
Invertebrate Immunity, Natural Transplantation Immunity, Somatic and Germ Cell Parasitism, and Transposon Defense


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
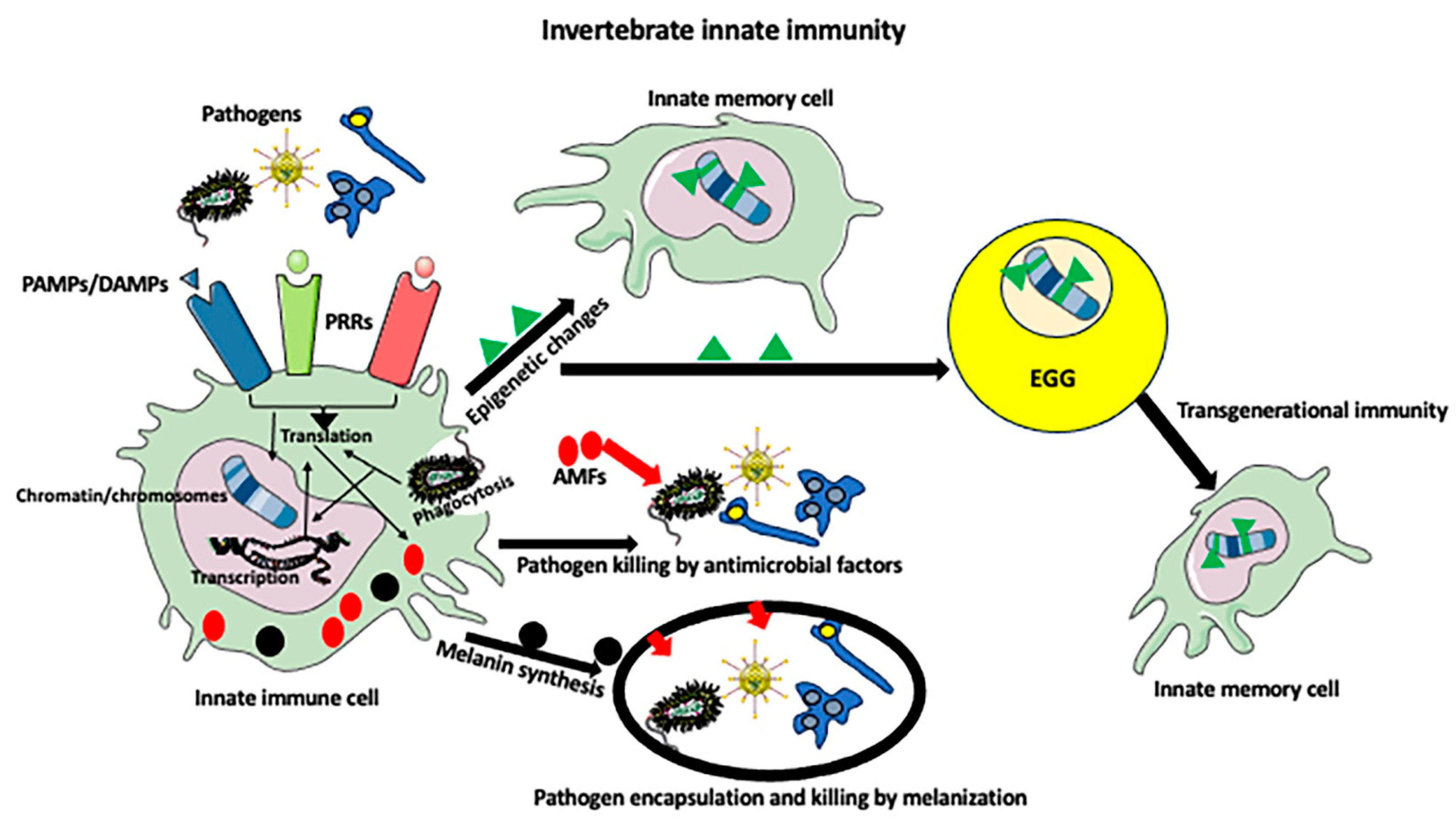
2. Invertebrate Immunity
3. Invertebrate Immune Cells
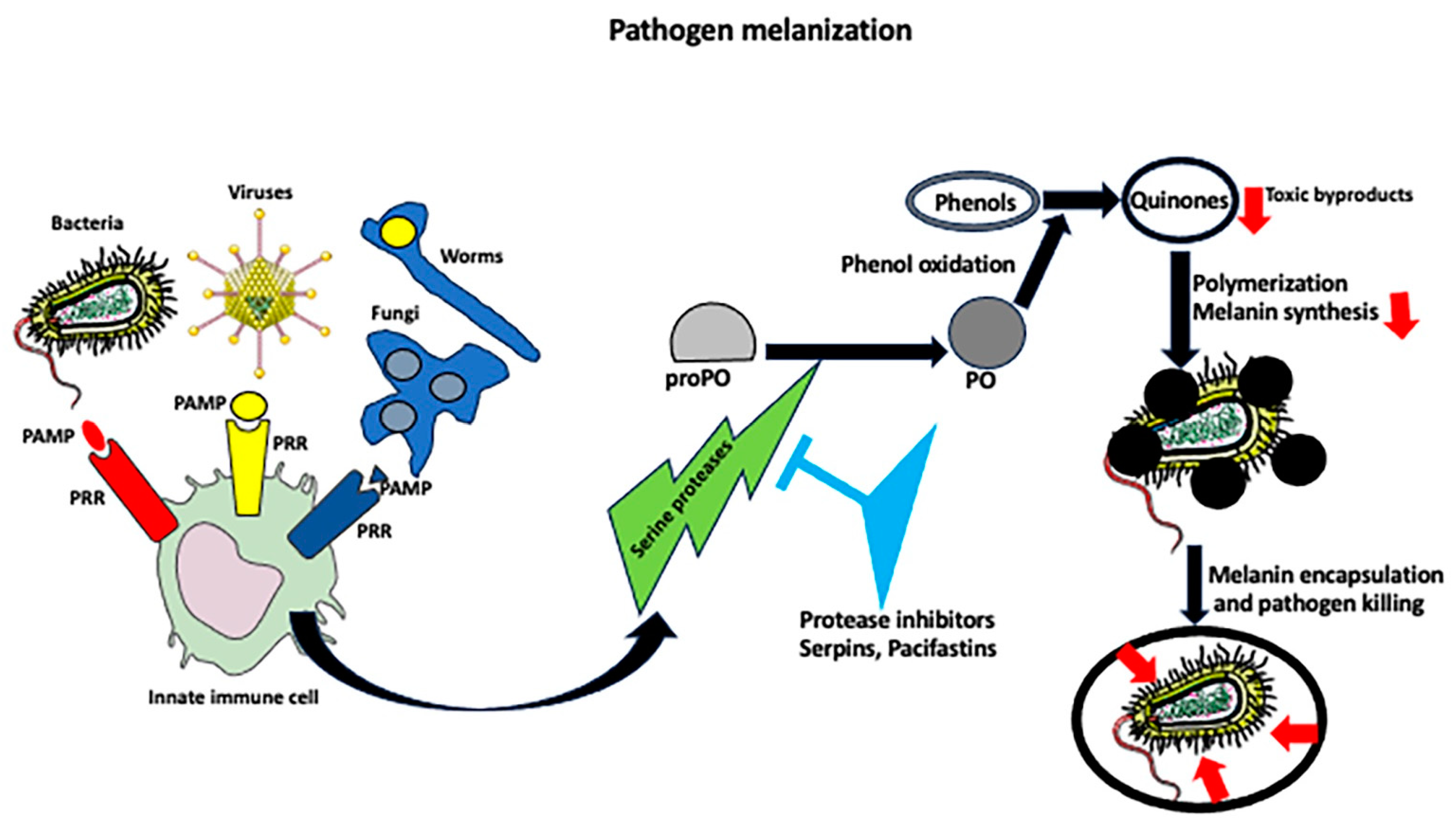
4. Melanization Immune Response
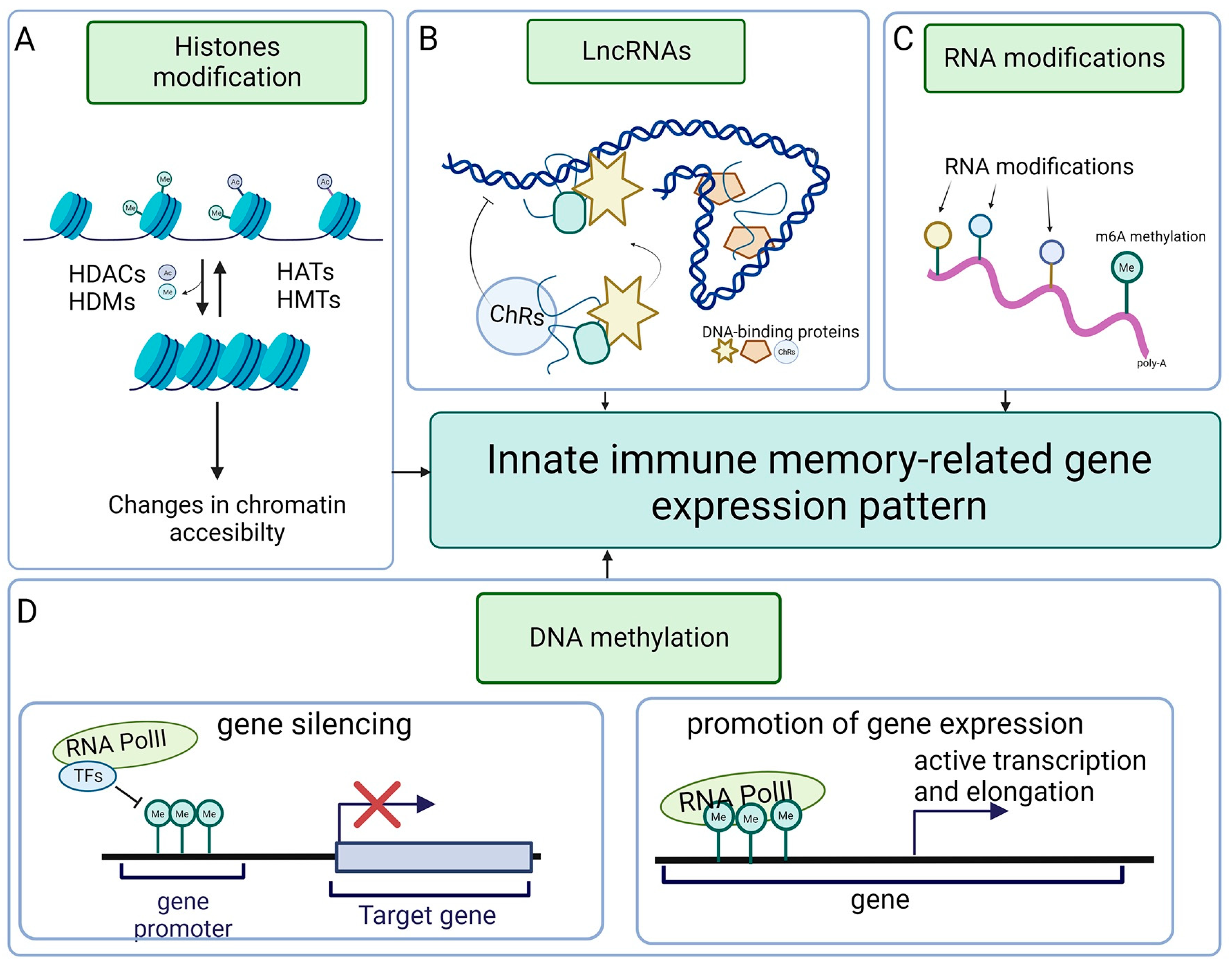
5. Innate Immune Memory
6. Epigenetics of Invertebrate Immunity
7. Intergenerational and Transgenerational Immunity
8. Mechanisms of Self and Non-Self-Recognition, Natural Transplantation, and Germ and Somatic Cell Parasitism
9. Defense against Transposons
10. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Černý, J.; Stříž, I. Adaptive innate immunity or innate adaptive immunity? Clin. Sci. 2019, 133, 1549–1565. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J. Specific memory within innate immune systems. Trends Immunol. 2005, 26, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Sadd, B.M.; Schmid-Hempel, P. Insect immunity shows specificity in protection upon secondary pathogen exposure. Curr. Biol. 2006, 16, 1206–1210. [Google Scholar] [CrossRef] [PubMed]
- Schulenburg, H.; Boehnisch, C.; Michiels, N.K. How do invertebrates generate a highly specific innate immune response? Mol. Immunol. 2007, 44, 3338–3344. [Google Scholar] [CrossRef]
- Mantovani, A.; Garlanda, C. Humoral Innate Immunity and Acute-Phase Proteins. N. Engl. J. Med. 2023, 388, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Kanellopoulos, J.M.; Ojcius, D.M. Development of humoral immunity. Biomed. J. 2019, 42, 207–208. [Google Scholar] [CrossRef] [PubMed]
- Foo, S.S.; Reading, P.C.; Jaillon, S.; Mantovani, A.; Mahalingam, S. Pentraxins and Collectins: Friend or Foe during Pathogen Invasion? Trends Microbiol. 2015, 23, 799–811. [Google Scholar] [CrossRef]
- Zhang, X.L.; Ali, M.A. Ficolins: Structure, function and associated diseases. Adv. Exp. Med. Biol. 2008, 632, 105–115. [Google Scholar]
- Zhu, Q.; Zhang, M.; Shi, M.; Liu, Y.; Zhao, Q.; Wang, W.; Zhang, G.; Yang, L.; Zhi, J.; Zhang, L.; et al. Human B cells have an active phagocytic capability and undergo immune activation upon phagocytosis of Mycobacterium tuberculosis. Immunobiology 2016, 221, 558–567. [Google Scholar] [CrossRef]
- Stögerer, T.; Stäger, S. Innate Immune Sensing by Cells of the Adaptive Immune System. Front. Immunol. 2020, 11, 1081. [Google Scholar] [CrossRef]
- Buchmann, K. Evolution of Innate Immunity: Clues from Invertebrates via Fish to Mammals. Front. Immunol. 2014, 5, 459. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Cho, B.; Lee, D.; Kim, H.; Shim, J.; Nam, J.W. Molecular traces of Drosophila hemocytes reveal transcriptomic conservation with vertebrate myeloid cells. PLoS Genet. 2023, 19, e1011077. [Google Scholar] [CrossRef] [PubMed]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems--not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef]
- Zhao, L.; Niu, J.; Feng, D.; Wang, X.; Zhang, R. Immune functions of pattern recognition receptors in Lepidoptera. Front. Immunol. 2023, 14, 1203061. [Google Scholar] [CrossRef] [PubMed]
- Werling, D.; Jungi, T.W. TOLL-like receptors linking innate and adaptive immune response. Vet. Immunol. Immunopathol. 2003, 91, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Arts, J.A.; Cornelissen, F.H.; Cijsouw, T.; Hermsen, T.; Savelkoul, H.F.; Stet, R.J. Molecular cloning and expression of a Toll receptor in the giant tiger shrimp, Penaeus monodon. Fish Shellfish Immunol. 2007, 23, 504–513. [Google Scholar] [CrossRef]
- Mekata, T.; Kono, T.; Yoshida, T.; Sakai, M.; Itami, T. Identification of cDNA encoding Toll receptor, MjToll gene from kuruma shrimp, Marsupenaeus japonicus. Fish Shellfish Immunol. 2008, 24, 122–133. [Google Scholar] [CrossRef]
- Yang, L.S.; Yin, Z.X.; Liao, J.X.; Huang, X.D.; Guo, C.J.; Weng, S.P.; Chan, S.M.; Yu, X.Q.; He, J.G. A Toll receptor in shrimp. Mol. Immunol. 2007, 44, 1999–2008. [Google Scholar] [CrossRef]
- Yang, C.; Zhang, J.; Li, F.; Ma, H.; Zhang, Q.; Jose Priya, T.A.; Zhang, X.; Xiang, J. A Toll receptor from Chinese shrimp Fenneropenaeus chinensis is responsive to Vibrio anguillarum infection. Fish Shellfish Immunol. 2008, 24, 564–574. [Google Scholar] [CrossRef]
- Alquraini, A.; El Khoury, J. Scavenger receptors. Curr. Biol. 2020, 30, R790–R795. [Google Scholar] [CrossRef]
- Canton, J.; Neculai, D.; Grinstein, S. Scavenger receptors in homeostasis and immunity. Nat. Rev. Immunol. 2013, 13, 621–634. [Google Scholar] [CrossRef] [PubMed]
- PrabhuDas, M.R.; Baldwin, C.L.; Bollyky, P.L.; Bowdish, D.M.E.; Drickamer, K.; Febbraio, M.; Herz, J.; Kobzik, L.; Krieger, M.; Loike, J.; et al. A Consensus Definitive Classification of Scavenger Receptors and Their Roles in Health and Disease. J. Immunol. 2017, 198, 3775–3789. [Google Scholar] [CrossRef]
- Zani, I.A.; Stephen, S.L.; Mughal, N.A.; Russell, D.; Homer-Vanniasinkam, S.; Wheatcroft, S.B.; Ponnambalam, S. Scavenger receptor structure and function in health and disease. Cells 2015, 4, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Patnaik, B.B.; Baliarsingh, S.; Patnaik, H.H.; Sang, M.K.; Park, J.E.; Cho, H.C.; Song, D.K.; Jeong, J.Y.; Hong, C.E.; et al. Transcriptome analysis of the endangered dung beetle Copris tripartitus (Coleoptera: Scarabaeidae) and characterization of genes associated to immunity, growth, and reproduction. BMC Genom. 2023, 24, 94. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Tang, Y.; Li, Z.; Wu, Q.; Qiao, X.; Wan, F.; Qian, W.; Liu, C. Investigation of Immune Responses in Giant African Snail, Achatina immaculata, against a Two-Round Lipopolysaccharide Challenge. Int. J. Mol. Sci. 2023, 24, 12191. [Google Scholar] [CrossRef]
- Pearson, A.; Lux, A.; Krieger, M. Expression cloning of dSR-CI, a class C macrophage-specific scavenger receptor from Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1995, 92, 4056–4060. [Google Scholar] [CrossRef]
- Shi, X.Z.; Yang, M.C.; Kang, X.L.; Li, Y.X.; Hong, P.P.; Zhao, X.F.; Vasta, G.R.; Wang, J.X. Scavenger receptor B2, a type III membrane pattern recognition receptor, senses LPS and activates the IMD pathway in crustaceans. Proc. Natl. Acad. Sci. USA 2023, 120, e2216574120. [Google Scholar] [CrossRef]
- Lyu, B.; Li, J.; Niemeyer, B.; Anderson, D.M.; Beerntsen, B.; Song, Q. Integrative analysis highlights molecular and immune responses of tick Amblyomma americanum to Escherichia coli challenge. Front. Cell. Infect. Microbiol. 2023, 13, 1236785. [Google Scholar] [CrossRef]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef]
- Alejandro, A.-D.; Lilia, J.-P.; Jesús, M.-B.; Henry, R.M. The IMD and Toll canonical immune pathways of Triatoma pallidipennis are preferentially activated by Gram-negative and Gram-positive bacteria, respectively, but cross-activation also occurs. Parasites Vectors 2022, 15, 256–268. [Google Scholar] [CrossRef]
- Belvin, M.P.; Anderson, K.V. A Conserved Signaling Pathway: The Drosophila Toll-Dorsal Pathway. Annu. Rev. Cell Dev. Biol. 1996, 12, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Gobert, V.; Gottar, M.; Matskevich, A.A.; Rutschmann, S.; Royet, J.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual Activation of the Drosophila Toll Pathway by Two Pattern Recognition Receptors. Science 2003, 302, 2126–2130. [Google Scholar] [CrossRef] [PubMed]
- Hetru, C.; Hoffmann, J.A. NF-kappaB in the Immune Response of Drosophila. Cold Spring Harb. Perspect. Biol. 2009, 1, a000232. [Google Scholar] [CrossRef]
- Jang, H.A.; Kojour, M.A.M.; Patnaik, B.B.; Han, Y.S.; Jo, Y.H. Current Status of Immune Deficiency Pathway in Tenebrio molitor Innate Immunity. Front. Immunol. 2022, 13, 906192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Wang, L.Z.; Gu, W.B.; Wang, C.; Zhu, Q.H.; Liu, Z.P.; Chen, Y.Y.; Shu, M.A. Identification and functional analysis of immune deficiency (IMD) from Scylla paramamosain: The first evidence of IMD signaling pathway involved in immune defense against bacterial infection in crab species. Fish Shellfish Immunol. 2018, 81, 150–160. [Google Scholar] [CrossRef]
- De Gregorio, E.; Spellman, P.T.; Tzou, P.; Rubin, G.M.; Lemaitre, B. The Toll and Imd pathways are the major regulators of the immune response in Drosophila. EMBO J. 2002, 21, 2568–2579. [Google Scholar] [CrossRef]
- Brianik, C.J.; Bouallegui, Y.; Allam, B. Triploid animals, a potential model for ETosis research: Influence of polyploidy on the formation and efficacy of extracellular traps in the eastern oyster. Fish Shellfish Immunol. 2023, 9, 108992. [Google Scholar] [CrossRef]
- Granger, V.; Faille, D.; Marani, V.; Noël, B.; Gallais, Y.; Szely, N.; Flament, H.; Pallardy, M.; Chollet-Martin, S.; de Chaisemartin, L. Human blood monocytes are able to form extracellular traps. J. Leukoc. Biol. 2017, 102, 775–781. [Google Scholar] [CrossRef]
- Kloc, M.; Uosef, A.; Kubiak, J.Z.; Ghobrial, R.M. Macrophage Proinflammatory Responses to Microorganisms and Transplanted Organs. Int. J. Mol. Sci. 2020, 21, 9669. [Google Scholar] [CrossRef]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.H. Adaptive evolution in two large families of ubiquitin-ligase adapters in nematodes and plants. Genome Res. 2006, 16, 1017–1030. [Google Scholar] [CrossRef] [PubMed]
- Cui, B.; Lin, H.; Yu, J.; Yu, J.; Hu, Z. Autophagy and the Immune Response. Adv. Exp. Med. Biol. 2019, 1206, 595–634. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.J.; Hansen, M.; Troemel, E. Autophagy and innate immunity: Insights from invertebrate model organisms. Autophagy 2018, 14, 233–242. [Google Scholar] [CrossRef]
- Bosch, T.C.; Augustin, R.; Anton-Erxleben, F.; Fraune, S.; Hemmrich, G.; Zill, H.; Rosenstiel, P.; Jacobs, G.; Schreiber, S.; Leippe, M.; et al. Uncovering the evolutionary history of innate immunity: The simple metazoan Hydra uses epithelial cells for host defence. Dev. Comp. Immunol. 2009, 33, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Chapman, J.A.; Kirkness, E.F.; Simakov, O.; Hampson, S.E.; Mitros, T.; Weinmaier, T.; Rattei, T.; Balasubramanian, P.G.; Borman, J.; Busam, D.; et al. The dynamic genome of Hydra. Nature 2010, 464, 592–596. [Google Scholar] [CrossRef]
- Schröder, K.; Bosch, T.C. The Origin of Mucosal Immunity: Lessons from the Holobiont Hydra. mBio 2016, 7, e01184-16. [Google Scholar] [CrossRef] [PubMed]
- Glauber, K.M.; Dana, C.E.; Steele, R.E. Hydra. Curr Biol. 2010, 20, R964–R965. [Google Scholar] [CrossRef]
- Hwang, J.S.; Ohyanagi, H.; Hayakawa, S.; Osato, N.; Nishimiya-Fujisawa, C.; Ikeo, K.; David, C.N.; Fujisawa, T.; Gojobori, T. The evolutionary emergence of cell type-specific genes inferred from the gene expression analysis of Hydra. Proc. Natl. Acad. Sci. USA 2007, 104, 14735–14740. [Google Scholar] [CrossRef]
- Boehm, A.M.; Khalturin, K.; Anton-Erxleben, F.; Hemmrich, G.; Klostermeier, U.C.; Lopez-Quintero, J.A.; Oberg, H.H.; Puchert, M.; Rosenstiel, P.; Wittlieb, J.; et al. FoxO is a critical regulator of stem cell maintenance in immortal Hydra. Proc. Natl. Acad. Sci. USA 2012, 109, 19697–19702. [Google Scholar] [CrossRef]
- Cho, B.; Yoon, S.H.; Lee, D.; Koranteng, F.; Tattikota, S.G.; Cha, N.; Shin, M.; Do, H.; Hu, Y.; Oh, S.Y.; et al. Single-cell transcriptome maps of myeloid blood cell lineages in Drosophila. Nat. Commun. 2020, 11, 4483. [Google Scholar] [CrossRef] [PubMed]
- Tattikota, S.G.; Cho, B.; Liu, Y.; Hu, Y.; Barrera, V.; Steinbaugh, M.J.; Yoon, S.H.; Comjean, A.; Li, F.; Dervis, F.; et al. A single-cell survey of Drosophila blood. eLife 2020, 9, e54818. [Google Scholar] [CrossRef] [PubMed]
- Rizki, M.T.; Rizki, R.M. Functional significance of the crystal cells in the larva of Drosophila melanogaster. J. Biophys. Biochem. Cytol. 1959, 5, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Stofanko, M.; Kwon, S.Y.; Badenhorst, P. Lineage tracing of lamellocytes demonstrates Drosophila macrophage plasticity. PLoS ONE 2010, 5, e14051. [Google Scholar] [CrossRef]
- Shan, T.; Wang, Y.; Bhattarai, K.; Jiang, H. An evolutionarily conserved serine protease network mediates melanization and Toll activation in Drosophila. Sci. Adv. 2023, 9, eadk2756. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, F.; Cao, X.; Huang, R.; Paskewitz, S.; Hartson, S.D.; Kanost, M.R.; Jiang, H. Inhibition of immune pathway-initiating hemolymph protease-14 by Manduca sexta serpin-12, a conserved mechanism for the regulation of melanization and Toll activation in insects. Insect. Biochem. Mol. Biol. 2020, 116, 103261. [Google Scholar] [CrossRef]
- Kan, H.; Kim, C.H.; Kwon, H.M.; Park, J.W.; Roh, K.B.; Lee, H.; Park, B.J.; Zhang, R.; Zhang, J.; Söderhäll, K.; et al. Molecular control of phenoloxidase-induced melanin synthesis in an insect. J. Biol. Chem. 2008, 283, 25316–25323. [Google Scholar] [CrossRef]
- Cattenoz, P.B.; Monticelli, S.; Pavlidaki, A.; Giangrande, A. Toward a Consensus in the Repertoire of Hemocytes Identified in Drosophila. Front. Cell Dev. Biol. 2021, 9, 643712. [Google Scholar] [CrossRef]
- Yang, P.; Chen, Y.; Huang, Z.; Xia, H.; Cheng, L.; Wu, H.; Zhang, Y.; Wang, F. Single-cell RNA sequencing analysis of shrimp immune cells identifies macrophage-like phagocytes. eLife 2022, 11, e80127. [Google Scholar] [CrossRef]
- Li, C.; Hong, P.P.; Yang, M.C.; Zhao, X.F.; Wang, J.X. FOXO regulates the expression of antimicrobial peptides and promotes phagocytosis of hemocytes in shrimp antibacterial immunity. PLoS Pathog. 2021, 17, e1009479. [Google Scholar] [CrossRef]
- Barretto, E.C.; Polan, D.M.; Beevor-Potts, A.N.; Lee, B.; Grewal, S.S. Tolerance to Hypoxia Is Promoted by FOXO Regulation of the Innate Immunity Transcription Factor NF-κB/Relish in Drosophila. Genetics 2020, 215, 1013–1025. [Google Scholar] [CrossRef]
- Dejean, A.S.; Hedrick, S.M.; Kerdiles, Y.M. Highly specialized role of Forkhead box O transcription factors in the immune system. Antioxid. Redox Signal. 2011, 14, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Rong, S.J.; Yang, C.L.; Wang, F.X.; Sun, F.; Luo, J.H.; Yue, T.T.; Yang, P.; Yu, Q.; Zhang, S.; Wang, C.Y. The Essential Role of FoxO1 in the Regulation of Macrophage Function. Biomed. Res. Int. 2022, 2022, 1068962. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.; Taketa, D.A.; Madhu, R.; Kassmer, S.; Loerke, D.; Valentine, M.T.; Tomaso, A.W. Vascular Aging in the Invertebrate Chordate, Botryllus schlosseri. Front. Mol. Biosci. 2021, 8, 626827. [Google Scholar] [CrossRef]
- Rosental, B.; Kowarsky, M.; Seita, J.; Corey, D.M.; Ishizuka, K.J.; Palmeri, K.J.; Chen, S.Y.; Sinha, R.; Okamoto, J.; Mantalas, G.; et al. Complex mammalian-like haematopoietic system found in a colonial chordate. Nature 2018, 564, 425–429. [Google Scholar] [CrossRef]
- Cinege, G.; Magyar, L.B.; Kovács, A.L.; Lerner, Z.; Juhász, G.; Lukacsovich, D.; Winterer, J.; Lukacsovich, T.; Hegedűs, Z.; Kurucz, É.; et al. Broad Ultrastructural and Transcriptomic Changes Underlie the Multinucleated Giant Hemocyte Mediated Innate Immune Response against Parasitoids. J. Innate Immun. 2022, 14, 335–354. [Google Scholar] [CrossRef] [PubMed]
- Kloc, M.; Uosef, A.; Subuddhi, A.; Kubiak, J.Z.; Piprek, R.P.; Ghobrial, R.M. Giant Multinucleated Cells in Aging and Senescence-An Abridgement. Biology 2022, 11, 1121. [Google Scholar] [CrossRef]
- Barletta, A.B.F.; Saha, B.; Trisnadi, N.; Talyuli, O.A.C.; Raddi, G.; Barillas-Mury, C. Hemocyte differentiation to the megacyte lineage enhances mosquito immunity against Plasmodium. eLife 2022, 11, e81116. [Google Scholar] [CrossRef]
- Nakhleh, J.; El Moussawi, L.; Osta, M.A. Chapter Three—The Melanization Response in Insect Immunity. In Advances in Insect Physiology; Ligoxygakis, P., Ed.; Academic Press: Cambridge, MA, USA, 2017; Volume 52, pp. 83–109. ISSN 0065-2806. ISBN 9780128117750. [Google Scholar] [CrossRef]
- Tang, H. Regulation and function of the melanization reaction in Drosophila. Fly 2009, 3, 105–111. [Google Scholar] [CrossRef]
- Marieshwari, B.N.; Bhuvaragavan, S.; Sruthi, K.; Mullainadhan, P.; Janarthanan, S. Insect phenoloxidase and its diverse roles: Melanogenesis and beyond. J. Comp. Physiol. B 2023, 193, 1–23. [Google Scholar] [CrossRef]
- Tang, H.; Kambris, Z.; Lemaitre, B.; Hashimoto, C. A serpin that regulates immune melanization in the respiratory system of Drosophila. Dev. Cell 2008, 15, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.F.Q.; Dragotakes, Q.; Kulkarni, M.; Hardwick, J.M.; Casadevall, A. Galleria mellonella immune melanization is fungicidal during infection. Commun. Biol. 2022, 5, 1364. [Google Scholar] [CrossRef] [PubMed]
- Tsatmali, M.; Ancans, J.; Thody, A.J. Melanocyte function and its control by melanocortin peptides. J. Histochem. Cytochem. 2002, 50, 125–133. [Google Scholar] [CrossRef]
- Ligoxygakis, P.; Pelte, N.; Ji, C.; Leclerc, V.; Duvic, B.; Belvin, M.; Jiang, H.; Hoffmann, J.A.; Reichhart, J.M. A serpin mutant links Toll activation to melanization in the host defence of Drosophila. EMBO J. 2002, 21, 6330–6337. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.G.; Light, A.; Nossal, G.J.; Tarlinton, D.M. The extent of affinity maturation differs between the memory and antibody-forming cell compartments in the primary immune response. EMBO J. 1997, 16, 2996–3006. [Google Scholar] [CrossRef]
- Cai, Z.; Xing, R.; Liu, J.; Xing, F. Commentary: PIRs Mediate Innate Myeloid Cell Memory to Nonself MHC Molecules. Front. Immunol. 2021, 12, 721344. [Google Scholar] [CrossRef]
- Dai, H.; Lan, P.; Zhao, D.; Abou-Daya, K.; Liu, W.; Chen, W.; Friday, A.J.; Williams, A.L.; Sun, T.; Chen, J.; et al. PIRs mediate innate myeloid cell memory to nonself MHC molecules. Science 2020, 368, 1122–1127. [Google Scholar] [CrossRef]
- Lanz-Mendoza, H.; Contreras-Garduño, J. Innate immune memory in invertebrates: Concept and potential mechanisms. Dev. Comp. Immunol. 2022, 127, 104285. [Google Scholar] [CrossRef]
- Wu, C.; Xu, Y.; Zhao, Y. Two kinds of macrophage memory: Innate and adaptive immune-like macrophage memory. Cell Mol. Immunol. 2022, 19, 852–854. [Google Scholar] [CrossRef]
- Yao, Y.; Jeyanathan, M.; Haddadi, S.; Barra, N.G.; Vaseghi-Shanjani, M.; Damjanovic, D.; Lai, R.; Afkhami, S.; Chen, Y.; Dvorkin-Gheva, A.; et al. Induction of autonomous memory alveolar macrophages requires T cell help and is critical to trained immunity. Cell 2018, 175, 1634–1650.e1617. [Google Scholar] [CrossRef]
- Arts, R.J.; Carvalho, A.; La Rocca, C.; Palma, C.; Rodrigues, F.; Silvestre, R.; Kleinnijenhuis, J.; Lachmandas, E.; Gonçalves, L.G.; Belinha, A.; et al. Immunometabolic pathways in BCG-induced trained immunity. Cell Rep. 2016, 17, 2562–2571. [Google Scholar] [CrossRef]
- Blok, B.A.; Arts, R.J.W.; van Crevel, R.; Benn, C.S.; Netea, M.G. Trained innate immunity as underlying mechanism for the long-term, nonspecific effects of vaccines. J. Leukoc. Biol. 2015, 98, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; van Loenhout, J.; de Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proc. Natl. Acad. Sci. USA 2012, 109, 17537–17542. [Google Scholar] [CrossRef] [PubMed]
- Melillo, D.; Marino, R.; Italiani, P.; Boraschi, D. Innate Immune Memory in Invertebrate Metazoans: A Critical Appraisal. Front. Immunol. 2018, 9, 1915. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Quintin, J.; Van Der Meer, J.W.M. Trained immunity: A memory for innate host defense. Cell Host. Microbe 2011, 9, 355–361. [Google Scholar] [CrossRef]
- Tribouley, J.; Tribouley-Duret, J.; Appriou, M. Effect of Bacillus Callmette Guerin (BCG) on the receptivity of nude mice to Schistosoma mansoni. C R. Seances Soc. Biol. Fil. 1978, 172, 902. [Google Scholar]
- van’t Wout, J.W.; Poell, R.; van Furth, R. The role of BCG/PPD-activated macrophages in resistance against systemic candidiasis in mice. Scand J. Immunol. 1992, 36, 713–719. [Google Scholar] [CrossRef]
- Ligoxygakis, P. Immunity: Insect immune memory goes viral. Curr. Biol. 2017, 27, R1218–R1220. [Google Scholar] [CrossRef]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating immune cells mediate a systemic RNAi-based adaptive antiviral response in Drosophila. Cell 2017, 169, 314–325. [Google Scholar] [CrossRef]
- Whitfield, Z.J.; Dolan, P.T.; Kunitomi, M.; Tassetto, M.; Seetin, M.G.; Oh, S.; Heiner, C.; Paxinos, E.; Andino, R. The diversity, structure, and function of heritable adaptive immunity sequences in the Aedes aegypti genome. Curr. Biol. 2017, 27, 3511–3519. [Google Scholar] [CrossRef]
- Conrath, U. Systemic acquired resistance. Plant Signal. Behav. 2006, 1, 179–184. [Google Scholar] [CrossRef]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.; Robin, G.P. Systemic signaling during plant defense. Curr. Opin. Plant Biol. 2013, 16, 527–533. [Google Scholar] [CrossRef]
- Luna, E.; Ton, J. The epigenetic machinery controlling transgenerational systemic acquired resistance. Plant Signal. Behav. 2012, 7, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Reimer-Michalski, E.-M.; Conrath, U. Innate immune memory in plants. Semin. Immunol. 2016, 28, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Loscalzo, J.; Handy, D.E. Epigenetic modifications: Basic mechanisms and role in cardiovascular disease (2013 Grover Conference series). Pulm. Circ. 2014, 4, 169–174. [Google Scholar] [CrossRef]
- Mehta, S.; Jeffrey, K.L. Beyond receptors and signaling: Epigenetic factors in the regulation of innate immunity. Immunol. Cell Biol. 2015, 93, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Jeffrey, K.L. Chapter 12—Immune System Disorders and Epigenetics. In Medical Epigenetics; Tollefsbol, T.O., Ed.; Academic Press: Cambridge, MA, USA, 2016; pp. 199–219. ISBN 9780128032398. [Google Scholar] [CrossRef]
- Rangan, K.J.; Reck-Peterson, S.L. RNA recoding in cephalopods tailors microtubule motor protein function. Cell 2023, 186, 2531–2543.e11. [Google Scholar] [CrossRef] [PubMed]
- Norouzitallab, P.; Baruah, K.; Biswas, P.; Vanrompay, D.; Bossier, P. Probing the phenomenon of trained immunity in invertebrates during a transgenerational study, using brine shrimp Artemia as a model system. Sci. Rep. 2016, 6, 21166. [Google Scholar] [CrossRef]
- Rodriguez, J.D.; Katz, D.J. Lineage Tracing and Single-Cell RNA-seq in C. elegans to Analyze Transgenerational Epigenetic Phenotypes Inherited from Germ Cells. Methods Mol. Biol. 2023, 2677, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Barreiro, L.B. The epigenetically-encoded memory of the innate immune system. Curr. Opin. Immunol. 2020, 65, 7–13. [Google Scholar] [CrossRef]
- Sun, S.; Aguirre-Gamboa, R.; Barreiro, L.B. Transmission of stimulus-induced epigenetic changes through cell division is coupled to continuous transcription factor activity. Front. Immunol. 2023, 14, 1129577. [Google Scholar] [CrossRef] [PubMed]
- del Rosario, R.C.H.; Poschmann, J.; Lim, C.; Cheng, C.Y.; Kumar, P.; Riou, C.; Ong, S.T.; Gerges, S.; Hajan, H.S.; Kumar, D.; et al. Histone acetylome-wide associations in immune cells from individuals with active Mycobacterium tuberculosis infection. Nat. Microbiol. 2022, 7, 312–326. [Google Scholar] [CrossRef]
- Hu, Y.T.; Wu, T.C.; Yang, E.C.; Wu, P.C.; Lin, P.T.; Wu, Y.L. Regulation of genes related to immune signaling and detoxification in Apis mellifera by an inhibitor of histone deacetylation. Sci. Rep. 2017, 7, 41255. [Google Scholar] [CrossRef] [PubMed]
- Amarante, A.M.; da Silva, I.C.A.; Carneiro, V.C.; Vicentino, A.R.R.; Pinto, M.A.; Higa, L.M.; Moharana, K.C.; Talyuli, O.A.C.; Venancio, T.M.; de Oliveira, P.L.; et al. Zika virus infection drives epigenetic modulation of immunity by the histone acetyltransferase CBP of Aedes aegypti. PLoS Negl. Trop. Dis. 2022, 16, e0010559. [Google Scholar] [CrossRef]
- Dicke, M.; Cusumano, A.; Poelman, E.H. Microbial Symbionts of Parasitoids. Annu. Rev. Entomol. 2020, 65, 171–190. [Google Scholar] [CrossRef] [PubMed]
- Norouzitallab, P.; Baruah, K.; Vandegehuchte, M.; Van Stappen, G.; Catania, F.; Vanden Bussche, J.; Vanhaecke, L.; Sorgeloos, P.; Bossier, P. Environmental heat stress induces epigenetic transgenerational inheritance of robustness in parthenogenetic Artemia model. FASEB J. 2014, 28, 3552–3563. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, L.; Wang, Y.; Huang, G.; Li, X.; Xie, Z.; Zhou, Z. Insights into the Role of DNA Methylation in Immune Cell Development and Autoimmune Disease. Front. Cell Dev. Biol. 2021, 9, 757318. [Google Scholar] [CrossRef] [PubMed]
- Klughammer, J.; Romanovskaia, D.; Nemc, A.; Posautz, A.; Seid, C.A.; Schuster, L.C.; Keinath, M.C.; Ramos, J.S.L.; Kosack, L.; Evankow, A.; et al. Comparative analysis of genome-scale, base-resolution DNA methylation profiles across 580 animal species. Nat. Commun. 2023, 14, 232. [Google Scholar] [CrossRef]
- Belk, J.A.; Daniel, B.; Satpathy, A.T. Epigenetic regulation of T cell exhaustion. Nat. Immunol. 2022, 23, 848–860. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Wang, W.; Li, J.X.; Yuan, P.; Liu, Y.; Li, Y.; Wang, L.; Song, L. The DNA cytosine-5-methyltransferase 3 (DNMT3) involved in regulation of CgIL-17 expression in the immune response of oyster Crassostrea gigas. Dev. Comp. Immunol. 2021, 123, 104092. [Google Scholar] [CrossRef]
- Gu, Z.; Yang, J.; Yang, M.; Deng, Y.; Jiao, Y. Immunomodulatory effects of decitabine in pearl oyster Pinctada fucata martensii. Fish Shellfish Immunol. 2022, 129, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Xiao, Q.; Yu, F.; Gan, Y.; Lu, C.; Peng, W.; Zhang, Y.; Luo, X.; Chen, N.; You, W.; et al. Comparative transcriptome and DNA methylation analysis of phenotypic plasticity in the pacific abalone (Haliotis discus hannai). Front. Physiol. 2021, 12, 683499. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Yang, H.; Zhao, H.; Chen, M.; Wang, B. Transcriptional changes in epigenetic modifiers associated with gene silencing in the intestine of the sea cucumber, Apostichopus japonicus (Selenka), during aestivation. Chin. J. Oceanol. Limnol. 2011, 29, 1267–1274. [Google Scholar] [CrossRef]
- Huang, H.; Wu, P.; Zhang, S.; Shang, Q.; Yin, H.; Hou, Q.; Zhong, J.; Guo, X. DNA methylomes and transcriptomes analysis reveal implication of host DNA methylation machinery in BmNPV proliferation in Bombyx mori. BMC Genom. 2019, 20, 736. [Google Scholar] [CrossRef]
- Kausar, S.; Abbas, M.N.; Gul, I.; Liu, R.; Li, Q.; Zhao, E.; Lv, M.; Cui, H. Molecular Identification of Two DNA Methyltransferase Genes and Their Functional Characterization in the Anti-Bacterial Immunity of Antheraea pernyi. Front. Immunol. 2022, 13, 855888. [Google Scholar] [CrossRef]
- Baradaran, E.; Moharramipour, S.; Asgari, S.; Mehrabadi, M. Induction of DNA methyltransferase genes in Helicoverpa armigera following injection of pathogenic bacteria modulates expression of antimicrobial peptides and affects bacterial proliferation. J. Insect. Physiol. 2019, 118, 103939. [Google Scholar] [CrossRef]
- Heitmueller, M.; Billion, A.; Dobrindt, U.; Vilcinskas, A.; Mukherjee, K. Epigenetic Mechanisms Regulate Innate Immunity against Uropathogenic and Commensal-Like Escherichia coli in the Surrogate Insect Model Galleria mellonella. Infect. Immun. 2017, 85, e00336-17. [Google Scholar] [CrossRef] [PubMed]
- Kunert, N.; Marhold, J.; Stanke, J.; Stach, D.; Lyko, F. A Dnmt2-like protein mediates DNA methylation in Drosophila. Development 2003, 130, 5083–5090. [Google Scholar] [CrossRef]
- Cui, L.; Ma, R.; Cai, J.; Guo, C.; Chen, Z.; Yao, L.; Wang, Y.; Fan, R.; Wang, X.; Shi, Y. RNA modifications: Importance in immune cell biology and related diseases. Signal Transduct. Target Ther. 2022, 7, 334. [Google Scholar] [CrossRef]
- Eisenberg, E. Proteome Diversification by RNA Editing. Methods Mol. Biol. 2021, 2181, 229–251. [Google Scholar] [CrossRef]
- Gabay, O.; Shoshan, Y.; Kopel, E.; Ben-Zvi, U.; Mann, T.D.; Bressler, N.; Cohen-Fultheim, R.; Schaffer, A.A.; Roth, S.H.; Tzur, Z.; et al. Landscape of adenosine-to-inosine RNA recoding across human tissues. Nat. Commun. 2022, 13, 1184. [Google Scholar] [CrossRef]
- Alon, S.; Garrett, S.C.; Levanon, E.Y.; Olson, S.; Graveley, B.R.; Rosenthal, J.J.C.; Eisenberg, E. The majority of transcripts in the squid nervous system are extensively recoded by A-to-I RNA editing. eLife 2015, 4, e05198. [Google Scholar] [CrossRef] [PubMed]
- Shu, B.; Zhou, Y.X.; Li, H.; Zhang, R.Z.; He, C.; Yang, X. The METTL3/MALAT1/PTBP1/USP8/TAK1 axis promotes pyroptosis and M1 polarization of macrophages and contributes to liver fibrosis. Cell Death Discov. 2021, 7, 368. [Google Scholar] [CrossRef]
- Yu, R.; Li, Q.; Feng, Z.; Cai, L.; Xu, Q. m6A Reader YTHDF2 Regulates LPS-Induced Inflammatory Response. Int. J. Mol. Sci. 2019, 20, 1323. [Google Scholar] [CrossRef]
- Castro-Vargas, C.; Linares-López, C.; López-Torres, A.; Wrobel, K.; Torres-Guzmán, J.C.; Hernández, G.A.G.; Wrobel, K.; Lanz-Mendoza, H.; Contreras-Garduño, J. Methylation on RNA: A Potential Mechanism Related to Immune Priming within But Not across Generations. Front. Microbiol. 2017, 8, 473. [Google Scholar] [CrossRef]
- Pigeault, R.; Garnier, R.; Rivero, A.; Gandon, S. Evolution of transgenerational immunity in invertebrates. Proc. Biol. Sci. 2016, 283, 20161136. [Google Scholar] [CrossRef]
- Tetreau, G.; Dhinaut, J.; Gourbal, B.; Moret, Y. Trans-generational Immune Priming in Invertebrates: Current Knowledge and Future Prospects. Front. Immunol. 2019, 10, 1938. [Google Scholar] [CrossRef] [PubMed]
- Tetreau, G.; Dhinaut, J.; Galinier, R.; Audant-Lacour, P.; Voisin, S.N.; Arafah, K.; Chogne, M.; Hilliou, F.; Bordes, A.; Sabarly, C.; et al. Deciphering the molecular mechanisms of mother-to-egg immune protection in the mealworm beetle Tenebrio molitor. PLoS Pathog. 2020, 16, e1008935. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.D.; Luallen, R.J. An organismal understanding of C. elegans innate immune responses, from pathogen recognition to multigenerational resistance. Semin. Cell Dev. Biol. 2023, 154, 77–84. [Google Scholar] [CrossRef]
- Fraune, S.; Augustin, R.; Bosch, T.C. Embryo protection in contemporary immunology: Why bacteria matter. Commun. Integr. Biol. 2011, 4, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Marré, J.; Traver, E.C.; Jose, A.M. Extracellular RNA is transported from one generation to the next in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2016, 113, 12496–12501. [Google Scholar] [CrossRef] [PubMed]
- Kloc, M. Seahorse Male Pregnancy as a Model System to Study Pregnancy, Immune Adaptations, and Environmental Effects. Int. J. Mol. Sci. 2023, 24, 9712. [Google Scholar] [CrossRef]
- Burton, N.O.; Riccio, C.; Dallaire, A.; Price, J.; Jenkins, B.; Koulman, A.; Miska, E.A. Cysteine synthases CYSL-1 and CYSL-2 mediate C. elegans heritable adaptation to P. vranovensis infection. Nat. Commun. 2020, 11, 1741. [Google Scholar] [CrossRef]
- Kelly, B.; Pearce, E.L. Amino Assets: How Amino Acids Support Immunity. Cell Metab. 2020, 32, 154–175. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Shao, Z.; Powell-Coffman, J.A. The Caenorhabditis elegans rhy-1 gene inhibits HIF-1 hypoxia-inducible factor activity in a negative feedback loop that does not include vhl. Genetics 2006, 174, 1205–1214. [Google Scholar] [CrossRef] [PubMed]
- Ashe, A.; Sarkies, P.; Le Pen, J.; Tanguy, M.; Miska, E.A. Antiviral RNA Interference against Orsay Virus Is neither Systemic nor Transgenerational in Caenorhabditis elegans. J. Virol. 2015, 89, 12035–12046. [Google Scholar] [CrossRef]
- Sterken, M.G.; Snoek, L.B.; Bosman, K.J.; Daamen, J.; Riksen, J.A.; Bakker, J.; Pijlman, G.P.; Kammenga, J.E. A heritable antiviral RNAi response limits Orsay virus infection in Caenorhabditis elegans N2. PLoS ONE 2014, 9, e89760. [Google Scholar] [CrossRef]
- Willis, A.R.; Sukhdeo, R.; Reinke, A.W. Remembering your enemies: Mechanisms of within-generation and multigenerational immune priming in Caenorhabditis elegans. FEBS J. 2021, 288, 1759–1770. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jia, L.; Yi, H.; Guo, G.; Huang, L.; Zhang, Y.; Jiao, Z.; Wu, J. Pre-exposure to Candida albicans induce trans-generational immune priming and gene expression of Musca domestica. Front. Microbiol. 2022, 13, 902496. [Google Scholar] [CrossRef]
- Dishaw, L.J.; Litman, G.W. Invertebrate allorecognition: The origins of histocompatibility. Curr. Biol. 2009, 19, R286–R288. [Google Scholar] [CrossRef]
- Buss, L.W.; Anderson, C.; Westerman, E.; Kritzberger, C.; Poudyal, M.; Moreno, M.A.; Lakkis, F.G. Allorecognition triggers autophagy and subsequent necrosis in the cnidarian Hydractinia symbiolongicarpus. PLoS ONE 2012, 7, e48914. [Google Scholar] [CrossRef] [PubMed]
- Rinkevich, B. The colonial urochordate Botryllus schlosseri: From stem cells and natural tissue transplantation to issues in evolutionary ecology. Bioessays 2002, 24, 730–740. [Google Scholar] [CrossRef]
- Rinkevich, B. Germ Cell Parasitism as an Ecological and Evolutionary Puzzle: Hitchhiking with Positively Selected Genotypes. OIKOS 2002, 96, 25–30. Available online: https://www.jstor.org/stable/3547273 (accessed on 1 November 2023). [CrossRef]
- Nydam, M.L.; Netuschil, N.; Sanders, E.; Langenbacher, A.; Lewis, D.D.; Taketa, D.A.; Marimuthu, A.; Gracey, A.Y.; De Tomaso, A.W. The candidate histocompatibility locus of a Basal chordate encodes two highly polymorphic proteins. PLoS ONE 2013, 8, e65980. [Google Scholar] [CrossRef]
- McKitrick, T.R.; Muscat, C.C.; Pierce, J.D.; Bhattacharya, D.; De Tomaso, A.W. Allorecognition in a basal chordate consists of independent activating and inhibitory pathways. Immunity 2011, 34, 616–626. [Google Scholar] [CrossRef]
- Nydam, M.L.; De Tomaso, A.W. The fester locus in Botryllus schlosseri experiences selection. BMC Evol. Biol. 2012, 12, 249. [Google Scholar] [CrossRef]
- Nicotra, M.L.; Powell, A.E.; Rosengarten, R.D.; Moreno, M.; Grimwood, J.; Lakkis, F.G.; Dellaporta, S.L.; Buss, L.W. A hypervariable invertebrate allodeterminant. Curr. Biol. 2009, 19, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.F.; Powell, A.E.; Rosengarten, R.D.; Nicotra, M.L.; Moreno, M.A.; Grimwood, J.; Lakkis, F.G.; Dellaporta, S.L.; Buss, L.W. Hydractinia allodeterminant alr1 resides in an immunoglobulin superfamily-like gene complex. Curr. Biol. 2010, 20, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsov, S.G.; Bosch, T.C. Self/nonself recognition in Cnidaria: Contact to allogeneic tissue does not result in elimination of nonself cells in Hydra vulgaris. Zoology 2003, 106, 109–116. [Google Scholar] [CrossRef]
- Carpenter, M.A.; Powell, J.H.; Ishizuka, K.J.; Palmeri, K.J.; Rendulic, S.; De Tomaso, A.W. Growth and long-term somatic and germline chimerism following fusion of juvenile Botryllus schlosseri. Biol. Bull. 2011, 220, 57–70. [Google Scholar] [CrossRef]
- Stoner, D.S.; Weissman, I.L. Somatic and germ cell parasitism in a colonial ascidian: Possible role for a highly polymorphic allorecognition system. Proc. Natl. Acad. Sci. USA 1996, 93, 15254–15259. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.J.; LaMarre, J. Transposons and the PIWI pathway: Genome defense in gametes and embryos. Reproduction 2018, 156, R111–R124. [Google Scholar] [CrossRef] [PubMed]
- Cox, D.N.; Chao, A.; Baker, J.; Chang, L.; Qiao, D.; Lin, H. A novel class of evolutionarily conserved genes defined by piwi are essential for stem cell self-renewal. Genes Dev. 1998, 12, 3715–3727. [Google Scholar] [CrossRef] [PubMed]
- Le Thomas, A.; Rogers, A.K.; Webster, A.; Marinov, G.K.; Liao, S.E.; Perkins, E.M.; Hur, J.K.; Aravin, A.A.; Tóth, K.F. Piwi induces piRNA-guided transcriptional silencing and establishment of a repressive chromatin state. Genes Dev. 2013, 27, 390–399. [Google Scholar] [CrossRef]
- Lin, H.; Spradling, A.C. A novel group of pumilio mutations affects the asymmetric division of germline stem cells in the Drosophila ovary. Development 1997, 124, 2463–2476. [Google Scholar] [CrossRef]
- Siomi, M.C.; Sato, K.; Pezic, D.; Aravin, A.A. PIWI-interacting small RNAs: The vanguard of genome defense. Nat. Rev. Mol. Cell Biol. 2011, 12, 246–258. [Google Scholar] [CrossRef]
- Tóth, K.F.; Pezic, D.; Stuwe, E.; Webster, A. The piRNA Pathway Guards the Germline Genome Against Transposable Elements. Adv. Exp. Med. Biol. 2016, 886, 51–77. [Google Scholar] [CrossRef]
- Fridrich, A.; Moran, Y. Some flies do not play ping-pong. PLoS Biol. 2023, 21, e3002152. [Google Scholar] [CrossRef]
- Gainetdinov, I.; Vega-Badillo, J.; Cecchini, K.; Bagci, A.; Colpan, C.; De, D.; Bailey, S.; Arif, A.; Wu, P.H.; MacRae, I.J.; et al. Relaxed targeting rules help PIWI proteins silence transposons. Nature 2023, 619, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Hasuwa, H.; Siomi, H. Identification of Mouse piRNA Pathway Components Using Anti-MIWI2 Antibodies. Methods Mol. Biol. 2017, 1463, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Brennecke, J.; Aravin, A.A.; Stark, A.; Dus, M.; Kellis, M.; Sachidanandam, R.; Hannon, G.J. Discrete small RNA-generating loci as master regulators of transposon activity in Drosophila. Cell 2007, 128, 1089–1103. [Google Scholar] [CrossRef] [PubMed]
- Casier, K.; Autaa, J.; Gueguen, N.; Delmarre, V.; Marie, P.P.; Ronsseray, S.; Carré, C.; Brasset, E.; Teysset, L.; Boivin, A. The histone demethylase Kdm3 prevents auto-immune piRNAs production in Drosophila. Sci. Adv. 2023, 9, eade3872. [Google Scholar] [CrossRef]
- Czech, B.; Hannon, G.J. One Loop to Rule Them All: The Ping-Pong Cycle and piRNA-Guided Silencing. Trends Biochem. Sci. 2016, 41, 324–337. [Google Scholar] [CrossRef] [PubMed]
- Seth, M.; Shirayama, M.; Gu, W.; Ishidate, T.; Conte, D., Jr.; Mello, C.C. The C. elegans CSR-1 argonaute pathway counteracts epigenetic silencing to promote germline gene expression. Dev. Cell 2013, 27, 656–663. [Google Scholar] [CrossRef]
- Wedeles, C.J.; Wu, M.Z.; Claycomb, J.M. A multitasking Argonaute: Exploring the many facets of C. elegans CSR. Chromosome Res. 2013, 21, 573–586. [Google Scholar] [CrossRef]
- Halic, M.; Moazed, D. 22G-RNAs in transposon silencing and centromere function. Mol. Cell 2009, 36, 170–171. [Google Scholar] [CrossRef]
- Lloyd, A.C.; Gregory, K.S.; Isaac, R.E.; Acharya, K.R. A Molecular Analysis of the Aminopeptidase P-Related Domain of PID-5 from Caenorhabditis elegans. Biomolecules 2023, 13, 1132. [Google Scholar] [CrossRef]
- Youngman, E.M.; Claycomb, J.M. From early lessons to new frontiers: The worm as a treasure trove of small RNA biology. Front. Genet. 2014, 5, 416. [Google Scholar] [CrossRef]
- Bamezai, S.; Rawat, V.P.; Buske, C. Concise review: The Piwi-piRNA axis: Pivotal beyond transposon silencing. Stem Cells 2012, 30, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Juliano, C.; Wang, J.; Lin, H. Uniting germline and stem cells: The function of Piwi proteins and the piRNA pathway in diverse organisms. Annu. Rev. Genet. 2011, 45, 447–469. [Google Scholar] [CrossRef]
- Kashima, M.; Miyata, A.; Shibata, N. Planarian PIWI-piRNA Interaction Analysis Using Immunoprecipitation and piRNA Sequencing. Methods Mol. Biol. 2022, 2509, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Santos, D.; Feng, M.; Kolliopoulou, A.; Taning, C.N.T.; Sun, J.; Swevers, L. What Are the Functional Roles of Piwi Proteins and piRNAs in Insects? Insects 2023, 14, 187. [Google Scholar] [CrossRef] [PubMed]
- Ganeshan, K.; Chawla, A. Metabolic regulation of immune responses. Annu. Rev. Immunol. 2014, 32, 609–634. [Google Scholar] [CrossRef]
- Bahat, A.; MacVicar, T.; Langer, T. Metabolism and Innate Immunity Meet at the Mitochondria. Front. Cell Dev. Biol. 2021, 9, 720490. [Google Scholar] [CrossRef]
- Ferreira, A.V.; Domiguéz-Andrés, J.; Netea, M.G. The Role of Cell Metabolism in Innate Immune Memory. J. Innate Immun. 2022, 14, 42–50, Erratum in J. Innate Immun. 2021, 13, 194. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kloc, M.; Halasa, M.; Kubiak, J.Z.; Ghobrial, R.M. Invertebrate Immunity, Natural Transplantation Immunity, Somatic and Germ Cell Parasitism, and Transposon Defense. Int. J. Mol. Sci. 2024, 25, 1072. https://doi.org/10.3390/ijms25021072
Kloc M, Halasa M, Kubiak JZ, Ghobrial RM. Invertebrate Immunity, Natural Transplantation Immunity, Somatic and Germ Cell Parasitism, and Transposon Defense. International Journal of Molecular Sciences. 2024; 25(2):1072. https://doi.org/10.3390/ijms25021072
Chicago/Turabian StyleKloc, Malgorzata, Marta Halasa, Jacek Z. Kubiak, and Rafik M. Ghobrial. 2024. "Invertebrate Immunity, Natural Transplantation Immunity, Somatic and Germ Cell Parasitism, and Transposon Defense" International Journal of Molecular Sciences 25, no. 2: 1072. https://doi.org/10.3390/ijms25021072