
Scavenging of Cation Radicals of the Visual Cycle Retinoids by Lutein, Zeaxanthin, Taurine, and Melanin
 , , and
, , and
Abstract
:1. Introduction
2. Results
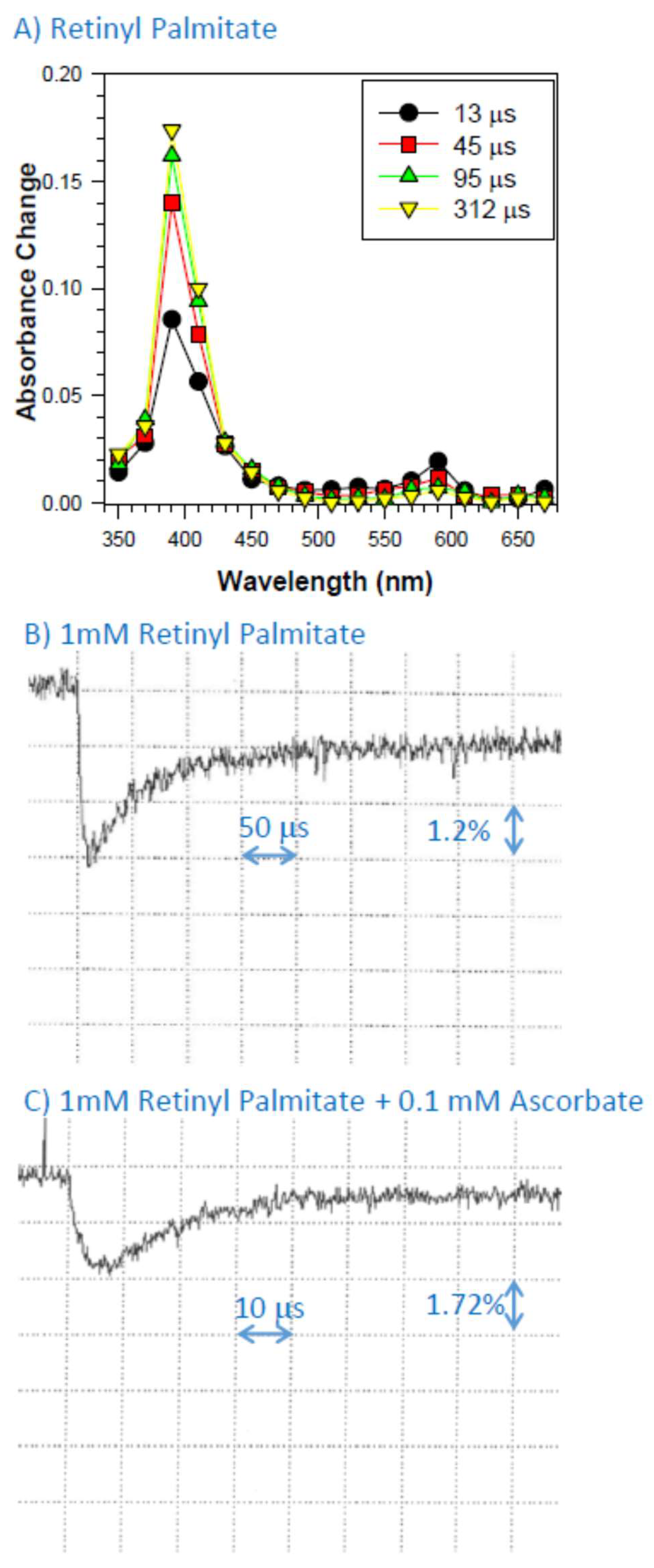
2.1. Formation of Cation Radical of Retinyl Palmitate in Benzene
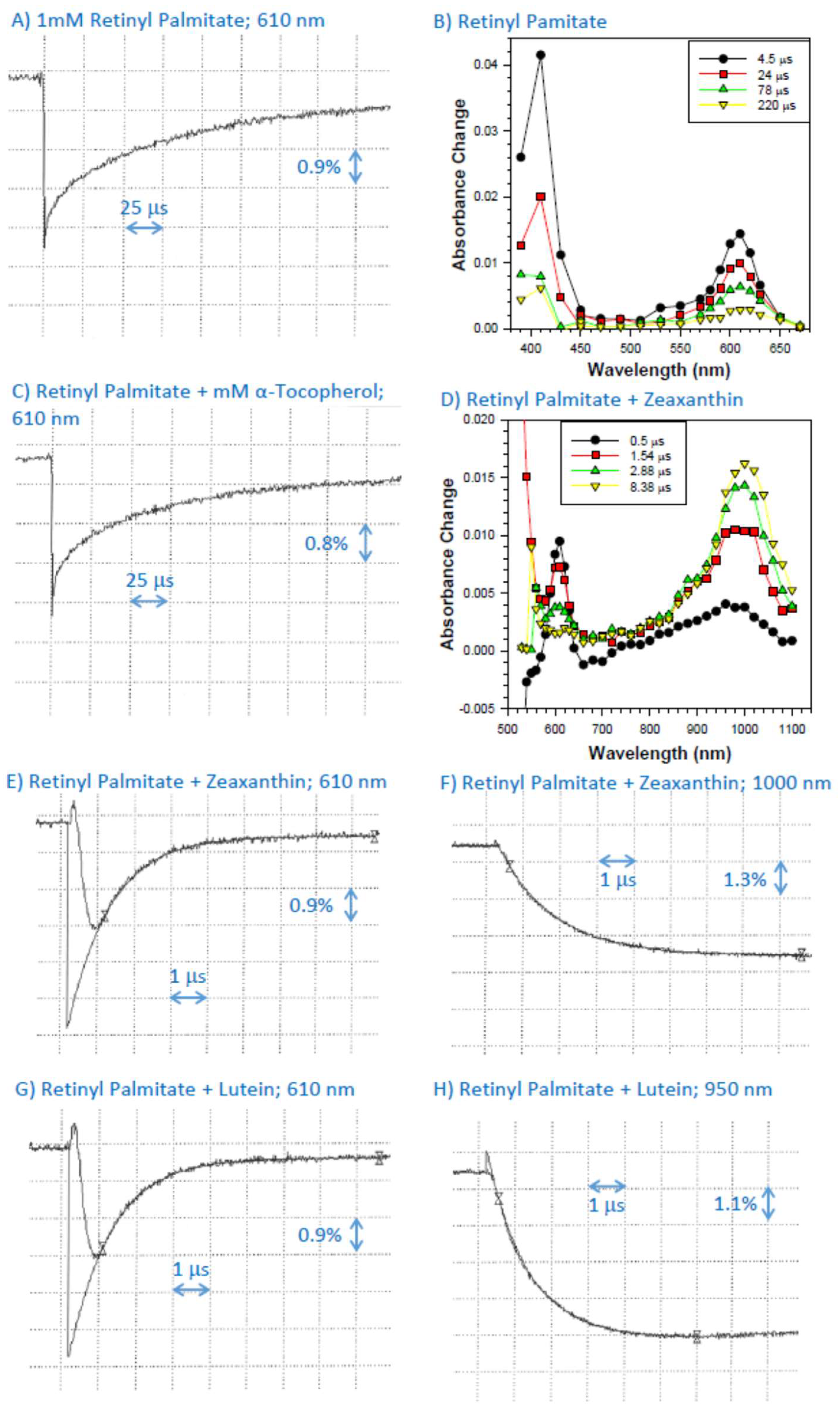
2.2. Interaction of Cation Radical of Retinyl Palmitate with α-Tocopherol, Lutein, and Zeaxanthin
2.3. Interaction of Retinyl Palmitate with Dibromine Radical Anions
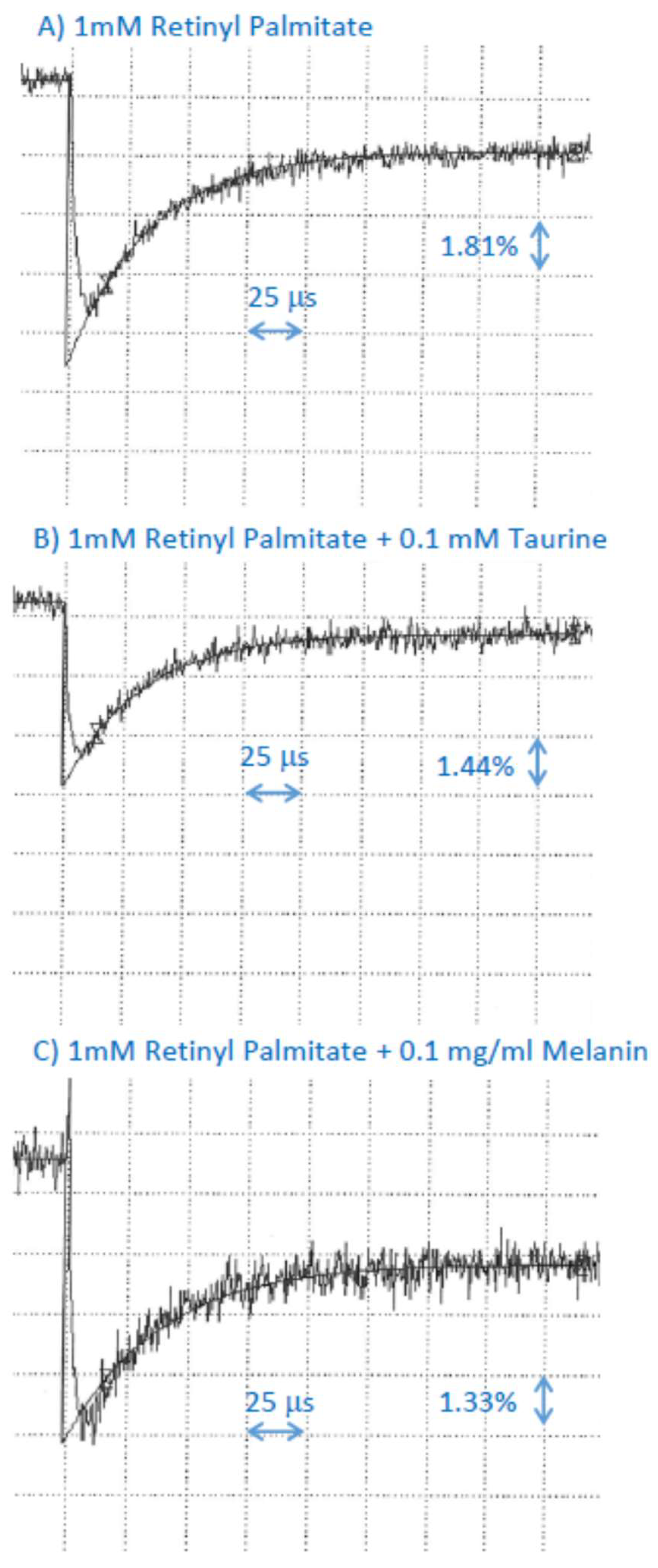
2.4. Interaction of Cation Radical of Retinyl Palmitate with Ascorbate, Taurine, and Dopa-Melanin
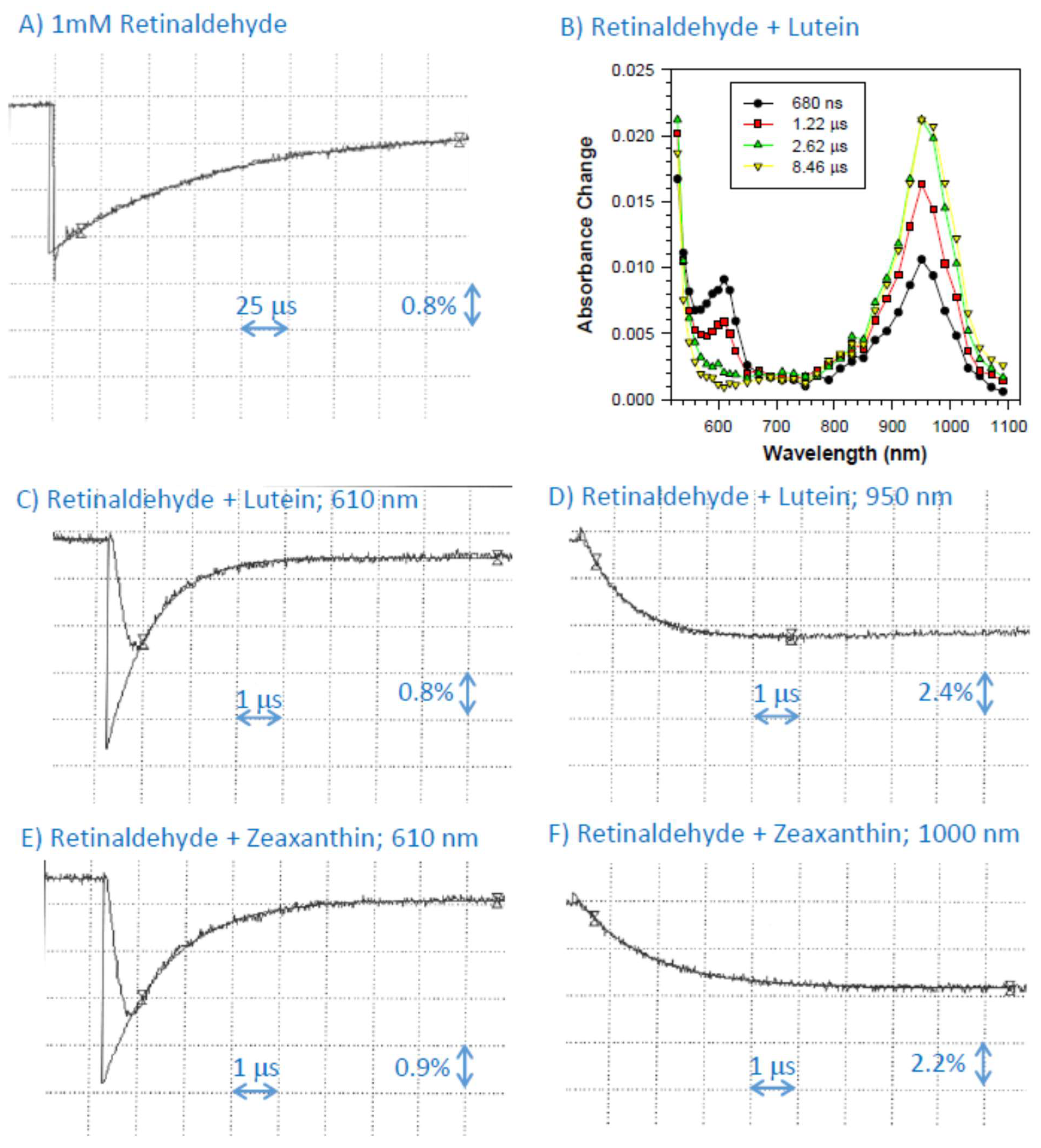
2.5. Interaction of Cation Radical of Retinaldehyde with Lutein, and Zeaxanthin
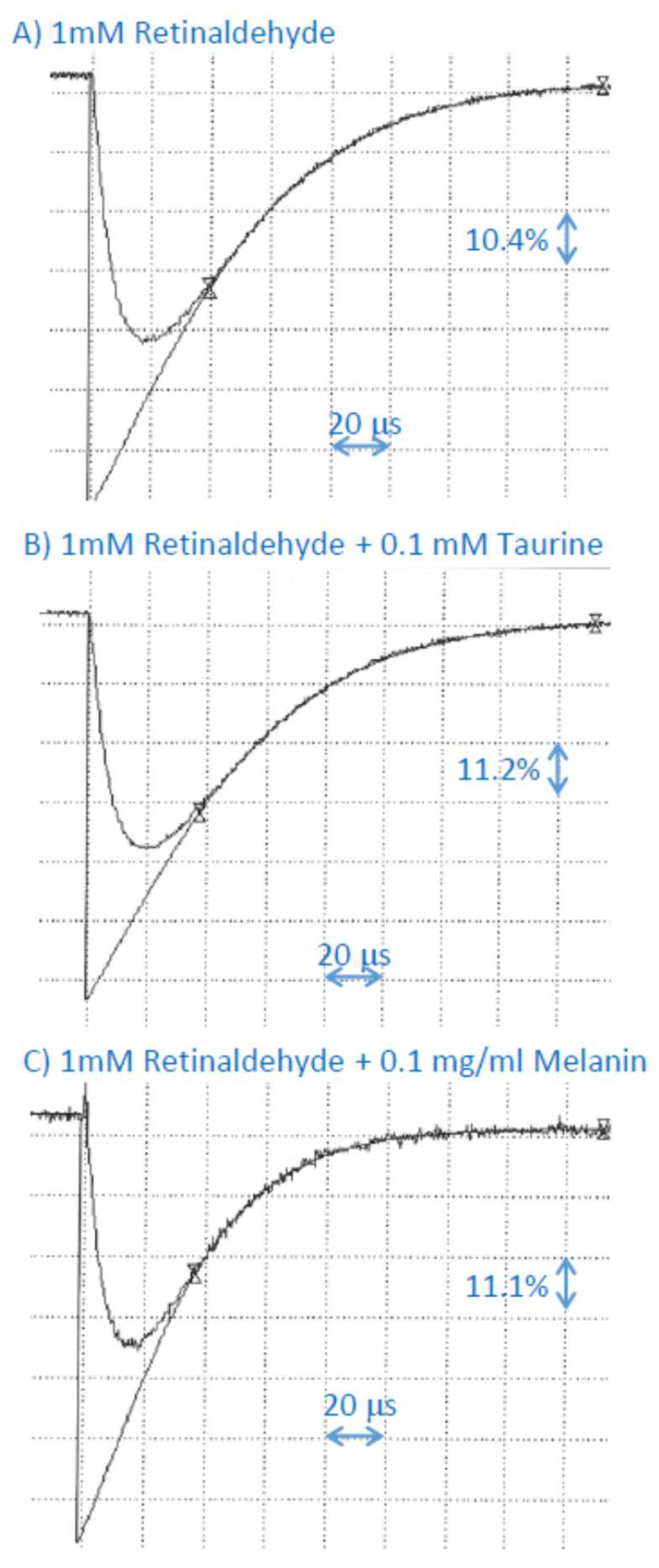
2.6. Interaction of Cation Radical of Retinaldehyde with Taurine, and Dopa-Melanin
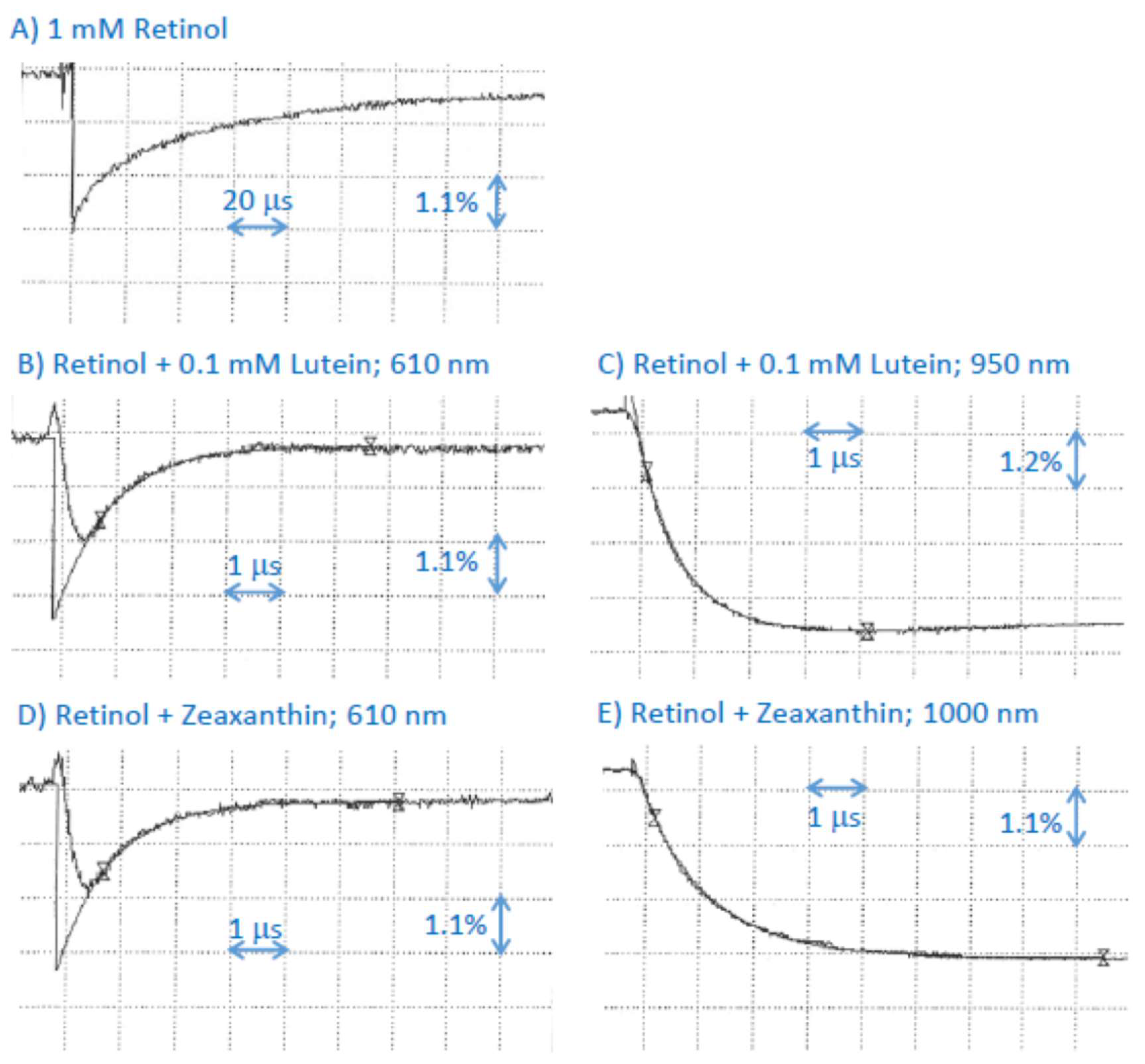
2.7. Interaction of Cation Radical of Retinol with Lutein, and Zeaxanthin
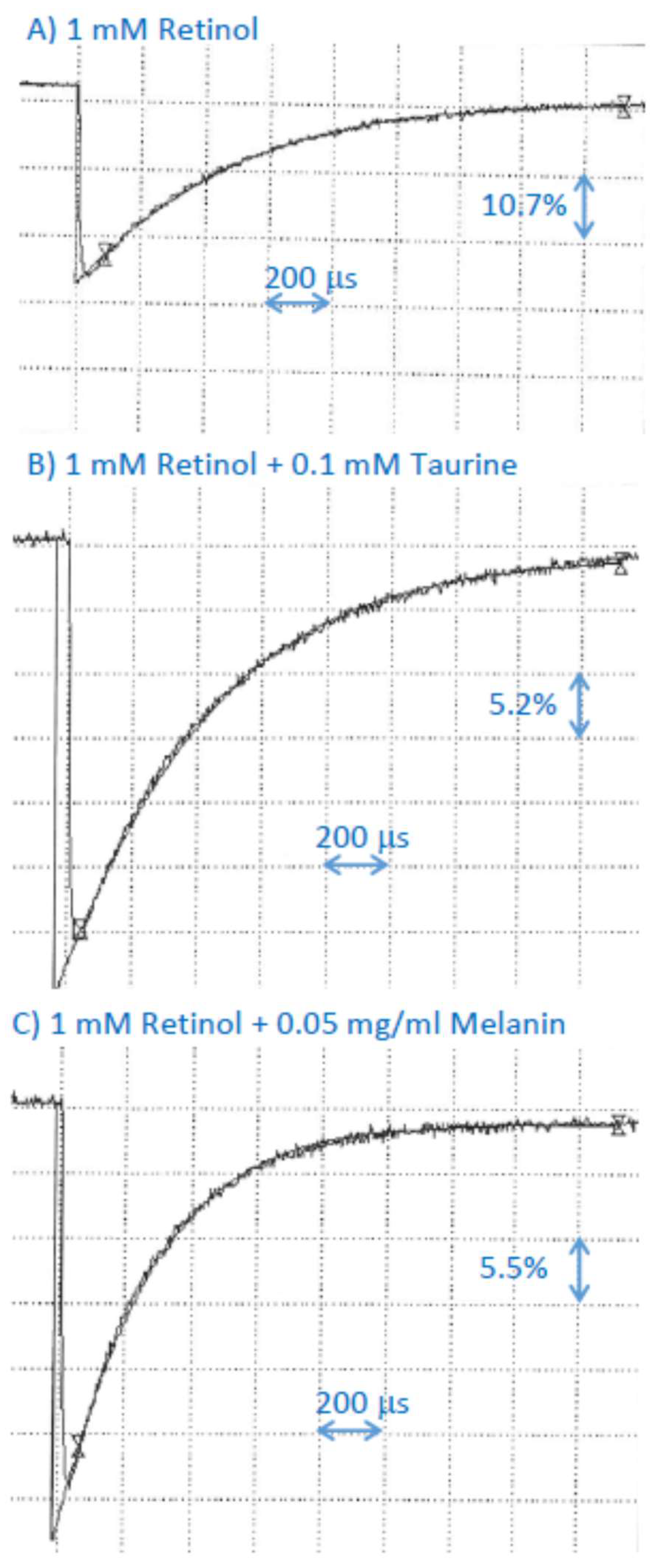
2.8. Interaction of Cation Radical of Retinol with Taurine, and Dopa-Melanin
3. Discussion
3.1. Scavenging of Retinyl Palmitate Cation Radicals by Retinal Antioxidants
3.2. Scavenging of Retinol Cation Radicals by Retinal Antioxidants
3.3. Scavenging of Retinaldehyde Cation Radicals by Retinal Antioxidants
3.4. Summary and Wider Physiological Relevance
4. Materials and Methods
4.1. Materials
4.2. Synthesis of Dopa-Melanin
4.3. Preparation of Triton X-100 Micelles with Retinoids
4.4. Generation of Retinoid Cation Radicals and Monitoring Their Interaction with Antioxidants
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiser, P.D.; Golczak, M.; Palczewski, K. Chemistry of the retinoid (visual) cycle. Chem. Rev. 2014, 114, 194–232. [Google Scholar] [CrossRef] [PubMed]
- Kiser, P.D.; Palczewski, K. Pathways and disease-causing alterations in visual chromophore production for vertebrate vision. J. Biol. Chem. 2021, 296, 100072. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Adler, L.t.; Goletz, P.; Gonzalez-Fernandez, F.; Thompson, D.A.; Koutalos, Y. Interphotoreceptor retinoid-binding protein removes all-trans-retinol and retinal from rod outer segments, preventing lipofuscin precursor formation. J. Biol. Chem. 2017, 292, 19356–19365. [Google Scholar] [CrossRef] [PubMed]
- Crouch, R.K.; Hazard, E.S.; Lind, T.; Wiggert, B.; Chader, G.; Corson, D.W. Interphotoreceptor retinoid-binding protein and alpha-tocopherol preserve the isomeric and oxidation state of retinol. Photochem. Photobiol. 1992, 56, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Hara, M.; Wu, W.; Malechka, V.V.; Takahashi, Y.; Ma, J.X.; Moiseyev, G. PNPLA2 mobilizes retinyl esters from retinosomes and promotes the generation of 11-cis-retinal in the visual cycle. Cell Rep. 2023, 42, 112091. [Google Scholar] [CrossRef] [PubMed]
- Tworak, A.; Kolesnikov, A.V.; Hong, J.D.; Choi, E.H.; Luu, J.C.; Palczewska, G.; Dong, Z.; Lewandowski, D.; Brooks, M.J.; Campello, L.; et al. Rapid RGR-dependent visual pigment recycling is mediated by the RPE and specialized Muller glia. Cell Rep. 2023, 42, 112982. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Choi, E.H.; Tworak, A.; Salom, D.; Leinonen, H.; Sander, C.L.; Hoang, T.V.; Handa, J.T.; Blackshaw, S.; Palczewska, G.; et al. Photic generation of 11-cis-retinal in bovine retinal pigment epithelium. J. Biol. Chem. 2019, 294, 19137–19154. [Google Scholar] [CrossRef]
- Morshedian, A.; Kaylor, J.J.; Ng, S.Y.; Tsan, A.; Frederiksen, R.; Xu, T.; Yuan, L.; Sampath, A.P.; Radu, R.A.; Fain, G.L.; et al. Light-Driven Regeneration of Cone Visual Pigments through a Mechanism Involving RGR Opsin in Muller Glial Cells. Neuron 2019, 102, 1172–1183.e5. [Google Scholar] [CrossRef]
- Rozanowska, M.; Rozanowski, B.; Boulton, M. Photobiology of the retina: Light damage to the retina. In Photobiological Sciences Online; Smith, K.C., Ed.; American Society for Photobiology: Herndon, VA, USA, 2009; Available online: http://www.photobiology.info (accessed on 22 January 2021).
- Różanowska, M.; Różanowski, B. Visual Transduction and Age-Related Changes in Lipofuscin. In Ophthalmology Research: The Visual Transduction Cascade; Tombran-Tink, J., Barnstable, C.J., Eds.; The Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 405–446. [Google Scholar]
- Hahn, P.; Milam, A.H.; Dunaief, J.L. Maculas affected by age-related macular degeneration contain increased chelatable iron in the retinal pigment epithelium and Bruch’s membrane. Arch. Ophthalmol. 2003, 121, 1099–1105. [Google Scholar] [CrossRef]
- Gu, X.; Meer, S.G.; Miyagi, M.; Rayborn, M.E.; Hollyfield, J.G.; Crabb, J.W.; Salomon, R.G. Carboxyethylpyrrole protein adducts and autoantibodies, biomarkers for age-related macular degeneration. J. Biol. Chem. 2003, 278, 42027–42035. [Google Scholar] [CrossRef]
- Crabb, J.W.; Miyagi, M.; Gu, X.; Shadrach, K.; West, K.A.; Sakaguchi, H.; Kamei, M.; Hasan, A.; Yan, L.; Rayborn, M.E.; et al. Drusen proteome analysis: An approach to the etiology of age-related macular degeneration. Proc. Natl. Acad. Sci. USA 2002, 99, 14682–14687. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef] [PubMed]
- Pennington, K.L.; DeAngelis, M.M. Epidemiology of age-related macular degeneration (AMD): Associations with cardiovascular disease phenotypes and lipid factors. Eye Vis. 2016, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Rein, D.B.; Wittenborn, J.S.; Burke-Conte, Z.; Gulia, R.; Robalik, T.; Ehrlich, J.R.; Lundeen, E.A.; Flaxman, A.D. Prevalence of Age-Related Macular Degeneration in the US in 2019. JAMA Ophthalmol. 2022, 140, 1202–1208. [Google Scholar] [CrossRef] [PubMed]
- Creuzot-Garcher, C.P.; Srour, M.; Baudin, F.; Daien, V.; Dot, C.; Nghiem-Buffet, S.; Girmens, J.F.; Coulombel, N.; Ponthieux, A.; Delcourt, C. Incidence and Prevalence of Neovascular Age-Related Macular Degeneration in France between 2008 and 2018: The LANDSCAPE Study. Ophthalmol. Sci. 2022, 2, 100114. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Cantrell, A.; Edge, R.; Land, E.J.; Sarna, T.; Truscott, T.G. Pulse radiolysis study of the interaction of retinoids with peroxyl radicals. Free Radic. Biol. Med. 2005, 39, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Belaidi, A.A.; Bush, A.I.; Ayton, S. Vitamin A metabolites inhibit ferroptosis. Biomed. Pharmacother. 2023, 164, 114930. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.K.; Land, E.J.; Truscott, T.G. Primary intermediates in the pulsed irradiation of retinoids. Photochem. Photobiol. 1982, 36, 139–145. [Google Scholar] [CrossRef]
- El-Agamey, A.; Fukuzumi, S. Laser flash photolysis study on the retinol radical cation in polar solvents. Org. Biomol. Chem. 2011, 9, 1046–6437. [Google Scholar] [CrossRef]
- El-Agamey, A.; Melo, T.B.; Sliwka, H.R. Exploring the reactivity of retinol radical cation toward organic and biological molecules: A laser flash photolysis study. J. Photochem. Photobiol. B 2017, 170, 33–39. [Google Scholar] [CrossRef]
- Artigas, J.M.; Felipe, A.; Navea, A.; Fandino, A.; Artigas, C. Spectral transmission of the human crystalline lens in adult and elderly persons: Color and total transmission of visible light. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4076–4084. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Handzel, K.; Boulton, M.E.; Rozanowski, B. Cytotoxicity of all-trans-retinal increases upon photodegradation. Photochem. Photobiol. 2012, 88, 1362–1372. [Google Scholar] [CrossRef] [PubMed]
- Cremers, F.P.M.; Lee, W.; Collin, R.W.J.; Allikmets, R. Clinical spectrum, genetic complexity and therapeutic approaches for retinal disease caused by ABCA4 mutations. Prog. Retin. Eye Res. 2020, 79, 100861. [Google Scholar] [CrossRef] [PubMed]
- Zampatti, S.; Peconi, C.; Calvino, G.; Ferese, R.; Gambardella, S.; Cascella, R.; Sebastiani, J.; Falsini, B.; Cusumano, A.; Giardina, E. A Splicing Variant in RDH8 Is Associated with Autosomal Recessive Stargardt Macular Dystrophy. Genes 2023, 14, 1659. [Google Scholar] [CrossRef] [PubMed]
- Molday, R.S.; Garces, F.A.; Scortecci, J.F.; Molday, L.L. Structure and function of ABCA4 and its role in the visual cycle and Stargardt macular degeneration. Prog. Retin. Eye Res. 2022, 89, 101036. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Edge, R.; Land, E.J.; Navaratnam, S.; Sarna, T.; Truscott, T.G. Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α, β, γ, and δ-Tocopherols. Int. J. Mol. Sci. 2019, 20, 2799. [Google Scholar] [CrossRef] [PubMed]
- Friedrichson, T.; Kalbach, H.L.; Buck, P.; van Kuijk, F.J. Vitamin E in macular and peripheral tissues of the human eye. Curr. Eye Res. 1995, 14, 693–701. [Google Scholar] [CrossRef]
- Ma, N.; Siegfried, C.; Kubota, M.; Huang, J.; Liu, Y.; Liu, M.; Dana, B.; Huang, A.; Beebe, D.; Yan, H.; et al. Expression Profiling of Ascorbic Acid-Related Transporters in Human and Mouse Eyes. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3440–3450. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study 2 Research, G.; Chew, E.Y.; Clemons, T.E.; Sangiovanni, J.P.; Danis, R.P.; Ferris, F.L., 3rd; Elman, M.J.; Antoszyk, A.N.; Ruby, A.J.; Orth, D.; et al. Secondary analyses of the effects of lutein/zeaxanthin on age-related macular degeneration progression: AREDS2 report No. 3. JAMA Ophthalmol. 2014, 132, 142–149. [Google Scholar] [CrossRef]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2017, 7, CD000254. [Google Scholar] [CrossRef]
- Chew, E.Y.; Clemons, T.E.; Agron, E.; Domalpally, A.; Keenan, T.D.L.; Vitale, S.; Weber, C.; Smith, D.C.; Christen, W.; Group, A.R. Long-term Outcomes of Adding Lutein/Zeaxanthin and omega-3 Fatty Acids to the AREDS Supplements on Age-Related Macular Degeneration Progression: AREDS2 Report 28. JAMA Ophthalmol. 2022, 140, 692–698. [Google Scholar] [CrossRef] [PubMed]
- Sommerburg, O.G.; Siems, W.G.; Hurst, J.S.; Lewis, J.W.; Kliger, D.S.; van Kuijk, F.J. Lutein and zeaxanthin are associated with photoreceptors in the human retina. Curr. Eye Res. 1999, 19, 491–495. [Google Scholar] [CrossRef] [PubMed]
- Rapp, L.M.; Maple, S.S.; Choi, J.H. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1200–1209. [Google Scholar]
- Rozanowska, M.B.; Czuba-Pelech, B.; Landrum, J.T.; Rozanowski, B. Comparison of Antioxidant Properties of Dehydrolutein with Lutein and Zeaxanthin, and their Effects on Cultured Retinal Pigment Epithelial Cells. Antioxidants 2021, 10, 753. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.B.; Czuba-Pelech, B.; Rozanowski, B. Is There an Optimal Combination of AREDS2 Antioxidants Zeaxanthin, Vitamin E and Vitamin C on Light-Induced Toxicity of Vitamin A Aldehyde to the Retina? Antioxidants 2022, 11, 1132. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, R.; Gorusupudi, A.; Bernstein, P.S. The macular carotenoids: A biochemical overview. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158617. [Google Scholar] [CrossRef]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef]
- Froger, N.; Moutsimilli, L.; Cadetti, L.; Jammoul, F.; Wang, Q.P.; Fan, Y.; Gaucher, D.; Rosolen, S.G.; Neveux, N.; Cynober, L.; et al. Taurine: The comeback of a neutraceutical in the prevention of retinal degenerations. Prog. Retin. Eye Res. 2014, 41, 44–63. [Google Scholar] [CrossRef]
- Garcia-Ayuso, D.; Di Pierdomenico, J.; Martinez-Vacas, A.; Vidal-Sanz, M.; Picaud, S.; Villegas-Perez, M.P. Taurine: A promising nutraceutic in the prevention of retinal degeneration. Neural Regen. Res. 2024, 19, 606–610. [Google Scholar] [CrossRef]
- Di Pierdomenico, J.; Martinez-Vacas, A.; Picaud, S.; Villegas-Perez, M.P.; Garcia-Ayuso, D. Taurine: An essential amino sulfonic acid for retinal health. Neural Regen. Res. 2023, 18, 807–808. [Google Scholar] [CrossRef]
- Różanowska, M. Properties and Functions of Ocular Melanins and Melanosomes. In Melanins and Melanosomes: Biosynthesis, Biogenesis, Physiological, and Pathological Functions; Riley, P.A., Borovansky, J., Eds.; John Wiley and Sons Inc.: Hoboken, NJ, USA, 2011; pp. 187–224. [Google Scholar]
- Rózanowska, M.B.; Pawlak, A.; Rózanowski, B. Products of docosahexaenoate oxidation as contributors to photosensitising properties of retinal lipofuscin. Int. J. Mol. Sci. 2021, 22, 3525. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.; Land, E.J.; McGarvey, D.J.; Truscott, T.G. Carotenoid triplet state lifetimes. J. Photochem. Photobiol. B 2000, 59, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.; Land, E.J.; McGarvey, D.; Mulroy, L.; Truscott, T.G. Relative one-electron reduction potentials of carotenoid radical cations and the interactions of carotenoids with the vitamin E radical cation. J. Am. Chem. Soc. 1998, 120, 4087–4090. [Google Scholar] [CrossRef]
- Polyakov, N.E.; Focsan, A.L.; Gao, Y.L.; Kispert, L.D. The Endless World of Carotenoids-Structural, Chemical and Biological Aspects of Some Rare Carotenoids. Int. J. Mol. Sci. 2023, 24, 9885. [Google Scholar] [CrossRef] [PubMed]
- Nagra, M.; Gilmartin, B.; Thai, N.J.; Logan, N.S. Determination of retinal surface area. J. Anat. 2017, 231, 319–324. [Google Scholar] [CrossRef]
- Li, B.; George, E.W.; Rognon, G.T.; Gorusupudi, A.; Ranganathan, A.; Chang, F.Y.; Shi, L.; Frederick, J.M.; Bernstein, P.S. Imaging lutein and zeaxanthin in the human retina with confocal resonance Raman microscopy. Proc. Natl. Acad. Sci. USA 2020, 117, 12352–12358. [Google Scholar] [CrossRef]
- Bhosale, P.; Zhao, D.Y.; Bernstein, P.S. HPLC measurement of ocular carotenoid levels in human donor eyes in the lutein supplementation era. Investig. Ophthalmol. Vis. Sci. 2007, 48, 543–549. [Google Scholar] [CrossRef]
- Shyam, R.; Gorusupudi, A.; Nelson, K.; Horvath, M.P.; Bernstein, P.S. RPE65 has an additional function as the lutein to meso-zeaxanthin isomerase in the vertebrate eye. Proc. Natl. Acad. Sci. USA 2017, 114, 10882–10887. [Google Scholar] [CrossRef]
- Poliakov, E.; Parikh, T.; Ayele, M.; Kuo, S.; Chander, P.; Gentleman, S.; Redmond, T.M. Aromatic lipophilic spin traps effectively inhibit RPE65 isomerohydrolase activity. Biochemistry 2011, 50, 6739–6741. [Google Scholar] [CrossRef]
- Kiser, P.D.; Zhang, J.; Badiee, M.; Li, Q.; Shi, W.; Sui, X.; Golczak, M.; Tochtrop, G.P.; Palczewski, K. Catalytic mechanism of a retinoid isomerase essential for vertebrate vision. Nat. Chem. Biol. 2015, 11, 409–415. [Google Scholar] [CrossRef]
- Kiser, P.D. Retinal pigment epithelium 65 kDa protein (RPE65): An update. Prog. Retin. Eye Res. 2022, 88, 101013. [Google Scholar] [CrossRef] [PubMed]
- Stuetz, W.; Weber, D.; Dolle, M.E.; Jansen, E.; Grubeck-Loebenstein, B.; Fiegl, S.; Toussaint, O.; Bernhardt, J.; Gonos, E.S.; Franceschi, C.; et al. Plasma Carotenoids, Tocopherols, and Retinol in the Age-Stratified (35–74 Years) General Population: A Cross-Sectional Study in Six European Countries. Nutrients 2016, 8, 614. [Google Scholar] [CrossRef] [PubMed]
- Jungert, A.; Neuhauser-Berthold, M. Interrelation between Plasma Concentrations of Vitamins C and E along the Trajectory of Ageing in Consideration of Lifestyle and Body Composition: A Longitudinal Study over Two Decades. Nutrients 2020, 12, 2944. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.; Padayatty, S.J.; Espey, M.G. Vitamin C: A concentration-function approach yields pharmacology and therapeutic discoveries. Adv. Nutr. 2011, 2, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Tveden-Nyborg, P. The Pharmacokinetics of Vitamin C. Nutrients 2019, 11, 2412. [Google Scholar] [CrossRef] [PubMed]
- Rice, M.E.; Russo-Menna, I. Differential compartmentalization of brain ascorbate and glutathione between neurons and glia. Neuroscience 1998, 82, 1213–1223. [Google Scholar] [CrossRef]
- Vachali, P.P.; Besch, B.M.; Gonzalez-Fernandez, F.; Bernstein, P.S. Carotenoids as possible interphotoreceptor retinoid-binding protein (IRBP) ligands: A surface plasmon resonance (SPR) based study. Arch. Biochem. Biophys. 2013, 539, 181–186. [Google Scholar] [CrossRef]
- Szuts, E.Z.; Harosi, F.I. Solubility of retinoids in water. Arch. Biochem. Biophys. 1991, 287, 297–304. [Google Scholar] [CrossRef]
- Rozanowski, B.; Cuenco, J.; Davies, S.; Shamsi, F.A.; Zadlo, A.; Dayhaw-Barker, P.; Rozanowska, M.; Sarna, T.; Boulton, M.E. The phototoxicity of aged human retinal melanosomes. Photochem. Photobiol. 2008, 84, 650–657. [Google Scholar] [CrossRef]
- Romano, F.; Lamanna, F.; Boon, C.J.; Siligato, A.; Kalra, G.; Agarwal, A.; Medori, C.; Bertelli, M.; Pellegrini, M.; Invernizzi, A.; et al. Clinical, Genotypic and Imaging Characterization of the spectrum of ABCA4 Retinopathies. Ophthalmol. Retina 2023, in press. [Google Scholar] [CrossRef]
- Maeda, T.; Golczak, M.; Maeda, A. Retinal photodamage mediated by all-trans-retinal. Photochem. Photobiol. 2012, 88, 1309–1319. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Sarna, T. Light-induced damage to the retina: Role of rhodopsin chromophore revisited. Photochem. Photobiol. 2005, 81, 1305–1330. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.; Grindle, J.; Ansell, P.L.; Borwein, B. Convolution in human rods: An ageing process. Br. J. Ophthalmol. 1979, 63, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Bazan, H.E.; Bazan, N.G.; Feeney-Burns, L.; Berman, E.R. Lipids in human lipofuscin-enriched subcellular fractions of two age populations. Comparison with rod outer segments and neural retina. Investig. Ophthalmol. Vis. Sci. 1990, 31, 1433–1443. [Google Scholar]
- Edge, R.; Truscott, T.G. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids-A Review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef]
- Różanowska, M.; Różanowski, B. Uptake and Photoprotection in Cultured RPE Cells. In Carotenoids: Physical, Chemical, and Biological Functions and Properties; Landrum, J.T., Ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 309–364. [Google Scholar]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.; Land, E.J.; Rozanowska, M.; Sarna, T.; Truscott, T.G. Carotenoid radical-melanin interactions. J. Phys. Chem. B 2000, 104, 7193–7196. [Google Scholar] [CrossRef]
- Rozanowska, M.; Bakker, L.; Boulton, M.E.; Rozanowski, B. Concentration dependence of vitamin C in combinations with vitamin E and zeaxanthin on light-induced toxicity to retinal pigment epithelial cells. Photochem. Photobiol. 2012, 88, 1408–1417. [Google Scholar] [CrossRef]
- Wrona, M.; Rozanowska, M.; Sarna, T. Zeaxanthin in combination with ascorbic acid or alpha-tocopherol protects ARPE-19 cells against photosensitized peroxidation of lipids. Free Radic. Biol. Med. 2004, 36, 1094–1101. [Google Scholar] [CrossRef]
- Wrona, M.; Korytowski, W.; Rozanowska, M.; Sarna, T.; Truscott, T.G. Cooperation of antioxidants in protection against photosensitized oxidation. Free Radic. Biol. Med. 2003, 35, 1319–1329. [Google Scholar] [CrossRef]
- Gao, Y.L.; Focsan, A.L.; Kispert, L.D. The effect of polarity of environment on the antioxidant activity of carotenoids. Chem. Phys. Lett. 2020, 761, 138098. [Google Scholar] [CrossRef]
- Arunkumar, R.; Gorusupudi, A.; Li, B.; Blount, J.D.; Nwagbo, U.; Kim, H.J.; Sparrow, J.R.; Bernstein, P.S. Lutein and zeaxanthin reduce A2E and iso-A2E levels and improve visual performance in Abca4−/−/Bco2−/− double knockout mice. Exp. Eye Res. 2021, 209, 108680. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, R.; Li, B.; Addo, E.K.; Hartnett, M.E.; Bernstein, P.S. Prenatal Carotenoid Supplementation With Lutein or Zeaxanthin Ameliorates Oxygen-Induced Retinopathy (OIR) in Bco2−/− Macular Pigment Mice. Investig. Ophthalmol. Vis. Sci. 2023, 64, 9. [Google Scholar] [CrossRef] [PubMed]
- Bandara, S.; Moon, J.; Ramkumar, S.; von Lintig, J. ASTER-B regulates mitochondrial carotenoid transport and homeostasis. J. Lipid Res. 2023, 64, 100369. [Google Scholar] [CrossRef] [PubMed]
- Bandara, S.; Ramkumar, S.; Imanishi, S.; Thomas, L.D.; Sawant, O.B.; Imanishi, Y.; von Lintig, J. Aster proteins mediate carotenoid transport in mammalian cells. Proc. Natl. Acad. Sci. USA 2022, 119, e2200068119. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; George, E.W.; Vachali, P.; Chang, F.Y.; Gorusupudi, A.; Arunkumar, R.; Giauque, N.A.; Wan, Z.; Frederick, J.M.; Bernstein, P.S. Mechanism for the selective uptake of macular carotenoids mediated by the HDL cholesterol receptor SR-BI. Exp. Eye Res. 2023, 229, 109429. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Nathans, J. ABCR, the ATP-binding cassette transporter responsible for Stargardt macular dystrophy, is an efficient target of all-trans-retinal-mediated photooxidative damage in vitro. Implications for retinal disease. J. Biol. Chem. 2001, 276, 11766–11774. [Google Scholar] [CrossRef]
- Weng, J.; Mata, N.L.; Azarian, S.M.; Tzekov, R.T.; Birch, D.G.; Travis, G.H. Insights into the function of Rim protein in photoreceptors and etiology of Stargardt’s disease from the phenotype in abcr knockout mice. Cell 1999, 98, 13–23. [Google Scholar] [CrossRef]
- Maeda, A.; Maeda, T.; Golczak, M.; Palczewski, K. Retinopathy in mice induced by disrupted all-trans-retinal clearance. J. Biol. Chem. 2008, 283, 26684–26693. [Google Scholar] [CrossRef]
- Zhao, J.; Liao, Y.; Chen, J.; Dong, X.; Gao, Z.; Zhang, H.; Wu, X.; Liu, Z.; Wu, Y. Aberrant Buildup of All-Trans-Retinal Dimer, a Nonpyridinium Bisretinoid Lipofuscin Fluorophore, Contributes to the Degeneration of the Retinal Pigment Epithelium. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1063–1075. [Google Scholar] [CrossRef]
- Lenis, T.L.; Hu, J.; Ng, S.Y.; Jiang, Z.; Sarfare, S.; Lloyd, M.B.; Esposito, N.J.; Samuel, W.; Jaworski, C.; Bok, D.; et al. Expression of ABCA4 in the retinal pigment epithelium and its implications for Stargardt macular degeneration. Proc. Natl. Acad. Sci. USA 2018, 115, E11120–E11127. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Tschulakow, A.; Taubitz, T.; Illing, B.; Biesemeier, A.; Julien-Schraermeyer, S.; Radu, R.A.; Jiang, Z.; Schraermeyer, U. Fundus autofluorescence, spectral-domain optical coherence tomography, and histology correlations in a Stargardt disease mouse model. FASEB J. 2020, 34, 3693–3714. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.; Banerjee, K.; Lehmann, G.L.; Almeida, D.; Hajjar, K.A.; Benedicto, I.; Jiang, Z.; Radu, R.A.; Thompson, D.H.; Rodriguez-Boulan, E.; et al. Lipofuscin causes atypical necroptosis through lysosomal membrane permeabilization. Proc. Natl. Acad. Sci. USA 2021, 118, e2100122118. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Taubitz, T.; Tschulakow, A.V.; Heiduschka, P.; Szewczyk, G.; Burnet, M.; Peters, T.; Biesemeier, A.; Sarna, T.; Schraermeyer, U.; et al. Removal of RPE lipofuscin results in rescue from retinal degeneration in a mouse model of advanced Stargardt disease: Role of reactive oxygen species. Free. Radic. Biol. Med. 2022, 182, 132–149. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Maeda, T.; Golczak, M.; Imanishi, Y.; Leahy, P.; Kubota, R.; Palczewski, K. Effects of potent inhibitors of the retinoid cycle on visual function and photoreceptor protection from light damage in mice. Mol. Pharmacol. 2006, 70, 1220–1229. [Google Scholar] [CrossRef]
- Maeda, T.; Maeda, A.; Matosky, M.; Okano, K.; Roos, S.; Tang, J.; Palczewski, K. Evaluation of potential therapies for a mouse model of human age-related macular degeneration caused by delayed all-trans-retinal clearance. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4917–4925. [Google Scholar] [CrossRef]
- Maeda, A.; Maeda, T.; Golczak, M.; Chou, S.; Desai, A.; Hoppel, C.L.; Matsuyama, S.; Palczewski, K. Involvement of all-transretinal in acute light-induced retinopathy of mice. J. Biol. Chem. 2009, 284, 15173–15183. [Google Scholar] [CrossRef]
- Maeda, A.; Golczak, M.; Chen, Y.; Okano, K.; Kohno, H.; Shiose, S.; Ishikawa, K.; Harte, W.; Palczewska, G.; Maeda, T.; et al. Primary amines protect against retinal degeneration in mouse models of retinopathies. Nat. Chem. Biol. 2011, 8, 170–178. [Google Scholar] [CrossRef]
- Shiose, S.; Chen, Y.; Okano, K.; Roy, S.; Kohno, H.; Tang, J.; Pearlman, E.; Maeda, T.; Palczewski, K.; Maeda, A. Toll-like receptor 3 is required for development of retinopathy caused by impaired all-trans-retinal clearance in mice. J. Biol. Chem. 2011, 286, 15543–15555. [Google Scholar] [CrossRef]
- Chen, Y.; Okano, K.; Maeda, T.; Chauhan, V.; Golczak, M.; Maeda, A.; Palczewski, K. Mechanism of all-trans-retinal toxicity with implications for Stargardt disease and age-related macular degeneration. J. Biol. Chem. 2012, 287, 5059–5069. [Google Scholar] [CrossRef]
- Okano, K.; Maeda, A.; Chen, Y.; Chauhan, V.; Tang, J.; Palczewska, G.; Sakai, T.; Tsuneoka, H.; Palczewski, K.; Maeda, T. Retinal cone and rod photoreceptor cells exhibit differential susceptibility to light-induced damage. J. Neurochem. 2012, 121, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sawada, O.; Kohno, H.; Le, Y.Z.; Subauste, C.; Maeda, T.; Maeda, A. Autophagy protects the retina from light-induced degeneration. J. Biol. Chem. 2013, 288, 7506–7518. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Palczewska, G.; Mustafi, D.; Golczak, M.; Dong, Z.; Sawada, O.; Maeda, T.; Maeda, A.; Palczewski, K. Systems pharmacology identifies drug targets for Stargardt disease-associated retinal degeneration. J. Clin. Investig. 2013, 123, 5119–5134. [Google Scholar] [CrossRef] [PubMed]
- Kohno, H.; Chen, Y.; Kevany, B.M.; Pearlman, E.; Miyagi, M.; Maeda, T.; Palczewski, K.; Maeda, A. Photoreceptor proteins initiate microglial activation via Toll-like receptor 4 in retinal degeneration mediated by all-trans-retinal. J. Biol. Chem. 2013, 288, 15326–15341. [Google Scholar] [CrossRef] [PubMed]
- Maeda, A.; Palczewska, G.; Golczak, M.; Kohno, H.; Dong, Z.; Maeda, T.; Palczewski, K. Two-photon microscopy reveals early rod photoreceptor cell damage in light-exposed mutant mice. Proc. Natl. Acad. Sci. USA 2014, 111, E1428–E1437. [Google Scholar] [CrossRef] [PubMed]
- Sawada, O.; Perusek, L.; Kohno, H.; Howell, S.J.; Maeda, A.; Matsuyama, S.; Maeda, T. All-trans-retinal induces Bax activation via DNA damage to mediate retinal cell apoptosis. Exp. Eye Res. 2014, 123, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yu, G.; Luo, C.; Maeda, A.; Zhang, N.; Sun, D.; Zhou, Z.; Puntel, A.; Palczewski, K.; Lu, Z.R. Synthesis and evaluation of a nanoglobular dendrimer 5-aminosalicylic Acid conjugate with a hydrolyzable schiff base spacer for treating retinal degeneration. ACS Nano 2014, 8, 153–161. [Google Scholar] [CrossRef]
- Yu, G.; Wu, X.; Ayat, N.; Maeda, A.; Gao, S.Q.; Golczak, M.; Palczewski, K.; Lu, Z.R. Multifunctional PEG retinylamine conjugate provides prolonged protection against retinal degeneration in mice. Biomacromolecules 2014, 15, 4570–4578. [Google Scholar] [CrossRef]
- Puntel, A.; Maeda, A.; Golczak, M.; Gao, S.Q.; Yu, G.; Palczewski, K.; Lu, Z.R. Prolonged prevention of retinal degeneration with retinylamine loaded nanoparticles. Biomaterials 2015, 44, 103–110. [Google Scholar] [CrossRef]
- Parmar, T.; Parmar, V.M.; Arai, E.; Sahu, B.; Perusek, L.; Maeda, A. Acute Stress Responses Are Early Molecular Events of Retinal Degeneration in Abca4−/−Rdh8−/− Mice After Light Exposure. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3257–3267. [Google Scholar] [CrossRef]
- Schur, R.M.; Gao, S.; Yu, G.; Chen, Y.; Maeda, A.; Palczewski, K.; Lu, Z.R. New GABA modulators protect photoreceptor cells from light-induced degeneration in mouse models. FASEB J. 2018, 32, 3289–3300. [Google Scholar] [CrossRef] [PubMed]
- Parmar, T.; Parmar, V.M.; Perusek, L.; Georges, A.; Takahashi, M.; Crabb, J.W.; Maeda, A. Lipocalin 2 Plays an Important Role in Regulating Inflammation in Retinal Degeneration. J. Immunol. 2018, 200, 3128–3141. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, J.; Wang, Y.; Liu, Z.; Wu, Y. Ferroptosis drives photoreceptor degeneration in mice with defects in all-trans-retinal clearance. J. Biol. Chem. 2021, 296, 100187. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Gao, S.Q.; Dong, Z.; Sheng, L.; Sun, D.; Zhang, N.; Zhang, J.; Margeivicus, S.; Fu, P.; Golczak, M.; et al. Peptide Derivatives of Retinylamine Prevent Retinal Degeneration with Minimal Side Effects on Vision in Mice. Bioconjug. Chem. 2021, 32, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Liao, C.; He, D.; Chen, J.; Han, J.; Lu, J.; Qin, K.; Liang, W.; Wu, X.; Liu, Z.; et al. Gasdermin E mediates photoreceptor damage by all-trans-retinal in the mouse retina. J. Biol. Chem. 2022, 298, 101553. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Tao, L.; Cai, B.; Chen, X.; Wang, Y.; Li, S.; Liao, C.; Chen, Y.; Chen, J.; Liu, Z.; et al. eIF2alpha incites photoreceptor cell and retina damage by all-trans-retinal. J. Biol. Chem. 2023, 299, 104686. [Google Scholar] [CrossRef]
- Chen, C.; Yang, K.; He, D.; Yang, B.; Tao, L.; Chen, J.; Wu, Y. Induction of ferroptosis by HO-1 contributes to retinal degeneration in mice with defective clearance of all-trans-retinal. Free Radic. Biol. Med. 2023, 194, 245–254. [Google Scholar] [CrossRef]
- Maeda, A.; Maeda, T.; Imanishi, Y.; Sun, W.; Jastrzebska, B.; Hatala, D.A.; Winkens, H.J.; Hofmann, K.P.; Janssen, J.J.; Baehr, W.; et al. Retinol dehydrogenase (RDH12) protects photoreceptors from light-induced degeneration in mice. J. Biol. Chem. 2006, 281, 37697–37704. [Google Scholar] [CrossRef]
- Maeda, A.; Golczak, M.; Maeda, T.; Palczewski, K. Limited roles of Rdh8, Rdh12, and Abca4 in all-trans-retinal clearance in mouse retina. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5435–5443. [Google Scholar] [CrossRef]
- Zobor, D.; Bruhwiler, B.; Zrenner, E.; Weisschuh, N.; Kohl, S. Genetic and Clinical Profile of Retinopathies Due to Disease-Causing Variants in Leber Congenital Amaurosis (LCA)-Associated Genes in a Large German Cohort. Int. J. Mol. Sci. 2023, 24, 8915. [Google Scholar] [CrossRef]
- Sen, P.; Srikrupa, N.; Maitra, P.; Srilekha, S.; Porkodi, P.; Gnanasekaran, H.; Bhende, M.; Khetan, V.; Mathavan, S.; Bhende, P.; et al. Next-generation sequencing--based genetic testing and phenotype correlation in retinitis pigmentosa patients from India. Indian. J. Ophthalmol. 2023, 71, 2512–2520. [Google Scholar] [CrossRef] [PubMed]
- Schlottmann, P.G.; Luna, J.D.; Labat, N.; Yadarola, M.B.; Bainttein, S.; Esposito, E.; Ibanez, A.; Barbaro, E.I.; Alvarez Mendiara, A.; Picotti, C.P.; et al. Nationwide genetic analysis of more than 600 families with inherited eye diseases in Argentina. NPJ Genom. Med. 2023, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.Y.; Chung, M.Y.; Chen, S.J. Pseudocoloboma-like maculopathy with biallelic RDH12 missense mutations. J. Med. Genet. 2023, 60, 859–865. [Google Scholar] [CrossRef] [PubMed]
- Daich Varela, M.; Moya, R.; Azevedo Souza Amaral, R.; Schlottmann, P.G.; Alvarez Mendiara, A.; Francone, A.; Guazi Resende, R.; Capalbo, L.; Gelvez, N.; Lopez, G.; et al. Clinical and Genetic Characterization of RDH12-Retinal Dystrophy in a South American Cohort. Ophthalmol. Retina 2023, in press. [Google Scholar] [CrossRef]
- Hanneken, A.; Neikirk, T.; Johnson, J.; Kono, M. Biochemical Measurements of Free Opsin in Macular Degeneration Eyes: Examining the 11-CIS Retinal Deficiency Hypothesis of Delayed Dark Adaptation (An American Ophthalmological Society Thesis). Trans. Am. Ophthalmol. Soc. 2017, 115, T1. [Google Scholar]
- Obana, A.; Gohto, Y.; Asaoka, R.; Gellermann, W.; Bernstein, P.S. Lutein and Zeaxanthin Distribution in the Healthy Macula and Its Association with Various Demographic Factors Examined in Pseudophakic Eyes. Antioxidants 2021, 10, 1857. [Google Scholar] [CrossRef]
- Rozanowska, M.; Sarna, T.; Land, E.J.; Truscott, T.G. Free radical scavenging properties of melanin interaction of eu- and pheo-melanin models with reducing and oxidising radicals. Free Radic. Biol. Med. 1999, 26, 518–525. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bimolecular Rates of Scavenging of Retinoid Radical Cations (109 M−1s−1) | |||
|---|---|---|---|
| Retinyl Palmitate | Retinaldehyde | Retinol | |
| Lutein | 8.85 ± 0.25 | 11.5 ± 1.4 | 12.6 ± 0.4 |
| Zeaxanthin | 6.39 ± 0.02 | 6.5 ± 0.3 | 7.9 ± 0.3 5.76 ± 0.50 in methanol a 2.50 ± 0.20 in benzonitrile a |
| α-Tocopherol | 0.027 ± 0.003 | 8.0 ± 0.3 b | 0.080 ± 0.045 a |
| Ascorbate | 0.58 ± 0.10 | 0.73 c 0.53 ± 0.08 in methanol d | 0.12 c |
| Taurine | <0.02 | <0.01 | <0.002 |
| Dopa-melanin | <0.02 | 0.016 ± 0.008 | 0.0051 ± 0.0001 |
| Effectiveness of Scavenging of Retinyl Palmitate Radical Cations in the RPE Scavenging Rate Constant × Antioxidant Concentration (103 s−1) | ||||
|---|---|---|---|---|
| Lutein/Zeaxanthin | ||||
| Normal Intake | Increased Intake | Maximum | ||
| [Macular RPE lutein] (µM) | 33.8 | 97.9 | 172 | |
| [Macular RPE lutein]kQ | 299 | 866 | 1520 | |
| [Macular RPE zeaxanthin] (µM) | 22.5 | 65.3 | 133 | |
| [Macular RPE zeaxanthin]kQ | 144 | 417 | 848 | |
| [Peripheral RPE lutein] (µM) | 1.14 | 3.30 | 5.8 | |
| [Peripheral RPE lutein]kQ | 10.1 | 29.2 | 51.3 | |
| [Peripheral RPE zeaxanthin] (µM) | 0.32 | 0.92 | 1.87 | |
| [Peripheral RPE zeaxanthin]kQ | 2.0 | 5.9 | 12.0 | |
| Vitamin E (α-tocopherol) | ||||
| Average | Maximum | |||
| [RPE Vitamin E] (µM) | 115 | 230 | ||
| [RPE Vitamin E]kQ | 3.11 | 6.21 | ||
| Vitamin C (ascorbate) | ||||
| [Ascorbate] (mM) | 1 mM | 2 mM | 10 mM | |
| [Ascorbate]kQ | 580 | 1160 | 5800 | |
| Effectiveness of Scavenging of Retinol Radical Cations in the RPE and Neural Retina Scavenging Rate Constant × Antioxidant Concentration (103 s−1) | ||||
|---|---|---|---|---|
| Lutein/Zeaxanthin | ||||
| Normal Intake | Increased Intake | Maximum | ||
| [Macular RPE lutein] (µM) | 33.8 | 97.9 | 172 | |
| [Macular RPE lutein]kQ | 425 | 1233 | 2169 | |
| [Macular RPE zeaxanthin] (µM) | 22.5 | 65.3 | 133 | |
| [Macular RPE zeaxanthin]kQ | 178 | 515 | 1049 | |
| [Peripheral RPE lutein] (µM) | 1.14 | 3.30 | 5.8 | |
| [Peripheral RPE lutein]kQ | 14 | 42 | 73 | |
| [Peripheral RPE zeaxanthin] (µM) | 0.32 | 0.92 | 1.87 | |
| [Peripheral RPE zeaxanthin]kQ | 2.5 | 7.3 | 14.8 | |
| [Macular POS lutein] (µM) | 18.8 | 54.5 | 95.9 | |
| [Macular POS lutein]kQ | 237 | 687 | 1209 | |
| [Macular POS zeaxanthin] (µM) | 13.7 | 39.7 | 80.7 | |
| [Macular POS zeaxanthin]kQ | 108 | 314 | 638 | |
| [Peripheral POS lutein] (µM) | 0.63 | 1.84 | 3.23 | |
| [Peripheral POS lutein]kQ | 8.0 | 23.1 | 40.7 | |
| [Peripheral POS zeaxanthin] (µM) | 0.19 | 0.56 | 1.14 | |
| [Peripheral POS zeaxanthin]kQ | 1.5 | 4.4 | 9.0 | |
| Vitamin E (α-tocopherol) | ||||
| Average | Maximum | |||
| [RPE Vitamin E] (µM) | 115 | 230 | ||
| [RPE Vitamin E]kQ | 9.2 | 18.4 | ||
| [neural vitamin E; macula] (µM) | 46 | 78 | ||
| [neural vitamin E; macula]kQ | 3.7 | 6.3 | ||
| [neural vitamin E; periphery] (µM) | 77 | 124 | ||
| [neural vitamin E; periphery]kQ | 6.1 | 9.9 | ||
| Vitamin C (ascorbate) | ||||
| [Ascorbate] (mM) | 1 mM | 2 mM | 10 mM | |
| [Ascorbate]kQ | 120 | 240 | 1200 | |
| Melanin in RPE melanosomes | ||||
| [Young RPE melanin] (mM) | 398 | |||
| [Young RPE melanin]kQ | 2030 | |||
| [Old RPE melanin] (mM) | 310 | |||
| [Old RPE melanin]kQ | 1581 | |||
| Effectiveness of Scavenging of Retinaldehyde Radical Cations in the RPE and Neural Retina Scavenging Rate Constant × Antioxidant Concentration (103 s−1) | ||||
|---|---|---|---|---|
| Lutein/Zeaxanthin | ||||
| Normal Intake | Increased Intake | Maximum | ||
| [Macular RPE lutein] (µM) | 33.8 | 97.9 | 172 | |
| [Macular RPE lutein]kQ | 388 | 1126 | 1979 | |
| [Macular RPE zeaxanthin] (µM) | 22.5 | 65.3 | 133 | |
| [Macular RPE zeaxanthin]kQ | 146 | 424 | 863 | |
| [Peripheral RPE lutein] (µM) | 1.14 | 3.30 | 5.8 | |
| [Peripheral RPE lutein]kQ | 13 | 38 | 67 | |
| [Peripheral RPE zeaxanthin] (µM) | 0.32 | 0.92 | 1.87 | |
| [Peripheral RPE zeaxanthin]kQ | 2.1 | 6.0 | 12.2 | |
| [Macular POS lutein] (µM) | 18.8 | 54.5 | 95.9 | |
| [Macular POS lutein]kQ | 217 | 627 | 1103 | |
| [Macular POS zeaxanthin] (µM) | 13.7 | 39.7 | 80.7 | |
| [Macular POS zeaxanthin]kQ | 89 | 258 | 525 | |
| [Peripheral POS lutein] (µM) | 0.63 | 1.84 | 3.23 | |
| [Peripheral POS lutein]kQ | 7.3 | 21.1 | 37.1 | |
| [Peripheral POS zeaxanthin] (µM) | 0.19 | 0.56 | 1.14 | |
| [Peripheral POS zeaxanthin]kQ | 1.3 | 3.6 | 7.4 | |
| Vitamin E (α-tocopherol) | ||||
| Average | Maximum | |||
| [RPE Vitamin E] (µM) | 115 | 230 | ||
| [RPE Vitamin E]kQ | 920 | 1840 | ||
| [neural vitamin E; macula] (µM) | 46 | 78 | ||
| [neural vitamin E; macula]kQ | 367 | 627 | ||
| [neural vitamin E; periphery] (µM) | 77 | 124 | ||
| [neural vitamin E; periphery]kQ | 613 | 991 | ||
| Vitamin C (ascorbate) | ||||
| [Ascorbate] (mM) | 1 mM | 2 mM | 10 mM | |
| [Ascorbate]kQ | 730 | 1460 | 7300 | |
| Melanin in RPE melanosomes | ||||
| [Young RPE melanin] (mM) | 398 | |||
| [Young RPE melanin]kQ | 6368 | |||
| [Old RPE melanin] (mM) | 310 | |||
| [Old RPE melanin]kQ | 4960 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozanowska, M.; Edge, R.; Land, E.J.; Navaratnam, S.; Sarna, T.; Truscott, T.G. Scavenging of Cation Radicals of the Visual Cycle Retinoids by Lutein, Zeaxanthin, Taurine, and Melanin. Int. J. Mol. Sci. 2024, 25, 506. https://doi.org/10.3390/ijms25010506
Rozanowska M, Edge R, Land EJ, Navaratnam S, Sarna T, Truscott TG. Scavenging of Cation Radicals of the Visual Cycle Retinoids by Lutein, Zeaxanthin, Taurine, and Melanin. International Journal of Molecular Sciences. 2024; 25(1):506. https://doi.org/10.3390/ijms25010506
Chicago/Turabian StyleRozanowska, Malgorzata, Ruth Edge, Edward J. Land, Suppiah Navaratnam, Tadeusz Sarna, and T. George Truscott. 2024. "Scavenging of Cation Radicals of the Visual Cycle Retinoids by Lutein, Zeaxanthin, Taurine, and Melanin" International Journal of Molecular Sciences 25, no. 1: 506. https://doi.org/10.3390/ijms25010506