
Cellular Distribution of Aquaporin 3, 7 and 9 in the Male Reproductive System: A Lesson from Bovine Study (Bos taurus)

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
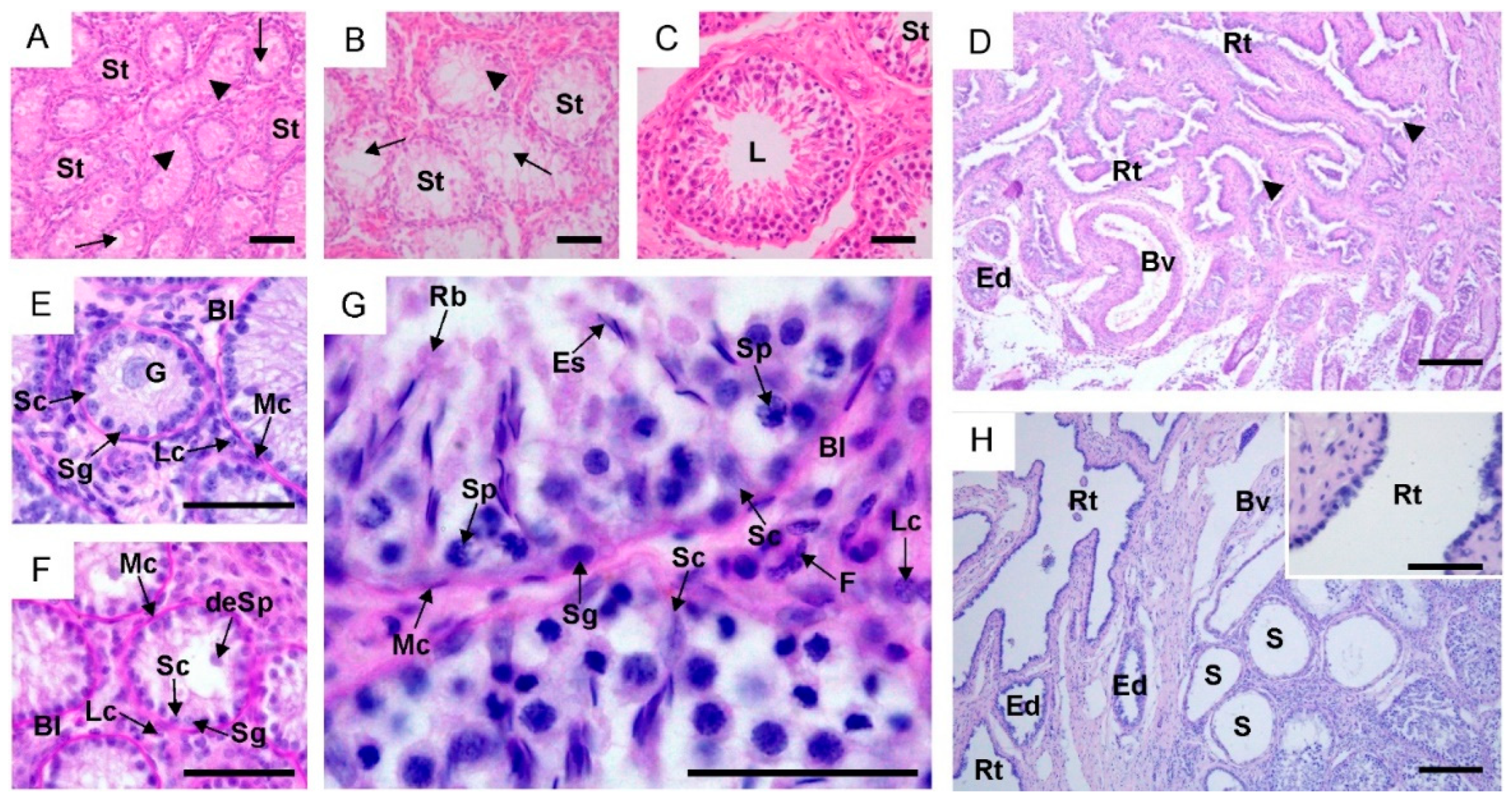
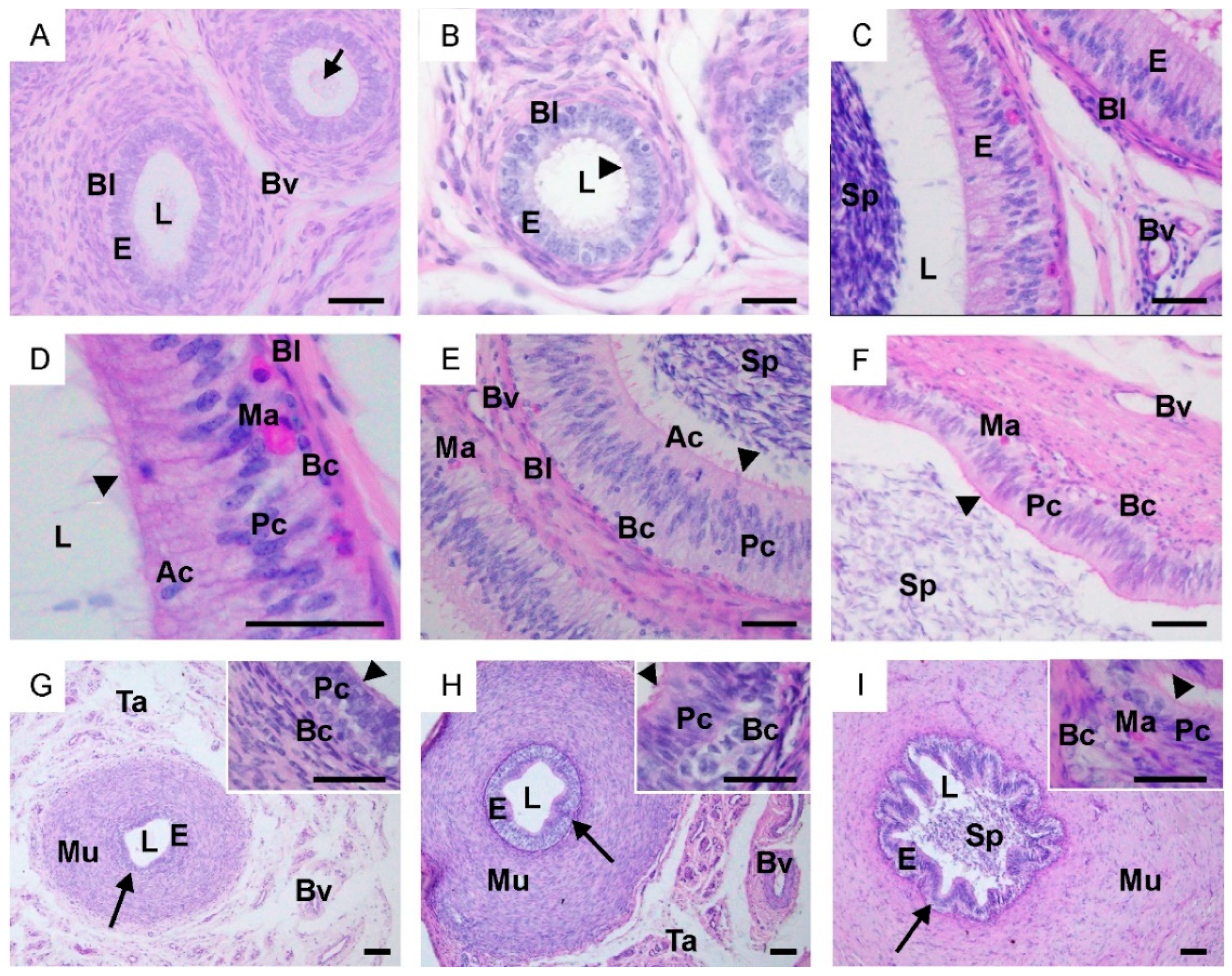
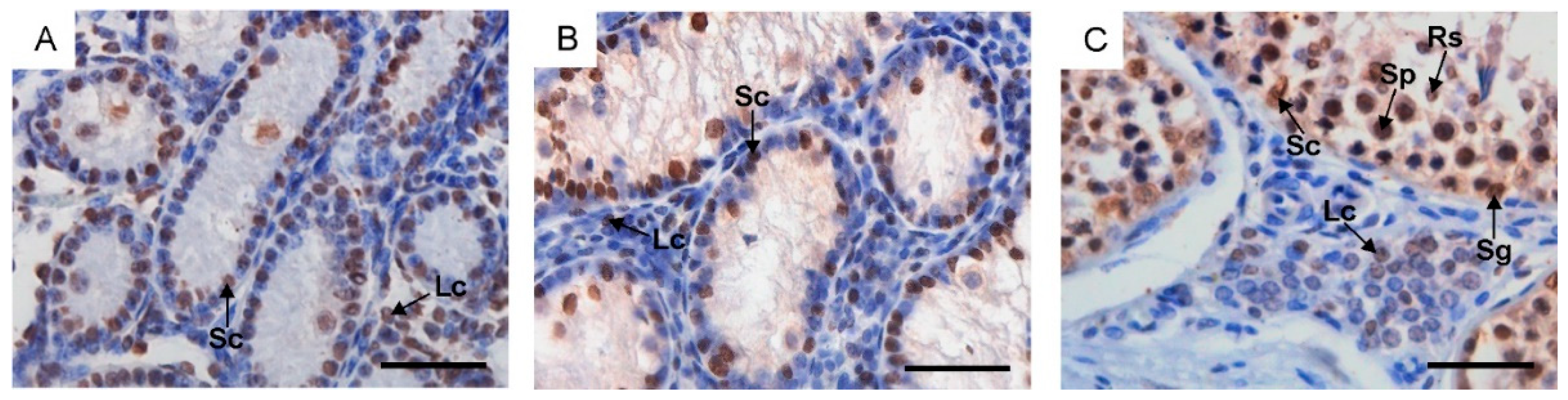
2.1. Morphological Changes in the Male Reproductive Tract, Collagen Distribution and Identification of Sertoli and Leydig Cells
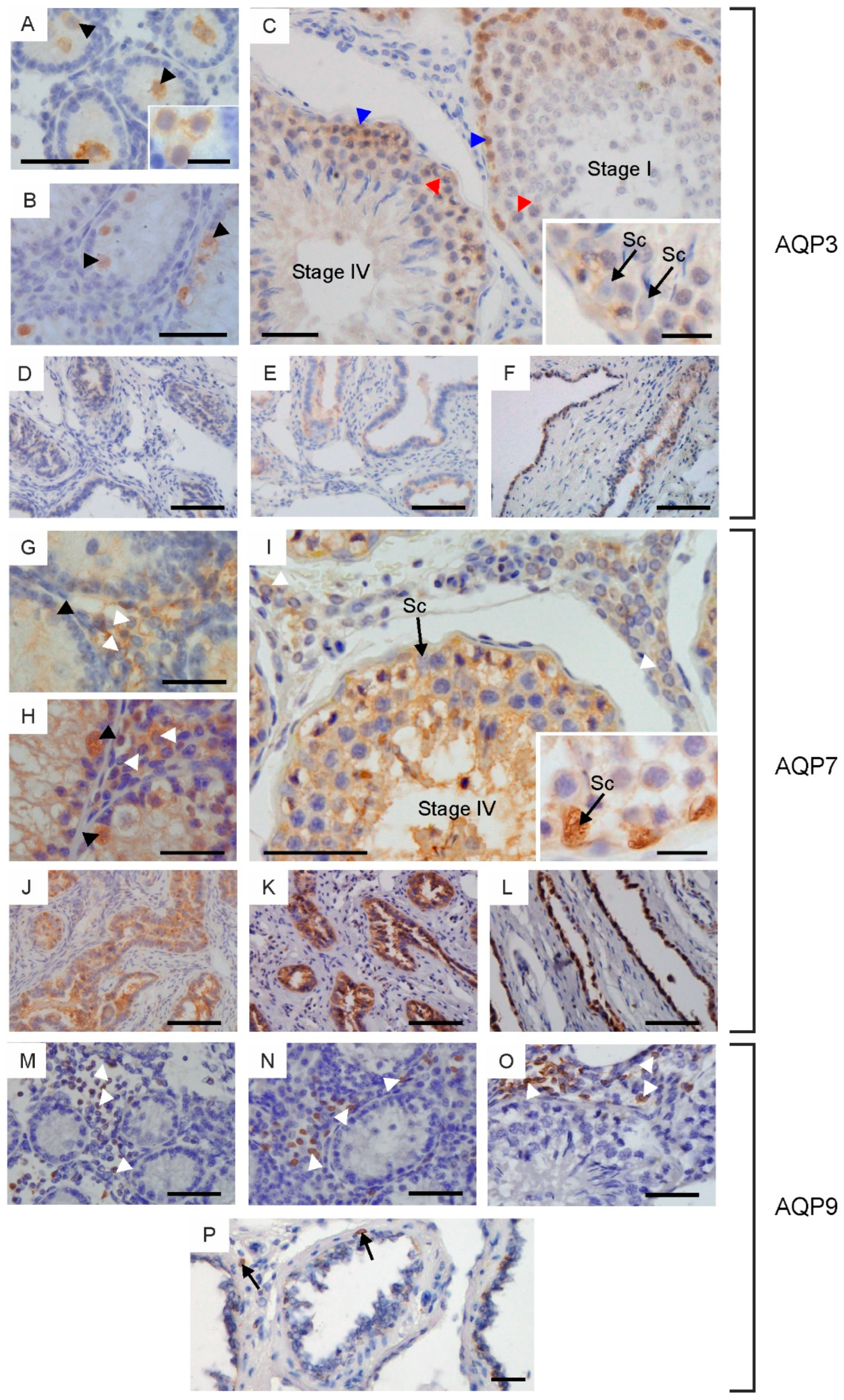
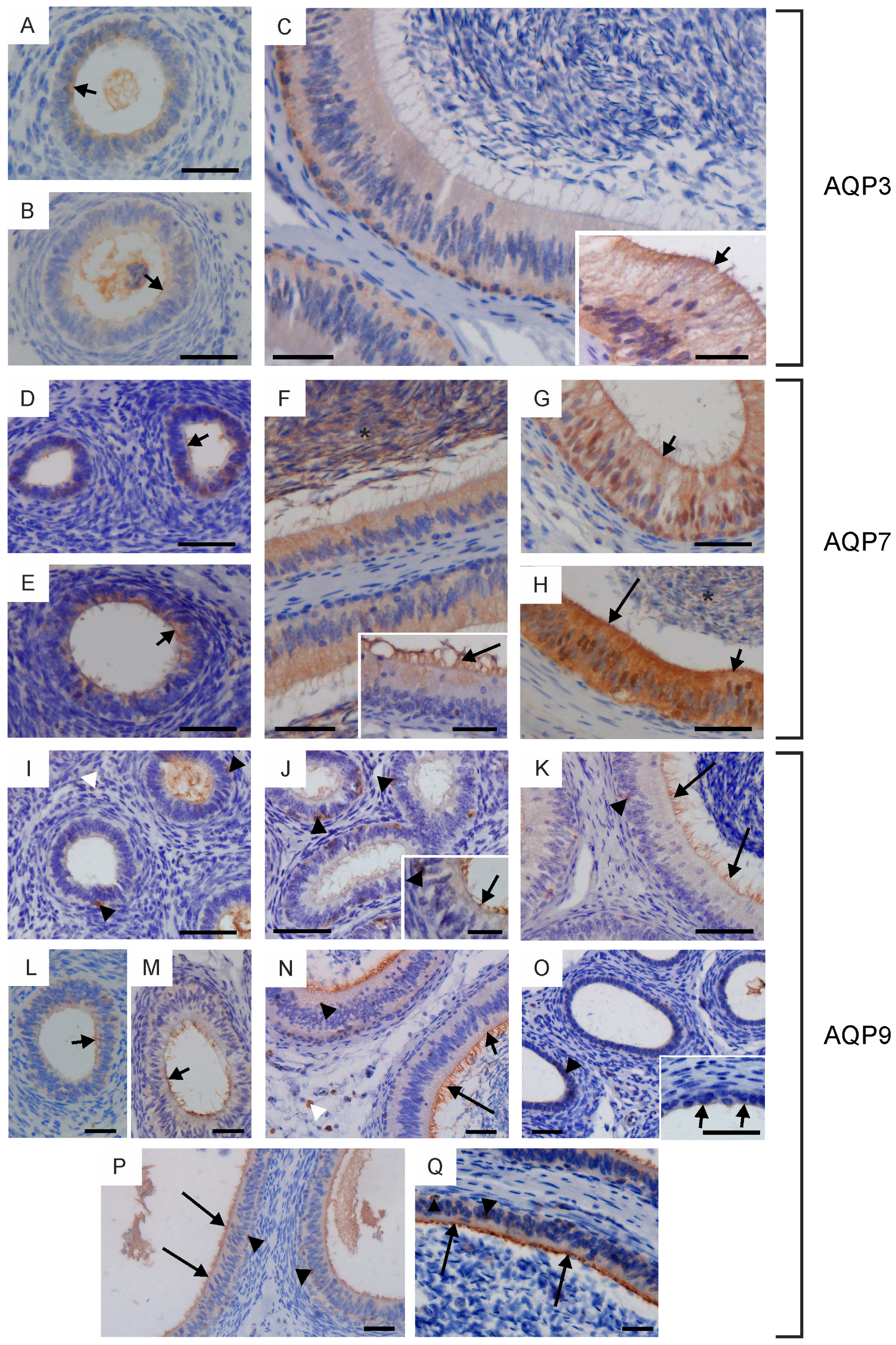
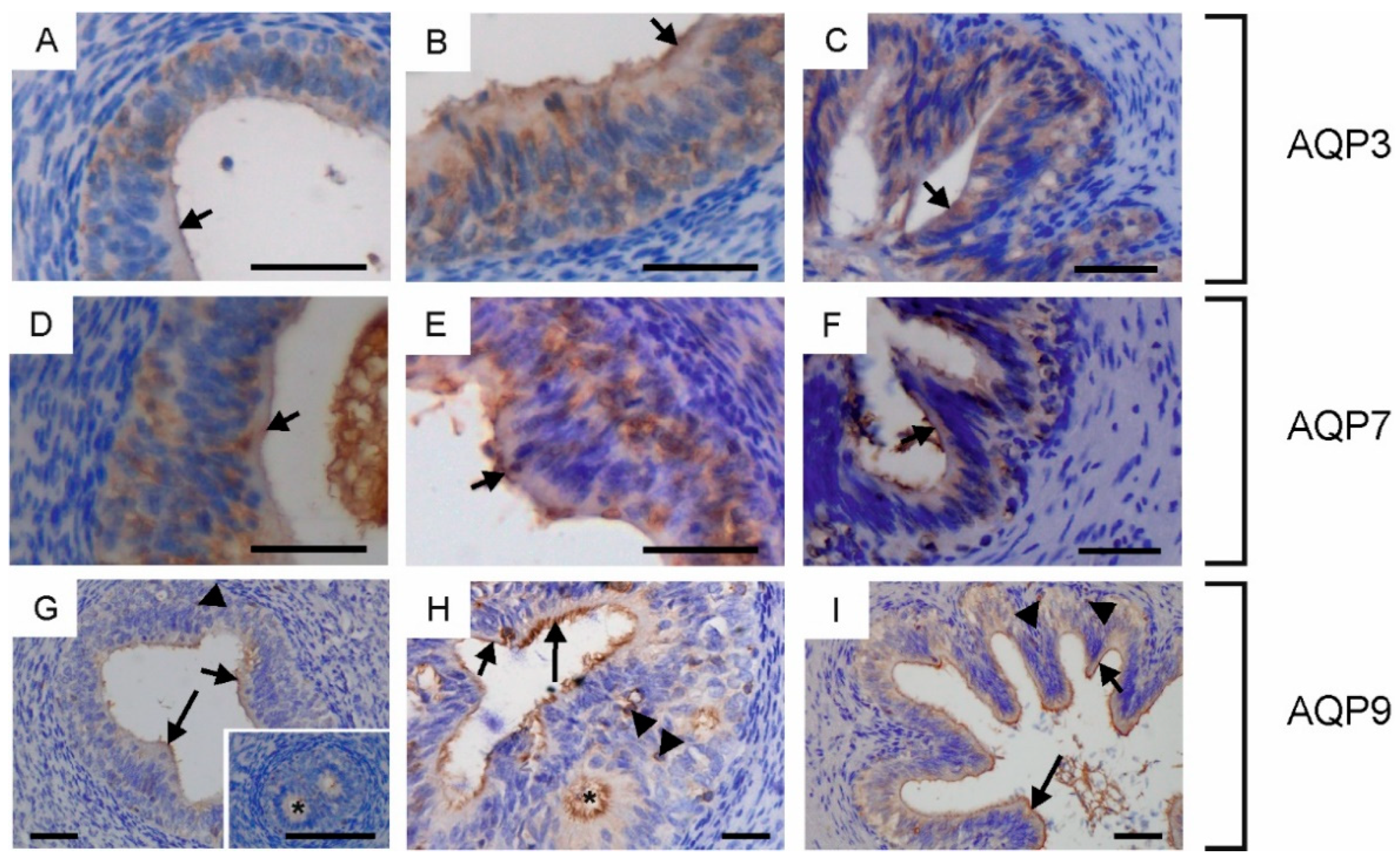
2.2. Immunolocalization of Aquaglyceroporins in Bovine Male Reproductive Tract
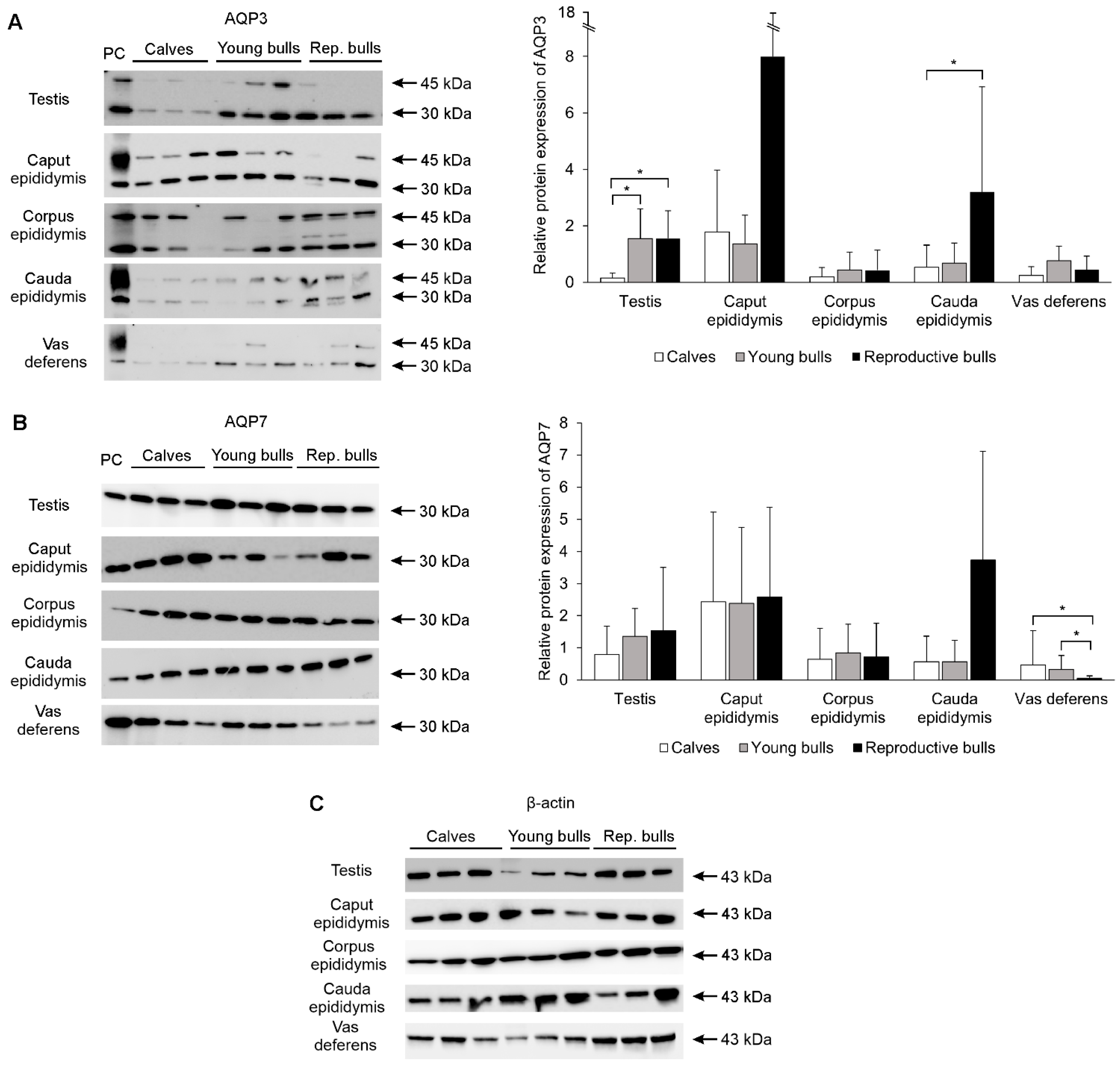
2.3. Immunoblotting and Densitometric Analysis of Aquaglyceroporins in the Bovine Male Reproductive System
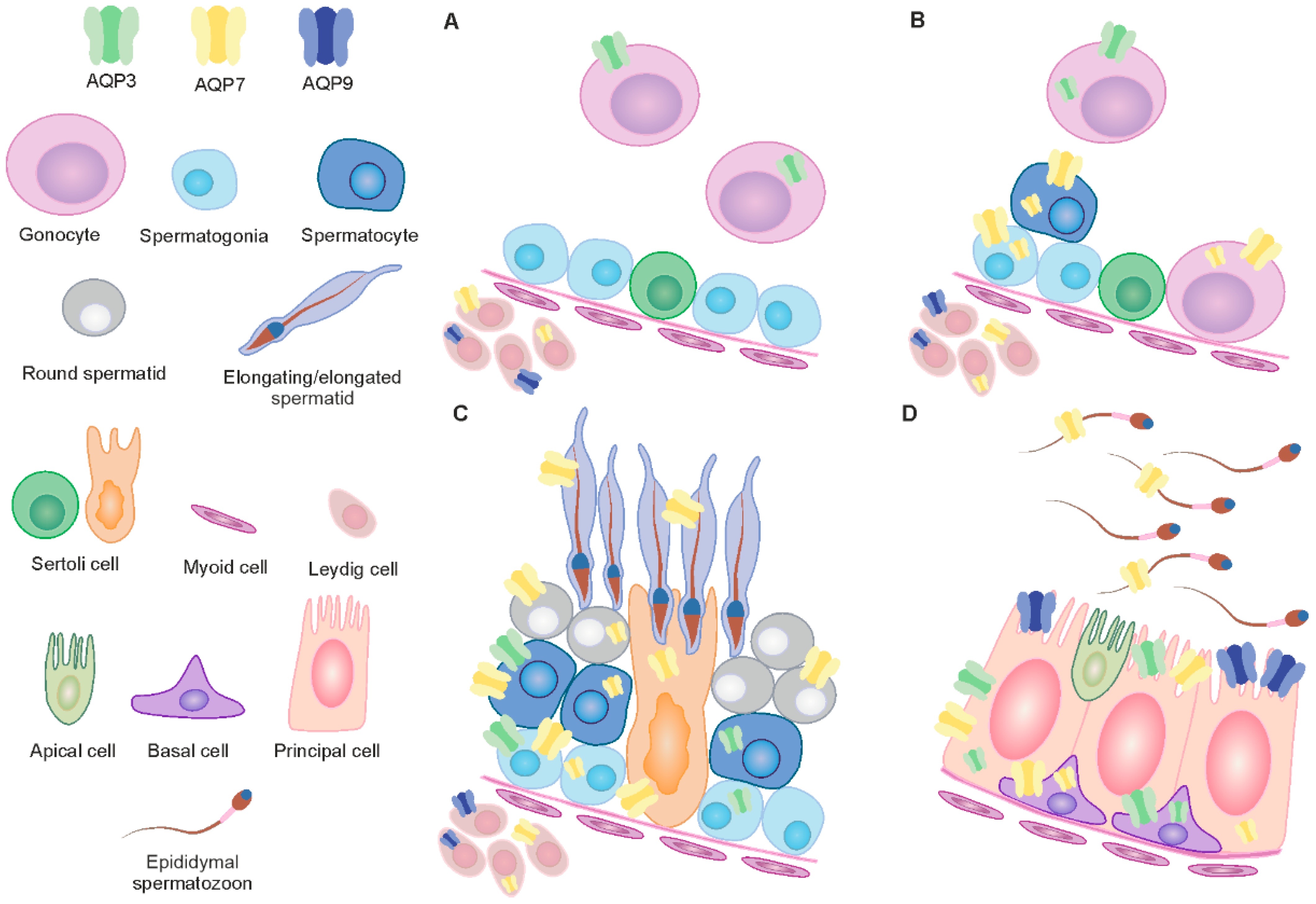
3. Discussion
4. Materials and Methods
4.1. Animals and Tissue Collection
4.2. Histological and Morphometric Analysis
4.3. Antibodies
4.4. Immunohistochemistry (IHC)
4.5. Western Blot (WB)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calamita, G.; Mazzone, A.; Bizzoca, A.; Svelto, M. Possible Involvement of Aquaporin-7 and -8 in Rat Testis Development and Spermatogenesis. Biochem. Biophys. Res. Commun. 2001, 288, 619–625. [Google Scholar] [CrossRef]
- Michałek, K.; Oberska, P. Aquaporins in the Male Reproductive System: A Chance for Paternity or a Road to Nowhere? Andrology 2023, 11, 970–972. [Google Scholar] [CrossRef]
- Ishibashi, K.; Hara, S.; Kondo, S. Aquaporin Water Channels in Mammals. Clin. Exp. Nephrol. 2009, 13, 107–117. [Google Scholar] [CrossRef]
- Gomes, D.; Agasse, A.; Thiébaud, P.; Delrot, S.; Gerós, H.; Chaumont, F. Aquaporins are Multifunctional Water and Solute Transporters Highly Divergent in Living Organisms. Biochim. Biophys. Acta 2009, 1788, 1213–1228. [Google Scholar] [CrossRef]
- Ishibashi, K.; Kuwahara, M.; Gu, Y.; Kageyama, Y.; Tohsaka, A.; Suzuki, F.; Marumo, F.; Sasaki, S. Cloning and Functional Expression of a New Water Channel Abundantly Expressed in the Testis Permeable to Water, Glycerol, and Urea. J. Biol. Chem. 1997, 272, 20782–20786. [Google Scholar] [CrossRef]
- Bernardino, R.L.; Carrageta, D.F.; Silva, A.M.; Calamita, G.; Alves, M.G.; Soveral, G.; Oliveira, P.F. Estrogen Modulates Glycerol Permeability in Sertoli Cells through Downregulation of Aquaporin-9. Cells 2018, 7, 153. [Google Scholar] [CrossRef]
- Hermo, L.; Krzeczunowicz, D.; Ruz, R. Cell Specificity of Aquaporins 0, 3, and 10 Expressed in the Testis, Efferent Ducts, and Epididymis of Adult Rats. J. Androl. 2004, 25, 494–505. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, S.; Bosi, G.; Accogli, G.; Desantis, S. Seasonal and Ageing-Depending Changes of Aquaporins 1 and 9 Expression in the Genital Tract of Buffalo Bulls (Bubalus bubalis). Reprod. Domest. Anim. 2016, 51, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, Y.; Morishita, Y.; Ishibashi, K. Aquaporin10 is a Pseudogene in Cattle and Their Relatives. Biochem. Biophys. Rep. 2015, 1, 16–21. [Google Scholar] [CrossRef]
- Amann, R.P. Reproductive Capacity of Dairy Bulls. III. The Effect of Ejaculation Frequency, Unilateral Vasectomy, and Age on Spermatogenesis. Am. J. Anat. 1962, 110, 49–67. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Choi, Y.R.; Kim, B.J.; Jung, S.E.; Kim, S.M.; Jin, J.H.; Yun, M.H.; Kim, S.U.; Kim, Y.H.; Hwang, S.; et al. GDNF Family Receptor Alpha 1 is a Reliable Marker of Undifferentiated Germ Cells in Bulls. Theriogenology 2019, 132, 172–181. [Google Scholar] [CrossRef]
- Ketola, I.; Pentikäinen, V.; Vaskivuo, T.; Ilvesmäki, V.; Herva, R.; Dunkel, L.; Tapanainen, J.S.; Toppari, J.; Heikinheimo, M. Expression of Transcription Factor GATA-4 During Human Testicular Development and Disease. J. Clin. Endocrinol. Metab. 2000, 85, 3925–3931. [Google Scholar] [CrossRef]
- Fossland, R.G.; Schultze, A.B. A Histological Study of the Postnatal Development of the Bovine Testis; Research Bulletin: Bulletin of the Agricultural Experiment Station of Nebraska No. 199; University of Nebraska: Lincoln, NE, USA, 1961; Volume 68. [Google Scholar]
- Goyal, H.O. Morphology of the Bovine Epididymis. Am. J. Anat. 1985, 172, 155–172. [Google Scholar] [CrossRef] [PubMed]
- Cornwall, G.A. New Insights into Epididymal Biology and Function. Hum. Reprod. Update 2009, 15, 213–227. [Google Scholar] [CrossRef]
- Hermo, L.; Badran, H.; Andonian, S. The Structural Organization and Functions of the Epithelium of the Vas Deferens. In The Epididymis: From Molecules to Clinical Practice; Robaire, B., Hinton, B.T., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 233–250. [Google Scholar]
- van Der Horst, G.; Seier, J.V.; Spinks, A.C.; Hendricks, S. The Maturation of Sperm Motility in the Epididymis and Vas Deferens of the Vervet Monkey, Cercopithecus aethiops. Int. J. Androl. 1999, 22, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Li, Y.; Yang, Q.; Xu, S.; Ma, S.; Yan, R.; Zhang, R.; Jia, G.; Ai, D.; Yang, Q. Gene Expression Dynamics During the Gonocyte to Spermatogonia Transition and Spermatogenesis in the Domestic Yak. J. Anim. Sci. Biotechnol. 2019, 10, 64. [Google Scholar] [CrossRef] [PubMed]
- Hara-Chikuma, M.; Verkman, A.S. Aquaporin-3 Facilitates Epidermal Cell Migration and Proliferation During Wound Healing. J. Mol. Med. 2008, 86, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Galán-Cobo, A.; Ramírez-Lorca, R.; Echevarría, M. Role of Aquaporins in Cell Proliferation: What Else Beyond Water Permeability? Channels 2016, 10, 185–201. [Google Scholar] [CrossRef]
- Zannetti, A.; Benga, G.; Brunetti, A.; Napolitano, F.; Avallone, L.; Pelagalli, A. Role of Aquaporins in the Physiological Functions of Mesenchymal Stem Cells. Cells 2020, 9, 2678. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Saadoun, S.; Verkman, A.S. Aquaporins and Cell Migration. Pflug. Arch. 2008, 456, 693–700. [Google Scholar] [CrossRef]
- O’Flaherty, C.; Boisvert, A.; Manku, G.; Culty, M. Protective Role of Peroxiredoxins Against Reactive Oxygen Species in Neonatal Rat Testicular Gonocytes. Antioxidants 2019, 9, 32. [Google Scholar] [CrossRef]
- Tuck, R.R.; Setchell, B.P.; Waites, G.M.; Young, J.A. The Composition of Fluid Collected by Micropuncture and Catheterization from the Seminiferous Tubules and Rete Testis of Rats. Pflug. Arch. 1970, 318, 225–243. [Google Scholar] [CrossRef]
- Nozaki, K.; Ishii, D.; Ishibashi, K. Intracellular Aquaporins: Clues for Intracellular Water Transport? Pflug. Arch. 2008, 456, 701–707. [Google Scholar] [CrossRef]
- Lie, S.; Wang, T.; Forbes, B.; Proud, C.G.; Petersen, J. The Ability to Utilize Ammonia as Nitrogen Source is Cell Type Specific and Intricately Linked to GDH, AMPK and mTORC1. Sci. Rep. 2019, 9, 1461. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Im, J.W.; Kim, D.H.; Bae, H.R. Differential Expressions of Aquaporin Subtypes in the Adult Mouse Testis. Dev. Reprod. 2022, 26, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Kageyama, Y.; Ishibashi, K.; Hayashi, T.; Xia, G.; Sasaki, S.; Kihara, K. Expression of Aquaporins 7 and 8 in the Developing Rat Testis. Andrologia 2001, 33, 165–169. [Google Scholar] [CrossRef]
- Saito, K.; Kageyama, Y.; Okada, Y.; Kawakami, S.; Kihara, K.; Ishibashi, K.; Sasaki, S. Localization of Aquaporin-7 in Human Testis and Ejaculated Sperm: Possible Involvement in Maintenance of Sperm Quality. J. Urol. 2004, 172 Pt 1, 2073–2076. [Google Scholar] [CrossRef]
- Yeung, C.H.; Callies, C.; Tüttelmann, F.; Kliesch, S.; Cooper, T.G. Aquaporins in the Human Testis and Spermatozoa—Identification, Involvement in Sperm Volume Regulation and Clinical Relevance. Int. J. Androl. 2010, 33, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Pelagalli, A.; Squillacioti, C.; Ali’, S.; Liguori, G.; Mirabella, N. Cellular Distribution of Aquaporins in Testes of Normal and Cryptorchid Dogs: A Preliminary Study on Dynamic Roles. Anim. Reprod. Sci. 2019, 204, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.D. Spermatid-Sertoli Tubulobulbar Complexes as Devices for Elimination of Cytoplasm from the Head Region Late Spermatids of the Rat. Anat. Rec. 1979, 194, 233–246. [Google Scholar] [CrossRef]
- Sprando, R.L.; Russell, L.D. Comparative Study of Cytoplasmic Elimination in Spermatids of Selected Mammalian Species. Am. J. Anat. 1987, 178, 72–80. [Google Scholar] [CrossRef]
- Calamita, G.; Mazzone, A.; Cho, Y.S.; Valenti, G.; Svelto, M. Expression and Localization of the Aquaporin-8 Water Channel in Rat Testis. Biol. Reprod. 2001, 64, 1660–1666. [Google Scholar] [CrossRef]
- França, L.R.; Hess, R.A.; Dufour, J.M.; Hofmann, M.C.; Griswold, M.D. The Sertoli Cell: One Hundred Fifty Years of Beauty and Plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef]
- Bernardino, R.L.; Marinelli, R.A.; Maggio, A.; Gena, P.; Cataldo, I.; Alves, M.G.; Svelto, M.; Oliveira, P.F.; Calamita, G. Hepatocyte and Sertoli Cell Aquaporins, Recent Advances and Research Trends. Int. J. Mol. Sci. 2016, 17, 1096. [Google Scholar] [CrossRef] [PubMed]
- Nihei, K.; Koyama, Y.; Tani, T.; Yaoita, E.; Ohshiro, K.; Adhikary, L.P.; Kurosaki, I.; Shirai, Y.; Hatakeyama, K.; Yamamoto, T. Immunolocalization of Aquaporin-9 in Rat Hepatocytes and Leydig Cells. Arch. Histol. Cytol. 2001, 64, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Nicòtina, P.A.; Romeo, C.; Arena, S.; Arena, F.; Maisano, D.; Zuccarello, B. Immunoexpression of Aquaporin-1 in Adolescent Varicocele Testes: Possible Significance for Fluid Reabsorption. Urology 2005, 65, 149–152. [Google Scholar] [CrossRef]
- Kannan, A.; Mariajoseph-Antony, L.F.; Panneerselvam, A.; Loganathan, C.; Kiduva Jothiraman, D.; Anbarasu, K.; Prahalathan, C. Aquaporin 9 Regulates Leydig Cell Steroidogenesis in Diabetes. Syst. Biol. Reprod. Med. 2022, 68, 213–226. [Google Scholar] [CrossRef]
- Hess, R.A.; Hermo, L. Rete Testis: Structure, Cell Biology and Site for Stem Cell Transplantation. In Encyclopedia of Reproduction; Skinner, M., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 263–269. [Google Scholar]
- Kulibin, A.Y.; Malolina, E.A. The Rete Testis: Development and Role in Testis Function. Russ. J. Dev. Biol. 2021, 52, 370–378. [Google Scholar] [CrossRef]
- Boj, M.; Chauvigné, F.; Cerdà, J. Aquaporin Biology of Spermatogenesis and Sperm Physiology in Mammals and Teleosts. Biol. Bull. 2015, 229, 93–108. [Google Scholar] [CrossRef]
- Domeniconi, R.F.; Orsi, A.M.; Justulin, L.A., Jr.; Leme Beu, C.C.; Felisbino, S.L. Immunolocalization of Aquaporins 1, 2 and 7 in Rete Testis, Efferent Ducts, Epididymis and Vas Deferens of Adult Dog. Cell Tissue Res. 2008, 332, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.Y.; Li, Y.; Bi, Z.W.; Yu, H.M.; Li, X.J. Expression and Immunohistochemical Localization of Aquaporin-1 in Male Reproductive Organs of the Mouse. Anat. Histol. Embryol. 2008, 37, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Schimming, B.C.; Pinheiro, P.; de Matteis, R.; Machado, C.M.; Domeniconi, R.F. Immunolocalization of Aquaporins 1 and 9 in the Ram Efferent Ducts and Epididymis. Reprod. Domest. Anim. 2015, 50, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Badran, H.H.; Hermo, L.S. Expression and Regulation of Aquaporins 1, 8, and 9 in the Testis, Efferent Ducts, and Epididymis of Adult Rats and during Postnatal Development. J. Androl. 2002, 23, 358–373. [Google Scholar] [CrossRef]
- Ribeiro, J.C.; Alves, M.G.; Yeste, M.; Cho, Y.S.; Calamita, G.; Oliveira, P.F. Aquaporins and (In)fertility: More Than Just Water Transport. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166039. [Google Scholar] [CrossRef]
- Pastor-Soler, N.; Bagnis, C.; Sabolic, I.; Tyszkowski, R.; McKee, M.; Van Hoek, A.; Breton, S.; Brown, D. Aquaporin 9 Expression along the Male Reproductive Tract. Biol. Reprod. 2001, 65, 384–393. [Google Scholar] [CrossRef]
- Domeniconi, R.F.; Orsi, A.M.; Justulin, L.A., Jr.; Beu, C.C.; Felisbino, S.L. Aquaporin 9 (AQP9) Localization in the Adult Dog Testis Excurrent Ducts by Immunohistochemistry. Anat. Rec. 2007, 290, 1519–1525. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Troedsson, M.H.; Rutllant, J. Region-Specific Expression of Aquaporin Subtypes in Equine Testis, Epididymis, and Ductus Deferens. Anat. Rec. 2013, 296, 1115–1126. [Google Scholar] [CrossRef]
- Castro, M.M.; Kim, B.; Hill, E.; Fialho, M.C.; Puga, L.C.; Freitas, M.B.; Breton, S.; Machado-Neves, M. The Expression Patterns of Aquaporin 9, Vacuolar H+-ATPase, and Cytokeratin 5 in the Epididymis of the Common Vampire Bat. Histochem. Cell Biol. 2017, 147, 39–48. [Google Scholar] [CrossRef]
- Hermo, L.; Schellenberg, M.; Liu, L.Y.; Dayanandan, B.; Zhang, T.; Mandato, C.A.; Smith, C.E. Membrane Domain Specificity in the Spatial Distribution of Aquaporins 5, 7, 9, and 11 in Efferent Ducts and Epididymis of Rats. J. Histochem. Cytochem. 2008, 56, 1121–1135. [Google Scholar] [CrossRef]
- Mobasheri, A.; Wray, S.; Marples, D. Distribution of AQP2 and AQP3 Water Channels in Human Tissue Microarrays. J. Mol. Histol. 2005, 36, 1–14. [Google Scholar] [CrossRef]
- Da Silva, N.; Silberstein, C.; Beaulieu, V.; Piétrement, C.; Van Hoek, A.N.; Brown, D.; Breton, S. Postnatal Expression of Aquaporins in Epithelial Cells of the Rat Epididymis. Biol. Reprod. 2006, 74, 427–438. [Google Scholar] [CrossRef] [PubMed]
- Schimming, B.C.; Baumam, C.; Pinheiro, P.; de Matteis, R.; Domeniconi, R.F. Aquaporin 9 Is Expressed in the Epididymis of Immature and Mature Pigs. Reprod. Domest. Anim. 2017, 52, 617–624. [Google Scholar] [CrossRef]
- Arrighi, S. Are the Basal Cells of the Mammalian Epididymis Still an Enigma? Reprod. Fertil. Dev. 2014, 26, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.L.; Campolina-Silva, G.H.; Nogueira, J.C.; Mahecha, G.A.; Oliveira, C.A. Differential Expression and Seasonal Variation on Aquaporins 1 and 9 in the Male Genital System of Big Fruit-Eating Bat Artibeus lituratus. Gen. Comp. Endocrinol. 2013, 186, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Koun, S.; Kim, J.D.; Rhee, M.; Kim, M.J.; Huh, T.L. Spatiotemporal Expression Pattern of the Zebrafish Aquaporin 8 Family during Early Developmental Stages. Gene Expr. Patterns 2016, 21, 1–6. [Google Scholar] [CrossRef]
- Forecki, J.; Van Antwerp, D.J.; Lujan, S.M.; Merzdorf, C.S. Roles for Xenopus Aquaporin-3b (aqp3.L) during Gastrulation: Fibrillar Fibronectin and Tissue Boundary Establishment in the Dorsal Margin. Dev. Biol. 2018, 433, 3–16. [Google Scholar] [CrossRef] [PubMed]
- Casotti, G.; Waldron, T.; Misquith, G.; Powers, D.; Slusher, L. Expression and Localization of an Aquaporin-1 Homologue in the Avian Kidney and Lower Intestinal Tract. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 147, 355–362. [Google Scholar] [CrossRef]
- Gamil, H.; Assaf, M.; Khater, M.; Fawzy, M. Abnormal Nuclear Expression of Aquaporin-3 in Lesional and Perilesional Skin of Vitiligo Patients: A Novel Immunohistochemical Finding. J. Cosmet. Dermatol. 2023, 22, 1063–1070. [Google Scholar] [CrossRef]
- Michałek, K.; Grabowska, M. Investigating Cellular Location of Aquaporins in the Bovine Kidney. A New View on Renal Physiology in Cattle. Res. Vet. Sci. 2019, 125, 162–169. [Google Scholar] [CrossRef]
- Michałek, K.; Laszczyńska, M.; Ciechanowicz, A.K.; Herosimczyk, A.; Rotter, I.; Oganowska, M.; Lepczyński, A. Immunohistochemical Identification of Aquaporin 2 in the Kidneys of Young Beef Cattle. Biotech. Histochem. 2014, 89, 342–347. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oberska, P.; Grabowska, M.; Marynowska, M.; Murawski, M.; Gączarzewicz, D.; Syczewski, A.; Michałek, K. Cellular Distribution of Aquaporin 3, 7 and 9 in the Male Reproductive System: A Lesson from Bovine Study (Bos taurus). Int. J. Mol. Sci. 2024, 25, 1567. https://doi.org/10.3390/ijms25031567
Oberska P, Grabowska M, Marynowska M, Murawski M, Gączarzewicz D, Syczewski A, Michałek K. Cellular Distribution of Aquaporin 3, 7 and 9 in the Male Reproductive System: A Lesson from Bovine Study (Bos taurus). International Journal of Molecular Sciences. 2024; 25(3):1567. https://doi.org/10.3390/ijms25031567
Chicago/Turabian StyleOberska, Patrycja, Marta Grabowska, Marta Marynowska, Maciej Murawski, Dariusz Gączarzewicz, Andrzej Syczewski, and Katarzyna Michałek. 2024. "Cellular Distribution of Aquaporin 3, 7 and 9 in the Male Reproductive System: A Lesson from Bovine Study (Bos taurus)" International Journal of Molecular Sciences 25, no. 3: 1567. https://doi.org/10.3390/ijms25031567