
Neural Stem Cells Transplanted into Rhesus Monkey Cortical Traumatic Brain Injury Can Survive and Differentiate into Neurons

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Induction and Identification of NSCs
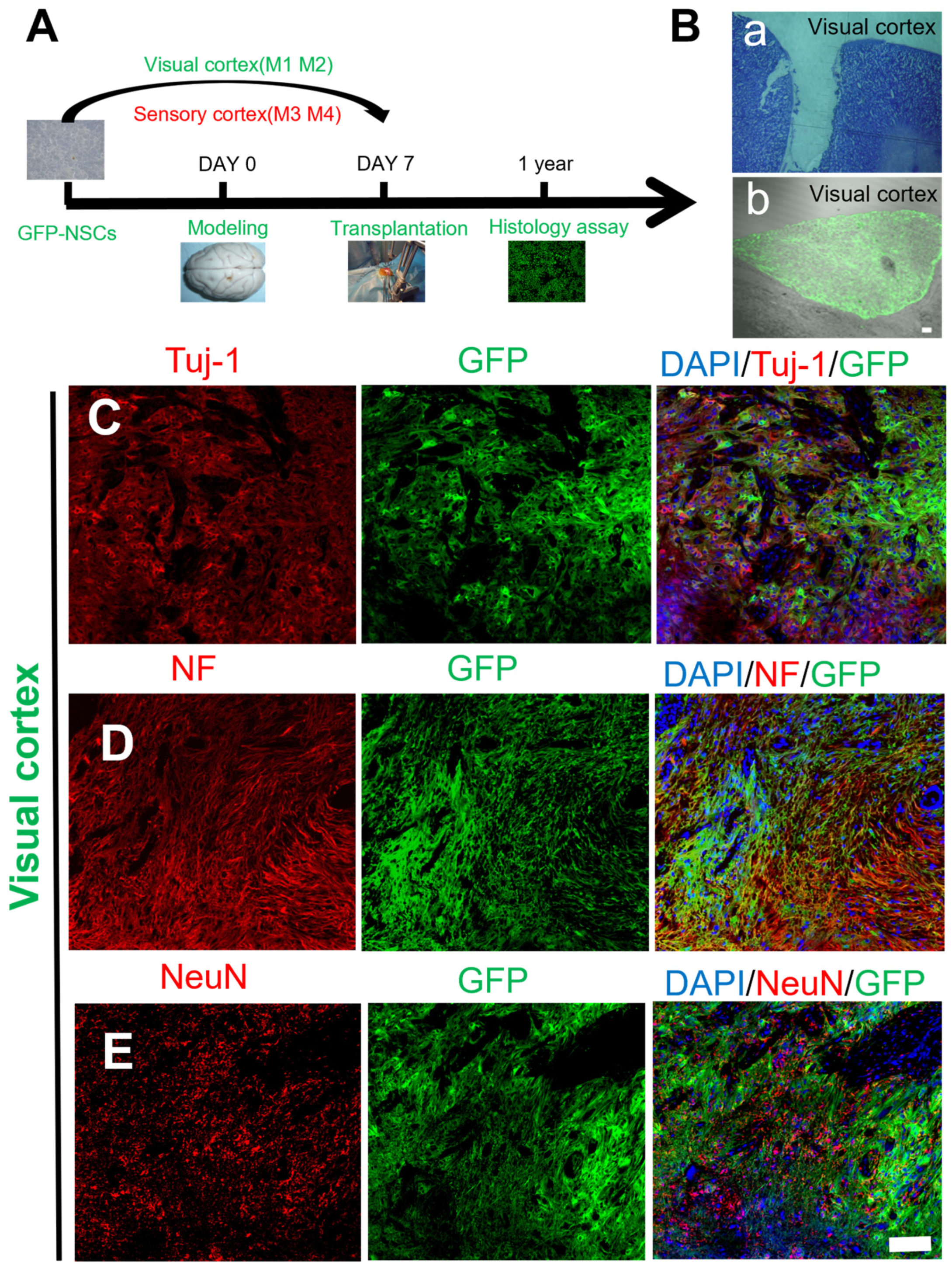
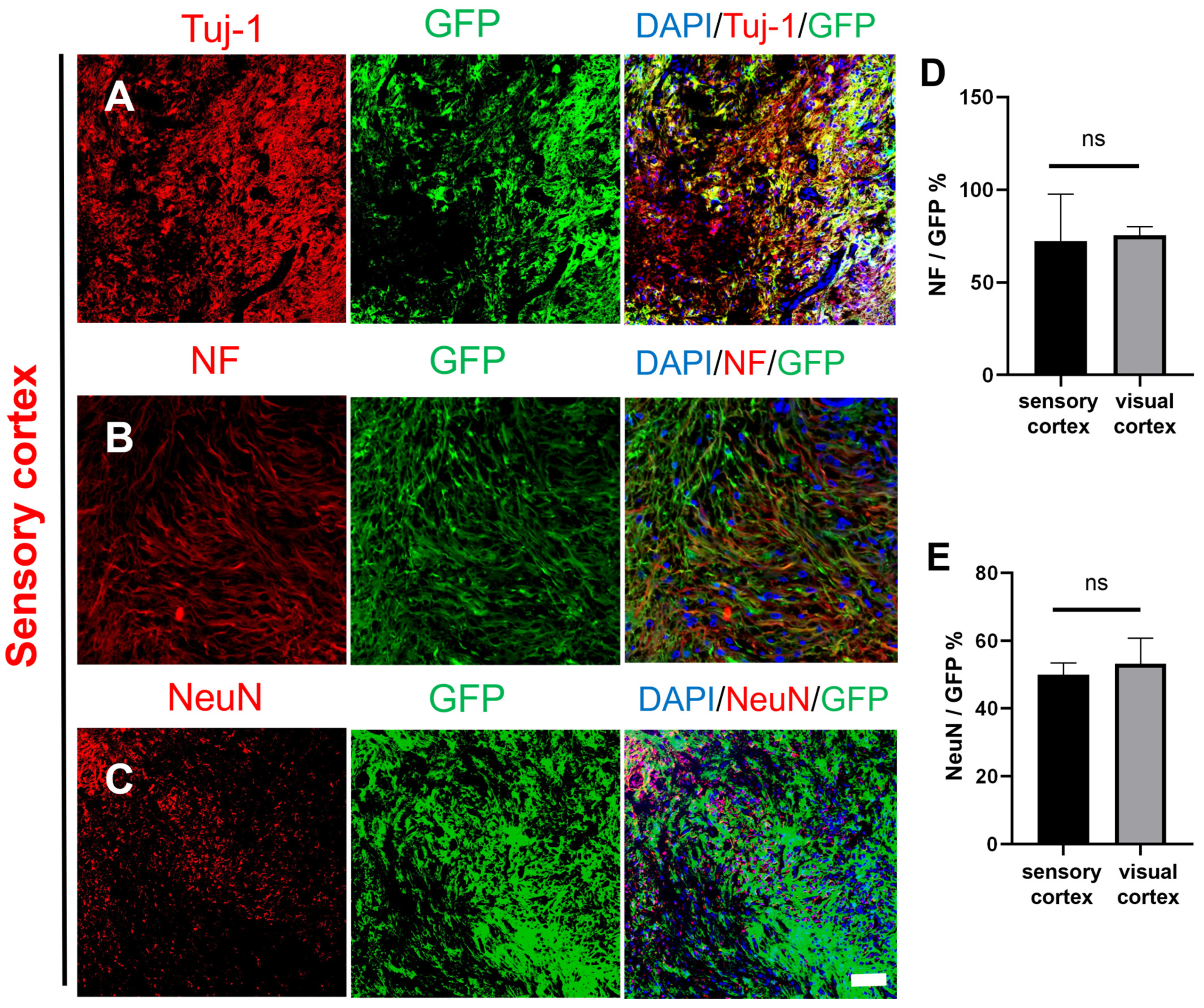
2.2. Survival and Differentiation of Grafted NSCs in the Site of TBI One Year after Transplantation
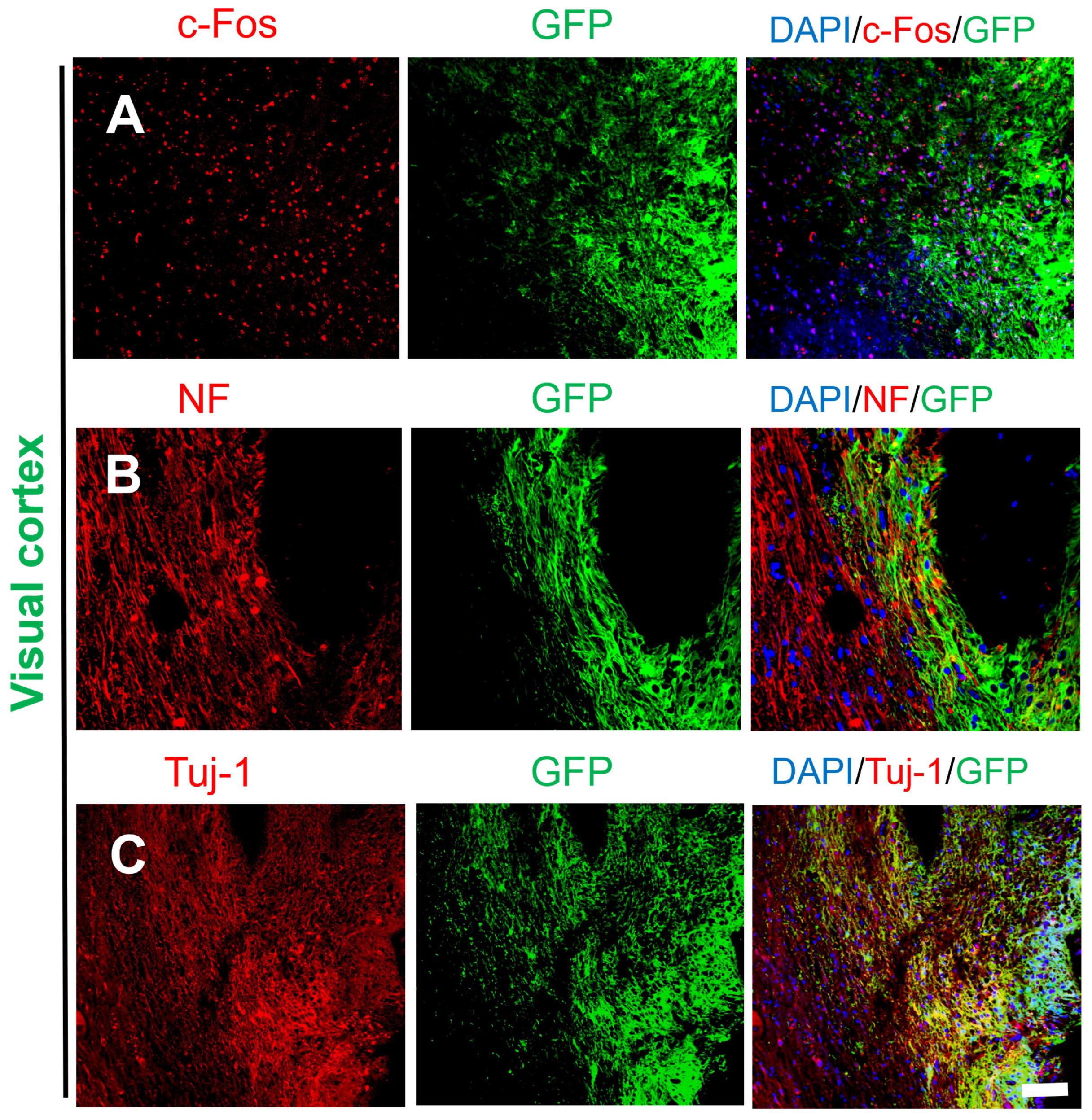
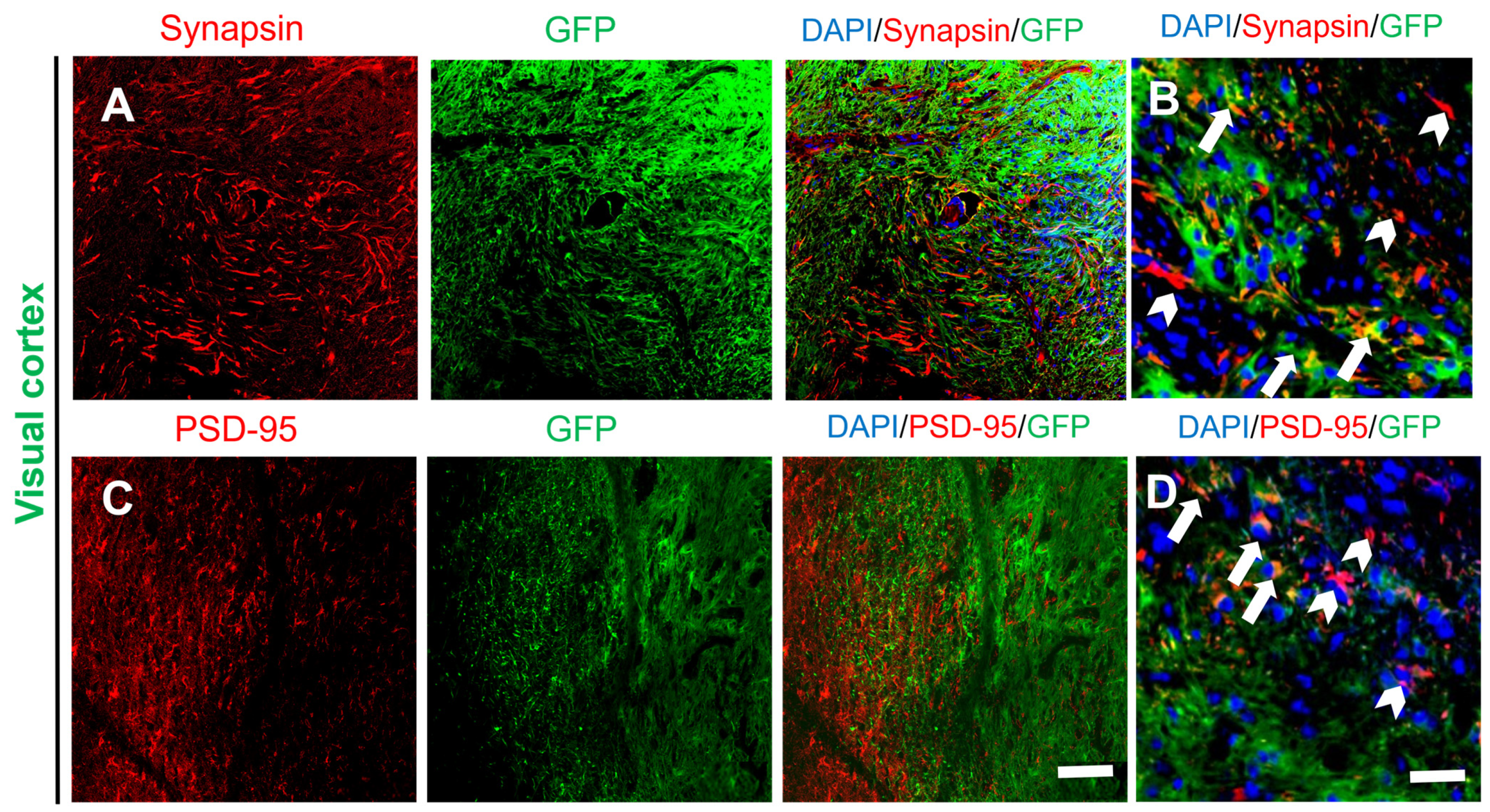
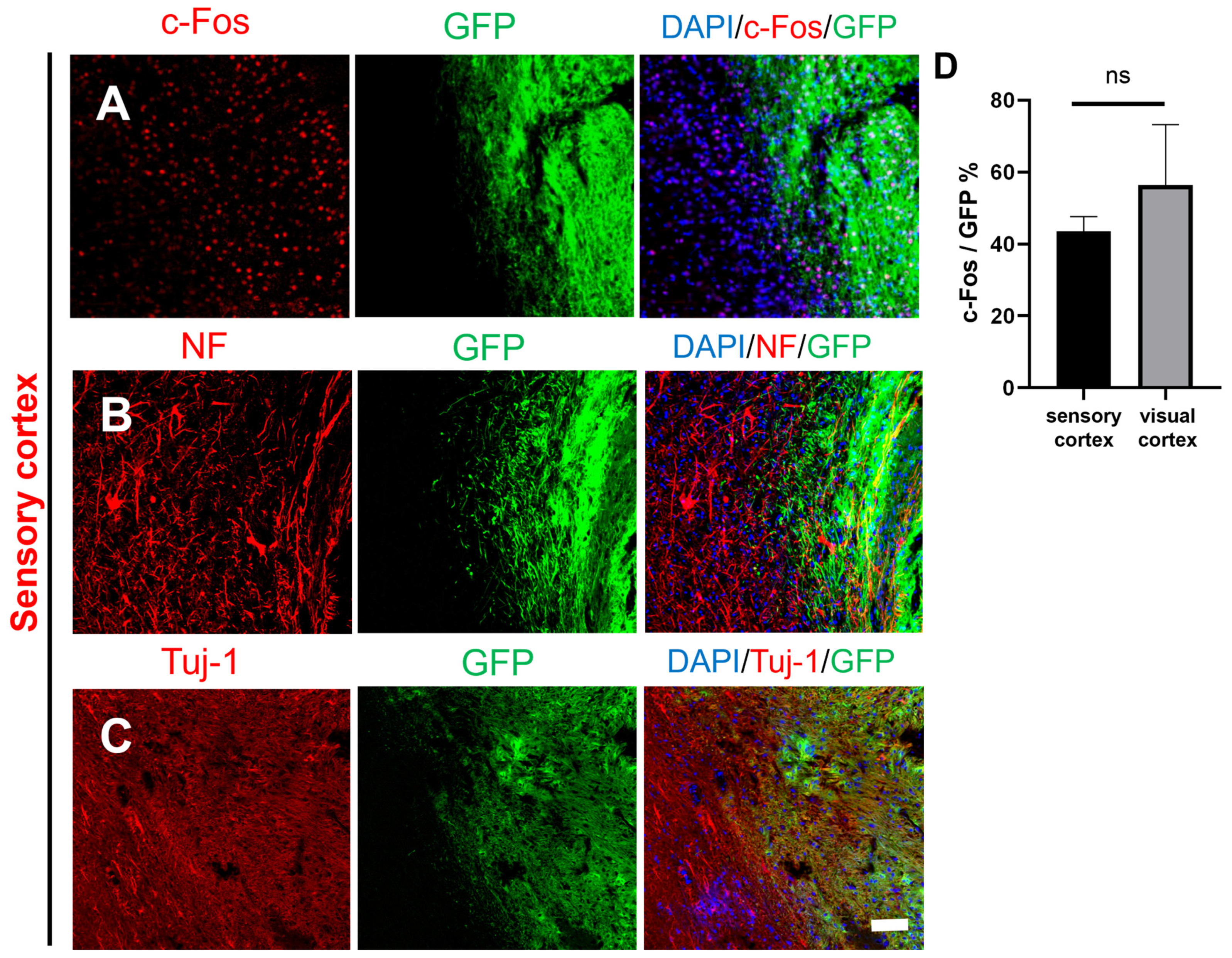
2.3. Activity and Migration of the Grafted NSCs
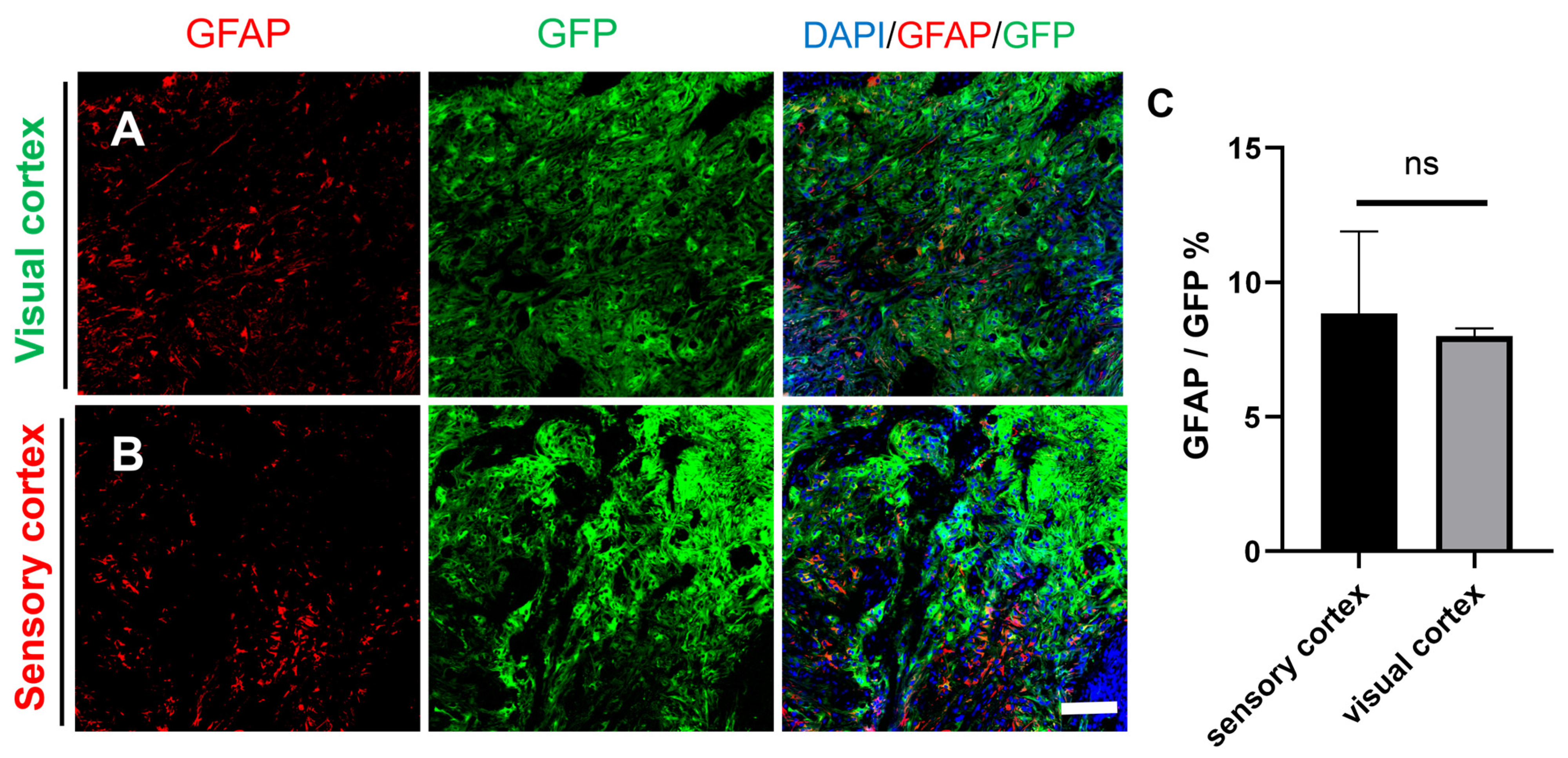
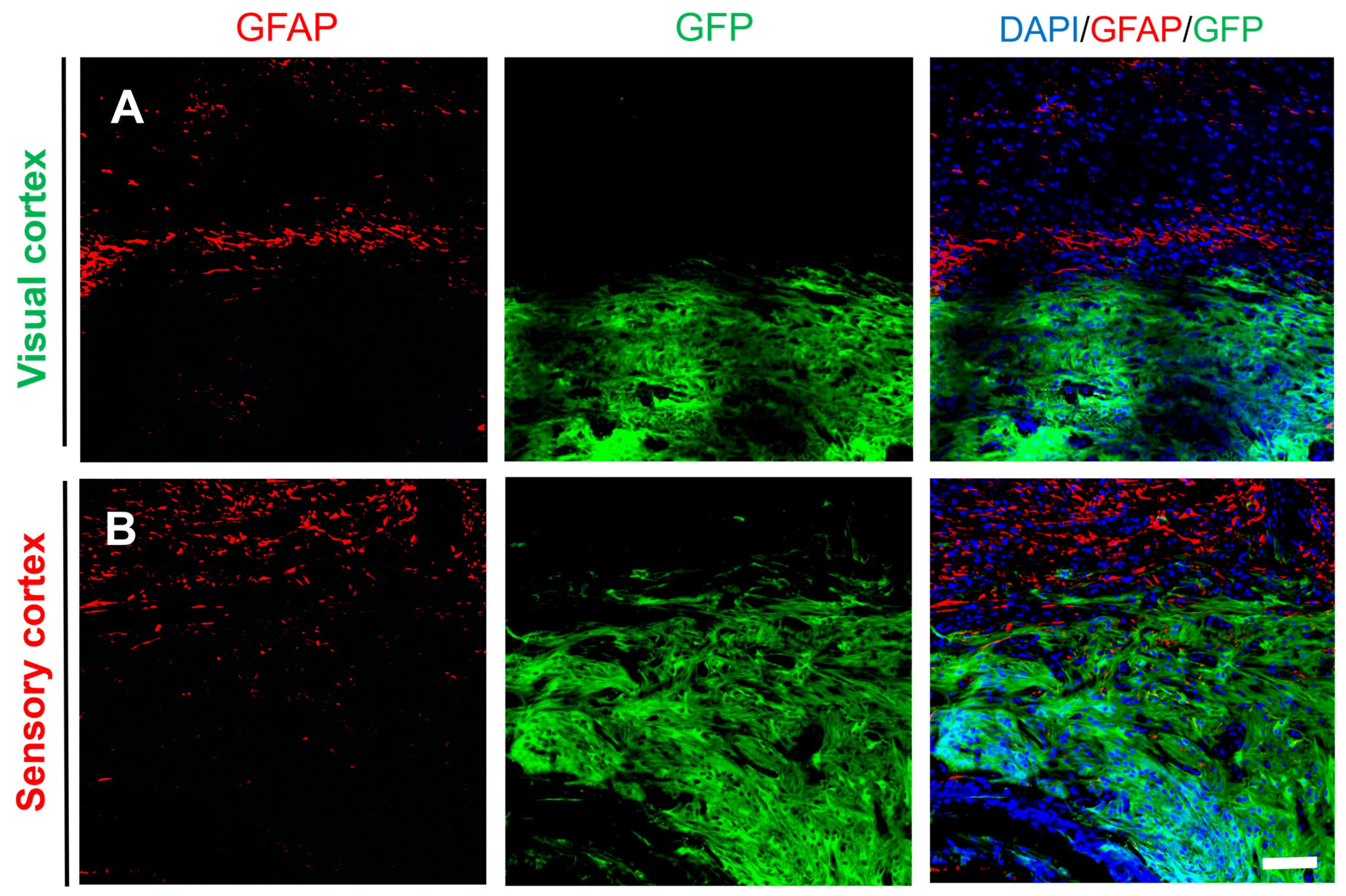
2.4. Differentiation of Grafted NSCs into Astrocytes
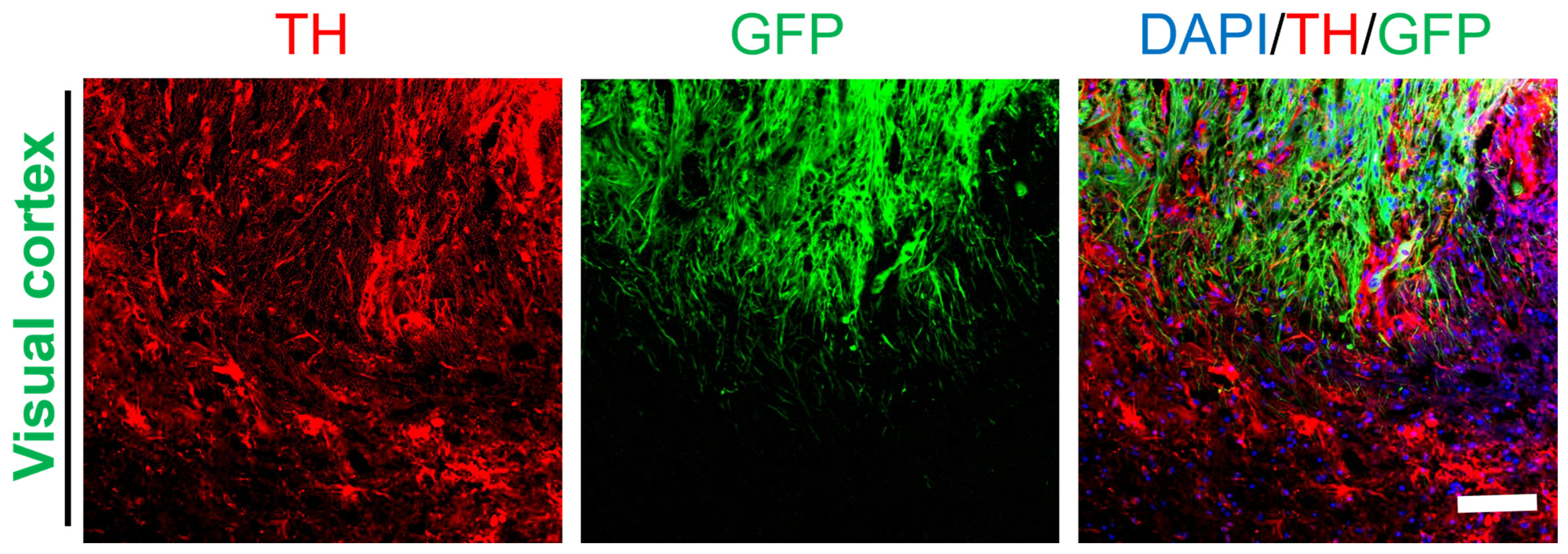
2.5. Differentiation of Grafted NSCs into Dopaminergic Neurons
3. Discussion
4. Materials and Methods
4.1. Embryonic Stem Cell Culture
4.2. Induction of NSC Differentiation
4.3. Induction of Dopaminergic Neurons Differentiation
4.4. Immunofluorescence Analysis of NSCs
4.5. Western Blot Analysis of NSCs
4.6. Brain Damage Surgery
4.7. Transplantation of NSCs
4.8. Toluidine Blue Staining
4.9. Identification of Cells by Immunofluorescence
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maas, A.I.R.; Menon, D.K.; Manley, G.T.; Abrams, M.; Åkerlund, C.; Andelic, N.; Aries, M.; Bashford, T.; Bell, M.J.; Bodien, Y.G.; et al. Traumatic brain injury: Progress and challenges in prevention, clinical care, and research. Lancet Neurol. 2022, 21, 1004–1060. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Z.; Wang, H.; Pei, Z.; Zhao, S. Molecular biomarkers of diffuse axonal injury: Recent advances and future perspectives. Expert. Rev. Mol. Diagn. 2024, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Howlett, J.R.; Nelson, L.D.; Stein, M.B. Mental Health Consequences of Traumatic Brain Injury. Biol. Psychiatry 2022, 91, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Fann, J.R.; Quinn, D.K.; Hart, T. Treatment of Psychiatric Problems After Traumatic Brain Injury. Biol. Psychiatry 2022, 91, 508–521. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, J.D. Traumatic Brain Injury and Treatment of Behavioral Health Conditions. Psychiatr. Serv. 2021, 72, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- McDonald, S.; Genova, H. The effect of severe traumatic brain injury on social cognition, emotion regulation, and mood. Handb. Clin. Neurol. 2021, 183, 235–260. [Google Scholar] [CrossRef]
- Zhou, Y.; Shao, A.; Xu, W.; Wu, H.; Deng, Y. Advance of Stem Cell Treatment for Traumatic Brain Injury. Front. Cell Neurosci. 2019, 13, 301. [Google Scholar] [CrossRef]
- Bonilla, C.; Zurita, M. Cell-Based Therapies for Traumatic Brain Injury: Therapeutic Treatments and Clinical Trials. Biomedicines 2021, 9, 669. [Google Scholar] [CrossRef]
- Velloso, F.J.; Shankar, S.; Parpura, V.; Rakic, P.; Levison, S.W. Neural Stem Cells in Adult Mammals are not Astrocytes. ASN Neuro 2022, 14, 17590914221134739. [Google Scholar] [CrossRef]
- De Gioia, R.; Biella, F.; Citterio, G.; Rizzo, F.; Abati, E.; Nizzardo, M.; Bresolin, N.; Comi, G.P.; Corti, S. Neural Stem Cell Transplantation for Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 3103. [Google Scholar] [CrossRef]
- Chrostek, M.R.; Fellows, E.G.; Guo, W.L.; Swanson, W.J.; Crane, A.T.; Cheeran, M.C.; Low, W.C.; Grande, A.W. Efficacy of Cell-Based Therapies for Traumatic Brain Injuries. Brain Sci. 2019, 9, 270. [Google Scholar] [CrossRef]
- Haus, D.L.; López-Velázquez, L.; Gold, E.M.; Cunningham, K.M.; Perez, H.; Anderson, A.J.; Cummings, B.J. Transplantation of human neural stem cells restores cognition in an immunodeficient rodent model of traumatic brain injury. Exp. Neurol. 2016, 281, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Badner, A.; Reinhardt, E.K.; Nguyen, T.V.; Midani, N.; Marshall, A.T.; Lepe, C.A.; Echeverria, K.; Lepe, J.J.; Torrecampo, V.; Bertan, S.H.; et al. Freshly Thawed Cryobanked Human Neural Stem Cells Engraft within Endogenous Neurogenic Niches and Restore Cognitive Function after Chronic Traumatic Brain Injury. J. Neurotrauma 2021, 38, 2731–2746. [Google Scholar] [CrossRef]
- Amirbekyan, M.; Adhikarla, V.; Cheng, J.P.; Moschonas, E.H.; Bondi, C.O.; Rockne, R.C.; Kline, A.E.; Gutova, M. Neuroprotective potential of intranasally delivered L-myc immortalized human neural stem cells in female rats after a controlled cortical impact injury. Sci. Rep. 2023, 13, 17874. [Google Scholar] [CrossRef]
- Xiong, L.L.; Hu, Y.; Zhang, P.; Zhang, Z.; Li, L.H.; Gao, G.D.; Zhou, X.F.; Wang, T.H. Neural Stem Cell Transplantation Promotes Functional Recovery from Traumatic Brain Injury via Brain Derived Neurotrophic Factor-Mediated Neuroplasticity. Mol. Neurobiol. 2018, 55, 2696–2711. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.K.; Wang, W.C.; Zhai, R.W.; Zhang, Y.H.; Yang, S.C.; Rizak, J.; Li, L.; Xu, L.Q.; Liu, L.; Pan, M.K.; et al. Neurons Differentiated from Transplanted Stem Cells Respond Functionally to Acoustic Stimuli in the Awake Monkey Brain. Cell Rep. 2016, 16, 1016–1025. [Google Scholar] [CrossRef]
- Li, H.; Gan, X.; Pan, L.; Zhang, Y.; Hu, X.; Wang, Z. EGF/bFGF promotes survival, migration and differentiation into neurons of GFP-labeled rhesus monkey neural stem cells xenografted into the rat brain. Biochem. Biophys. Res. Commun. 2022, 620, 76–82. [Google Scholar] [CrossRef]
- Hu, Z.; Gajavelli, S.; Spurlock, M.S.; Mahavadi, A.; Quesada, L.S.; Gajavelli, G.R.; Andreoni, C.B.; Di, L.; Janecki, J.; Lee, S.W.; et al. Human neural stem cell transplant location-dependent neuroprotection and motor deficit amelioration in rats with penetrating traumatic brain injury. J. Trauma. Acute Care Surg. 2020, 88, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Gutman, A.; Wasylyk, B. Nuclear targets for transcription regulation by oncogenes. Trends Genet. 1991, 7, 49–54. [Google Scholar] [CrossRef]
- Ferreira, A.; Rapoport, M. The synapsins: Beyond the regulation of neurotransmitter release. Cell Mol. Life Sci. 2002, 59, 589–595. [Google Scholar] [CrossRef]
- Hunt, C.A.; Schenker, L.J.; Kennedy, M.B. PSD-95 is associated with the postsynaptic density and not with the presynaptic membrane at forebrain synapses. J. Neurosci. 1996, 16, 1380–1388. [Google Scholar] [CrossRef]
- Skardelly, M.; Gaber, K.; Burdack, S.; Scheidt, F.; Hilbig, H.; Boltze, J.; Förschler, A.; Schwarz, S.; Schwarz, J.; Meixensberger, J.; et al. Long-term benefit of human fetal neuronal progenitor cell transplantation in a clinically adapted model after traumatic brain injury. J. Neurotrauma 2011, 28, 401–414. [Google Scholar] [CrossRef]
- Chen, T.; Wu, Y.; Wang, Y.; Zhu, J.; Chu, H.; Kong, L.; Yin, L.; Ma, H. Brain-Derived Neurotrophic Factor Increases Synaptic Protein Levels via the MAPK/Erk Signaling Pathway and Nrf2/Trx Axis Following the Transplantation of Neural Stem Cells in a Rat Model of Traumatic Brain Injury. Neurochem. Res. 2017, 42, 3073–3083. [Google Scholar] [CrossRef] [PubMed]
- Pang, A.L.; Xiong, L.L.; Xia, Q.J.; Liu, F.; Wang, Y.C.; Liu, F.; Zhang, P.; Meng, B.L.; Tan, S.; Wang, T.H. Neural Stem Cell Transplantation Is Associated with Inhibition of Apoptosis, Bcl-xL Upregulation, and Recovery of Neurological Function in a Rat Model of Traumatic Brain Injury. Cell Transplant. 2017, 26, 1262–1275. [Google Scholar] [CrossRef] [PubMed]
- Daadi, M.M.; Barberi, T.; Shi, Q.; Lanford, R.E. Nonhuman primate models in translational regenerative medicine. Stem Cells Dev. 2014, 23 (Suppl. S1), 83–87. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.J.; Zhang, P.; Hu, Y.Q.; Zhang, H.T.; Hong, S.Q.; Zhou, H.L.; Zhang, M.Y.; Xu, R.X. Neural stem-like cells derived from human amnion tissue are effective in treating traumatic brain injury in rat. Neurochem. Res. 2013, 38, 1022–1033. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.Q.; He, X.F.; Liang, F.Y.; Guo, Y.; Sunnassee, G.; Chen, J.; Cao, X.M.; Chen, Y.Y.; Pan, G.J.; Pei, Z.; et al. Transplanted human neural precursor cells integrate into the host neural circuit and ameliorate neurological deficits in a mouse model of traumatic brain injury. Neurosci. Lett. 2018, 674, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Jgamadze, D.; Lim, J.T.; Zhang, Z.; Harary, P.M.; Germi, J.; Mensah-Brown, K.; Adam, C.D.; Mirzakhalili, E.; Singh, S.; Gu, J.B.; et al. Structural and functional integration of human forebrain organoids with the injured adult rat visual system. Cell Stem Cell 2023, 30, 137–152. [Google Scholar] [CrossRef]
- Gao, J.; Grill, R.J.; Dunn, T.J.; Bedi, S.; Labastida, J.A.; Hetz, R.A.; Xue, H.; Thonhoff, J.R.; DeWitt, D.S.; Prough, D.S.; et al. Human Neural Stem Cell Transplantation-Mediated Alteration of Microglial/Macrophage Phenotypes after Traumatic Brain Injury. Cell Transplant. 2016, 25, 1863–1877. [Google Scholar] [CrossRef]
- Lee, J.Y.; Acosta, S.; Tuazon, J.P.; Xu, K.; Nguyen, H.; Lippert, T.; Liska, M.G.; Semechkin, A.; Garitaonandia, I.; Gonzalez, R.; et al. Human parthenogenetic neural stem cell grafts promote multiple regenerative processes in a traumatic brain injury model. Theranostics 2019, 9, 1029–1046. [Google Scholar] [CrossRef]
- Imai, R.; Tamura, R.; Yo, M.; Sato, M.; Fukumura, M.; Takahara, K.; Kase, Y.; Okano, H.; Toda, M. Neuroprotective Effects of Genome-Edited Human iPS Cell-Derived Neural Stem/Progenitor Cells on Traumatic Brain Injury. Stem Cells 2023, 41, 603–616. [Google Scholar] [CrossRef]
- Alizada, M.; Lin, S.; Gao, H. Recent advances in the treatment of traumatic brain injury with autologous and non-autologous multipotent stem and progenitor cells: Preclinical models and clinical trials. Folia Neuropathol. 2021, 59, 298–316. [Google Scholar] [CrossRef] [PubMed]
- Wianny, F.; Bernat, A.; Huissoud, C.; Marcy, G.; Markossian, S.; Cortay, V.; Giroud, P.; Leviel, V.; Kennedy, H.; Savatier, P.; et al. Derivation and cloning of a novel rhesus embryonic stem cell line stably expressing tau-green fluorescent protein. Stem Cells 2008, 26, 1444–1453. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Tang, X.; Bai, Y.F.; Wang, S.; An, J.; Wu, Y.; Xu, Z.D.; Zhang, Y.A.; Chen, Z. Dopaminergic precursors differentiated from human blood-derived induced neural stem cells improve symptoms of a mouse Parkinson’s disease model. Theranostics 2018, 8, 4679–4694. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Shi, L.; Huang, T.; Luo, Y.; Chen, Y.; Li, S.; Wang, Z. Neural Stem Cells Transplanted into Rhesus Monkey Cortical Traumatic Brain Injury Can Survive and Differentiate into Neurons. Int. J. Mol. Sci. 2024, 25, 1642. https://doi.org/10.3390/ijms25031642
Liu S, Shi L, Huang T, Luo Y, Chen Y, Li S, Wang Z. Neural Stem Cells Transplanted into Rhesus Monkey Cortical Traumatic Brain Injury Can Survive and Differentiate into Neurons. International Journal of Molecular Sciences. 2024; 25(3):1642. https://doi.org/10.3390/ijms25031642
Chicago/Turabian StyleLiu, Shuyi, Liping Shi, Tianzhuang Huang, Yuyi Luo, Yongchang Chen, Shangang Li, and Zhengbo Wang. 2024. "Neural Stem Cells Transplanted into Rhesus Monkey Cortical Traumatic Brain Injury Can Survive and Differentiate into Neurons" International Journal of Molecular Sciences 25, no. 3: 1642. https://doi.org/10.3390/ijms25031642