mediated Cytotoxicity. Front. Immunol. 2022, 13, 853094. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, W.-T.; Fang, K.; Chen, Z.-H.; Sun, Y.-M.; Han, C.; Sun, L.-Y.; Luo, X.-Q.; Chen, Y.-Q. MIR-708 Promotes Phagocytosis to Eradicate T-ALL Cells by Targeting CD47. Mol. Cancer 2018, 17, 12. [Google Scholar] [CrossRef]
- Lin, Y.-Z.; Liu, S.-H.; Wu, W.-R.; Shen, Y.-C.; Wang, Y.-L.; Liao, C.-C.; Lin, P.-L.; Chang, H.; Liu, L.-C.; Cheng, W.-C.; et al. miR-4759 Suppresses Breast Cancer through Immune Checkpoint Blockade. Comput. Struct. Biotechnol. J. 2021, 20, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Incorvaia, L.; Fanale, D.; Badalamenti, G.; Brando, C.; Bono, M.; De Luca, I.; Algeri, L.; Bonasera, A.; Corsini, L.R.; Scurria, S.; et al. A “Lymphocyte MicroRNA Signature” as Predictive Biomarker of Immunotherapy Response and Plasma PD-1/PD-L1 Expression Levels in Patients with Metastatic Renal Cell Carcinoma: Pointing towards Epigenetic Reprogramming. Cancers 2020, 12, 3396. [Google Scholar] [CrossRef] [PubMed]
- Bustos, M.A.; Gross, R.; Rahimzadeh, N.; Cole, H.; Tran, L.T.; Tran, K.D.; Takeshima, L.; Stern, S.L.; O’Day, S.; Hoon, D.S.B. A Pilot Study Comparing the Efficacy of Lactate Dehydrogenase Levels Versus Circulating Cell-Free microRNAs in Monitoring Responses to Checkpoint Inhibitor Immunotherapy in Metastatic Melanoma Patients. Cancers 2020, 12, 3361. [Google Scholar] [CrossRef]
- Xia, W.; Chen, H.; Chen, D.; Ye, Y.; Xie, C.; Hou, M. PD-1 Inhibitor Inducing Exosomal miR-34a-5p Expression Mediates the Cross Talk between Cardiomyocyte and Macrophage in Immune Checkpoint Inhibitor-Related Cardiac Dysfunction. J. Immunother. Cancer 2020, 8, e001293. [Google Scholar] [CrossRef]
- Xia, W.; Zou, C.; Chen, H.; Xie, C.; Hou, M. Immune Checkpoint Inhibitor Induces Cardiac Injury through Polarizing Macrophages via Modulating microRNA-34a/Kruppel-like Factor 4 Signaling. Cell Death Dis. 2020, 11, 575. [Google Scholar] [CrossRef]
- Marschner, D.; Falk, M.; Javorniczky, N.R.; Hanke-Müller, K.; Rawluk, J.; Schmitt-Graeff, A.; Simonetta, F.; Haring, E.; Dicks, S.; Ku, M.; et al. MicroRNA-146a Regulates Immune-Related Adverse Events Caused by Immune Checkpoint Inhibitors. JCI Insight 2020, 5, 132334. [Google Scholar] [CrossRef]
- Zhou, J.; Xiang, H.; Cao, Z. Dual Mechanism of Let-7i in Tumor Progression. Front. Oncol. 2023, 13, 1253191. [Google Scholar] [CrossRef]
- Zhang, L.; Huang, J.; Yang, N.; Greshock, J.; Megraw, M.S.; Giannakakis, A.; Liang, S.; Naylor, T.L.; Barchetti, A.; Ward, M.R.; et al. microRNAs Exhibit High Frequency Genomic Alterations in Human Cancer. Proc. Natl. Acad. Sci. USA 2006, 103, 9136–9141. [Google Scholar] [CrossRef]
- Piskounova, E.; Polytarchou, C.; Thornton, J.E.; Hagan, J.P.; LaPierre, R.J.; Pothoulakis, C.; Iliopoulos, D.; Gregory, R.I. Oncogenic Lin28A and Lin28B Inhibit Let-7 microRNA Biogenesis by Distinct Mechanisms. Cell 2011, 147, 1066–1079. [Google Scholar] [CrossRef]
- Lu, L.; Katsaros, D.; de la Longrais, I.A.R.; Sochirca, O.; Yu, H. Hypermethylation of Let-7a-3 in Epithelial Ovarian Cancer Is Associated with Low Insulin-like Growth Factor-II Expression and Favorable Prognosis. Cancer Res. 2007, 67, 10117–10122. [Google Scholar] [CrossRef] [PubMed]
- Baer, C.; Squadrito, M.L.; Laoui, D.; Thompson, D.; Hansen, S.K.; Kiialainen, A.; Hoves, S.; Ries, C.H.; Ooi, C.-H.; De Palma, M. Suppression of microRNA Activity Amplifies IFN-γ-Induced Macrophage Activation and Promotes Anti-Tumour Immunity. Nat. Cell Biol. 2016, 18, 790–802. [Google Scholar] [CrossRef]
- Hsu, J.-M.; Xia, W.; Hsu, Y.-H.; Chan, L.-C.; Yu, W.-H.; Cha, J.-H.; Chen, C.-T.; Liao, H.-W.; Kuo, C.-W.; Khoo, K.-H.; et al. STT3-Dependent PD-L1 Accumulation on Cancer Stem Cells Promotes Immune Evasion. Nat. Commun. 2018, 9, 1908. [Google Scholar] [CrossRef]
- Chen, Y.; Xie, C.; Zheng, X.; Nie, X.; Wang, Z.; Liu, H.; Zhao, Y. LIN28/Let-7/PD-L1 Pathway as a Target for Cancer Immunotherapy. Cancer Immunol. Res. 2019, 7, 487–497. [Google Scholar] [CrossRef]
- Palamarchuk, A.; Tsyba, L.; Tomasello, L.; Pekarsky, Y.; Croce, C.M. PDCD1 (PD-1) Is a Direct Target of miR-15a-5p and miR-16-5p. Signal Transduct. Target. Ther. 2022, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.C.; Cheng, Y.Y.; Williams, M.; Kirschner, M.B.; Madore, J.; Lum, T.; Sarun, K.H.; Linton, A.; McCaughan, B.; Klebe, S.; et al. Tumor Suppressor microRNAs Contribute to the Regulation of PD-L1 Expression in Malignant Pleural Mesothelioma. J. Thorac. Oncol. 2017, 12, 1421–1433. [Google Scholar] [CrossRef]
- van Zandwijk, N.; Pavlakis, N.; Kao, S.C.; Linton, A.; Boyer, M.J.; Clarke, S.; Huynh, Y.; Chrzanowska, A.; Fulham, M.J.; Bailey, D.L.; et al. Safety and Activity of microRNA-Loaded Minicells in Patients with Recurrent Malignant Pleural Mesothelioma: A First-in-Man, Phase 1, Open-Label, Dose-Escalation Study. Lancet Oncol. 2017, 18, 1386–1396. [Google Scholar] [CrossRef]
- Cui, Y.; Shen, T.; Xu, F.; Zhang, J.; Wang, Y.; Wu, J.; Bu, H.; Fu, D.; Fang, B.; Lv, H.; et al. KCNN4 May Weaken Anti-Tumor Immune Response via Raising Tregs and Diminishing Resting Mast Cells in Clear Cell Renal Cell Carcinoma. Cancer Cell Int. 2022, 22, 211. [Google Scholar] [CrossRef]
- Chiang, E.Y.; Li, T.; Jeet, S.; Peng, I.; Zhang, J.; Lee, W.P.; DeVoss, J.; Caplazi, P.; Chen, J.; Warming, S.; et al. Potassium Channels Kv1.3 and KCa3.1 Cooperatively and Compensatorily Regulate Antigen-Specific Memory T Cell Functions. Nat. Commun. 2017, 8, 14644. [Google Scholar] [CrossRef]
- Segovia, M.; Russo, S.; Jeldres, M.; Mahmoud, Y.D.; Perez, V.; Duhalde, M.; Charnet, P.; Rousset, M.; Victoria, S.; Veigas, F.; et al. Targeting TMEM176B Enhances Antitumor Immunity and Augments the Efficacy of Immune Checkpoint Blockers by Unleashing Inflammasome Activation. Cancer Cell 2019, 35, 767–781. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Li, Q.; Luo, M.; Zhong, Z.; Zhou, S.; Jiang, L.; Shen, N.; Geng, Z.; Cheng, H.; Meng, L.; et al. Leukemia Cell-Derived Microvesicles Induce T Cell Exhaustion via miRNA Delivery. Oncoimmunology 2018, 7, e1448330. [Google Scholar] [CrossRef] [PubMed]
- Quagliariello, V.; Passariello, M.; Di Mauro, A.; Cipullo, C.; Paccone, A.; Barbieri, A.; Palma, G.; Luciano, A.; Buccolo, S.; Bisceglia, I.; et al. Immune Checkpoint Inhibitor Therapy Increases Systemic SDF-1, Cardiac DAMPs Fibronectin-EDA, S100/Calgranulin, Galectine-3, and NLRP3-MyD88-Chemokine Pathways. Front. Cardiovasc. Med. 2022, 9, 930797. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A Costimulatory Molecule for T Cell Activation and IFN-Gamma Production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef]
- Yang, S.; Wei, W.; Zhao, Q. B7-H3, a Checkpoint Molecule, as a Target for Cancer Immunotherapy. Int. J. Biol. Sci. 2020, 16, 1767–1773. [Google Scholar] [CrossRef]
- He, Y.; Sun, M.M.; Zhang, G.G.; Yang, J.; Chen, K.S.; Xu, W.W.; Li, B. Targeting PI3K/Akt Signal Transduction for Cancer Therapy. Signal Transduct. Target. Ther. 2021, 6, 425. [Google Scholar] [CrossRef]
- Zhu, J.; Chen, L.; Zou, L.; Yang, P.; Wu, R.; Mao, Y.; Zhou, H.; Li, R.; Wang, K.; Wang, W.; et al. MiR-20b, -21, and -130b Inhibit PTEN Expression Resulting in B7-H1 over-Expression in Advanced Colorectal Cancer. Hum. Immunol. 2014, 75, 348–353. [Google Scholar] [CrossRef]
- Dastmalchi, N.; Hosseinpourfeizi, M.A.; Khojasteh, S.M.B.; Baradaran, B.; Safaralizadeh, R. Tumor Suppressive Activity of miR-424-5p in Breast Cancer Cells through Targeting PD-L1 and Modulating PTEN/PI3K/AKT/mTOR Signaling Pathway. Life Sci. 2020, 259, 118239. [Google Scholar] [CrossRef]
- Wang, B.; Zhang, Q. The Expression and Clinical Significance of Circulating microRNA-21 in Serum of Five Solid Tumors. J. Cancer Res. Clin. Oncol. 2012, 138, 1659–1666. [Google Scholar] [CrossRef]
- Matsuhashi, S.; Manirujjaman, M.; Hamajima, H.; Ozaki, I. Control Mechanisms of the Tumor Suppressor PDCD4: Expression and Functions. Int. J. Mol. Sci. 2019, 20, 2304. [Google Scholar] [CrossRef]
- Betzler, A.C.; Theodoraki, M.-N.; Schuler, P.J.; Döscher, J.; Laban, S.; Hoffmann, T.K.; Brunner, C. NF-κB and Its Role in Checkpoint Control. Int. J. Mol. Sci. 2020, 21, 3949. [Google Scholar] [CrossRef]
- Svoronos, A.A.; Engelman, D.M.; Slack, F.J. OncomiR or Tumor Suppressor? The Duplicity of MicroRNAs in Cancer. Cancer Res. 2016, 76, 3666–3670. [Google Scholar] [CrossRef] [PubMed]
- Huber, V.; Vallacchi, V.; Fleming, V.; Hu, X.; Cova, A.; Dugo, M.; Shahaj, E.; Sulsenti, R.; Vergani, E.; Filipazzi, P.; et al. Tumor-Derived microRNAs Induce Myeloid Suppressor Cells and Predict Immunotherapy Resistance in Melanoma. J. Clin. Investig. 2018, 128, 5505–5516. [Google Scholar] [CrossRef]
- Kara, G.; Calin, G.A.; Ozpolat, B. RNAi-Based Therapeutics and Tumor Targeted Delivery in Cancer. Adv. Drug Deliv. Rev. 2022, 182, 114113. [Google Scholar] [CrossRef]
- Atwa, S.M.; Handoussa, H.; Hosny, K.M.; Odenthal, M.; El Tayebi, H.M. Pivotal Role of Long Non-Coding Ribonucleic Acid-X-Inactive Specific Transcript in Regulating Immune Checkpoint Programmed Death Ligand 1 through a Shared Pathway between miR-194-5p and miR-155-5p in Hepatocellular Carcinoma. World J. Hepatol. 2020, 12, 1211–1227. [Google Scholar] [CrossRef]
- Jiang, X.; Zhou, T.; Xiao, Y.; Yu, J.; Dou, S.; Chen, G.; Wang, R.; Xiao, H.; Hou, C.; Wang, W.; et al. Tim-3 Promotes Tumor-Promoting M2 Macrophage Polarization by Binding to STAT1 and Suppressing the STAT1-miR-155 Signaling Axis. Oncoimmunology 2016, 5, e1211219. [Google Scholar] [CrossRef]
- Dudda, J.C.; Salaun, B.; Ji, Y.; Palmer, D.C.; Monnot, G.C.; Merck, E.; Boudousquie, C.; Utzschneider, D.T.; Escobar, T.M.; Perret, R.; et al. MicroRNA-155 Is Required for Effector CD8+ T Cell Responses to Virus Infection and Cancer. Immunity 2013, 38, 742–753. [Google Scholar] [CrossRef]
- Huffaker, T.B.; Hu, R.; Runtsch, M.C.; Bake, E.; Chen, X.; Zhao, J.; Round, J.L.; Baltimore, D.; O’Connell, R.M. Epistasis between microRNAs 155 and 146a during T Cell-Mediated Antitumor Immunity. Cell Rep. 2012, 2, 1697–1709. [Google Scholar] [CrossRef] [PubMed]
- Jonas, K.; Prinz, F.; Ferracin, M.; Krajina, K.; Pasculli, B.; Deutsch, A.; Madl, T.; Rinner, B.; Slaby, O.; Klec, C.; et al. MiR-4649-5p Acts as a Tumor-Suppressive microRNA in Triple Negative Breast Cancer by Direct Interaction with PIP5K1C, Thereby Potentiating Growth-Inhibitory Effects of the AKT Inhibitor Capivasertib. Breast Cancer Res. 2023, 25, 119. [Google Scholar] [CrossRef]
- Afra, F.; Mahboobipour, A.A.; Salehi Farid, A.; Ala, M. Recent Progress in the Immunotherapy of Hepatocellular Carcinoma: Non-Coding RNA-Based Immunotherapy May Improve the Outcome. Biomed. Pharmacother. 2023, 165, 115104. [Google Scholar] [CrossRef]
- Saini, S.; Yamamura, S.; Majid, S.; Shahryari, V.; Hirata, H.; Tanaka, Y.; Dahiya, R. MicroRNA-708 Induces Apoptosis and Suppresses Tumorigenicity in Renal Cancer Cells. Cancer Res. 2011, 71, 6208–6219. [Google Scholar] [CrossRef]
- de Oliveira, J.C.; Scrideli, C.A.; Brassesco, M.S.; Yunes, J.A.; Brandalise, S.R.; Tone, L.G. MiR-708-5p Is Differentially Expressed in Childhood Acute Lymphoblastic Leukemia but Not Strongly Associated to Clinical Features. Pediatr. Blood Cancer 2015, 62, 177–178. [Google Scholar] [CrossRef]
- Khan, M.J.; Singh, P.; Dohare, R.; Jha, R.; Rahmani, A.H.; Almatroodi, S.A.; Ali, S.; Syed, M.A. Inhibition of miRNA-34a Promotes M2 Macrophage Polarization and Improves LPS-Induced Lung Injury by Targeting Klf4. Genes 2020, 11, 966. [Google Scholar] [CrossRef]
- Zhong, L.; Altan, M.; Shannon, V.R.; Sheshadri, A. Immune-Related Adverse Events: Pneumonitis. Immunotherapy 2020, 1244, 255–269. [Google Scholar] [CrossRef]
- Mockly, S.; Seitz, H. Synthetic miR-34a against Solid Tumours: A Predictable Failure. Br. J. Cancer 2023, 128, 478–480. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.S.; Kang, Y.-K.; Borad, M.; Sachdev, J.; Ejadi, S.; Lim, H.Y.; Brenner, A.J.; Park, K.; Lee, J.-L.; Kim, T.-Y.; et al. Phase 1 Study of MRX34, a Liposomal miR-34a Mimic, in Patients with Advanced Solid Tumours. Br. J. Cancer 2020, 122, 1630–1637. [Google Scholar] [CrossRef]
- Ivanova, E.; Asadullina, D.; Rakhimov, R.; Izmailov, A.; Izmailov, A.; Gilyazova, G.; Galimov, S.; Pavlov, V.; Khusnutdinova, E.; Gilyazova, I. Exosomal miRNA-146a Is Downregulated in Clear Cell Renal Cell Carcinoma Patients with Severe Immune-Related Adverse Events. Non-Coding RNA Res. 2022, 7, 159–163. [Google Scholar] [CrossRef]
- Shimada, B.K.; Yang, Y.; Zhu, J.; Wang, S.; Suen, A.; Kronstadt, S.M.; Jeyaram, A.; Jay, S.M.; Zou, L.; Chao, W. Extracellular miR-146a-5p Induces Cardiac Innate Immune Response and Cardiomyocyte Dysfunction. Immunohorizons 2020, 4, 561–572. [Google Scholar] [CrossRef]
- Mann, M.; Mehta, A.; Zhao, J.L.; Lee, K.; Marinov, G.K.; Garcia-Flores, Y.; Lu, L.-F.; Rudensky, A.Y.; Baltimore, D. An NF-κB-microRNA Regulatory Network Tunes Macrophage Inflammatory Responses. Nat. Commun. 2017, 8, 851. [Google Scholar] [CrossRef]
- Li, M.; Shan, W.; Hong, B.; Zou, J.; Li, H.; Han, D.; Zhang, Y.; Li, L.; Li, D.; Lin, W. Circulating miR-92b and miR-375 for Monitoring the Chemoresistance and Prognosis of Small Cell Lung Cancer. Sci. Rep. 2020, 10, 12705. [Google Scholar] [CrossRef]
- Ahmad, N.; Haider, S.; Jagannathan, S.; Anaissie, E.; Driscoll, J.J. MicroRNA Theragnostics for the Clinical Management of Multiple Myeloma. Leukemia 2014, 28, 732–738. [Google Scholar] [CrossRef]
- García-Giménez, J.L.; Mena-Mollá, S.; Beltrán-García, J.; Sanchis-Gomar, F. Challenges in the Analysis of Epigenetic Biomarkers in Clinical Samples. Clin. Chem. Lab. Med. 2017, 55, 1474–1477. [Google Scholar] [CrossRef] [PubMed]
- Androvic, P.; Benesova, S.; Rohlova, E.; Kubista, M.; Valihrach, L. Small RNA-Sequencing for Analysis of Circulating miRNAs: Benchmark Study. J. Mol. Diagn. 2022, 24, 386–394. [Google Scholar] [CrossRef] [PubMed]
- Fawzy, M.S.; Ibrahiem, A.T.; Bayomy, N.A.; Makhdoom, A.K.; Alanazi, K.S.; Alanazi, A.M.; Mukhlef, A.M.; Toraih, E.A. MicroRNA-155 and Disease-Related Immunohistochemical Parameters in Cutaneous Melanoma. Diagnostics 2023, 13, 1205. [Google Scholar] [CrossRef]
- Jiang, S.; Zhang, H.-W.; Lu, M.-H.; He, X.-H.; Li, Y.; Gu, H.; Liu, M.-F.; Wang, E.-D. MicroRNA-155 Functions as an OncomiR in Breast Cancer by Targeting the Suppressor of Cytokine Signaling 1 Gene. Cancer Res. 2010, 70, 3119–3127. [Google Scholar] [CrossRef]
- Yang, Y.; Liu, H.; Chen, Y.; Xiao, N.; Zheng, Z.; Liu, H.; Wan, J. Liquid Biopsy on the Horizon in Immunotherapy of Non-Small Cell Lung Cancer: Current Status, Challenges, and Perspectives. Cell Death Dis. 2023, 14, 230. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Giménez, J.L.; Saadi, W.; Ortega, A.L.; Lahoz, A.; Suay, G.; Carretero, J.; Pereda, J.; Fatmi, A.; Pallardó, F.V.; Mena-Molla, S. miRNAs Related to Immune Checkpoint Inhibitor Response: A Systematic Review. Int. J. Mol. Sci. 2024, 25, 1737. https://doi.org/10.3390/ijms25031737
García-Giménez JL, Saadi W, Ortega AL, Lahoz A, Suay G, Carretero J, Pereda J, Fatmi A, Pallardó FV, Mena-Molla S. miRNAs Related to Immune Checkpoint Inhibitor Response: A Systematic Review. International Journal of Molecular Sciences. 2024; 25(3):1737. https://doi.org/10.3390/ijms25031737
Chicago/Turabian StyleGarcía-Giménez, José Luis, Wiam Saadi, Angel L. Ortega, Agustin Lahoz, Guillermo Suay, Julián Carretero, Javier Pereda, Ahlam Fatmi, Federico V. Pallardó, and Salvador Mena-Molla. 2024. "miRNAs Related to Immune Checkpoint Inhibitor Response: A Systematic Review" International Journal of Molecular Sciences 25, no. 3: 1737. https://doi.org/10.3390/ijms25031737