
Leptin-Mediated Induction of IL-6 Expression in Hofbauer Cells Contributes to Preeclampsia Pathogenesis
 , ,
, ,
Abstract
:1. Introduction
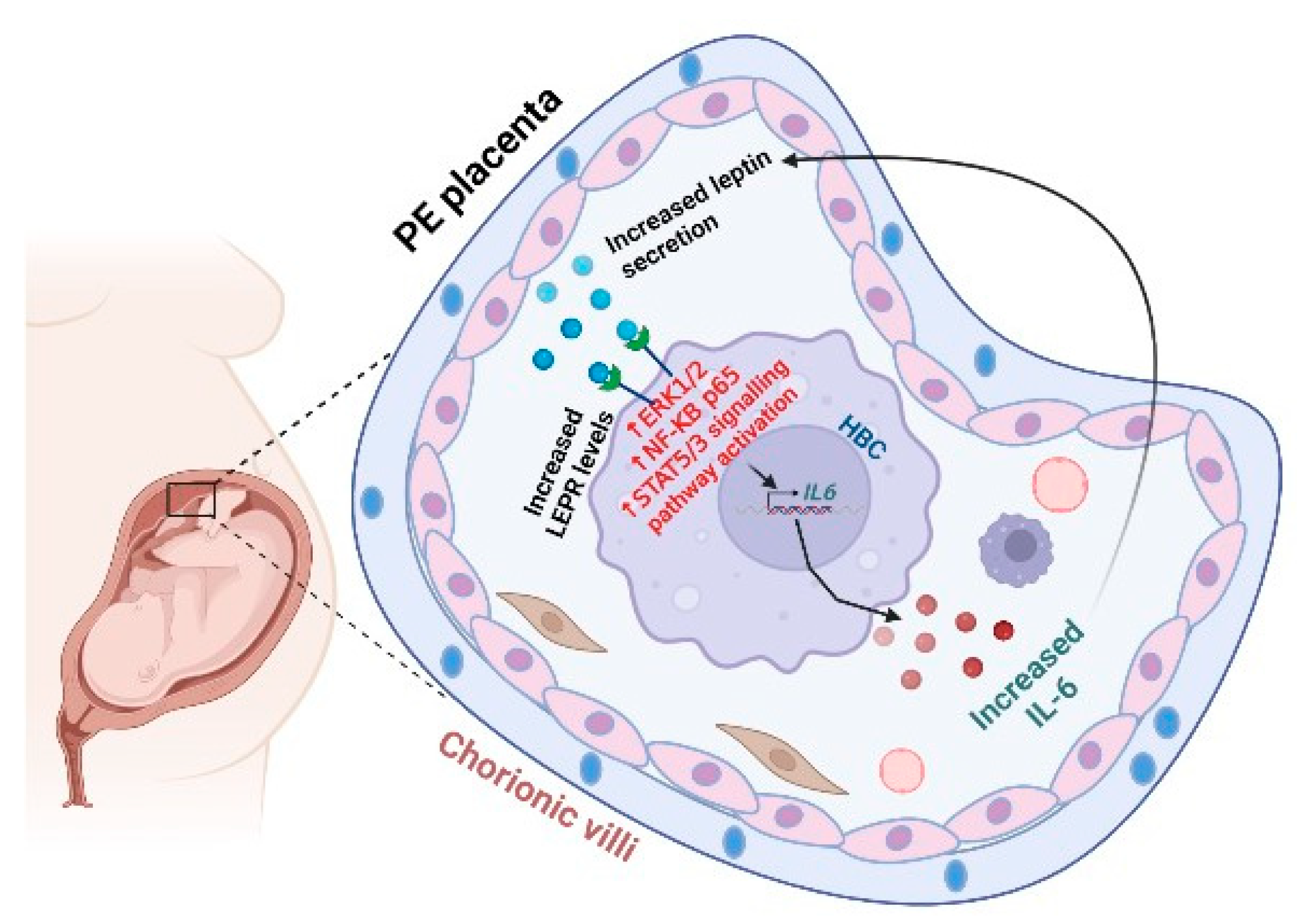
2. Results
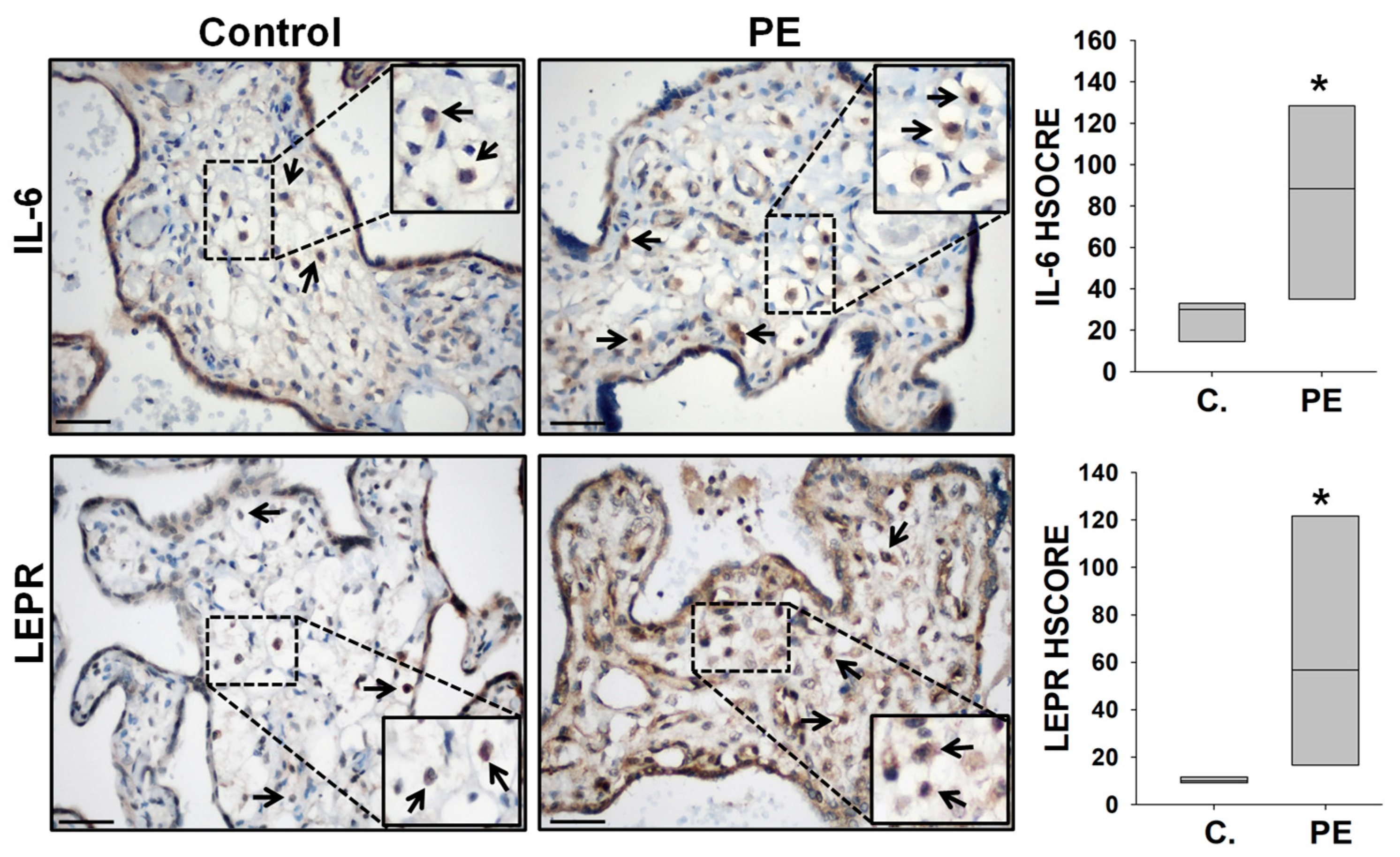
2.1. Elevated IL-6 and LEPR Immunoreactivity in Hofbauer Cells of PE Placentas
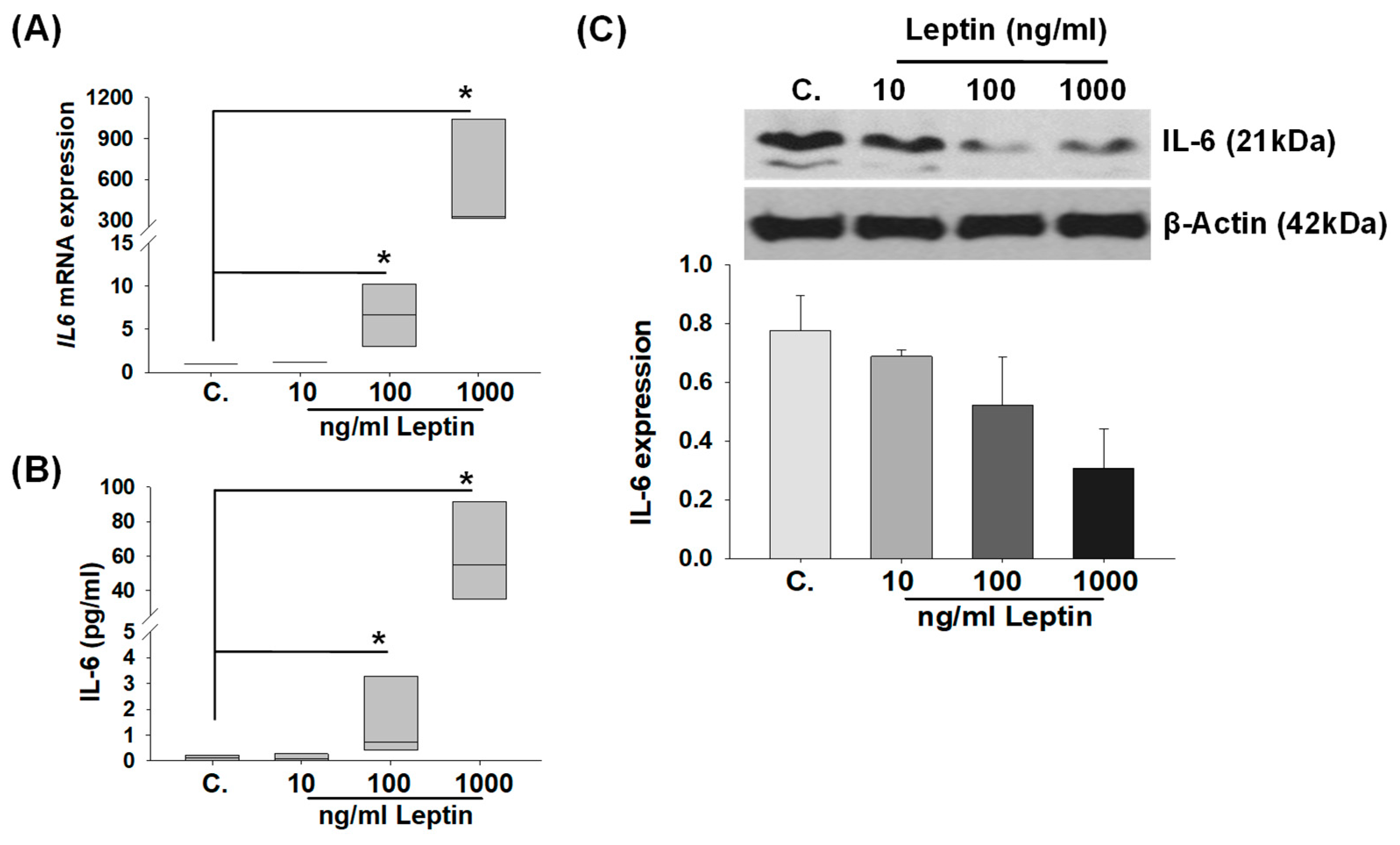
2.2. In Vitro Regulation of IL-6 Levels by Leptin Treatment in HBCs
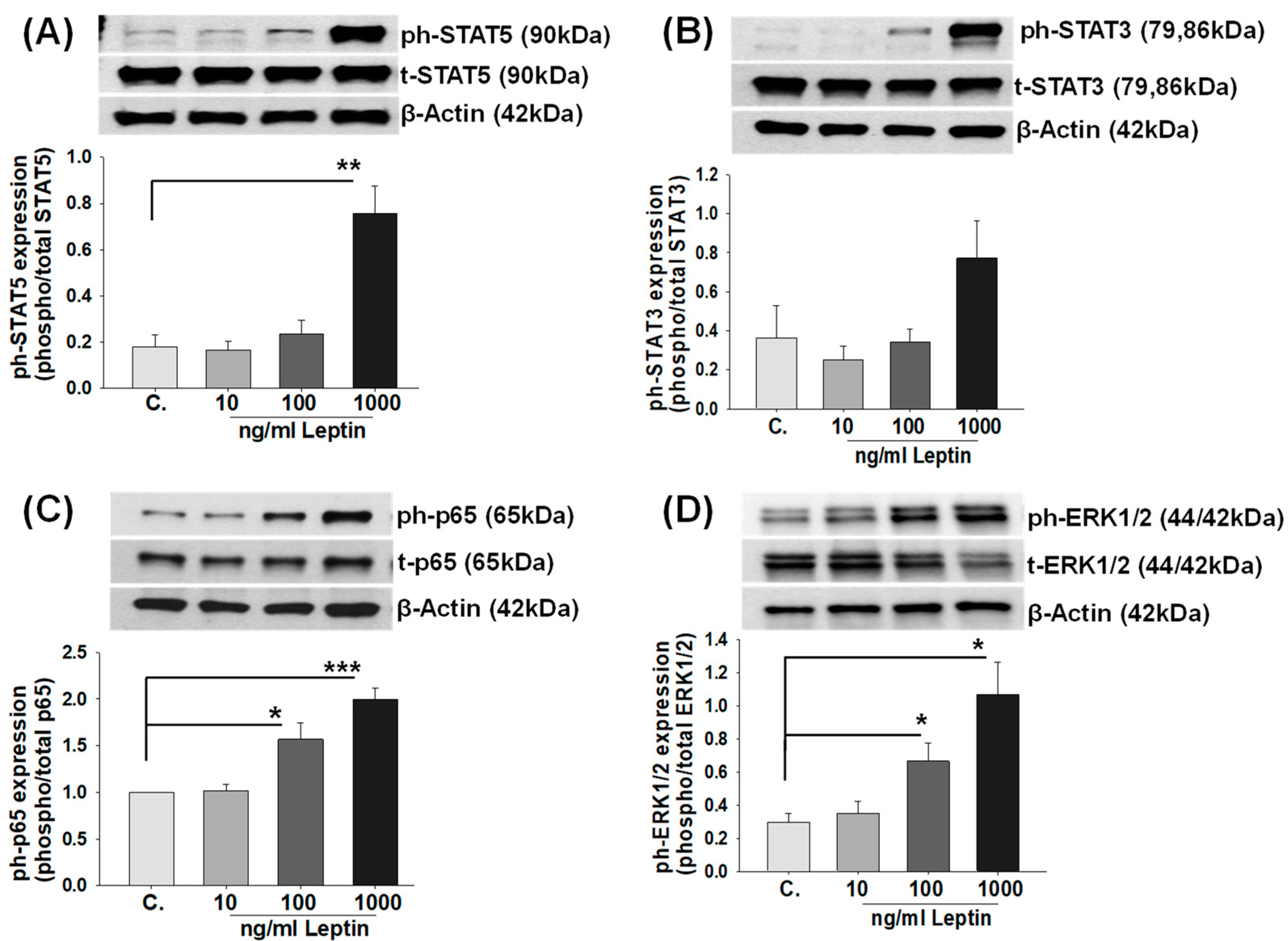
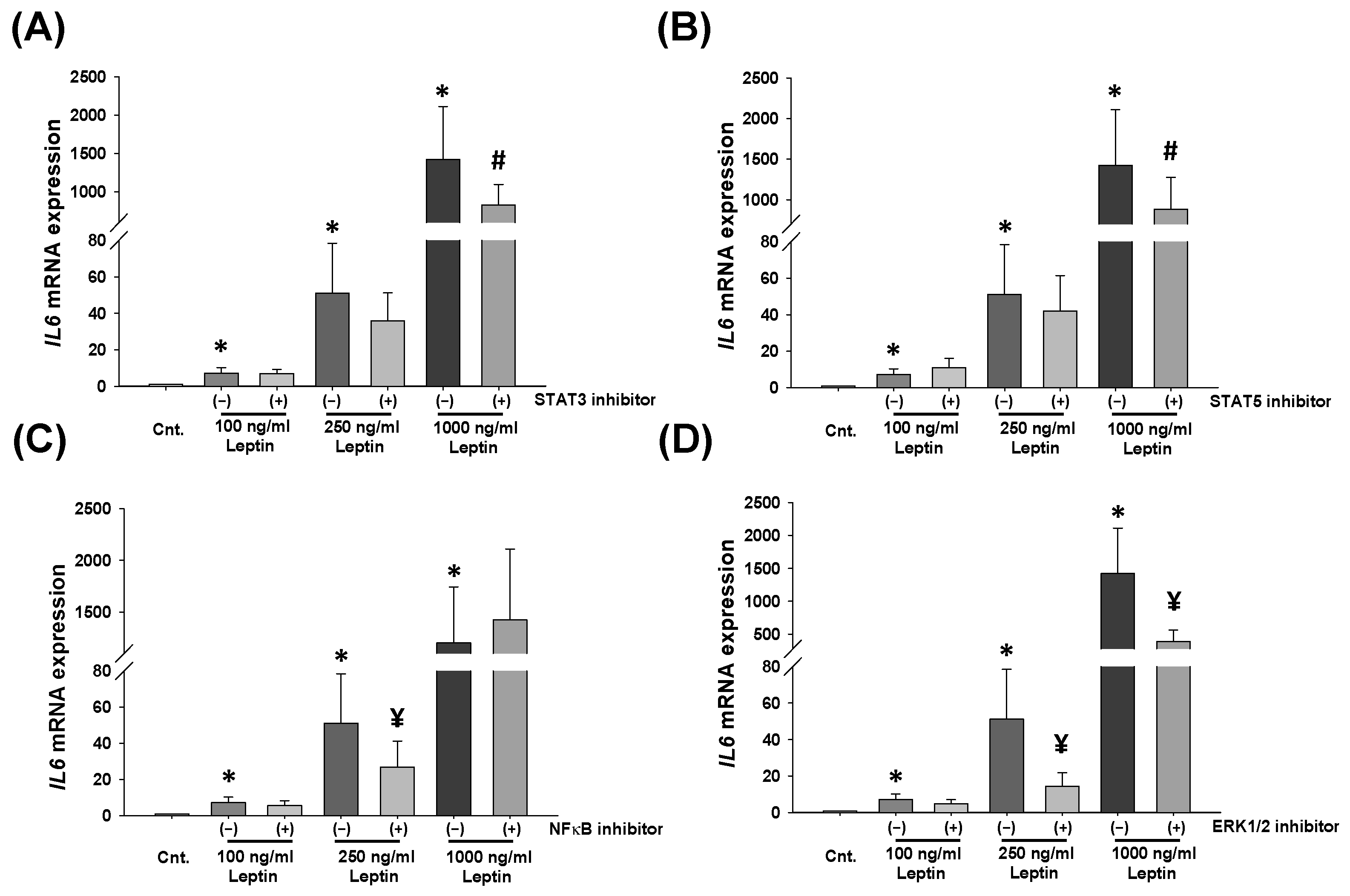
2.3. Leptin Activates STATs, NF-κB, and ERK1/2 Signaling Pathways in HBCs
3. Discussion
4. Materials and Methods
4.1. Placental Tissue Collection
4.2. Immunohistochemistry
4.3. Isolation and Culture of Hofbauer Cells
4.3.1. Experimental Setup
4.3.2. Western Blot Analysis
4.3.3. Enzyme-Linked Immunosorbent Assay (ELISA)
4.3.4. Quantitative Real-Time PCR
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perez-Perez, A.; Sanchez-Jimenez, F.; Maymo, J.; Duenas, J.L.; Varone, C.; Sanchez-Margalet, V. Role of leptin in female reproduction. Clin. Chem. Lab. Med. 2015, 53, 15–28. [Google Scholar] [CrossRef] [PubMed]
- de Knegt, V.E.; Hedley, P.L.; Kanters, J.K.; Thagaard, I.N.; Krebs, L.; Christiansen, M.; Lausten-Thomsen, U. The Role of Leptin in Fetal Growth during Pre-Eclampsia. Int. J. Mol. Sci. 2021, 22, 4569. [Google Scholar] [CrossRef] [PubMed]
- Fruhbeck, G. Intracellular signalling pathways activated by leptin. Biochem. J. 2006, 393 Pt 1, 7–20. [Google Scholar] [CrossRef] [PubMed]
- Barrichon, M.; Hadi, T.; Wendremaire, M.; Ptasinski, C.; Seigneuric, R.; Marcion, G.; Delignette, M.; Marchet, J.; Dumas, M.; Sagot, P.; et al. Dose-dependent biphasic leptin-induced proliferation is caused by non-specific IL-6/NF-kappaB pathway activation in human myometrial cells. Br. J. Pharmacol. 2015, 172, 2974–2990. [Google Scholar] [CrossRef] [PubMed]
- Lappas, M.; Permezel, M.; Rice, G.E. Leptin and adiponectin stimulate the release of proinflammatory cytokines and prostaglandins from human placenta and maternal adipose tissue via nuclear factor-kappaB, peroxisomal proliferator-activated receptor-gamma and extracellularly regulated kinase 1/2. Endocrinology 2005, 146, 3334–3342. [Google Scholar]
- Tang, C.H.; Lu, D.Y.; Yang, R.S.; Tsai, H.Y.; Kao, M.C.; Fu, W.M.; Chen, Y.F. Leptin-induced IL-6 production is mediated by leptin receptor, insulin receptor substrate-1, phosphatidylinositol 3-kinase, Akt, NF-kappaB, and p300 pathway in microglia. J. Immunol. 2007, 179, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.H.; Liu, S.C.; Tsai, C.H.; Fong, Y.C.; Wang, S.J.; Chang, Y.S.; Tang, C.H. Leptin induces IL-6 expression through OBRl receptor signaling pathway in human synovial fibroblasts. PLoS ONE 2013, 8, e75551. [Google Scholar] [CrossRef]
- Al-Hussaniy, H.A.; Alburghaif, A.H.; Naji, M.A. Leptin hormone and its effectiveness in reproduction, metabolism, immunity, diabetes, hopes and ambitions. J. Med. Life 2021, 14, 600–605. [Google Scholar] [CrossRef]
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The Importance of Leptin to Reproduction. Endocrinology 2021, 162, bqaa204. [Google Scholar] [CrossRef]
- Maurya, R.; Bhattacharya, P.; Dey, R.; Nakhasi, H.L. Leptin Functions in Infectious Diseases. Front. Immunol. 2018, 9, 2741. [Google Scholar] [CrossRef]
- Munzberg, H.; Morrison, C.D. Structure, production and signaling of leptin. Metab. Clin. Exp. 2015, 64, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Masuzaki, H.; Ogawa, Y.; Sagawa, N.; Hosoda, K.; Matsumoto, T.; Mise, H.; Nishimura, H.; Yoshimasa, Y.; Tanaka, I.; Mori, T.; et al. Nonadipose tissue production of leptin: Leptin as a novel placenta-derived hormone in humans. Nat. Med. 1997, 3, 1029–1033. [Google Scholar] [CrossRef] [PubMed]
- Senaris, R.; Garcia-Caballero, T.; Casabiell, X.; Gallego, R.; Castro, R.; Considine, R.V.; Dieguez, C.; Casanueva, F.F. Synthesis of leptin in human placenta. Endocrinology 1997, 138, 4501–4504. [Google Scholar] [CrossRef] [PubMed]
- Anim-Nyame, N.; Sooranna, S.R.; Steer, P.J.; Johnson, M.R. Longitudinal analysis of maternal plasma leptin concentrations during normal pregnancy and pre-eclampsia. Hum. Reprod. 2000, 15, 2033–2036. [Google Scholar] [CrossRef] [PubMed]
- Hauguel-de Mouzon, S.; Lepercq, J.; Catalano, P. The known and unknown of leptin in pregnancy. Am. J. Obstet. Gynecol. 2006, 194, 1537–1545. [Google Scholar] [CrossRef]
- Schanton, M.; Maymo, J.L.; Perez-Perez, A.; Sanchez-Margalet, V.; Varone, C.L. Involvement of leptin in the molecular physiology of the placenta. Reproduction 2018, 155, R1–R12. [Google Scholar] [CrossRef]
- Challier, J.; Galtier, M.; Bintein, T.; Cortez, A.; Lepercq, J.; Hauguel-de Mouzon, S. Placental leptin receptor isoforms in normal and pathological pregnancies. Placenta 2003, 24, 92–99. [Google Scholar] [CrossRef]
- Castellucci, M.; De Matteis, R.; Meisser, A.; Cancello, R.; Monsurro, V.; Islami, D.; Sarzani, R.; Marzioni, D.; Cinti, S.; Bischof, P. Leptin modulates extracellular matrix molecules and metalloproteinases: Possible implications for trophoblast invasion. Mol. Hum. Reprod. 2000, 6, 951–958. [Google Scholar] [CrossRef]
- Henson, M.C.; Swan, K.F.; O’Neil, J.S. Expression of placental leptin and leptin receptor transcripts in early pregnancy and at term. Obstet. Gynecol. 1998, 92, 1020–1028. [Google Scholar]
- Perez-Perez, A.; Maymo, J.; Duenas, J.L.; Goberna, R.; Calvo, J.C.; Varone, C.; Sanchez-Margalet, V. Leptin prevents apoptosis of trophoblastic cells by activation of MAPK pathway. Arch. Biochem. Biophys. 2008, 477, 390–395. [Google Scholar] [CrossRef]
- Teppa, R.J.; Ness, R.B.; Crombleholme, W.R.; Roberts, J.M. Free leptin is increased in normal pregnancy and further increased in preeclampsia. Metab. Clin. Exp. 2000, 49, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Gao, J.; Qu, Y.; Wang, S.; Wang, X.; Liu, J. Serum levels of leptin, adiponectin and resistin in relation to clinical characteristics in normal pregnancy and preeclampsia. Clin. Chim. Acta Int. J. Clin. Chem. 2016, 458, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Kalinderis, M.; Papanikolaou, A.; Kalinderi, K.; Vyzantiadis, T.A.; Ioakimidou, A.; Tarlatzis, B.C. Serum levels of leptin and IP-10 in preeclampsia compared to controls. Arch. Gynecol. Obstet. 2015, 292, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Taylor, B.D.; Ness, R.B.; Olsen, J.; Hougaard, D.M.; Skogstrand, K.; Roberts, J.M.; Haggerty, C.L. Serum leptin measured in early pregnancy is higher in women with preeclampsia compared with normotensive pregnant women. Hypertension 2015, 65, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Sibai, B.; Dekker, G.; Kupferminc, M. Pre-eclampsia. Lancet 2005, 365, 785–799. [Google Scholar] [CrossRef] [PubMed]
- Redman, C.W.; Sargent, I.L. Pre-eclampsia, the placenta and the maternal systemic inflammatory response—A review. Placenta 2003, 24 (Suppl. A), S21–S27. [Google Scholar] [CrossRef]
- Pijnenborg, R.; Bland, J.M.; Robertson, W.B.; Brosens, I. Uteroplacental arterial changes related to interstitial trophoblast migration in early human pregnancy. Placenta 1983, 4, 397–413. [Google Scholar] [CrossRef]
- Zhou, Y.; Damsky, C.H.; Fisher, S.J. Preeclampsia is associated with failure of human cytotrophoblasts to mimic a vascular adhesion phenotype. One cause of defective endovascular invasion in this syndrome? J. Clin. Investig. 1997, 99, 2152–2164. [Google Scholar] [CrossRef]
- Soleymanlou, N.; Jurisica, I.; Nevo, O.; Ietta, F.; Zhang, X.; Zamudio, S.; Post, M.; Caniggia, I. Molecular evidence of placental hypoxia in preeclampsia. J. Clin. Endocrinol. Metab. 2005, 90, 4299–4308. [Google Scholar] [CrossRef]
- Benyo, D.F.; Smarason, A.; Redman, C.W.; Sims, C.; Conrad, K.P. Expression of inflammatory cytokines in placentas from women with preeclampsia. J. Clin. Endocrinol. Metab. 2001, 86, 2505–2512. [Google Scholar] [CrossRef]
- Conrad, K.P.; Miles, T.M.; Benyo, D.F. Circulating levels of immunoreactive cytokines in women with preeclampsia. Am. J. Reprod. Immunol. 1998, 40, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Vince, G.S.; Starkey, P.M.; Austgulen, R.; Kwiatkowski, D.; Redman, C.W. Interleukin-6, tumour necrosis factor and soluble tumour necrosis factor receptors in women with pre-eclampsia. Br. J. Obstet. Gynaecol. 1995, 102, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, Y.; Ruber, M.; Matthiesen, L.; Berg, G.; Nieminen, K.; Sharma, S.; Ernerudh, J.; Ekerfelt, C. Cytokine mapping of sera from women with preeclampsia and normal pregnancies. J. Reprod. Immunol. 2006, 70, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Takacs, P.; Green, K.L.; Nikaeo, A.; Kauma, S.W. Increased vascular endothelial cell production of interleukin-6 in severe preeclampsia. Am. J. Obstet. Gynecol. 2003, 188, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Mise, H.; Sagawa, N.; Matsumoto, T.; Yura, S.; Nanno, H.; Itoh, H.; Mori, T.; Masuzaki, H.; Hosoda, K.; Ogawa, Y.; et al. Augmented placental production of leptin in preeclampsia: Possible involvement of placental hypoxia. J. Clin. Endocrinol. Metab. 1998, 83, 3225–3229. [Google Scholar] [CrossRef]
- Cameo, P.; Bischof, P.; Calvo, J.C. Effect of leptin on progesterone, human chorionic gonadotropin, and interleukin-6 secretion by human term trophoblast cells in culture. Biol. Reprod. 2003, 68, 472–477. [Google Scholar] [CrossRef]
- Thomas, J.R.; Naidu, P.; Appios, A.; McGovern, N. The Ontogeny and Function of Placental Macrophages. Front. Immunol. 2021, 12, 771054. [Google Scholar] [CrossRef]
- Luizon, M.R.; Palei, A.C.; Sandrim, V.C.; Amaral, L.M.; Machado, J.S.; Lacchini, R.; Cavalli, R.C.; Duarte, G.; Tanus-Santos, J.E. Tissue inhibitor of matrix metalloproteinase-1 polymorphism, plasma TIMP-1 levels, and antihypertensive therapy responsiveness in hypertensive disorders of pregnancy. Pharmacogenomics J. 2014, 14, 535–541. [Google Scholar] [CrossRef]
- Plaks, V.; Rinkenberger, J.; Dai, J.; Flannery, M.; Sund, M.; Kanasaki, K.; Ni, W.; Kalluri, R.; Werb, Z. Matrix metalloproteinase-9 deficiency phenocopies features of preeclampsia and intrauterine growth restriction. Proc. Natl. Acad. Sci. USA 2013, 110, 11109–11114. [Google Scholar] [CrossRef]
- Khan, S.; Katabuchi, H.; Araki, M.; Nishimura, R.; Okamura, H. Human villous macrophage-conditioned media enhance human trophoblast growth and differentiation in vitro. Biol. Reprod. 2000, 62, 1075–1083. [Google Scholar] [CrossRef]
- Pavlov, O.; Pavlova, O.; Ailamazyan, E.; Selkov, S. Characterization of cytokine production by human term placenta macrophages in vitro. Am. J. Reprod. Immunol. 2008, 60, 556–567. [Google Scholar] [CrossRef] [PubMed]
- Young, O.M.; Tang, Z.; Niven-Fairchild, T.; Tadesse, S.; Krikun, G.; Norwitz, E.R.; Mor, G.; Abrahams, V.M.; Guller, S. Toll-like receptor-mediated responses by placental Hofbauer cells (HBCs): A potential pro-inflammatory role for fetal M2 macrophages. Am. J. Reprod. Immunol. 2015, 73, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Laivuori, H.; Gallaher, M.J.; Collura, L.; Crombleholme, W.R.; Markovic, N.; Rajakumar, A.; Hubel, C.A.; Roberts, J.M.; Powers, R.W. Relationships between maternal plasma leptin, placental leptin mRNA and protein in normal pregnancy, pre-eclampsia and intrauterine growth restriction without pre-eclampsia. Mol. Hum. Reprod. 2006, 12, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, H.; Pryor-Koishi, K.; Kato, T.; Kowa, H.; Kurahashi, H.; Udagawa, Y. Microarray analysis of differentially expressed fetal genes in placental tissue derived from early and late onset severe pre-eclampsia. Placenta 2007, 28, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed]
- Maymo, J.L.; Perez Perez, A.; Gambino, Y.; Calvo, J.C.; Sanchez-Margalet, V.; Varone, C.L. Review: Leptin gene expression in the placenta--regulation of a key hormone in trophoblast proliferation and survival. Placenta 2011, 32 (Suppl. S2), S146–S153. [Google Scholar] [CrossRef] [PubMed]
- Molvarec, A.; Szarka, A.; Walentin, S.; Beko, G.; Karadi, I.; Prohaszka, Z.; Rigo, J., Jr. Serum leptin levels in relation to circulating cytokines, chemokines, adhesion molecules and angiogenic factors in normal pregnancy and preeclampsia. Reprod. Biol. Endocrinol. 2011, 9, 124. [Google Scholar] [CrossRef] [PubMed]
- Sitras, V.; Paulssen, R.H.; Gronaas, H.; Leirvik, J.; Hanssen, T.A.; Vartun, A.; Acharya, G. Differential placental gene expression in severe preeclampsia. Placenta 2009, 30, 424–433. [Google Scholar] [CrossRef]
- Redman, C.W.; Sargent, I.L. Placental stress and pre-eclampsia: A revised view. Placenta 2009, 30 (Suppl. A), S38–S42. [Google Scholar] [CrossRef]
- Li, R.H.; Poon, S.C.; Yu, M.Y.; Wong, Y.F. Expression of placental leptin and leptin receptors in preeclampsia. Int. J. Gynecol. Pathol. 2004, 23, 378–385. [Google Scholar] [CrossRef]
- Tsai, P.J.; Davis, J.; Bryant-Greenwood, G. Systemic and placental leptin and its receptors in pregnancies associated with obesity. Reprod. Sci. 2015, 22, 189–197. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Yen, C.F.; Basar, M.; Kayisli, U.A.; Martel, M.; Buhimschi, I.; Buhimschi, C.; Huang, S.J.; Krikun, G.; Schatz, F. Preeclampsia-related inflammatory cytokines regulate interleukin-6 expression in human decidual cells. Am. J. Pathol. 2008, 172, 1571–1579. [Google Scholar] [CrossRef] [PubMed]
- Gray, G.; Scroggins, D.G.; Wilson, K.T.; Scroggins, S.M. Cellular Immunotherapy in Mice Prevents Maternal Hypertension and Restores Anti-Inflammatory Cytokine Balance in Maternal and Fetal Tissues. Int. J. Mol. Sci. 2023, 24, 13594. [Google Scholar] [CrossRef]
- Ma, Y.; Ye, Y.; Zhang, J.; Ruan, C.C.; Gao, P.J. Immune imbalance is associated with the development of preeclampsia. Medicine 2019, 98, e15080. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, R.; Jain, A.K.; Mittal, P.; Kohli, M.; Jawanjal, P.; Rath, G. Association of pro- and anti-inflammatory cytokines in preeclampsia. J. Clin. Lab. Anal. 2019, 33, e22834. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Romero, R.; Kim, M.R.; Kim, Y.M.; Friel, L.; Espinoza, J.; Kim, C.J. Involvement of Hofbauer cells and maternal T cells in villitis of unknown aetiology. Histopathology 2008, 52, 457–464. [Google Scholar] [CrossRef]
- Castellucci, M.; Zaccheo, D.; Pescetto, G. A three-dimensional study of the normal human placental villous core. I. The Hofbauer cells. Cell Tissue Res. 1980, 210, 235–247. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage plasticity and polarization in tissue repair and remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Joerink, M.; Rindsjo, E.; van Riel, B.; Alm, J.; Papadogiannakis, N. Placental macrophage (Hofbauer cell) polarization is independent of maternal allergen-sensitization and presence of chorioamnionitis. Placenta 2011, 32, 380–385. [Google Scholar] [CrossRef]
- Sisino, G.; Bouckenooghe, T.; Aurientis, S.; Fontaine, P.; Storme, L.; Vambergue, A. Diabetes during pregnancy influences Hofbauer cells, a subtype of placental macrophages, to acquire a pro-inflammatory phenotype. Biochim. Biophys. Acta 2013, 1832, 1959–1968. [Google Scholar] [CrossRef]
- Tamblyn, J.A.; Lissauer, D.M.; Powell, R.; Cox, P.; Kilby, M.D. The immunological basis of villitis of unknown etiology—Review. Placenta 2013, 34, 846–855. [Google Scholar] [CrossRef] [PubMed]
- Hendrix, P.; Tang, Z.; Silasi, M.; Racicot, K.E.; Mor, G.; Abrahams, V.M.; Guller, S. Herpesvirus-infected Hofbauer cells activate endothelial cells through an IL-1beta-dependent mechanism. Placenta 2020, 91, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Schliefsteiner, C.; Ibesich, S.; Wadsack, C. Placental Hofbauer Cell Polarization Resists Inflammatory Cues In Vitro. Int. J. Mol. Sci. 2020, 21, 736. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Buhimschi, I.A.; Buhimschi, C.S.; Tadesse, S.; Norwitz, E.; Niven-Fairchild, T.; Huang, S.T.; Guller, S. Decreased levels of folate receptor-beta and reduced numbers of fetal macrophages (Hofbauer cells) in placentas from pregnancies with severe pre-eclampsia. Am. J. Reprod. Immunol. 2013, 70, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, A.; Nwabuobi, C.; Guzeloglu-Kayisli, O.; Schatz, F.; Lockwood, C.J.; Kayisli, U.A. Comparison of Fetal and Maternal Macrophage Numbers in Normal and Preeclamptic Human Placentas. In 66th Annual Meeting of the Society for Reproductive Investigation (SRI); Taylor, H.S., Ed.; Springer: Berlin/Heidelberg, Germany; Paris, France, 2019; Volume 26, p. 182A. [Google Scholar]
- La Cava, A.; Alviggi, C.; Matarese, G. Unraveling the multiple roles of leptin in inflammation and autoimmunity. J. Mol. Med. 2004, 82, 4–11. [Google Scholar] [CrossRef] [PubMed]
- La Cava, A.; Matarese, G. The weight of leptin in immunity. Nat. Rev. Immunol. 2004, 4, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Meisser, A.; Cameo, P.; Islami, D.; Campana, A.; Bischof, P. Effects of interleukin-6 (IL-6) on cytotrophoblastic cells. Mol. Hum. Reprod. 1999, 5, 1055–1058. [Google Scholar] [CrossRef]
- Stefaniak, M.; Dmoch-Gajzlerska, E.; Mazurkiewicz, B.; Gajzlerska-Majewska, W. Maternal serum and cord blood leptin concentrations at delivery. PLoS ONE 2019, 14, e0224863. [Google Scholar] [CrossRef]
- Highman, T.J.; Friedman, J.E.; Huston, L.P.; Wong, W.W.; Catalano, P.M. Longitudinal changes in maternal serum leptin concentrations, body composition, and resting metabolic rate in pregnancy. Am. J. Obstet. Gynecol. 1998, 178, 1010–1015. [Google Scholar] [CrossRef]
- Jansson, N.; Greenwood, S.L.; Johansson, B.R.; Powell, T.L.; Jansson, T. Leptin stimulates the activity of the system A amino acid transporter in human placental villous fragments. J. Clin. Endocrinol. Metab. 2003, 88, 1205–1211. [Google Scholar] [CrossRef]
- Chardonnens, D.; Cameo, P.; Aubert, M.L.; Pralong, F.P.; Islami, D.; Campana, A.; Gaillard, R.C.; Bischof, P. Modulation of human cytotrophoblastic leptin secretion by interleukin-1alpha and 17beta-oestradiol and its effect on HCG secretion. Mol. Hum. Reprod. 1999, 5, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- von Versen-Hoynck, F.; Rajakumar, A.; Parrott, M.S.; Powers, R.W. Leptin affects system A amino acid transport activity in the human placenta: Evidence for STAT3 dependent mechanisms. Placenta 2009, 30, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Pineiro, V.; Casabiell, X.; Peino, R.; Lage, M.; Camina, J.P.; Menendez, C.; Baltar, J.; Dieguez, C.; Casanueva, F. Dihydrotestosterone, stanozolol, androstenedione and dehydroepiandrosterone sulphate inhibit leptin secretion in female but not in male samples of omental adipose tissue in vitro: Lack of effect of testosterone. J. Endocrinol. 1999, 160, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Halleux, C.M.; Servais, I.; Reul, B.A.; Detry, R.; Brichard, S.M. Multihormonal control of ob gene expression and leptin secretion from cultured human visceral adipose tissue: Increased responsiveness to glucocorticoids in obesity. J. Clin. Endocrinol. Metab. 1998, 83, 902–910. [Google Scholar] [CrossRef] [PubMed]
- Cauzac, M.; Czuba, D.; Girard, J.; Hauguel-de Mouzon, S. Transduction of leptin growth signals in placental cells is independent of JAK-STAT activation. Placenta 2003, 24, 378–384. [Google Scholar] [CrossRef] [PubMed]
- Ozmen, A.; Guzeloglu-Kayisli, O.; Tabak, S.; Guo, X.; Semerci, N.; Nwabuobi, C.; Larsen, K.; Wells, A.; Uyar, A.; Arlier, S.; et al. Preeclampsia is Associated with Reduced ISG15 Levels Impairing Extravillous Trophoblast Invasion. Front. Cell Dev. Biol. 2022, 10, 898088. [Google Scholar] [CrossRef] [PubMed]
- Canfield, J.; Arlier, S.; Mong, E.F.; Lockhart, J.; VanWye, J.; Guzeloglu-Kayisli, O.; Schatz, F.; Magness, R.R.; Lockwood, C.J.; Tsibris, J.C.M.; et al. Decreased LIN28B in preeclampsia impairs human trophoblast differentiation and migration. FASEB J. 2019, 33, 2759–2769. [Google Scholar] [CrossRef]
- ACOG practice bulletin. Diagnosis and management of preeclampsia and eclampsia. Number 33, January 2002. American College of Obstetricians and Gynecologists. Int. J. Gynaecol. Obstet. Off. Organ Int. Fed. Gynaecol. Obstet. 2002, 77, 67–75. [Google Scholar]
- ACOG. ACOG Practice Bulletin No. 202 Summary: Gestational Hypertension and Preeclampsia. Obstet. Gynecol. 2019, 133, 211–214. [Google Scholar]
- Lockwood, C.J.; Kayisli, U.A.; Stocco, C.; Murk, W.; Vatandaslar, E.; Buchwalder, L.F.; Schatz, F. Abruption-induced preterm delivery is associated with thrombin-mediated functional progesterone withdrawal in decidual cells. Am. J. Pathol. 2012, 181, 2138–2148. [Google Scholar] [CrossRef]
- Tang, Z.; Tadesse, S.; Norwitz, E.; Mor, G.; Abrahams, V.M.; Guller, S. Isolation of hofbauer cells from human term placentas with high yield and purity. Am. J. Reprod. Immunol. 2011, 66, 336–348. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (Mean ± SD; n = 7) | Preeclampsia (Mean ± SD; n = 8) | p Value | |
|---|---|---|---|
| Gestational Age | 38.15 ± 1.15 | 36.73 ± 1.72 | p = 0.089 |
| BMI | 34.57 ± 13.47 | 34.12 ± 1.1 | p = 0.232 |
| Maternal Age | 32.714 ± 5.99 | 28.62 ± 4.59 | p = 0.159 |
| Birth Weight | 3415.00 ± 507.08 | 2670.62 ± 621.43 | p = 0.026 |
| Antibody | Company and Catalog Number | Application Dilution |
|---|---|---|
| Phospho-ERK1/2 | Cell Signaling #4370. | WB 1:1000 |
| Total-ERK1/2 | Cell Signaling #9102 | WB 1:1000 |
| Phospho-STAT3 | Cell Signaling #9145 | WB 1:1000 |
| Total-STAT3 | Cell Signaling #9139 | WB 1:1000 |
| Phospho-STAT5 | Cell Signaling #9359 | WB 1:1000 |
| Total-STAT5 | Cell Signaling #94205 | WB 1:1000 |
| Phospho-p65 | Cell Signaling #3033 | WB 1:1000 |
| Total-p65 | Cell Signaling #8242 | WB 1:1000 |
| IL-6 | Santa Cruz #sc-28343 | WB 1:1000 |
| Β-Actin | Cell Signaling #5125 | WB 1:1000 |
| IL-6 | R&D #AF-206-NA | IHC: 10 μg/mL |
| LepR (ObR) | Santa Cruz #sc-8391(B-3) | IHC 1:100 |
| CD68 | Santa Cruz #sc-20060 (KP1) | IHC 1:1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozmen, A.; Nwabuobi, C.; Tang, Z.; Guo, X.; Larsen, K.; Guller, S.; Blas, J.; Moore, M.; Kayisli, U.A.; Lockwood, C.J.; et al. Leptin-Mediated Induction of IL-6 Expression in Hofbauer Cells Contributes to Preeclampsia Pathogenesis. Int. J. Mol. Sci. 2024, 25, 135. https://doi.org/10.3390/ijms25010135
Ozmen A, Nwabuobi C, Tang Z, Guo X, Larsen K, Guller S, Blas J, Moore M, Kayisli UA, Lockwood CJ, et al. Leptin-Mediated Induction of IL-6 Expression in Hofbauer Cells Contributes to Preeclampsia Pathogenesis. International Journal of Molecular Sciences. 2024; 25(1):135. https://doi.org/10.3390/ijms25010135
Chicago/Turabian StyleOzmen, Asli, Chinedu Nwabuobi, Zhonghua Tang, Xiaofang Guo, Kellie Larsen, Seth Guller, Jacqueline Blas, Monica Moore, Umit A. Kayisli, Charles J. Lockwood, and et al. 2024. "Leptin-Mediated Induction of IL-6 Expression in Hofbauer Cells Contributes to Preeclampsia Pathogenesis" International Journal of Molecular Sciences 25, no. 1: 135. https://doi.org/10.3390/ijms25010135