
mRNA Levels of Aromatase, 5α-Reductase Isozymes, and Prostate Cancer-Related Genes in Plucked Hair from Young Men with Androgenic Alopecia
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
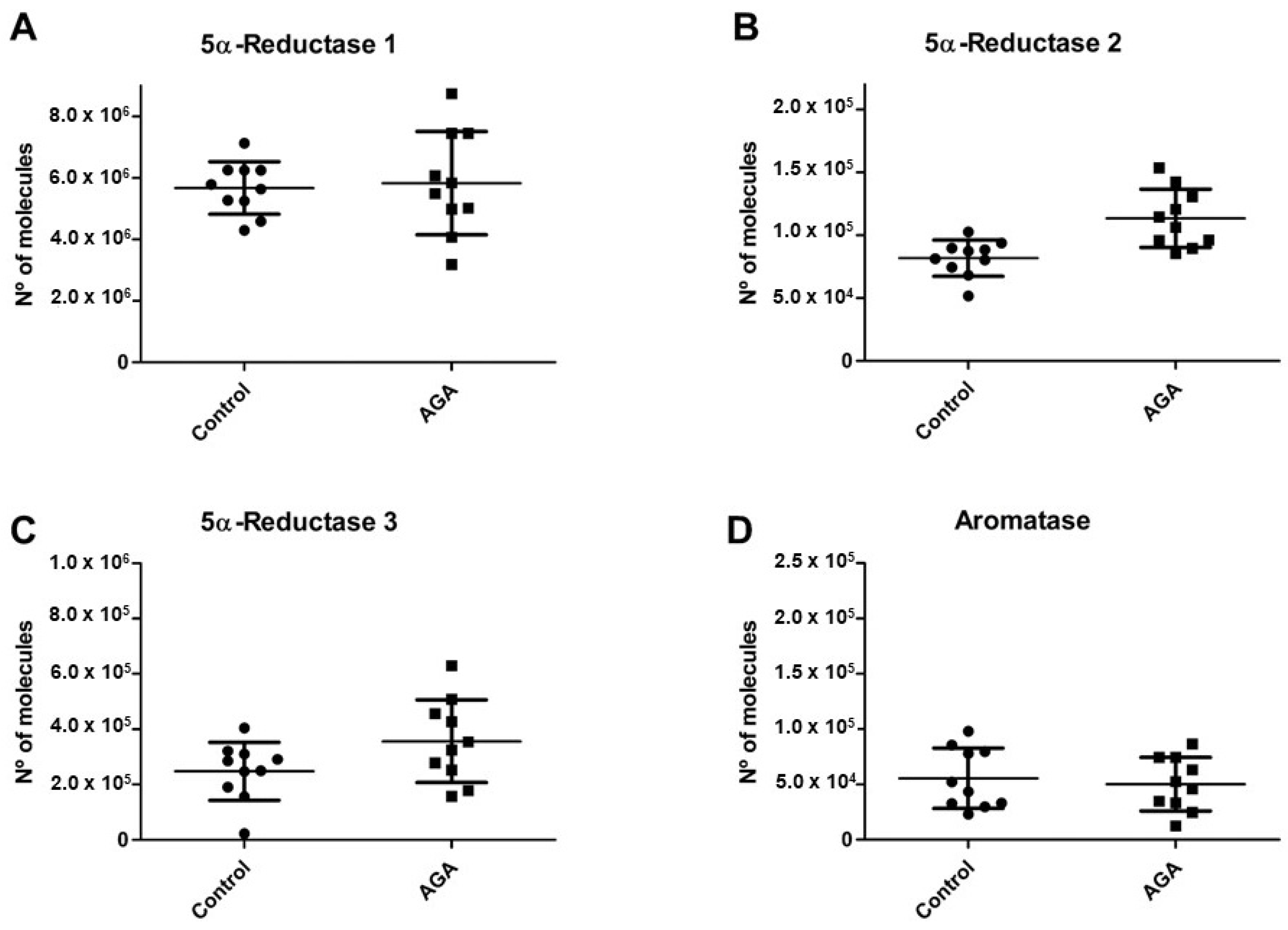
2.1. 5α-R Isozymes and Aromatase mRNA Levels
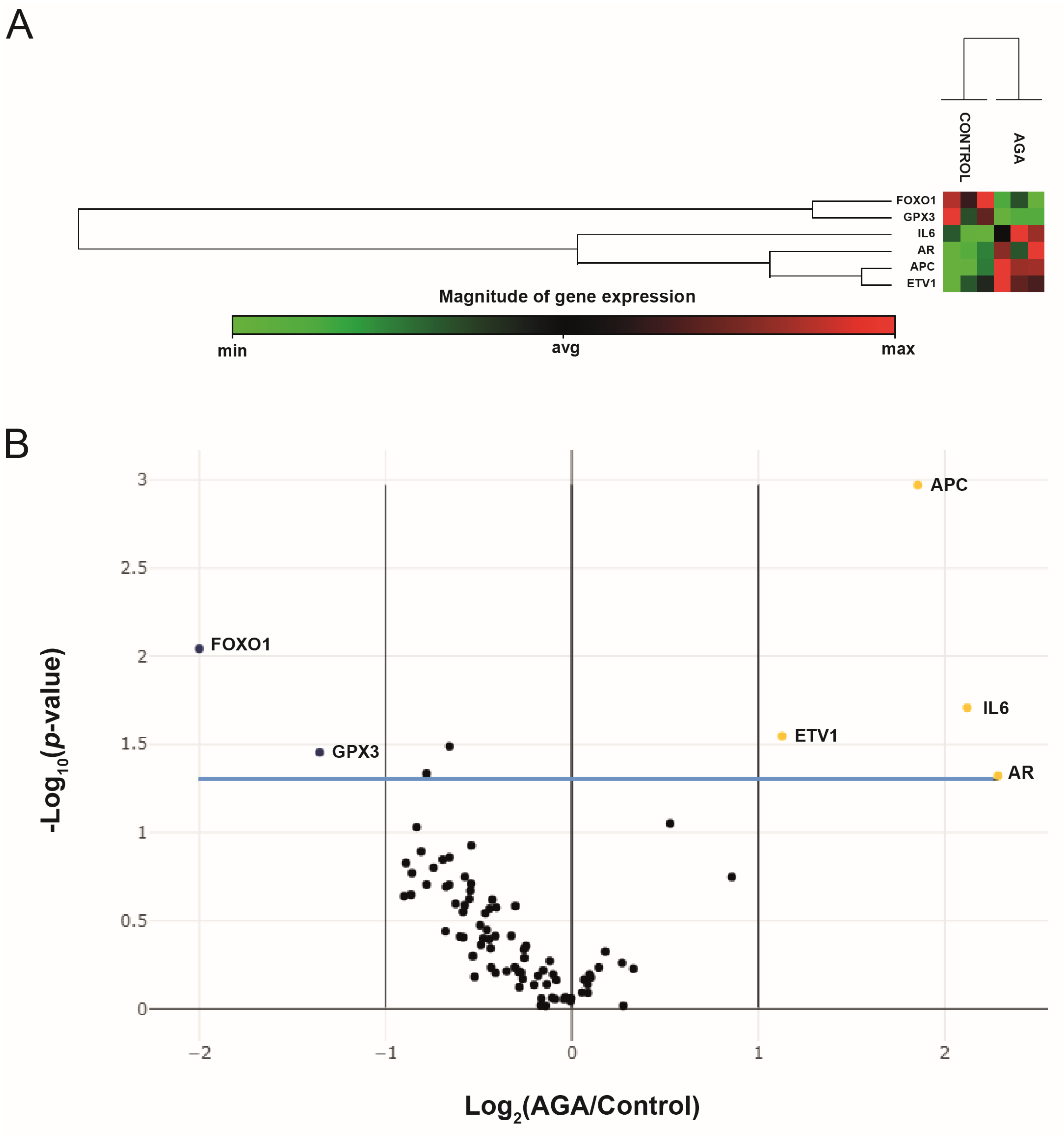
2.2. Analysis of Gene Expression Profiles via PCR Array
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Subjects and Sampling
5.2. RNA Isolation
5.3. Reverse Transcription and Quantitative Real-Time PCR
5.4. Human PCa PCR Array
5.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pierard-Franchimont, C.; Pierard, G.E. Teloptosis, a turning point in hair shedding biorhythms. Dermatology 2001, 203, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, J.B. Patterned loss of hair in man; types and incidence. Ann. N. Y. Acad Sci. 1951, 53, 708–728. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J. Androgenetic alopecia: Current status. Indian J. Dermatol. 2005, 50, 179–190. [Google Scholar]
- Aukerman, E.L.; Jafferany, M. The psychological consequences of androgenetic alopecia: A systematic review. J. Cosmet. Dermatol. 2023, 22, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xie, X.; Zhang, G.; Zhang, Y. Comorbidities in Androgenetic Alopecia: A Comprehensive Review. Dermatol. Ther. 2022, 12, 2233–2247. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Jacobo, L.; Villarreal-Villarreal, C.D.; Ortiz-Lopez, R.; Ocampo-Candiani, J.; Rojas-Martínez, A. Genetic and molecular aspects of androgenetic alopecia. Indian J. Dermatol. Venereol. Leprol. 2018, 84, 263–268. [Google Scholar] [CrossRef] [PubMed]
- English, R.S. A hypothetical pathogenesis model for androgenic alopecia: Clarifying the dihydrotestosterone paradox and rate-limiting recovery factors. Med. Hypotheses 2018, 111, 73–81. [Google Scholar] [CrossRef]
- Lolli, F.; Pallotti, F.; Rossi, A.; Fortuna, M.C.; Caro, G.; Lenzi, A.; Sansone, A.; Lombardo, F. Androgenetic alopecia: A review. Endocrine 2017, 57, 9–17. [Google Scholar] [CrossRef]
- Ellis, J.A.; Stebbing, M.; Harrap, S.B. Polymorphism of the androgen receptor gene is associated with male pattern baldness. J. Invest. Dermatol. 2001, 116, 452–455. [Google Scholar] [CrossRef]
- Sánchez, P.; Serrano-Falcón, C.; Torres, J.M.; Serrano, S.; Ortega, E. 5α-Reductase isozymes and aromatase mRNA levels in plucked hair from young women with female pattern hair loss. Arch. Dermatol. Res. 2018, 310, 77–83. [Google Scholar] [CrossRef]
- Gupta, A.K.; Venkataraman, M.; Talukder, M.; Bamimore, M.A. Relative efficacy of minoxidil and the 5-α reductase inhibitors in androgenetic alopecia treatment of male patients: A network meta-analysis. JAMA Dermatol. 2022, 158, 266–274. [Google Scholar] [CrossRef]
- Imperato-McGineley, J.; Guerrero, L.; Gautier, T.; Peterson, R.E. Steroid 5a-reductase deficiency in man: An inherited form of male pseudohermaphroditism. Science 1974, 86, 1213–1215. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, M.E.; Penneys, N.S. Immunohistochemical distribution of aromatase and 3B-hydroxysteroid dehydrogenase in human hair follicle and sebaceous gland. J. Cutan. Pathol. 1992, 19, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Fujita, K.; Nonomura, N. Role of Androgen Receptor in Prostate Cancer: A Review. World J. Mens. Health 2019, 37, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, P.; Huang, H.; Ye, X.; Chen, W.; Xu, G.; Zhang, F. Androgen receptor regulates eIF5A2 expression and promotes prostate cancer metastasis via EMT. Cell Death Discov. 2021, 7, 373. [Google Scholar] [CrossRef] [PubMed]
- Emond, J.P.; Lacombe, L.; Caron, P.; Turcotte, V.; Simonyan, D.; Aprikian, A.; Saad, F.; Carmel, M.; Chevalier, S.; Guillemette, C.; et al. Urinary oestrogen steroidome as an indicator of the risk of localised prostate cancer progression. Br. J. Cancer 2021, 125, 78–84. [Google Scholar] [CrossRef]
- Lichtenstein, P.; Holm, N.V.; Verkasalo, P.K.; Iliadou, A.; Kaprio, J.; Koskenvuo, M.; Pukkala, E.; Skytthe, A.; Hemminki, K. Environmental and heritable factors in the causation of cancer--analyses of cohorts of twins from Sweden, Denmark, and Finland. N. Engl. J. Med. 2000, 343, 78–85. [Google Scholar] [CrossRef]
- Nyholt, D.R.; Gillespie, N.A.; Heath, A.C.; Martin, N.G. Genetic basis of male pattern baldness. J. Investig. Dermatol. 2003, 121, 1561–1564. [Google Scholar]
- Al Edwan, G.; Bhindi, B.; Margel, D.; Chadwick, K.; Finelli, A.; Zlotta, A.; Trachtenberg, J.; Fleshner, N. The association of male pattern baldness and risk of cancer and high-grade disease among men presenting for prostate biopsy. Can. Urol. Assoc. J. 2016, 10, 424–427. [Google Scholar] [CrossRef]
- Denmark-Wahnefried, W.; Schildkraut, J.M.; Thompson, D.; Lesko, S.M.; McIntyre, L.; Schwingl, P.; Paulson, D.F.; Robertson, C.N.; Anderson, E.E.; Waltherm, P.J. Early onset baldness and prostate cancer risk. Cancer Epidemiol. Biomarkers Prev. 2000, 9, 325–328. [Google Scholar]
- He, H.; Xie, B.; Xie, L. Male pattern baldness and incidence of prostate cancer: A systematic review and meta-analysis. Medicine 2018, 97, e11379. [Google Scholar] [CrossRef] [PubMed]
- Khaled, H.N.; Allah, A.M.A.; Abdelhameed, A.A.; Shehata, W.A. Role of serum androgens and prostate-specific antigen levels in men with androgenetic alopecia. Egypt. J. Dermat. Vener. 2020, 40, 106–111. [Google Scholar] [CrossRef]
- Rokni, G.R.; Gorji, A.M.H.; Sharifian, M.; Talarposhti, A.H.R.; Barzgarnejad, A. The relationship between androgenic alopecia and prostate cancer. Int. J. Med. Res. Health Sci. 2016, 5, 23–29. [Google Scholar]
- Yassa, M.; Saliou, M.; De Rycke, Y.; Hemery, C.; Henni, M.; Bachaud, J.M.; Thiounn, N.; Cosset, J.M.; Giraud, P. Male pattern baldness and the risk of prostate cancer. Ann. Oncol. 2011, 22, 1824–1827. [Google Scholar] [CrossRef] [PubMed]
- Cremers, R.G.; Aben, K.K.; Vermeulen, S.H.; den Heijer, M.; van Oort, I.M.; Kiemeney, L.A. Androgenic alopecia is not useful as an indicator of men at high risk of prostate cancer. Eur. J. Cancer 2010, 46, 3294–3299. [Google Scholar] [CrossRef]
- Liang, W.J.; Song, L.Y.; Peng, Z.; Zou, Y.; Dai, S.M. Possible association between androgenic alopecia and risk of prostate cancer and testicular germ cell tumor: A systematic review and meta-analysis. BMC Cancer 2018, 18, 279. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.L.; Page, S.T.; Lin, D.W.; Stanford, J.L. Male pattern baldness and prostate cancer risk in a population-based case-control study. Cancer Epidemiol. 2010, 34, 131–135. [Google Scholar] [CrossRef]
- Sawaya, M.E.; Price, V.H. Different levels of 5alpha-reductase type I and II, aromatase, and androgen receptor in hair follicles of women and men with androgenetic alopecia. J. Investig. Dermatol. 1997, 109, 296–300. [Google Scholar] [CrossRef]
- Asada, Y.; Sonoda, T.; Ojiro, M.; Kurata, S.; Sato, T.; Ezaki, T.; Takayasu, S. 5 alpha-reductase type 2 is constitutively expressed in the dermal papilla and connective tissue sheath of the hair follicle in vivo but not during culture in vitro. J. Clin. Endocrinol. Metab. 2001, 86, 2875–2880. [Google Scholar]
- Nakanishi, S.; Adachi, I.; Takayasu, K. Expression of androgen receptor, type I and type II 5α-reductase in human dermal papilla cellse. In Hair Research for the Next Millennium; Neste, D., Ed.; VREPB: Amsterdam, The Netherlands, 1996; pp. 333–337. [Google Scholar]
- Piraccini, B.M.; Blume-Peytavi, U.; Scarci, F.; Jansat, J.M.; Falqués, M.; Otero, R.; Tamarit, M.L. Topical Finasteride Study Group. Efficacy and safety of topical finasteride spray solution for male androgenetic alopecia: A phase III, randomized, controlled clinical trial. J. Eur. Acad. Dermatol. Venereol. 2022, 36, 286–294. [Google Scholar] [CrossRef]
- Roberts, J.L.; Fiedler, V.; Imperato-McGinley, J.; Whiting, D.; Olsen, E.; Shupack, J.; Stough, D.; DeVillez, R.; Rietschel, R.; Savin, R.; et al. Clinical dose ranging studies with finasteride, a type 2 5alpha-reductase inhibitor, in men with male pattern hair loss. J. Am. Acad. Dermatol. 1999, 41, 555–563. [Google Scholar] [PubMed]
- Azzouni, F.; Godoy, A.; Li, Y.; Mohler, J. The 5 Alpha-Reductase Isozyme Family: A Review of Basic Biology and Their Role in Human Diseases. Adv. Urol. 2012, 2012, 530121. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Song, S.; Gao, Z.; Wu, J.; Ma, J.; Cui, Y. The efficacy and safety of dutasteride compared with finasteride in treating men with androgenetic alopecia: A systematic review and meta-analysis. Clin. Interv. Aging 2019, 14, 399–406. [Google Scholar] [CrossRef] [PubMed]
- LaTulippe, E.; Satagopan, J.; Smith, A.; Scher, H.; Scardino, P.; Reuter, V.; Gerald, W.L. Comprehensive gene expression analysis of prostate cancer reveals distinct transcriptional programs associated with metastatic disease. Cancer Res. 2002, 62, 4499–4506. [Google Scholar] [PubMed]
- Visakorpi, T.; Hyytinen, E.; Koivisto, P.; Tanner, M.; Keinänen, R.; Palmberg, C.; Palotie, A.; Tammela, T.; Isola, J.; Kallioniemi, O.P. In vivo amplification of the androgen receptor gene and progression of human prostate cancer. Nat. Genet. 1995, 9, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hayes, V.M.; Severi, G.; Eggleton, S.A.; Padilla, E.J.; Southey, M.C.; Sutherland, R.L.; Hopper, J.L.; Giles, G.G. The E211 G>A androgen receptor polymorphism is associated with a decreased risk of metastatic prostate cancer and androgenetic alopecia. Cancer Epidemiol. Biomark. Prev. 2005, 14, 993–996. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wang, L.H.; Farrar, W.L. Interleukin 6 activates androgen receptor-mediated gene expression through a signal transducer and activator of transcription 3-dependent pathway in LNCaP prostate cancer cells. Cancer Res. 2000, 60, 2132–2135. [Google Scholar]
- Lou, W.; Ni, Z.; Dyer, K.; Tweardy, D.J.; Gao, A.C. Interleukin-6 induces prostate cancer cell growth accompanied by activation of stat3 signaling pathway. Prostate 2000, 42, 239–242. [Google Scholar] [CrossRef]
- Chun, J.Y.; Nadiminty, N.; Dutt, S.; Lou, W.; Yang, J.C.; Kung, H.J.; Evans, C.P.; Gao, A.C. Interleukin-6 regulates androgen synthesis in prostate cancer cells. Clin. Cancer Res. 2009, 15, 4815–4822. [Google Scholar] [CrossRef]
- Kwack, M.H.; Ahn, J.S.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dihydrotestosterone-inducible IL-6 inhibits elongation of human hair shafts by suppressing matrix cell proliferation and promotes regression of hair follicles in mice. J. Investig. Dermatol. 2012, 132, 43–49. [Google Scholar] [CrossRef]
- Chaiyana, W.; Punyoyai, C.; Somwongin, S.; Leelapornpisid, P.; Ingkaninan, K.; Waranuch, N.; Srivilai, J.; Thitipramote, N.; Wisuitiprot, W.; Schuster, R.; et al. Inhibition of 5α-Reductase, IL-6 Secretion, and Oxidation Process of Equisetum debile Roxb. ex Vaucher Extract as Functional Food and Nutraceuticals Ingredients. Nutrients 2017, 9, 1105. [Google Scholar] [CrossRef] [PubMed]
- Culig, Z. Interleukin-6 Function and Targeting in Prostate Cancer. Adv. Exp. Med. Biol. 2021, 1290, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Yamada, T. Molecular biology of the Ets family of transcription factors. Gene 2003, 303, 11–34. [Google Scholar] [CrossRef]
- Baena, E.; Shao, Z.; Linn, D.E.; Glass, K.; Hamblen, M.J.; Fujiwara, Y.; Kim, J.; Nguyen, M.; Zhang, X.; Godinho, F.J.; et al. ETV1 directs androgen metabolism and confers aggressive prostate cancer in targeted mice and patients. Genes Dev. 2013, 27, 683–698. [Google Scholar] [CrossRef] [PubMed]
- Khatiwada, P.; Kannan, A.; Malla, M.; Dreier, M.; Shemshedini, L. Androgen up-regulation of Twist1 gene expression is mediated by ETV1. PeerJ 2020, 8, e8921. [Google Scholar] [CrossRef] [PubMed]
- Chew, E.G.Y.; Tan, J.H.J.; Bahta, A.W.; Ho, B.S.; Liu, X.; Lim, T.C.; Sia, Y.Y.; Bigliardi, P.L.; Heilmann, S.; Wan, A.C.A.; et al. Differential Expression between Human Dermal Papilla Cells from Balding and Non-Balding Scalps Reveals New Candidate Genes for Androgenetic Alopecia. J. Investig. Dermatol. 2016, 13, 1559–1567. [Google Scholar] [CrossRef]
- Shiota, M.; Yokomizo, A.; Tada, Y.; Inokuchi, J.; Kashiwagi, E.; Masubuchi, D.; Eto, M.; Uchiumi, T.; Naito, S. Castration resistance of prostate cancer cells caused by castration-induced oxidative stress through Twist1 and androgen receptor overexpression. Oncogene 2010, 29, 237–250. [Google Scholar] [CrossRef]
- Stefanski, C.D.; Prosperi, J.R. Wnt-Independent and Wnt-Dependent Effects of APC Loss on the Chemotherapeutic Response. Int. J. Mol. Sci. 2020, 21, 7844. [Google Scholar] [CrossRef]
- Peifer, M.; Polakis, P. Wnt signalling in oncogenesis and embryogenesis—A look outside the nucleus. Science 2000, 287, 1606–1609. [Google Scholar] [CrossRef]
- Hawkshaw, N.J.; Hardman, J.A.; Alam, M.; Jimenez, F.; Paus, R. Deciphering the molecular morphology of the human hair cycle: Wnt signalling during the telogen-anagen transformation. Br. J. Dermatol. 2020, 182, 1184–1193. [Google Scholar] [CrossRef]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Qureshi, W.M.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef]
- Thomas, A.; Seshamma, T.R.; Trivikram, G.; Sarah, E.M. WNT signals are required for the initiation of hair follicle development. Dev. Cell 2002, 2, 643–653. [Google Scholar]
- Hwang, J.H.; Lee, H.Y.; Chung, K.B.; Lee, H.J.; Kim, J.; Song, K.; Kim, D.Y. Non-thermal atmospheric pressure plasma activates Wnt/β-catenin signaling in dermal papilla cells. Sci. Rep. 2021, 11, 16125. [Google Scholar] [CrossRef]
- Leirós, G.J.; Attorresi, A.I.; Balañá, M.E. Hair follicle stem cell differentiation is inhibited through cross-talk between Wnt/β-catenin and androgen signalling in dermal papilla cells from patients with androgenetic alopecia. Br. J. Dermatol. 2012, 166, 1035–1042. [Google Scholar] [CrossRef]
- Duan, X.; Kong, Z.; Liu, Y.; Zeng, Z.; Li, S.; Wu, W.; Ji, W.; Yang, B.; Zhao, Z.; Zeng, G. β-arrestin2 contributes to cell viability and proliferation via the down-regulation of FOXO1 in castration-resistant prostate cancer. J. Cell. Physiol. 2015, 230, 2371–2381. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Wu, J.; Zhang, T.; Yang, F.; Shao, J.; He, C.; Qin, L. Clinical Evaluation of FOXO1 as a Tumor Suppressor in Prostate Cancer. Comput. Math. Methods Med. 2021, 2021, 8773423. [Google Scholar] [CrossRef] [PubMed]
- Furuyama, T.; Kitayama, K.; Shimoda, Y.; Ogawa, M.; Sone, K.; Yoshida-Araki, K.; Hisatsune, H.; Nishikawa, S.; Nakayama, K.; Nakayama, K.; et al. Abnormal angiogenesis in Foxo1 (Fkhr)-deficient mice. J. Biol. Chem. 2004, 279, 34741–34749. [Google Scholar] [CrossRef] [PubMed]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Investig. 2001, 107, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.P.; Yu, G.; Tseng, G.; Cieply, K.; Nelson, J.; Defrances, M.; Zarnegar, R.; Michalopoulos, G.; Luo, J.H. Glutathione peroxidase 3, deleted or methylated in prostate cancer, suppresses prostate cancer growth and metastasis. Cancer Res. 2007, 67, 8043–8050. [Google Scholar] [CrossRef] [PubMed]
- Hinnant, T.; Lechler, T. Hair Follicle Stem Cells Feel the Pressure. Cell Stem Cell. 2022, 29, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Jadkauskaite, L.; Coulombe, P.A.; Schafer, M.; DinkovaKostova, A.T.; Paus, R.; Haslam, I.S. Oxidative Stress Management in the Hair Follicle: Could Targeting Nrf2 Counter Age-Related Hair. Disorders and Beyond? Bioessays 2017, 39, 1700029. [Google Scholar] [CrossRef]
- Serrano-Falcón, C.; Fernández-Pugnaire, M.A.; Serrano-Ortega, S. Hair and scalp evaluation: The trichogram. Actas Dermosifiliogr. 2013, 104, 867–876. [Google Scholar] [CrossRef]
- Castro, B.; Sánchez, P.; Torres, J.M.; Preda, O.; del Moral, R.G.; Ortega, E. Bisphenol A exposure during adulthood alters expression of aromatase and 5α-reductase isozymes in rat prostate. PLoS ONE 2013, 8, e55905. [Google Scholar] [CrossRef] [PubMed]
- Fronhoffs, S.; Totzke, G.; Stier, S.; Wernert, N.; Rothe, M.; Brüning, T.; Koch, B.; Sachinidis, A.; Vetter, H.; Ko, Y. A method for the rapid construction of cRNA standard curves in quantitative realtime reverse transcription polymerase chain reaction. Mol. Cell Probes 2002, 16, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Roy. Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Gene Symbol | Description | Fold Changes | p |
|---|---|---|---|
| APC | Adenomatous polyposis coli | 3.62 | 0.001070 |
| AR | Androgen receptor | 4.87 | 0.047702 |
| ETV1 | Ets variant 1 | 2.18 | 0.028468 |
| IL6 | Interleukin 6 | 4.35 | 0.019576 |
| FOXO1 | Forkhead box O1 | −4.00 | 0.009073 |
| GPX3 | Glutathione peroxidase 3 | −2.56 | 0.035205 |
| Differentially Methylated Promoters | APC, AR, CAV1, CCNA1, CDH1 (E-Cadherin), CDKN2A (P16INK4A), DKK3, DLC1, EDNRB, GPX3, GSTP1, MGMT (AGT), MSX1, PDLIM4 (RIL), PTGS2 (COX2), RARB, RASSF1, SFRP1, SLC5A8, TIMP2, TNFRSF10D, ZNF185. |
| Upregulated in Prostate Cancer | ARNTL(BMAL1), CAMSAP1, DDX11, ECT2, ETV1, HAL, IGFBP5, KLK3, MTO1, PDPK1, RBM39, SOCS3, SOX4, SUPT7L. |
| Downregulated in Prostate Cancer | CCND2, CLN3, GCA, IGF1, LGALS4, LOXL1, PPP2R1B, SFRP1, SLC5A8, TFPI2, USP5. |
| Metastatic Potential | CREB1, KLHL13, MAX, NDRG3, PES1, SCAF11, SEPT7. |
| Androgen Receptor Signaling | AR, CAV1, CCND1, DAXX, EGFR(ERBB1), FOXO1, GNRH1, IGF1, IL6, NFKB1, NRIP1, PTEN, SHBG, TGFB1I1, TIMP2, TIMP3, VEGFA. |
| AKT & PI3 Kinase Signaling | AKT1, AR, BCL2, CCND1, CCND2, CDH1 (E-Cadherin), CDKN2A (P16INK4A), EGFR(ERBB1), FOXO1, GNRH1, IGF1, IL6, MAPK1 (ERK2), NFKB1, PDPK1, PTEN, TIMP2, TIMP3, TNFRSF10D, TP53 (p53), VEGFA. |
| PTEN Signaling | AKT1, EGFR (ERBB1), GNRH1, IGF1, IL6, MAPK1 (ERK2), PDPK1, PTEN, TIMP2, TIMP3, TP53 (p53), VEGFA. |
| Apoptosis | BCL2, CASP3, CDKN2A (P16INK4A), EGFR (ERBB1), ETV1, GNRH1, IGF1, IL6, MAPK1 (ERK2), NFKB1, PTEN, TIMP2, TIMP3, TP53 (p53), VEGFA. |
| Cell Cycle | APC, BCL2, CASP3, CAV2, CCNA1, CCND1, CCND2, CDKN2A (P16INK4A), EGFR (ERBB1), IGF1, PPP2R1B, PTEN, PTGS1 (COX1), PTGS2 (COX2), TP53 (p53). |
| Transcription Factors | AR, ARNTL (BMAL1), CDKN2A (P16INK4A), CREB1, DAXX, EGR3, ERG, ETV1, FOXO1, MAX, MSX1, NFKB1, NKX3-1, NRIP1, RARB, RBM39, SOX4, SREBF1, SUPT7L, TP53 (p53). |
| Fatty Acid Metabolism | ACACA, CAMKK1, FASN, HMGCR, IGF1, PRKAB1, SREBF1, STK11 (LKB1). |
| Other Prostate Cancer Genes | MKI67, TMPRSS2. |
| Primer | Forward | Reverse | Size | Annealing Tª (°C) |
|---|---|---|---|---|
| SRD5A1 | AGCCATTGTGCAGTGTATGC | AGCCTCCCCTTGGTATTTTG | 136 | 55 |
| SRD5A2 | TGAATACCCTGATGGGTGG | CAAGCCACCTTGTGGAATC | 154 | 54 |
| SRD5A3 | TCCTTCTTTGCCCAAACATC | CTGATGCTCTCCCTTTACGC | 212 | 60 |
| CYP19A1 | TATTAGGGCCCTGTGTCTGC | TGGGTTGGGACTTTTCCTCC | 193 | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, P.; Serrano Falcón, C.; Martínez Rodríguez, S.; Torres, J.M.; Serrano, S.; Ortega, E. mRNA Levels of Aromatase, 5α-Reductase Isozymes, and Prostate Cancer-Related Genes in Plucked Hair from Young Men with Androgenic Alopecia. Int. J. Mol. Sci. 2023, 24, 17461. https://doi.org/10.3390/ijms242417461
Sánchez P, Serrano Falcón C, Martínez Rodríguez S, Torres JM, Serrano S, Ortega E. mRNA Levels of Aromatase, 5α-Reductase Isozymes, and Prostate Cancer-Related Genes in Plucked Hair from Young Men with Androgenic Alopecia. International Journal of Molecular Sciences. 2023; 24(24):17461. https://doi.org/10.3390/ijms242417461
Chicago/Turabian StyleSánchez, Pilar, Cristina Serrano Falcón, Sergio Martínez Rodríguez, Jesús M. Torres, Salvio Serrano, and Esperanza Ortega. 2023. "mRNA Levels of Aromatase, 5α-Reductase Isozymes, and Prostate Cancer-Related Genes in Plucked Hair from Young Men with Androgenic Alopecia" International Journal of Molecular Sciences 24, no. 24: 17461. https://doi.org/10.3390/ijms242417461