
Identification and Expression Analysis of the Nucleotidyl Transferase Protein (NTP) Family in Soybean (Glycine max) under Various Abiotic Stresses

Abstract
:1. Introduction
2. Results
2.1. Identification and Characterization of GmNTP Family Genes
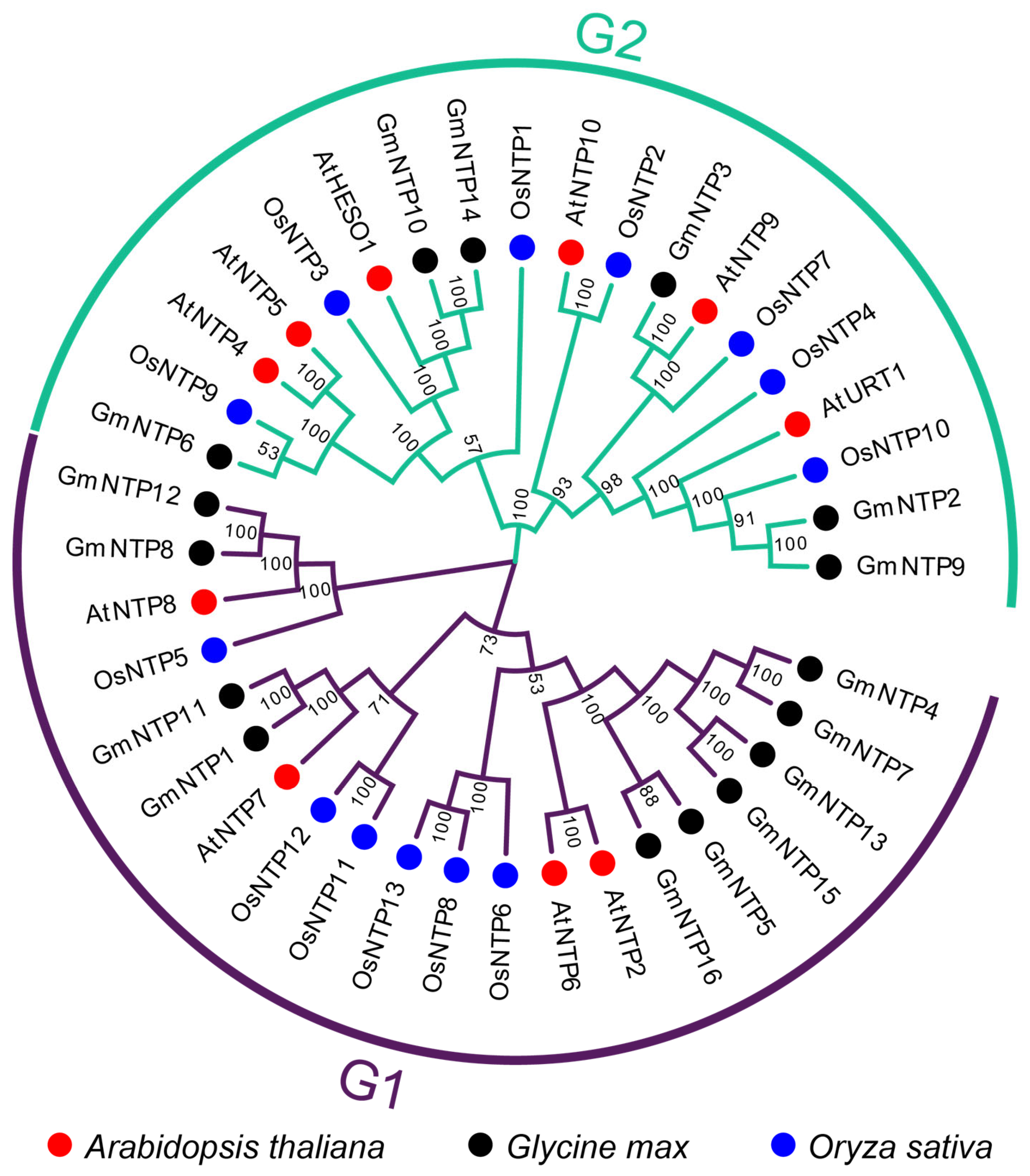
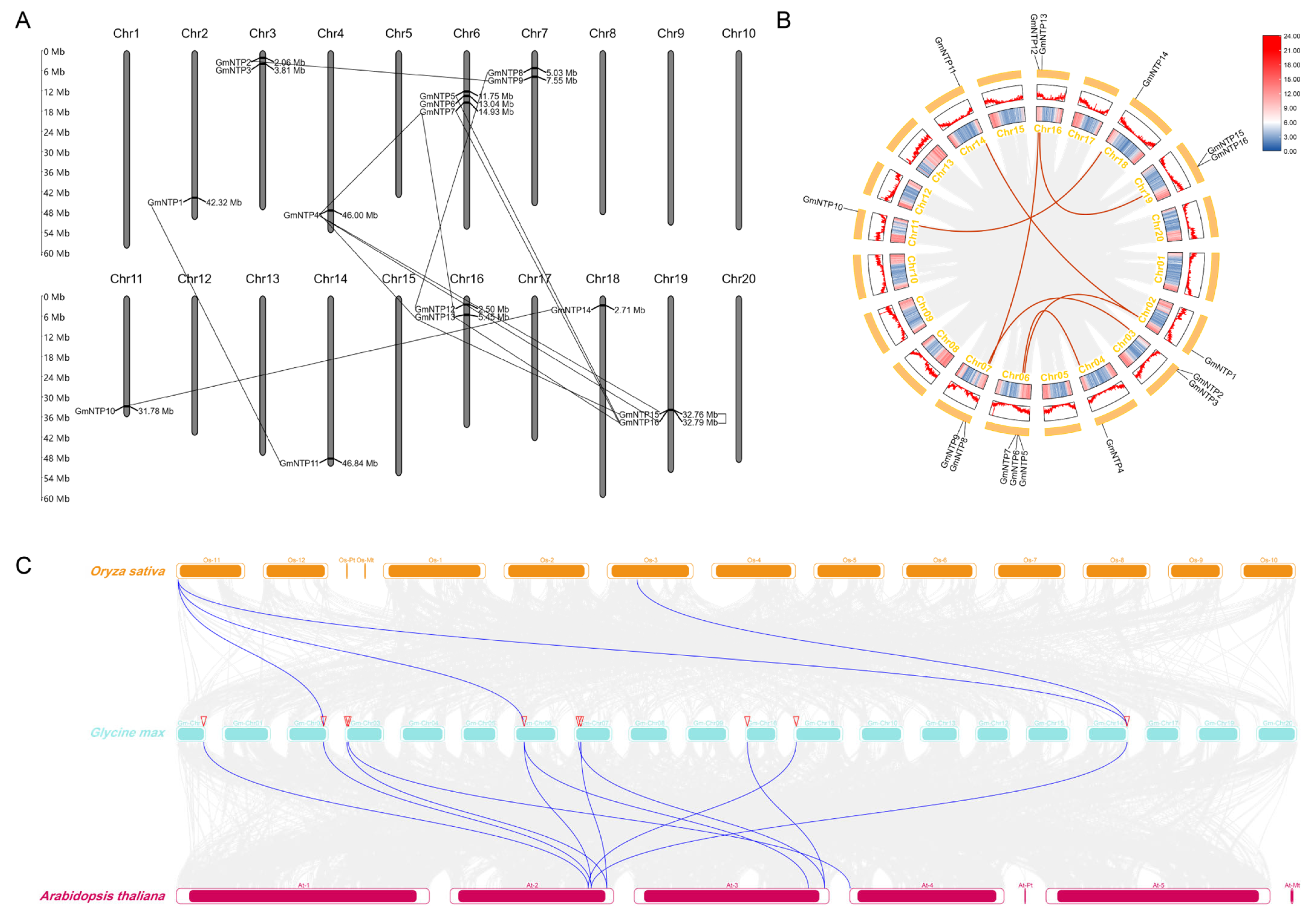
2.2. Phylogenetic Evolution and Chromosomal Localization of GmNTP Genes
2.3. Collinearity Analysis and Evolutionary Relationship of NTP Genes
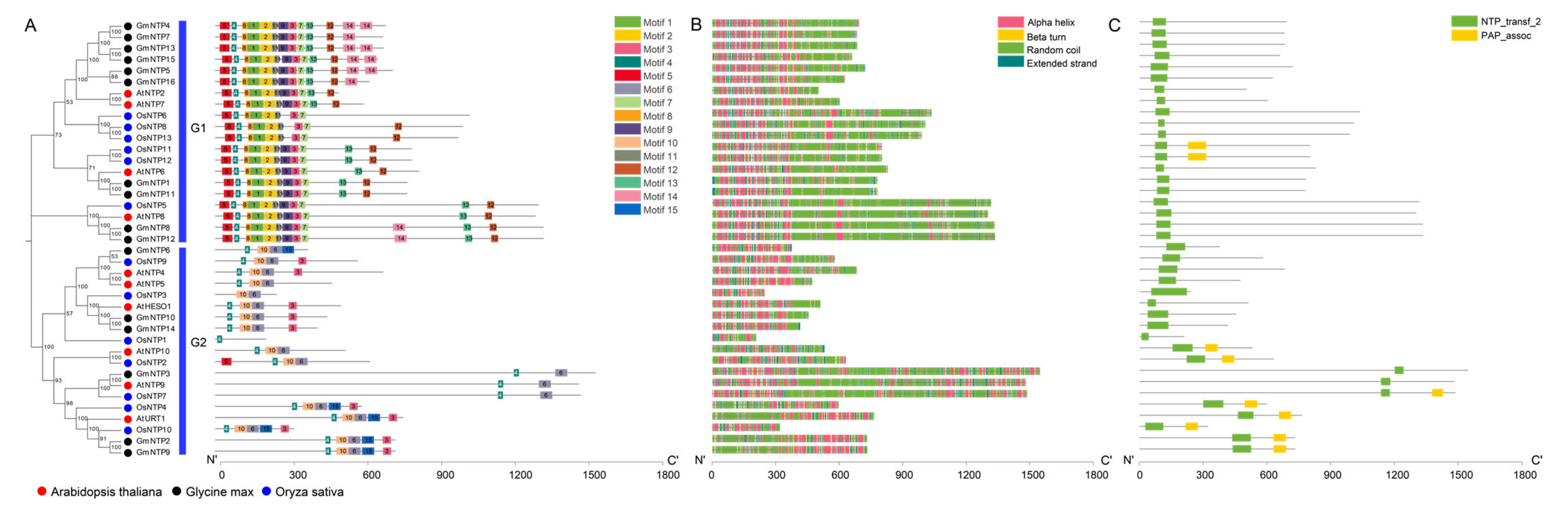
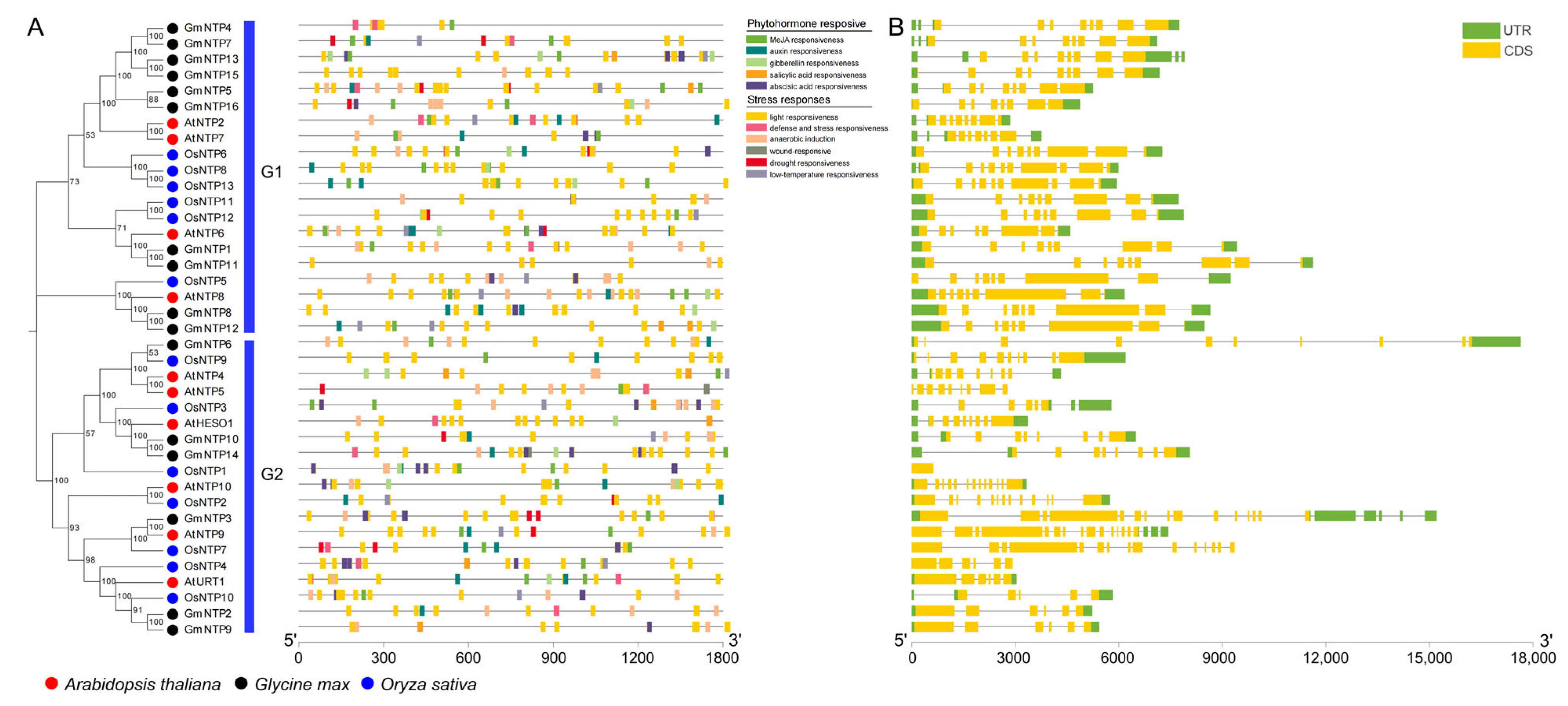
2.4. Conserved Motifs Analysis of NTP Genes
2.5. Secondary Structure and Conserved Domain Analysis of GmNTP
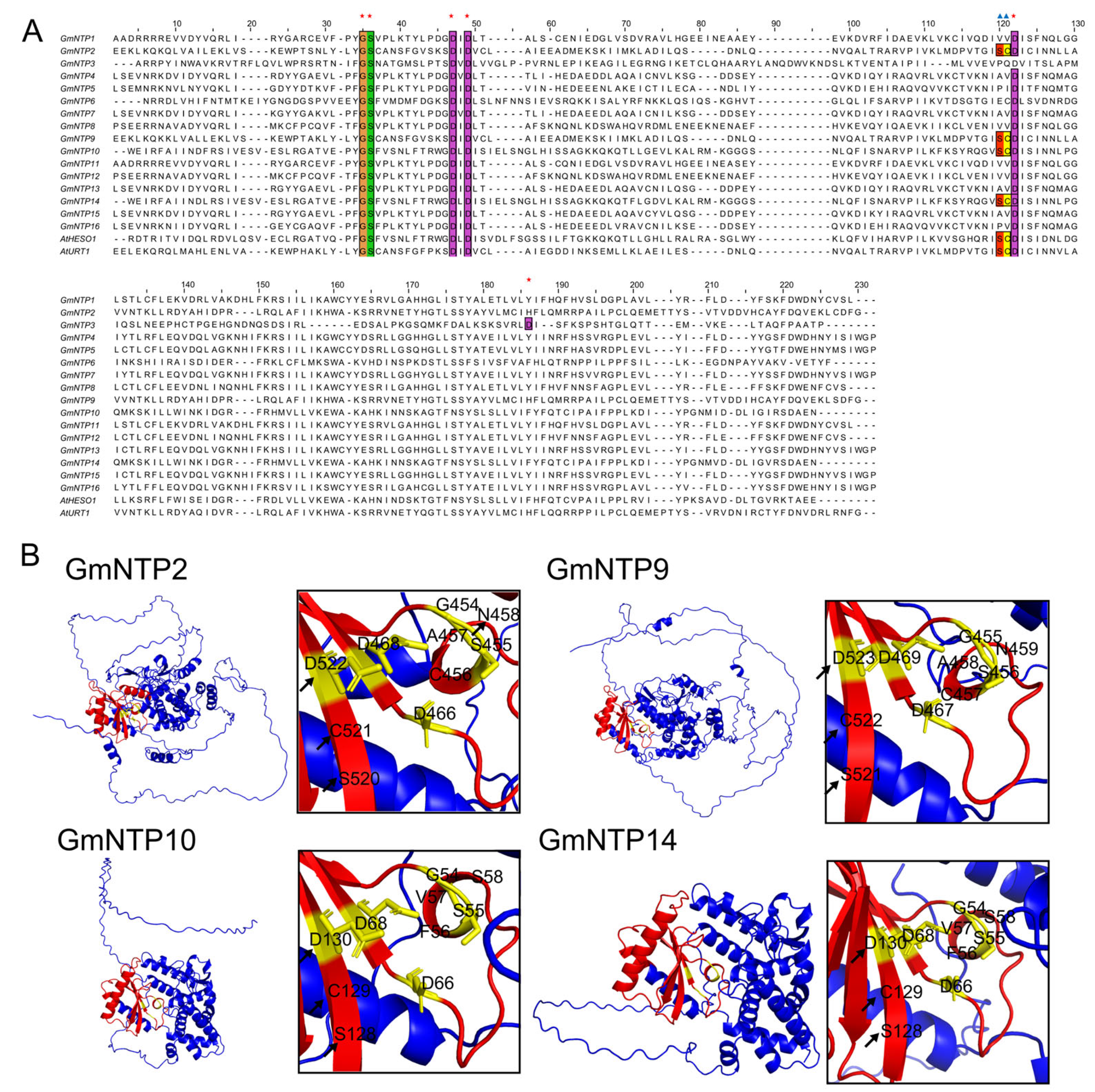
2.6. Tertiary Structure Prediction and Analysis
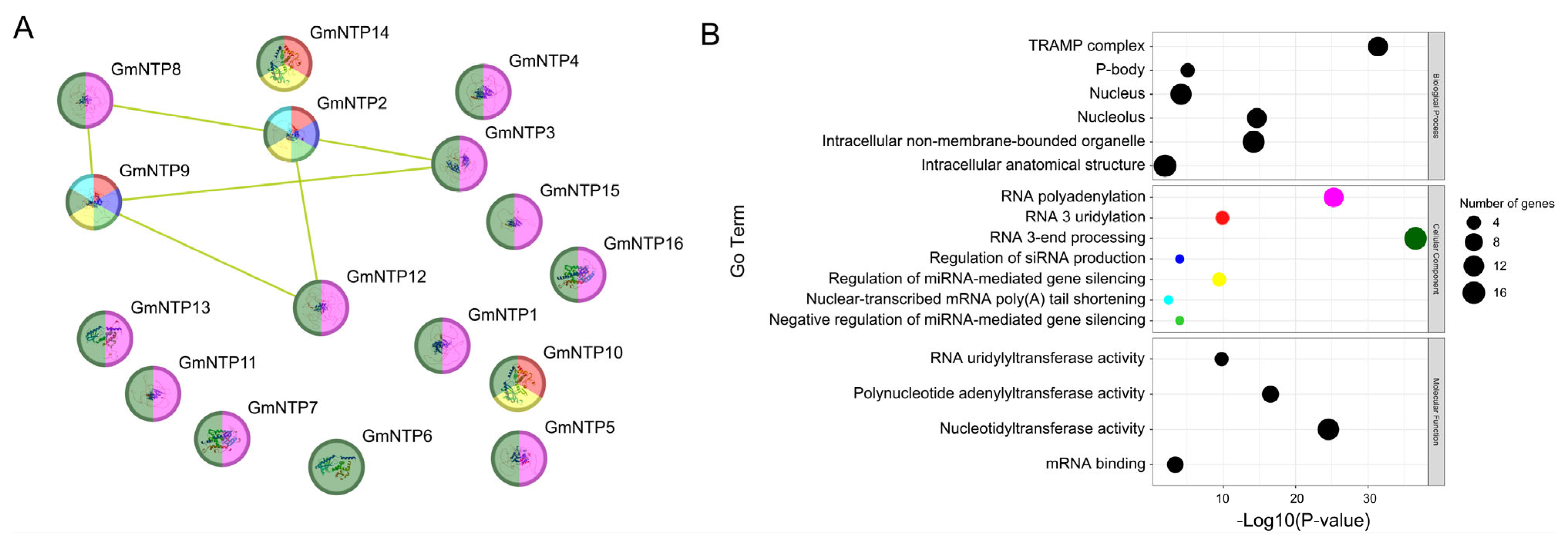
2.7. Protein Interaction Networks and Gene Co-Expression Networks Analysis of GmNTP
2.8. Cis-Element Analysis and Gene Structure of GmNTP Genes
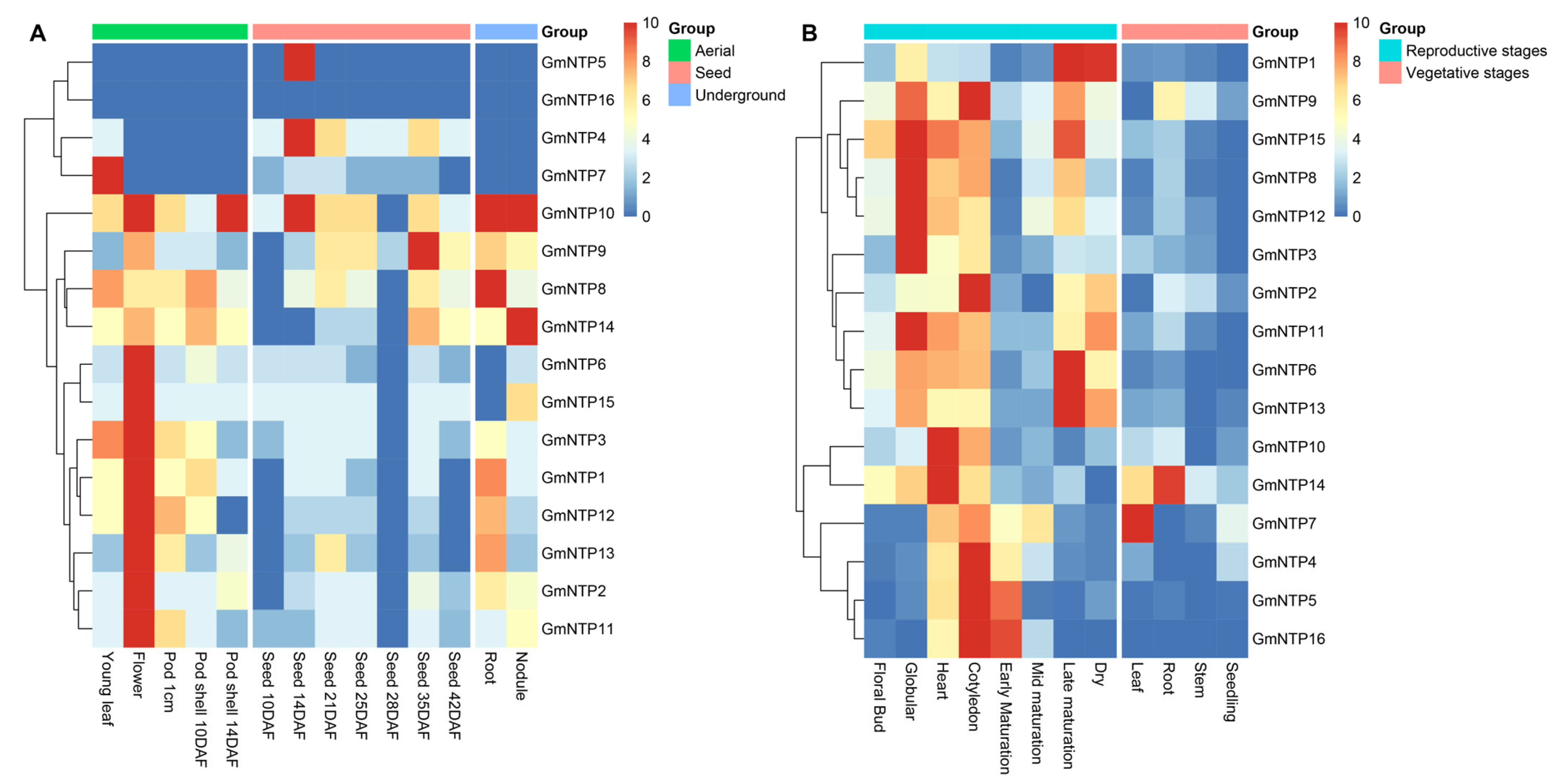
2.9. Expression Analysis of GmNTP Genes at Different Tissues and Developmental Stages
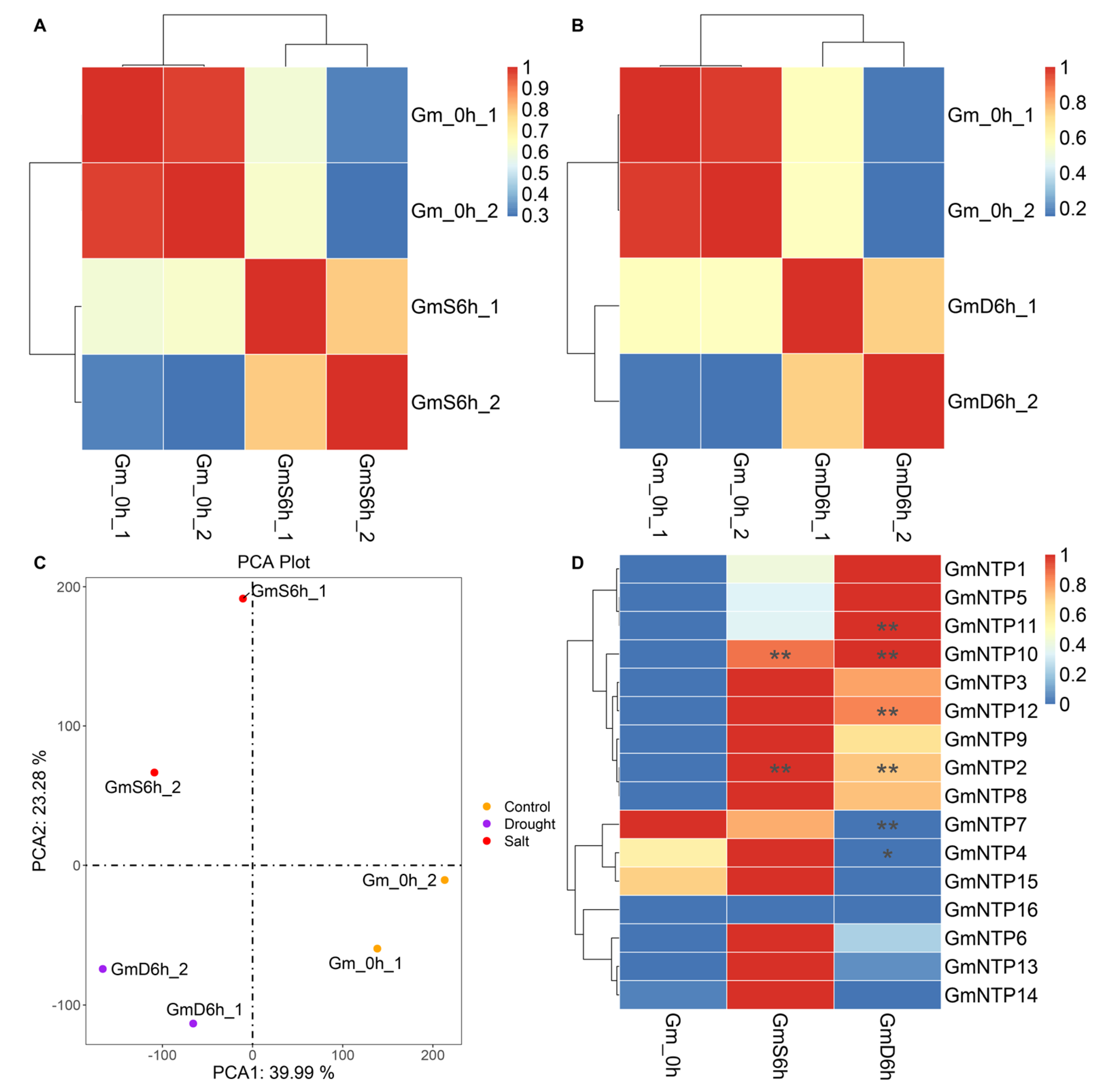
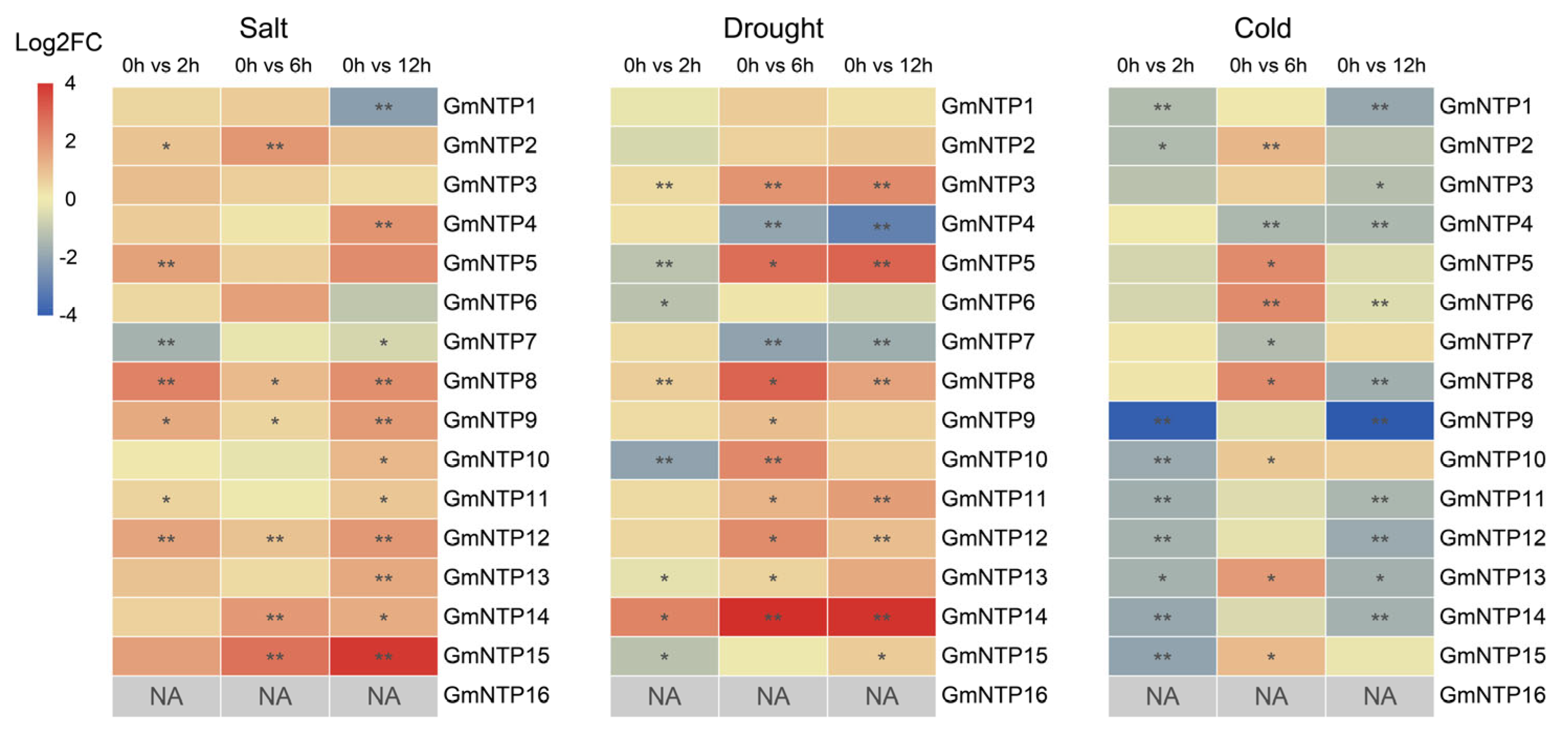
2.10. Expression Analysis of GmNTP Genes under Abiotic Stress
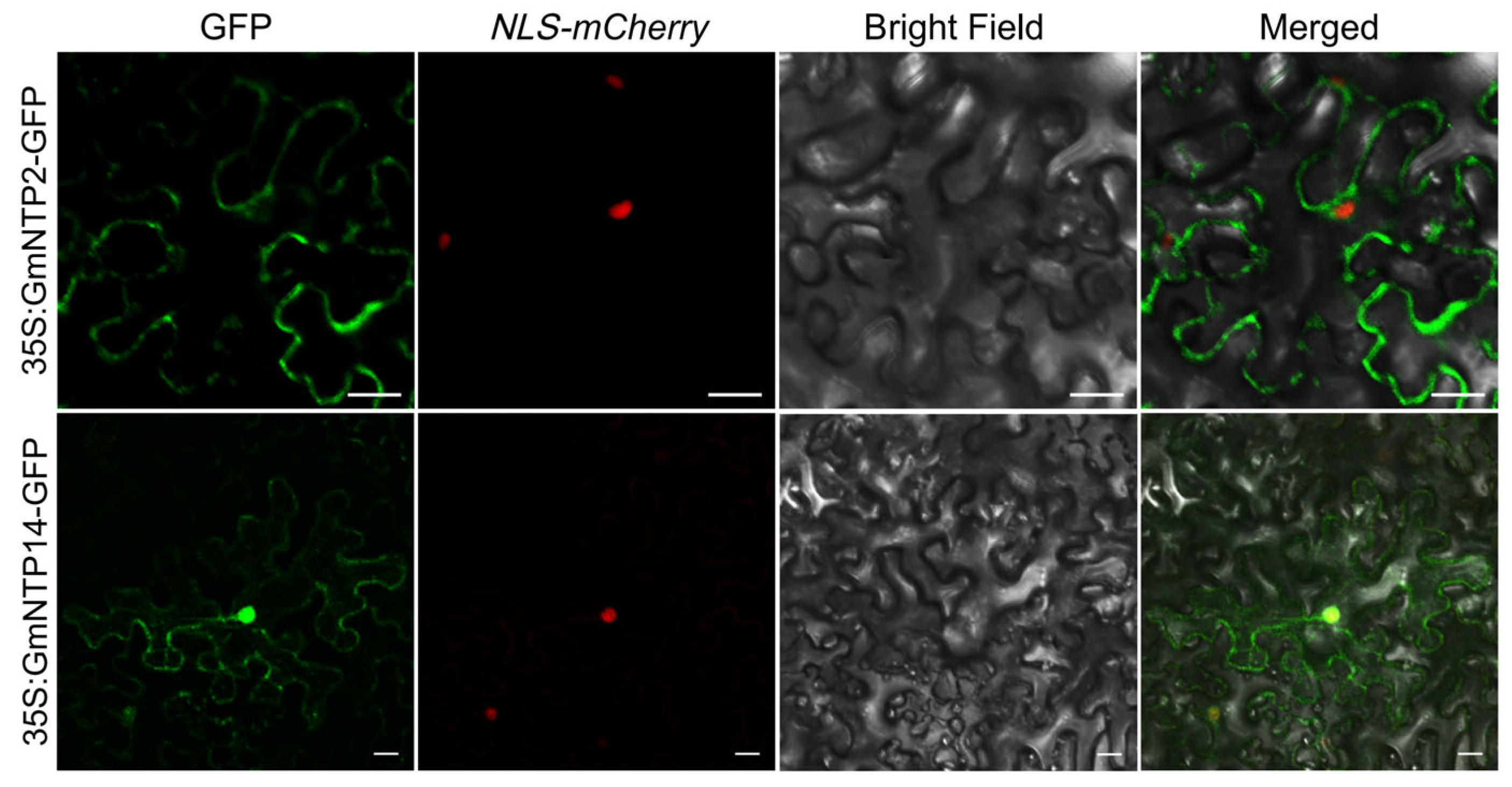
2.11. Subcellular Localization of GmNTP2 and GmNTP14
3. Discussion
4. Materials and Methods
4.1. Identification and Phylogenetic Tree Construction of GmNTPs
4.2. Chromosomal Location and Collinearity Analysis
4.3. Conserved Motifs, Secondary Structure, and Conserved Domains Analysis of GmNTPs
4.4. Gene Structure and Cis-Acting Elements Analysis of GmNTPs
4.5. Tertiary Structure Prediction of GmNTP Proteins
4.6. Protein Interaction Network and Gene Co-Expression Analysis of GmNTP
4.7. Expression Profiles of GmNTP Genes at Different Tissues and Developmental Stages
4.8. Plant Materials and Stress Treatments
4.9. Transcriptome Sequencing, Data Processing, and Gene Expression Analysis
4.10. RNA Extraction and qRT-PCR Analysis
4.11. Construction of Vectors and Subcellular Localization
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, H.; Song, J.; Yue, L.; Mo, X.; Song, J.; Mo, B. Identification and expression profiling of Oryza sativa nucleotidyl transferase protein (NTP) genes under various stress conditions. Gene 2017, 628, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.; Han, B.W.; Hsiao, C.; Zamore, P.D.; Weng, Z.; Hung, J. Tailor: A computational framework for detecting non-templated tailing of small silencing RNAs. Nucleic Acids Res. 2015, 43, e109. [Google Scholar] [CrossRef] [PubMed]
- Knouf, E.C.; Wyman, S.K.; Tewari, M. The human TUT1 nucleotidyl transferase as a global regulator of microRNA abundance. PLoS ONE 2013, 8, e69630. [Google Scholar] [CrossRef] [PubMed]
- Ohnishi, Y.; Totoki, Y.; Toyoda, A.; Watanabe, T.; Yamamoto, Y.; Tokunaga, K.; Sakaki, Y.; Sasaki, H.; Hohjoh, H. Small RNA class transition from siRNA/piRNA to miRNA during pre-implantation mouse development. Nucleic Acids Res. 2010, 38, 5141–5151. [Google Scholar] [CrossRef] [PubMed]
- Song, J.B.; Gao, S.; Sun, D.; Li, H.; Shu, X.X.; Yang, Z.M. miR394 and LCR are involved in Arabidopsis salt and drought stress responses in an abscisic acid-dependent manner. BMC Plant Biol. 2013, 13, 210. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Li, B.; Iwakawa, H.; Pan, Y.; Tang, X.; Ling-hu, Q.; Liu, Y.; Sheng, S.; Feng, L.; Zhang, H.; et al. Plant 22-nt siRNAs mediate translational repression and stress adaptation. Nature 2020, 581, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Javaid, A.; Naqvi, S.H.; Batcho, A.; Kayani, W.K.; Lal, A.; Sajid, I.A.; Nwogwugwu, J.O. Biotic stress triggered small RNA and RNAi defense response in plants. Mol. Biol. Rep. 2020, 47, 5511–5522. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Yu, Y.; Zhai, J.; Ramachandran, V.; Dinh, T.T.; Meyers, B.C.; Mo, B.; Chen, X. The Arabidopsis Nucleotidyl Transferase HESO1 Uridylates Unmethylated Small RNAs to Trigger Their Degradation. Curr. Biol. 2012, 22, 689–694. [Google Scholar] [CrossRef]
- Ibrahim, F.; Rymarquis, L.A.; Kim, E.; Becker, J.; Balassa, E.; Green, P.J.; Cerutti, H. Uridylation of mature miRNAs and siRNAs by the MUT68 nucleotidyltransferase promotes their degradation in Chlamydomonas. Proc. Natl. Acad. Sci. USA 2010, 107, 3906–3911. [Google Scholar] [CrossRef]
- Lu, S.; Sun, Y.H.; Chiang, V.L. Adenylation of plant miRNAs. Nucleic Acids Res. 2009, 37, 1878–1885. [Google Scholar] [CrossRef]
- D’Ambrogio, A.; Gu, W.; Udagawa, T.; Mello, C.C.; Richter, J.D. Specific miRNA Stabilization by Gld2-Catalyzed Monoadenylation. Cell Rep. 2012, 2, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Wang, X.; Song, B.; Gao, L.; Mo, X.; Yue, L.; Yang, H.; Lu, J.; Ren, G.; Mo, B.; et al. Prevalent cytidylation and uridylation of precursor miRNAs in Arabidopsis. Nat. Plants 2019, 5, 1260–1272. [Google Scholar] [CrossRef] [PubMed]
- Wyman, S.K.; Knouf, E.C.; Parkin, R.K.; Fritz, B.R.; Lin, D.W.; Dennis, L.M.; Krouse, M.A.; Webster, P.J.; Tewari, M. Post-transcriptional generation of miRNA variants by multiple nucleotidyl transferases contributes to miRNA transcriptome complexity. Genome Res. 2011, 21, 1450–1461. [Google Scholar] [CrossRef] [PubMed]
- Heo, I.; Joo, C.; Kim, Y.; Ha, M.; Yoon, M.; Cho, J.; Yeom, K.; Han, J.; Kim, V.N. TUT4 in Concert with Lin28 Suppresses MicroRNA Biogenesis through Pre-MicroRNA Uridylation. Cell 2009, 138, 696–708. [Google Scholar] [CrossRef] [PubMed]
- Rissland, O.S.; Mikulasova, A.; Norbury, C.J. Efficient RNA Polyuridylation by Noncanonical Poly(A) Polymerases. Mol. Cell. Biol. 2007, 27, 3612–3624. [Google Scholar] [CrossRef] [PubMed]
- Bortolamiol-Becet, D.; Hu, F.; Jee, D.; Wen, J.; Okamura, K.; Lin, C.; Ameres, S.L.; Lai, E.C. Selective Suppression of the Splicing-Mediated MicroRNA Pathway by the Terminal Uridyltransferase Tailor. Mol. Cell 2015, 59, 217–228. [Google Scholar] [CrossRef]
- van Wolfswinkel, J.C.; Claycomb, J.M.; Batista, P.J.; Mello, C.C.; Berezikov, E.; Ketting, R.F. CDE-1 Affects Chromosome Segregation through Uridylation of CSR-1-Bound siRNAs. Cell 2009, 139, 135–148. [Google Scholar] [CrossRef]
- Yamashita, S.; Tomita, K. Mechanism of U6 snRNA oligouridylation by human TUT1. Nat. Commun. 2023, 14, 4686. [Google Scholar] [CrossRef]
- Menezes, M.R.; Balzeau, J.; Hagan, J.P. 3′ RNA Uridylation in Epitranscriptomics, Gene Regulation, and Disease. Front. Mol. Biosci. 2018, 5, 61. [Google Scholar] [CrossRef]
- Yamashita, S.; Nagaike, T.; Tomita, K. Crystal structure of the Lin28-interacting module of human terminal uridylyltransferase that regulates let-7 expression. Nat. Commun. 2019, 10, 1960. [Google Scholar] [CrossRef]
- Kim, B.; Ha, M.; Loeff, L.; Chang, H.; Simanshu, D.K.; Li, S.; Fareh, M.; Patel, D.J.; Joo, C.; Kim, V.N. TUT7 controls the fate of precursor microRNAs by using three different uridylation mechanisms. EMBO J. 2015, 34, 1801–1815. [Google Scholar] [CrossRef] [PubMed]
- Katoh, T.; Sakaguchi, Y.; Miyauchi, K.; Suzuki, T.; Kashiwabara, S.; Baba, T.; Suzuki, T. Selective stabilization of mammalian microRNAs by 3′ adenylation mediated by the cytoplasmic poly(A) polymerase GLD-2. Gene Dev. 2009, 23, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Mo, B.; Chen, X. Mechanisms that impact microRNA stability in plants. RNA Biol. 2012, 9, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Song, J.; Wang, X.; Mo, X.; Wang, S.; Xu, X.; Lao, K.; Mo, B. Nucleotidyl transferase proteins and the biological responses of Zea mays to abiotic stress. J. Shenzhen Univ. Sci. Eng. 2018, 35, 536. [Google Scholar] [CrossRef]
- Ren, Y.; Ma, X.; Song, B.; Yang, X.; Chen, Y.; Yu, Y.; Chen, X.; Mo, B.; Wang, X. HEN1 SUPPRESSOR1 stabilizes polymerase IV RNAs via uridylation in Arabidopsis. Plant Physiol. 2023, 193, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Tu, B.; Liu, L.; Xu, C.; Zhai, J.; Li, S.; Lopez, M.A.; Zhao, Y.; Yu, Y.; Ramachandran, V.; Ren, G.; et al. Distinct and Cooperative Activities of HESO1 and URT1 Nucleotidyl Transferases in MicroRNA Turnover in Arabidopsis. PLoS Genet. 2015, 11, e1005119. [Google Scholar] [CrossRef] [PubMed]
- Scheer, H.; de Almeida, C.; Ferrier, E.; Simonnot, Q.; Poirier, L.; Pflieger, D.; Sement, F.M.; Koechler, S.; Piermaria, C.; Krawczyk, P.; et al. The TUTase URT1 connects decapping activators and prevents the accumulation of excessively deadenylated mRNAs to avoid siRNA biogenesis. Nat. Commun. 2021, 12, 1298. [Google Scholar] [CrossRef]
- Zuber, H.; Scheer, H.; Joly, A.; Gagliardi, D. Respective Contributions of URT1 and HESO1 to the Uridylation of 5′ Fragments Produced From RISC-Cleaved mRNAs. Front. Plant Sci. 2018, 9, 1438. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, S.; Dou, Y.; Zhang, C.; Chen, X.; Yu, B.; Ren, G. Synergistic and Independent Actions of Multiple Terminal Nucleotidyl Transferases in the 3′ Tailing of Small RNAs in Arabidopsis. PLoS Genet. 2015, 11, e1005091. [Google Scholar] [CrossRef]
- Gu, Z.; Cavalcanti, A.; Chen, F.; Bouman, P.; Li, W. Extent of Gene Duplication in the Genomes of Drosophila, Nematode, and Yeast. Mol. Biol. Evol. 2002, 19, 256–262. [Google Scholar] [CrossRef]
- Yang, X.; Kalluri, U.C.; Jawdy, S.; Gunter, L.E.; Yin, T.; Tschaplinski, T.J.; Weston, D.J.; Ranjan, P.; Tuskan, G.A. The F-Box Gene Family Is Expanded in Herbaceous Annual Plants Relative to Woody Perennial Plants. Plant Physiol. 2008, 148, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wen, X.; Huang, X.; Wei, M.; Chen, H.; Yu, Y.; Dai, S. Genome-Wide Identification and Characterization of TCP Gene Family Members in Melastoma candidum. Molecules 2022, 27, 9036. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Hu, Q.; Cheng, L.; Jiang, Y.; Lv, M.; Liu, Y.; Li, F.; Shi, Y.; Gong, Q. Crystal structure of Arabidopsis terminal uridylyl transferase URT1. Biochem. Biophys. Res. Commun. 2020, 524, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Aphasizhev, R.; Simpson, L. RNA-editing Terminal Uridylyl Transferase 1. J. Biol. Chem. 2004, 279, 24123–24130. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.; Ernst, N.L.; Turley, S.; Stuart, K.D.; Hol, W.G. Structural basis for UTP specificity of RNA editing TUTases from Trypanosoma brucei. EMBO J. 2005, 24, 4007–4017. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, F.; Jiang, Y.; Yu, H.; Xie, C.; Shi, Y.; Gong, Q. Structural insights into a unique preference for 3′ terminal guanine of mirtron in Drosophila TUTase tailor. Nucleic Acids Res. 2019, 47, 495–508. [Google Scholar] [CrossRef]
- Falk, S.; Weir, J.R.; Hentschel, J.; Reichelt, P.; Bonneau, F.; Conti, E. The Molecular Architecture of the TRAMP Complex Reveals the Organization and Interplay of Its Two Catalytic Activities. Mol. Cell 2014, 55, 856–867. [Google Scholar] [CrossRef]
- Wang, X.; Kong, W.; Wang, Y.; Wang, J.; Zhong, L.; Lao, K.; Dong, X.; Zhang, D.; Huang, H.; Mo, B.; et al. Uridylation and the SKI complex orchestrate the Calvin cycle of photosynthesis through RNA surveillance of TKL1 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2022, 119, e2205842119. [Google Scholar] [CrossRef]
- Khalid, M.; Rehman, H.M.; Ahmed, N.; Nawaz, S.; Saleem, F.; Ahmad, S.; Uzair, M.; Rana, I.A.; Atif, R.M.; Zaman, Q.U.; et al. Using Exogenous Melatonin, Glutathione, Proline, and Glycine Betaine Treatments to Combat Abiotic Stresses in Crops. Int. J. Mol. Sci. 2022, 23, 12913. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, Y.; Mou, F.; Tian, Y.; Chen, L.; Zhang, S.; Jiang, Q.; Li, X. Genome-Wide Small RNA Analysis of Soybean Reveals Auxin-Responsive microRNAs that are Differentially Expressed in Response to Salt Stress in Root Apex. Front. Plant Sci. 2016, 6, 1273. [Google Scholar] [CrossRef]
- Ramesh, S.V.; Govindasamy, V.; Rajesh, M.K.; Sabana, A.A.; Praveen, S. Stress-responsive miRNAome of Glycine max (L.) Merrill: Molecular insights and way forward. Planta 2019, 249, 1267–1284. [Google Scholar] [CrossRef] [PubMed]
- Cadavid, I.C.; Da Fonseca, G.C.; Margis, R. HDAC inhibitor affects soybean miRNA482bd expression under salt and osmotic stress. J. Plant Physiol. 2020, 253, 153261. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, T.; Zhang, Y.; Li, Y. Overexpression of soybean miR172c confers tolerance to water deficit and salt stress, but increases ABA sensitivity in transgenic Arabidopsis thaliana. J. Exp. Bot. 2016, 68, w404. [Google Scholar] [CrossRef] [PubMed]
- Sahito, Z.A.; Wang, L.; Sun, Z.; Yan, Q.; Zhang, X.; Jiang, Q.; Ullah, I.; Tong, Y.; Li, X. The miR172c-NNC1 module modulates root plastic development in response to salt in soybean. BMC Plant Biol. 2017, 17, 229. [Google Scholar] [CrossRef] [PubMed]
- Mishra, V.; Singh, A.; Gandhi, N.; Sarkar Das, S.; Yadav, S.; Kumar, A.; Sarkar, A.K. A unique miR775-GALT9 module regulates leaf senescence in Arabidopsis during post-submergence recovery by modulating ethylene and the abscisic acid pathway. Development 2022, 149, dev199974. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Ding, H.; Zhu, J.K.; Zhang, F.; Li, W.X. Involvement of miR169 in the nitrogen-starvation responses in Arabidopsis. New Phytol. 2011, 190, 906–915. [Google Scholar] [CrossRef]
- Jagadeeswaran, G.; Saini, A.; Sunkar, R. Biotic and abiotic stress down-regulate miR398 expression in Arabidopsis. Planta 2009, 229, 1009–1014. [Google Scholar] [CrossRef]
- Ma, C.; Burd, S.; Lers, A. miR408 is involved in abiotic stress responses in Arabidopsis. Plant J. 2015, 84, 169–187. [Google Scholar] [CrossRef]
- Yu, Y.; Ji, L.; Le, B.H.; Zhai, J.; Chen, J.; Luscher, E.; Gao, L.; Liu, C.; Cao, X.; Mo, B.; et al. Correction: ARGONAUTE10 promotes the degradation of miR165/6 through the SDN1 and SDN2 exonucleases in Arabidopsis. PLoS Biol. 2021, 19, e3001120. [Google Scholar] [CrossRef]
- Li, X.; Xie, X.; Li, J.; Cui, Y.; Hou, Y.; Zhai, L.; Wang, X.; Fu, Y.; Liu, R.; Bian, S. Conservation and diversification of the miR166 family in soybean and potential roles of newly identified miR166s. BMC Plant Biol. 2017, 17, 32. [Google Scholar] [CrossRef]
- Yadav, A.; Kumar, S.; Verma, R.; Lata, C.; Sanyal, I.; Rai, S.P. microRNA 166: An evolutionarily conserved stress biomarker in land plants targeting HD-ZIP family. Physiol. Mol. Biol. Plants 2021, 27, 2471–2485. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, R.; Ting, C.S.; King, J. Whole Proteome pI Values Correlate with Subcellular Localizations of Proteins for Organisms within the Three Domains of Life. Genome Res. 2001, 11, 703–709. [Google Scholar] [CrossRef]
- Demuth, J.P.; Hahn, M.W. The life and death of gene families. Bioessays 2009, 31, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, J.P.; Seddon, A.E.; Moghe, G.D.; Simenc, M.C.; Shiu, S. Characteristics of Plant Essential Genes Allow for within- and between-Species Prediction of Lethal Mutant Phenotypes. Plant Cell 2015, 27, 2133–2147. [Google Scholar] [CrossRef] [PubMed]
- Aphasizhev, R.; Aphasizheva, I.; Simpson, L. Multiple terminal uridylyltransferases of trypanosomes. FEBS Lett. 2004, 572, 15–18. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, R.; Jiang, K.; Qi, J.; Hu, Y.; Guo, J.; Zhu, R.; Zhang, T.; Egan, A.N.; Yi, T.; et al. Nuclear phylotranscriptomics and phylogenomics support numerous polyploidization events and hypotheses for the evolution of rhizobial nitrogen-fixing symbiosis in Fabaceae. Mol. Plant 2021, 14, 748–773. [Google Scholar] [CrossRef]
- Yan, Z.; Hossain, M.S.; Wang, J.; Valdés-López, O.; Liang, Y.; Libault, M.; Qiu, L.; Stacey, G. miR172 Regulates Soybean Nodulation. Mol. Plant-Microbe Interact. 2013, 26, 1371–1377. [Google Scholar] [CrossRef]
- Kulcheski, F.R.; de Oliveira, L.F.; Molina, L.G.; Almerão, M.P.; Rodrigues, F.A.; Marcolino, J.; Barbosa, J.F.; Stolf-Moreira, R.; Nepomuceno, A.L.; Marcelino-Guimarães, F.C.; et al. Identification of novel soybean microRNAs involved in abiotic and biotic stresses. BMC Genom. 2011, 12, 307. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Y.; Lu, G.; Sun, X.; Feng, Y.; Yan, S.; Zhang, H.; Jiang, Q.; Zhang, H.; Hu, Z.; et al. Genome-wide identification of microRNAs and phased siRNAs in soybean roots under long-term salt stress. Genes Genom. 2020, 42, 1239–1249. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, Y.; Li, K.; Zou, Y.; Chen, L.; Li, X. Identification of Cold-Responsive miRNAs and Their Target Genes in Nitrogen-Fixing Nodules of Soybean. Int. J. Mol. Sci. 2014, 15, 13596–13614. [Google Scholar] [CrossRef]
- Yu, Y.; Ni, Z.; Wang, Y.; Wan, H.; Hu, Z.; Jiang, Q.; Sun, X.; Zhang, H. Overexpression of soybean miR169c confers increased drought stress sensitivity in transgenic Arabidopsis thaliana. Plant Sci. 2019, 285, 68–78. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Subramanian, B.; Gao, S.; Lercher, M.J.; Hu, S.; Chen, W. Evolview v3: A webserver for visualization, annotation, and management of phylogenetic trees. Nucleic Acids Res. 2019, 47, W270–W275. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Chao, J.T.; Kong, Y.Z.; Wang, Q.; Sun, Y.H.; Gong, D.P.; Lv, J.; Liu, G.S. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Yi Chuan 2015, 37, 91–97. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhang, Z. KaKs_Calculator 3.0: Calculating Selective Pressure on Coding and Non-coding Sequences. Genom. Proteom. Bioinform. 2022, 20, 536–540. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Geourjon, C.; Deléage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef]
- Lescot, M. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef]
- Bramucci, E.; Paiardini, A.; Bossa, F.; Pascarella, S. PyMod: Sequence similarity searches, multiple sequence-structure alignments, and homology modeling within PyMOL. BMC Bioinform. 2012, 13, S2. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2023, 51, D638–D646. [Google Scholar] [CrossRef]
- Obayashi, T.; Hibara, H.; Kagaya, Y.; Aoki, Y.; Kinoshita, K. ATTED-II v11: A Plant Gene Coexpression Database Using a Sample Balancing Technique by Subagging of Principal Components. Plant Cell. Physiol. 2022, 63, 869–881. [Google Scholar] [CrossRef]
- Franz, M.; Lopes, C.T.; Fong, D.; Kucera, M.; Cheung, M.; Siper, M.C.; Huck, G.; Dong, Y.; Sumer, O.; Bader, G.D. Cytoscape.js 2023 update: A graph theory library for visualization and analysis. Bioinformatics 2023, 39, btad031. [Google Scholar] [CrossRef]
- Danzer, J.; Mellott, E.; Bui, A.Q.; Le, B.H.; Martin, P.; Hashimoto, M.; Perez-Lesher, J.; Chen, M.; Pelletier, J.M.; Somers, D.A.; et al. Down-Regulating the Expression of 53 Soybean Transcription Factor Genes Uncovers a Role for SPEECHLESS in Initiating Stomatal Cell Lineages during Embryo Development. Plant Physiol. 2015, 168, 1025–1035. [Google Scholar] [CrossRef]
- Jiang, H.; Lei, R.; Ding, S.W.; Zhu, S. Skewer: A fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chou, K.; Shen, H. Cell-PLoc: A package of Web servers for predicting subcellular localization of proteins in various organisms. Nat. Protoc. 2008, 3, 153–162. [Google Scholar] [CrossRef]
- Blatt, M.R.; Grefen, C. Applications of fluorescent marker proteins in plant cell biology. Methods Mol. Biol. 2014, 1062, 487–507. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome | Domain | Other Domain | Length (aa) | Mass (Da) | pI | Putative Localization |
|---|---|---|---|---|---|---|---|---|
| GmNTP1 | Glyma.02G235800 | 2 | NTP_transf_2 | Unknown | 782 | 87,145 | 6.03 | Nucleus |
| GmNTP2 | Glyma.03G020100 | 3 | NTP_transf_2 | PAP_assoc | 731 | 81,750 | 7.1 | Chloroplast |
| GmNTP3 | Glyma.03G032900 | 3 | NTP_transf_2 | Unknown | 1547 | 170,960 | 6.72 | Nucleus |
| GmNTP4 | Glyma.04G189100 | 4 | NTP_transf_2 | Unknown | 692 | 78,386 | 5.3 | Chloroplast |
| GmNTP5 | Glyma.06G144300 | 6 | NTP_transf_2 | Unknown | 721 | 82,370 | 5.81 | Chloroplast |
| GmNTP6 | Glyma.06G158600 | 6 | NTP_transf_2 | Unknown | 376 | 43,316 | 8.93 | Chloroplast |
| GmNTP7 | Glyma.06G176600 | 6 | NTP_transf_2 | Unknown | 682 | 77,424 | 5.54 | Cytoplasm |
| GmNTP8 | Glyma.07G056700 | 7 | NTP_transf_2 | Unknown | 1334 | 148,607 | 5.82 | Nucleus |
| GmNTP9 | Glyma.07G082200 | 7 | NTP_transf_2 | PAP_assoc | 732 | 81,792 | 7.74 | Chloroplast |
| GmNTP10 | Glyma.11G222700 | 11 | NTP_transf_2 | Unknown | 455 | 51,096 | 9.65 | Chloroplast |
| GmNTP11 | Glyma.14G203800 | 14 | NTP_transf_2 | Unknown | 780 | 86,580 | 6.17 | Nucleus |
| GmNTP12 | Glyma.16G025800 | 16 | NTP_transf_2 | Unknown | 1335 | 148,724 | 5.81 | Nucleus |
| GmNTP13 | Glyma.16G055700 | 16 | NTP_transf_2 | Unknown | 684 | 77,377 | 5.58 | Chloroplast |
| GmNTP14 | Glyma.18G034900 | 18 | NTP_transf_2 | Unknown | 415 | 46,676 | 9.68 | Chloroplast |
| GmNTP15 | Glyma.19G092300 | 19 | NTP_transf_2 | Unknown | 660 | 75,146 | 5.94 | Chloroplast |
| GmNTP16 | Glyma.19G092500 | 19 | NTP_transf_2 | Unknown | 626 | 71,256 | 5.38 | Cytoplasm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, L.; Li, C.; Qin, A.; Liu, Z.; Li, X.; Zeng, L.; Yu, H.; Wang, Y.; Song, J.; Chen, R. Identification and Expression Analysis of the Nucleotidyl Transferase Protein (NTP) Family in Soybean (Glycine max) under Various Abiotic Stresses. Int. J. Mol. Sci. 2024, 25, 1115. https://doi.org/10.3390/ijms25021115
Kang L, Li C, Qin A, Liu Z, Li X, Zeng L, Yu H, Wang Y, Song J, Chen R. Identification and Expression Analysis of the Nucleotidyl Transferase Protein (NTP) Family in Soybean (Glycine max) under Various Abiotic Stresses. International Journal of Molecular Sciences. 2024; 25(2):1115. https://doi.org/10.3390/ijms25021115
Chicago/Turabian StyleKang, Liqing, Changgen Li, Aokang Qin, Zehui Liu, Xuanyue Li, Liming Zeng, Hongyang Yu, Yihua Wang, Jianbo Song, and Rongrong Chen. 2024. "Identification and Expression Analysis of the Nucleotidyl Transferase Protein (NTP) Family in Soybean (Glycine max) under Various Abiotic Stresses" International Journal of Molecular Sciences 25, no. 2: 1115. https://doi.org/10.3390/ijms25021115