
The WMI Rat of Premature Cognitive Aging Presents Intrinsic Vulnerability to Oxidative Stress in Primary Neurons and Astrocytes Compared to Its Nearly Isogenic WLI Control
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
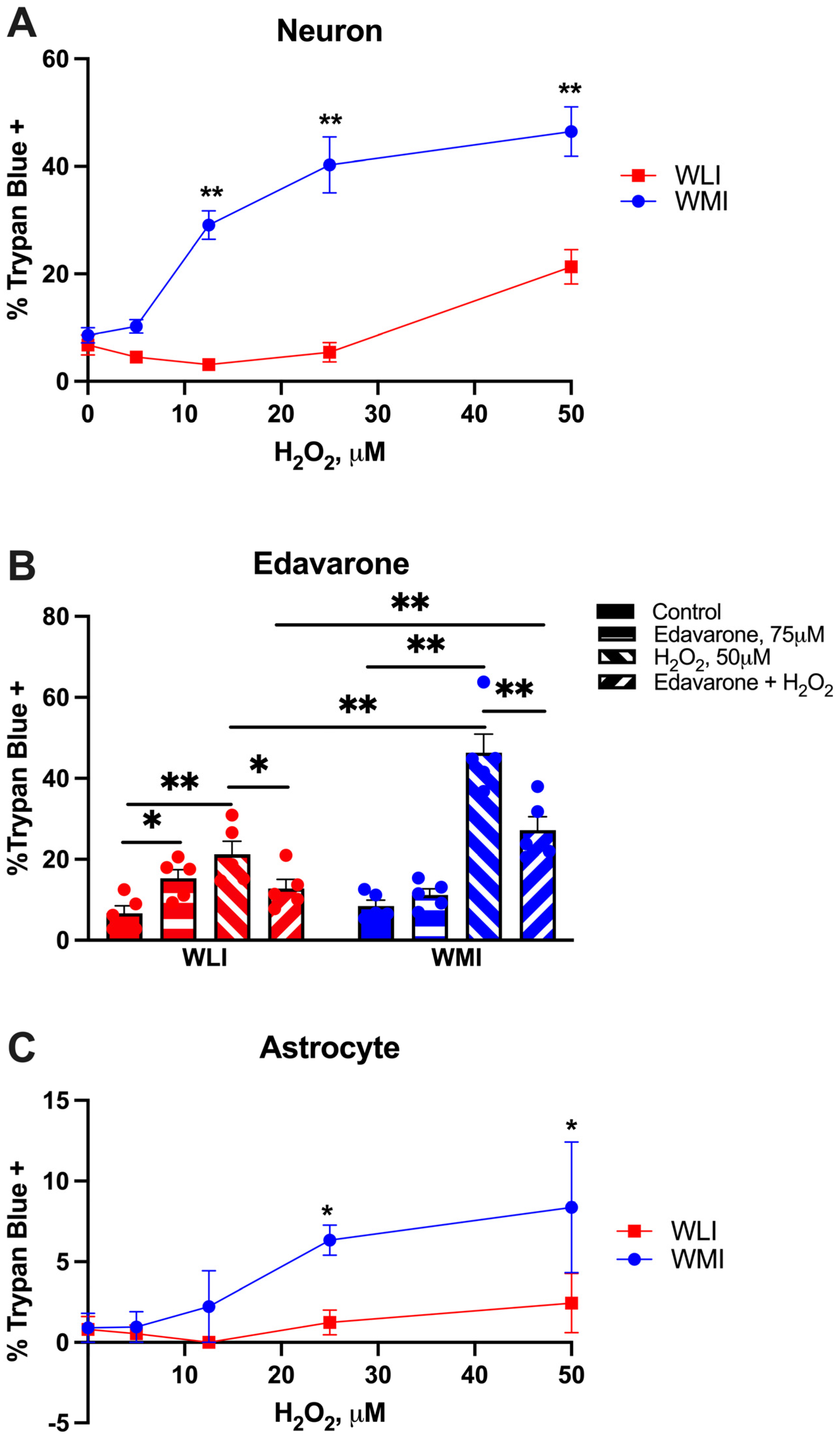
2.1. Cell Viability in Response to H2O2
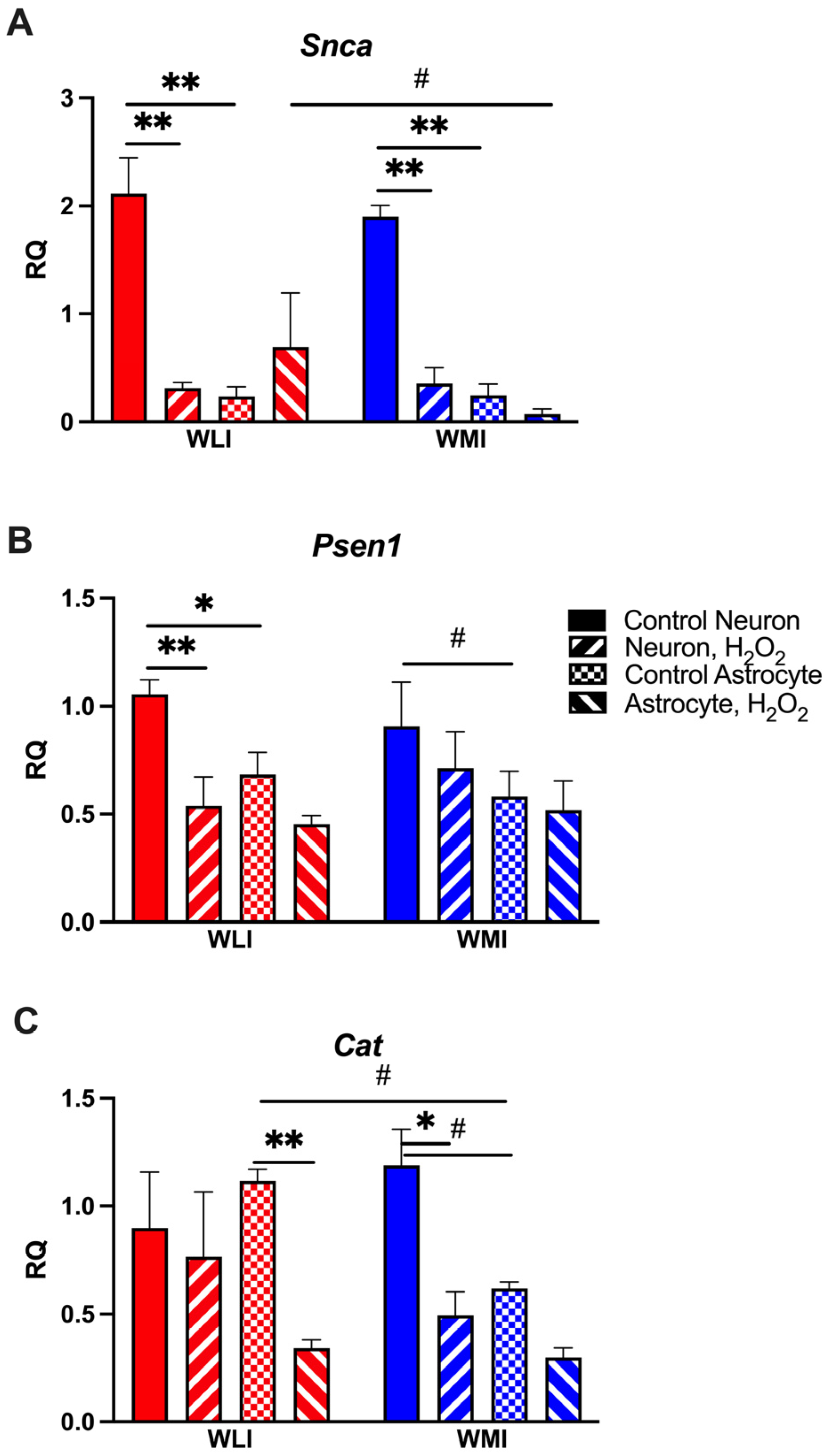
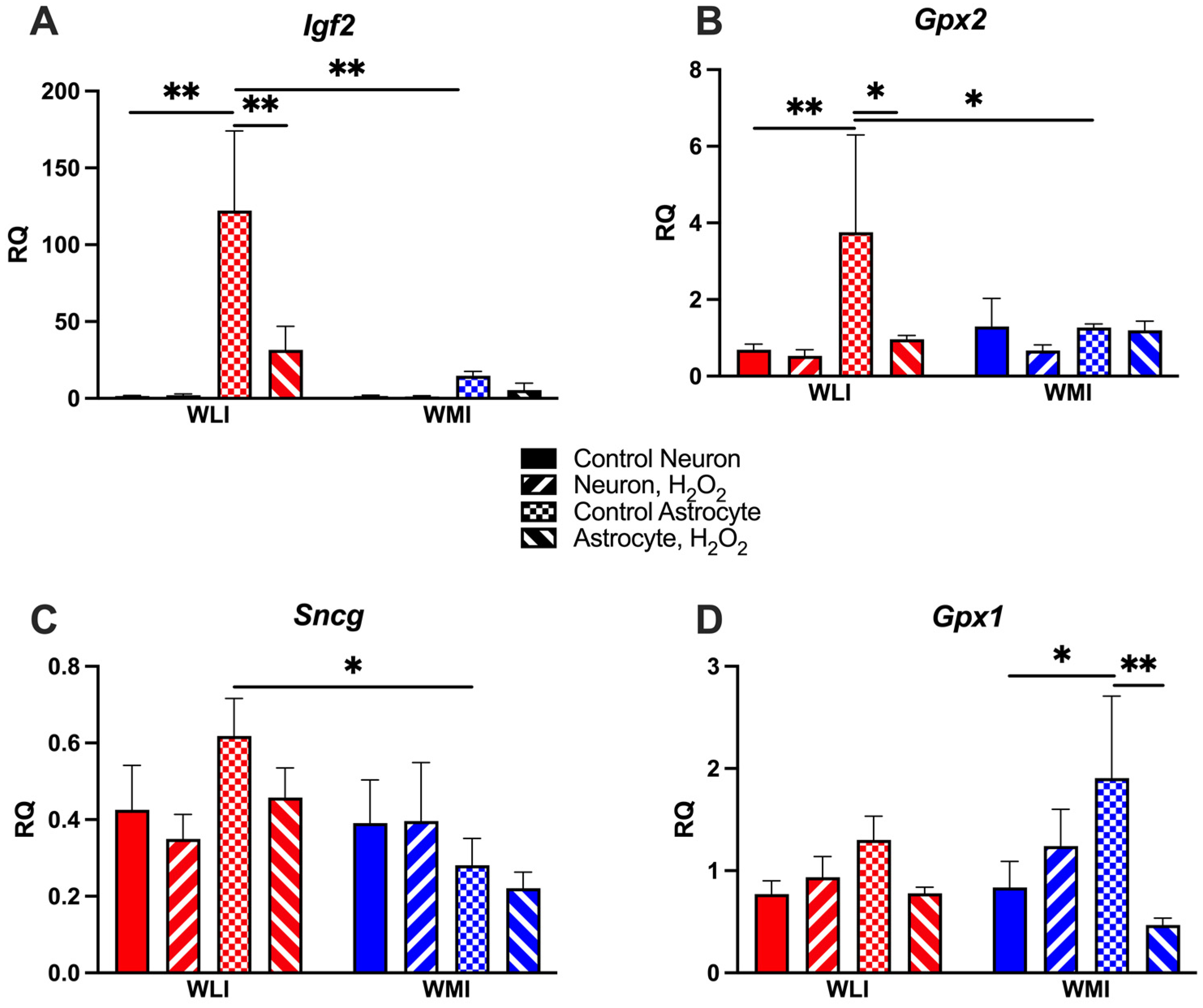
2.2. Neuron-Specific Gene Expression Responses to OS
2.3. Astrocyte-Specific Gene Expression Responses to OS
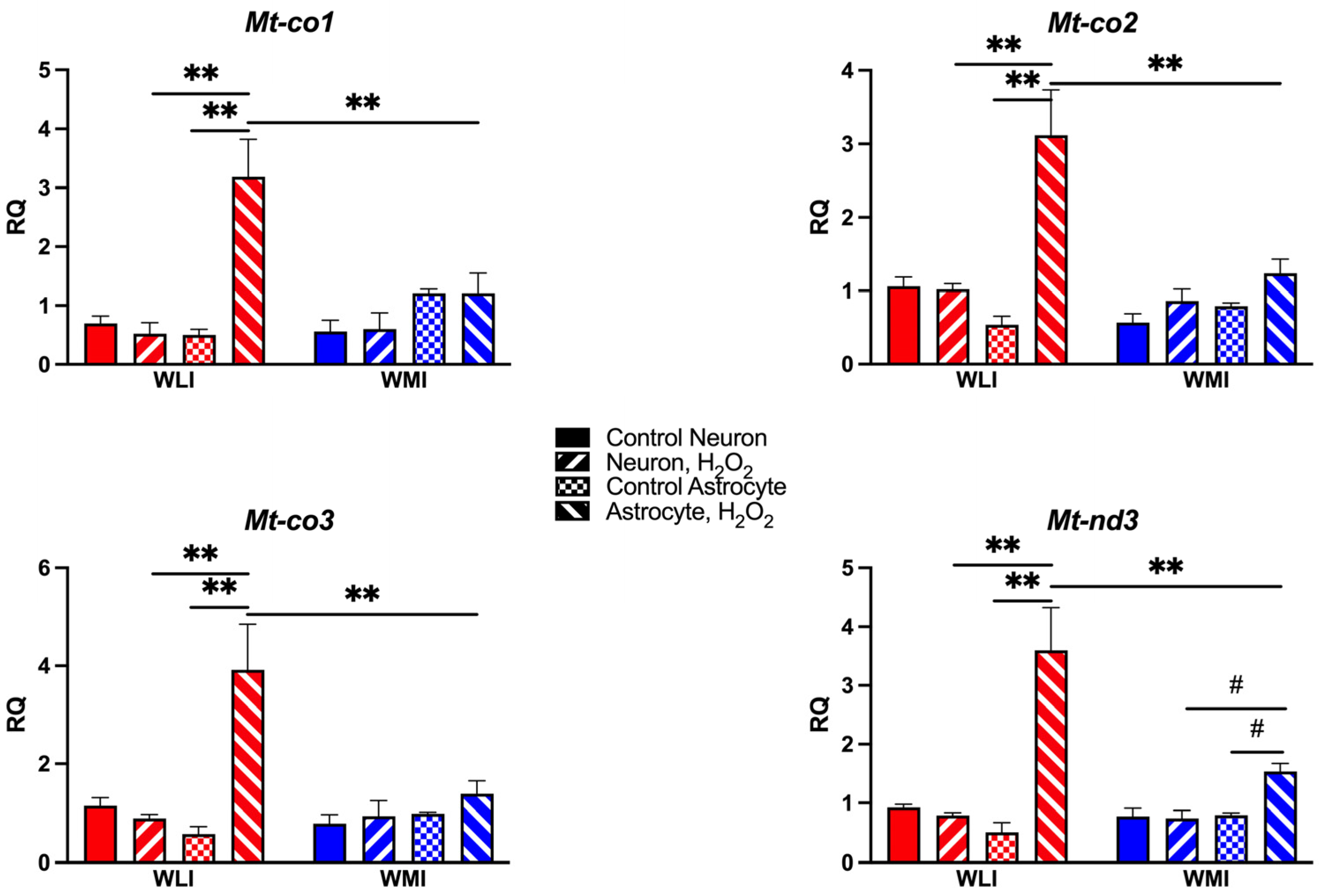
2.4. Mitochondrial Gene Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Hippocampal Culture Preparation
4.3. Preparation of Astrocyte Cultures
4.4. Preparation of N2 Astrocyte-Conditioned Medium
4.5. Drug Treatments
4.6. Determination of Cell Survival
4.7. RNA Extraction and Rreverse Transcription
4.8. Real-Time Reverse Transcription-Polymerase Chain Reaction [RT-qPCR]
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Weger, M.; Alpern, D.; Cherix, A.; Ghosal, S.; Grosse, J.; Russeil, J.; Gruetter, R.; de Kloet, E.R.; Deplancke, B.; Sandi, C. Mitochondrial gene signature in the prefrontal cortex for differential susceptibility to chronic stress. Sci. Rep. 2020, 10, 18308. [Google Scholar] [CrossRef]
- Lindqvist, D.; Dhabhar, F.S.; James, S.J.; Hough, C.M.; Jain, F.A.; Bersani, F.S.; Reus, V.I.; Verhoeven, J.E.; Epel, E.S.; Mahan, L.; et al. Oxidative stress, inflammation and treatment response in major depression. Psychoneuroendocrinology 2017, 76, 197–205. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress [O&NS] pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar] [CrossRef]
- Beal, M.F. Oxidatively modified proteins in aging and disease. Free Radic. Biol. Med. 2002, 32, 797–803. [Google Scholar] [CrossRef]
- Lovell, M.A.; Markesbery, W.R. Oxidative damage in mild cognitive impairment and early Alzheimer’s disease. J. Neurosci. Res. 2007, 85, 3036–3040. [Google Scholar] [CrossRef] [PubMed]
- Petersen, R.C.; Thomas, R.G.; Grundman, M.; Bennett, D.; Doody, R.; Ferris, S.; Galasko, D.; Jin, S.; Kaye, J.; Levey, A.; et al. Vitamin E and Donepezil for the Treatment of Mild Cognitive Impairment. N. Engl. J. Med. 2005, 352, 2379–2388. [Google Scholar] [CrossRef]
- Mecocci, P.; Boccardi, V.; Cecchetti, R.; Bastiani, P.; Scamosci, M.; Ruggiero, C.; Baroni, M. A Long Journey into Aging, Brain Aging, and Alzheimer’s Disease Following the Oxidative Stress Tracks. J. Alzheimer’s Dis. 2018, 62, 1319–1335. [Google Scholar] [CrossRef]
- Lim, P.H.; Wert, S.L.; Tunc-Ozcan, E.; Marr, R.; Ferreira, A.; Redei, E.E. Premature hippocampus-dependent memory decline in middle-aged females of a genetic rat model of depression. Behav. Brain Res. 2018, 353, 242–249. [Google Scholar] [CrossRef]
- Will, C.C.; Aird, F.; Redei, E.E. Selectively bred Wistar–Kyoto rats: An animal model of depression and hyper-responsiveness to antidepressants. Mol. Psychiatry 2003, 8, 925–932. [Google Scholar] [CrossRef]
- de Jong, T.V.; Kim, P.; Guryev, V.; Mulligan, M.K.; Williams, R.W.; Redei, E.E.; Chen, H. Whole genome sequencing of nearly isogenic WMI and WLI inbred rats identifies genes potentially involved in depression and stress reactivity. Sci. Rep. 2021, 11, 14774. [Google Scholar] [CrossRef] [PubMed]
- Redei, E.E.; Udell, M.E.; Woods, L.C.S.; Chen, H. The Wistar Kyoto Rat: A Model of Depression Traits. Curr. Neuropharmacol. 2023, 21, 1884–1905. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Pryzbyl, K.J.; Bigio, E.H.; Weintraub, S.; Mesulam, M.-M.; Redei, E.E. Reduced Hippocampal and Anterior Cingulate Expression of Antioxidant Enzymes and Membrane Progesterone Receptors in Alzheimer’s Disease with Depression. J. Alzheimer’s Dis. 2022, 89, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.; Delgado-Esteban, M.; Bolaños, J.P.; Medina, J.M. Oxygen and glucose deprivation induces mitochondrial dysfunction and oxidative stress in neurones but not in astrocytes in primary culture. J. Neurochem. 2002, 81, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Günther, M.; Al Nimer, F.; Piehl, F.; Risling, M.; Mathiesen, T. Susceptibility to Oxidative Stress Is Determined by Genetic Background in Neuronal Cell Cultures. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed]
- Nemesh, M.; Ji, M.T.; Przybyl, K.J.; Harter, A.M.; Kim, C.S.; Yamazaki, A.; Redei, E.E. Enriched Environment Reversed Learning and Memory Deficit of Genetically Stress Reactive Female WMI Rats at Middle Age; Program No. 664.15. 2022 Neuroscience Meeting Planner; Society for Neuroscience: San Diego, CA, USA, 2022; Online. [Google Scholar]
- Han, F.-F.; Wang, H.-X.; Wu, J.-J.; Yao, W.; Hao, C.-F.; Pei, J.-J. Depressive symptoms and cognitive impairment: A 10-year follow-up study from the Survey of Health, Ageing and Retirement in Europe. Eur. Psychiatry 2021, 64, 55. [Google Scholar] [CrossRef] [PubMed]
- Richmond-Rakerd, L.S.; D’souza, S.; Milne, B.J.; Caspi, A.; Moffitt, T.E. Longitudinal Associations of Mental Disorders With Dementia: 30-Year Analysis of 1.7 Million New Zealand Citizens. JAMA Psychiatry 2022, 79, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Caruso, G.; Grasso, M.; Fidilio, A.; Torrisi, S.A.; Musso, N.; Geraci, F.; Tropea, M.R.; Privitera, A.; Tascedda, F.; Puzzo, D.; et al. Antioxidant Activity of Fluoxetine and Vortioxetine in a Non-Transgenic Animal Model of Alzheimer’s Disease. Front. Pharmacol. 2021, 12, 809541. [Google Scholar] [CrossRef]
- Eid, R.S.; Gobinath, A.R.; Galea, L.A.M. Sex differences in depression: Insights from clinical and preclinical studies. Prog. Neurobiol. 2019, 176, 86–102. [Google Scholar] [CrossRef]
- Schuch, J.J.; Roest, A.M.; Nolen, W.A.; Penninx, B.W.; de Jonge, P. Gender differences in major depressive disorder: Results from the Netherlands study of depression and anxiety. J. Affect. Disord. 2014, 156, 156–163. [Google Scholar] [CrossRef]
- Ferretti, M.T.; Iulita, M.F.; Cavedo, E.; Chiesa, P.A.; Schumacher Dimech, A.; Santuccione Chadha, A.; Baracchi, F.; Girouard, H.; Misoch, S.; Giacobini, E.; et al. Sex differences in Alzheimer disease—The gateway to precision medicine. Nat. Rev. Neurol. 2018, 14, 457–469. [Google Scholar] [CrossRef]
- Zhu, D.; Montagne, A.; Zhao, Z. Alzheimer’s pathogenic mechanisms and underlying sex difference. Cell. Mol. Life Sci. 2021, 78, 4907–4920. [Google Scholar] [CrossRef]
- Sundermann, E.E.; Katz, M.J.; Lipton, R.B. Sex Differences in the Relationship between Depressive Symptoms and Risk of Amnestic Mild Cognitive Impairment. Am. J. Geriatr. Psychiatry 2016, 25, 13–22. [Google Scholar] [CrossRef]
- Russell, J.K.; Jones, C.K.; Newhouse, P.A. The Role of Estrogen in Brain and Cognitive Aging. Neurotherapeutics 2019, 16, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Bove, R.; Secor, E.; Chibnik, L.B.; Barnes, L.L.; Schneider, J.A.; Bennett, D.A.; De Jager, P.L. Age at surgical menopause influences cognitive decline and Alzheimer pathology in older women. Neurology 2014, 82, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Correia, S.C.; Santos, R.X.; Cardoso, S.; Carvalho, C.; Santos, M.S.; Oliveira, C.R.; Moreira, P.I. Effects of Estrogen in the Brain: Is it a Neuroprotective Agent in Alzheimers Disease? Curr. Aging Sci. 2010, 3, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Suzuki, S.; Brown, C.M. Estradiol: A hormone with diverse and contradictory neuroprotective actions. Dialogues Clin. Neurosci. 2009, 11, 297–303. [Google Scholar] [CrossRef]
- Prokai-Tatrai, K.; Perjesi, P.; Rivera-Portalatin, N.M.; Simpkins, J.W.; Prokai, L. Mechanistic investigations on the antioxidant action of a neuroprotective estrogen derivative. Steroids 2008, 73, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Zárate, S.; Stevnsner, T.; Gredilla, R. Role of Estrogen and Other Sex Hormones in Brain Aging. Neuroprotection and DNA Repair. Front. Aging Neurosci. 2017, 9, 430. [Google Scholar] [CrossRef] [PubMed]
- Cha, S.J.; Kim, K. Effects of the Edaravone, a Drug Approved for the Treatment of Amyotrophic Lateral Sclerosis, on Mitochondrial Function and Neuroprotection. Antioxidants 2022, 11, 195. [Google Scholar] [CrossRef]
- Luo, L.; Song, Z.; Li, X.; Huiwang; Zeng, Y.; Qinwang; Meiqi; He, J. Efficacy and safety of edaravone in treatment of amyotrophic lateral sclerosis—A systematic review and meta-analysis. Neurol. Sci. 2019, 40, 235–241. [Google Scholar] [CrossRef]
- Song, Y.; Bei, Y.; Xiao, Y.; Tong, H.-D.; Wu, X.-Q.; Chen, M.-T. Edaravone, a free radical scavenger, protects neuronal cells’ mitochondria from ischemia by inactivating another new critical factor of the 5-lipoxygenase pathway affecting the arachidonic acid metabolism. Brain Res. 2018, 1690, 96–104. [Google Scholar] [CrossRef]
- Liu, Z.; Yang, C.; Meng, X.; Li, Z.; Lv, C.; Cao, P. Neuroprotection of edaravone on the hippocampus of kainate-induced epilepsy rats through Nrf2/HO-1 pathway. Neurochem. Int. 2018, 112, 159–165. [Google Scholar] [CrossRef]
- Pritam, P.; Deka, R.; Bhardwaj, A.; Srivastava, R.; Kumar, D.; Jha, A.K.; Jha, N.K.; Villa, C.; Jha, S.K. Antioxidants in Alzheimer’s Disease: Current Therapeutic Significance and Future Prospects. Biology 2022, 11, 212. [Google Scholar] [CrossRef]
- Wang, X.; Michaelis, E.K. Selective neuronal vulnerability to oxidative stress in the brain. Front. Aging Neurosci. 2010, 2, 12. [Google Scholar] [CrossRef]
- Baxter, P.S.; Hardingham, G.E. Adaptive regulation of the brain’s antioxidant defences by neurons and astrocytes. Free Radic. Biol. Med. 2016, 100, 147–152. [Google Scholar] [CrossRef]
- Pei, J.; Pan, X.; Wei, G.; Hua, Y. Research progress of glutathione peroxidase family [GPX] in redoxidation. Front. Pharmacol. 2023, 14, 1147414. [Google Scholar] [CrossRef]
- Malysheva, O.V.; Ordyan, N.E. Insulin-Like Growth Factor 2: New Roles for a Known Molecule. Neurosci. Behav. Physiol. 2022, 52, 175–182. [Google Scholar] [CrossRef]
- Huang, X.; Yan, P.; Song, X.; Zhang, S.; Deng, Y.; Huang, C.; Zhao, X.; Liu, S.; Cheng, X.; Liao, D. MT-CO1 expression in nine organs and tissues of different-aged MRL/lpr mice: Investigation of mitochondrial respiratory chain dysfunction at organ level in systemic lupus erythematosus pathogenesis. Arch. Rheumatol. 2022, 37, 504–516. [Google Scholar] [CrossRef] [PubMed]
- Herst, P.M.; Rowe, M.R.; Carson, G.M.; Berridge, M.V. Functional Mitochondria in Health and Disease. Front. Endocrinol. 2017, 8, 296. [Google Scholar] [CrossRef] [PubMed]
- Esteitie, N.; Hinttala, R.; Wibom, R.; Nilsson, H.; Hance, N.; Naess, K.; Teär-Fahnehjelm, K.; Von Döbeln, U.; Majamaa, K.; Larsson, N.G. Secondary metabolic effects in complex I deficiency. Ann. Neurol. 2005, 58, 544–552. [Google Scholar] [CrossRef]
- Bottenstein, J.E.; Sato, G.H. Growth of a rat neuroblastoma cell line in serum-free supplemented medium. Proc. Natl. Acad. Sci. USA 1979, 76, 514–517. [Google Scholar] [CrossRef]
- Ferreira, A.; Sinjoanu, R.C.; Nicholson, A.; Kleinschmidt, S. Aβ toxicity in primary cultured neurons. Methods Mol. Biol. 2011, 160, 141–154. [Google Scholar] [CrossRef]
- Ferreira, A.; Loomis, P.A. Isolation and culture of primary neural cells. In Cells: A Laboratory Manual; Spector, D., Goldman, R., Leinwand, L., Eds.; Cold Spring Harbor Laboratory Press: Woodbury, NY, USA, 1998; pp. 9.1–9.9. [Google Scholar]
- Gao, Z.; Huang, K.; Xu, H. Protective effects of flavonoids in the roots of scutellaria baicalensis georgi against hydrogen peroxide-induced oxidative stress in hs-sy5y cells. Pharmacol. Res. 2001, 43, 173–178. [Google Scholar] [CrossRef]
- Fisk, L.; Nalivaeva, N.N.; Boyle, J.P.; Peers, C.S.; Turner, A.J. Effects of Hypoxia and Oxidative Stress on Expression of Neprilysin in Human Neuroblastoma Cells and Rat Cortical Neurones and Astrocytes. Neurochem. Res. 2016, 32, 1741–1748. [Google Scholar] [CrossRef] [PubMed]
- Xing, G.; Evbuomwan, O.; Li, Z.; Sharma, P. In Vitro Cytoprotection Of Resveratrol Against H2O2-induced Oxidative Stress And Injury In Astrocytes. J. Behav. Ther. Ment. Health 2016, 1, 49–58. [Google Scholar] [CrossRef]
- Li, Q.; Qiu, Z.; Lu, Y.; Lu, P.; Wen, J.; Wang, K.; Zhao, X.; Li, R.; Zhang, H.; Zhang, Y.; et al. Edaravone protects primary-cultured rat cortical neurons from ketamine-induced apoptosis via reducing oxidative stress and activating PI3K/Akt signal pathway. Mol. Cell. Neurosci. 2019, 100, 103399. [Google Scholar] [CrossRef]
- Ying, H.S.; Gottron, F.J.; Choi, D.W. Assessment of Cell Viability in Primary Neuronal Cultures. Curr. Protoc. Neurosci. 2001, 7, 18. [Google Scholar] [CrossRef] [PubMed]
- Meckes, J.K.; Lim, P.H.; Wert, S.L.; Luo, W.; Gacek, S.A.; Platt, D.; Jankord, R.; Saar, K.; Redei, E.E. Brain region-specific expression of genes mapped within quantitative trait loci for behavioral responsiveness to acute stress in Fisher 344 and Wistar Kyoto male rats. PLoS ONE 2018, 13, e0194293. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, A.; Harter, A.; Afreen, S.; Kanai, K.; Batori, S.; Redei, E.E. The WMI Rat of Premature Cognitive Aging Presents Intrinsic Vulnerability to Oxidative Stress in Primary Neurons and Astrocytes Compared to Its Nearly Isogenic WLI Control. Int. J. Mol. Sci. 2024, 25, 1692. https://doi.org/10.3390/ijms25031692
Ferreira A, Harter A, Afreen S, Kanai K, Batori S, Redei EE. The WMI Rat of Premature Cognitive Aging Presents Intrinsic Vulnerability to Oxidative Stress in Primary Neurons and Astrocytes Compared to Its Nearly Isogenic WLI Control. International Journal of Molecular Sciences. 2024; 25(3):1692. https://doi.org/10.3390/ijms25031692
Chicago/Turabian StyleFerreira, Adriana, Aspen Harter, Sana Afreen, Karoly Kanai, Sandor Batori, and Eva E. Redei. 2024. "The WMI Rat of Premature Cognitive Aging Presents Intrinsic Vulnerability to Oxidative Stress in Primary Neurons and Astrocytes Compared to Its Nearly Isogenic WLI Control" International Journal of Molecular Sciences 25, no. 3: 1692. https://doi.org/10.3390/ijms25031692