
The Role of Short-Chain Fatty Acids and Altered Microbiota Composition in Autism Spectrum Disorder: A Comprehensive Literature Review


Abstract
:1. Introduction
2. Short-Chain Fatty Acids
2.1. Classification and Sources
2.2. Metabolism and Distribution
2.3. SCFA Levels in ASD
3. Microbiota
3.1. Overview
3.2. Microbiota Shift in Children with ASD

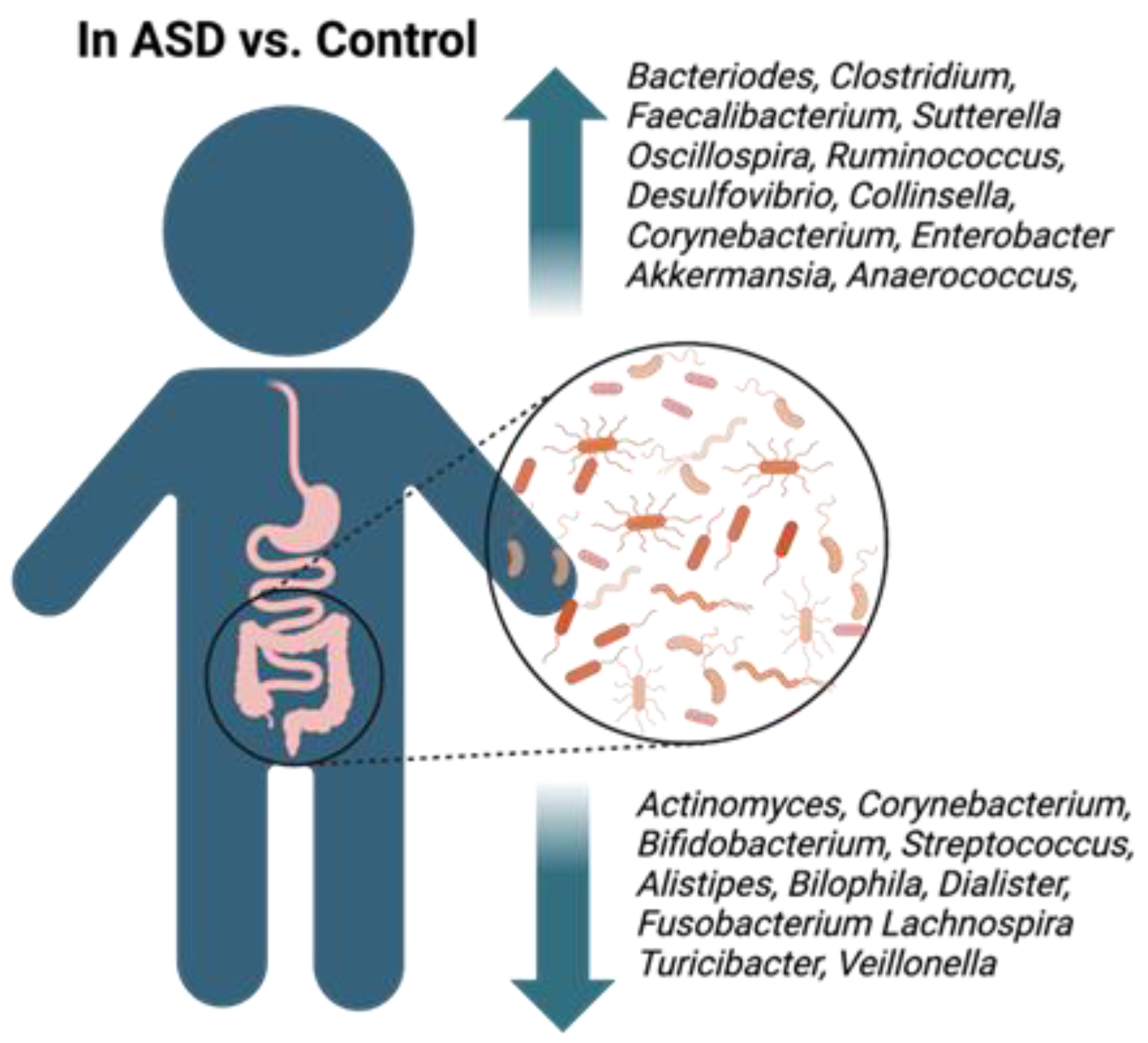{kind=link}
{kind=link}
| Author Year | Study Design | Change in ASD vs. Control | Other Findings | ||||
|---|---|---|---|---|---|---|---|
| Number of Participants | Age (Years) | Sample Source | Assessment Type | Increase | Decrease | ||
| P: Phylum O: Order F: Family G: Genus S: Species | |||||||
| Coretti et al., 2018 [41] | ASD: 11 CON: 14 | 2–4 | Stool | V3–V4 16S rRNA Illumina Miseq System | P: Bacteroidetes, Parabacteroidetes G: Bacteroides, Faecalibacterium Oscillospira, Ruminococcus | P: Actinobacteria G: Actinomyces, Corynebacterium, Bifidobacterium | Increased BA and PPA in ASD |
| Finegold et al., 2010 [44] | ASD: 33 CON: 15 (including 7 siblings of ASD and 8 nonrelated subjects) | 2–14 | Stool | bTEFAP FLX sequencer | P: Bacteroidetes, Proteobacteria G: Desulfovibrio, Turicibacter Bacteroides Parabacteroides S: Desulfovibrio piger, Desulfovibrio Desulfovibrio intestinalis, Bacteroides vulgatus | P: Firmicutes Actinobacteria G: Weissella, Costridium, Actinomyces, Corynebacterium, Bifidobacterium, Ruminococcus Streptococcus, Dialister S: Dialister invisus, Bifidobacterium longum, Clostridium leptum | Very high level of Bacteroides in severe cases of ASD |
| Parracho et al., 2005 [58] | ASD: 58 CON: 22 (12 siblings of ASD and 10 not related) | ASD: 3–16 CON: 2–13 | Stool | FISH 16S rRNA oligonucleotide probes | S: Clostridium histolyticum | A high portion of the ASD group had GI issues | |
| Strati et al., 2017 [20] | ASD: 40 CON: 40 | 4–17 | Stool | V3–V5 16S rRNA. GS FLX + system | G: Collinsella, Corynebacterium, Dorea, Lactobacillus | G: Alistipes, Bilophila, Dialister, Parabacteroides, and Veillonella | ASD altered microbiota, constipation is an important factor |
| De Angelis et al., 2013 [42] | ASD: 10 CON: 10 siblings | 4–10 | Stool | bTEFAP 454 FLX Sequencer | P: Bacteroidetes, G: Bacteroides Clostridium Roseburia Enterobacter Akkermansia | P: Fusobacteria, Verrucomicrobia G: Eubacterium, Fusobacterium, Lachnospira, Turicibacter, Bifidobacterium | Increase in PPA and AA |
| Wang et al., 2020 [65] | ASD: 26 CON: 24 | 3–9 | Stool | V1-V2 16S rRNA Illumina HiSeq sequencer | F: Rikenellaceae, G: Ruminococcus, Victivallales Oscillospira, Odoribacter, Cetobacterium, | P: Actinobacteria O: Bifidobacteriales, F: Bifidobacteriaceae Veillonellaceae, G: Bifidobacterium, S: B. adolescentis, B. longum | Decrease in PPA in ASD Odoribacter: common SCFA producer |
| Li et al., 2019 [55] | ASD: 59 children and their mothers CON: 30 children and their mothers | Children: 2–10 Mothers: 26–42 | Stool | V1-V2 16S rRNA Illumina HiSeq sequencer | Children- G: Enhydrobacter, Chryseobacterium, Streptococcus, Acinetobacter, Clostridium S: Acinetobacter rhizosphaerae, Acinetobacter johnsonii Mothers-F: Moraxellaceae Enterobacteriaceae G: Acinetobacter | Children-S: Prevotella melaninogenica Mothers- G: Faecalibacterium | Assessment of mother–child gut microbiome profile. There is a clear correlation; however, a unique bacteria profile is still present in ASD children. |
| Kushak et al., 2017 [69] | ASD: 21 CON: 19 Both ASD and CON with GI symptoms | ASD: 14.43 ± 1.07 CON: 16.05 ± 1.25 | Duodenum, endoscopic biopsy | 16S rRNA 454 FLX Sequencer | G: Burkholderia, Oscillospira, Actinomyces, Neisseria, Peptostreptococcus, Ralstonia, | G: Neisseria, Devosia, Prevotella, Bacteroides, Streptococcus | Differences in bacteria associated with disaccharidase activity |
| Williams et al., 2011 [71] | ASD: 15 CON: 7 Both ASD and CON children had GI issues | 3–6 | Biopsy of ileal and cecal tissues | V2 16S rRNA 454 FLX Sequencer | O: Clostridiale F: Lachnospiraceae, Ruminococcaceae, Alcaligenaceae, Methylobacteriaceae | P: Bacteroidetes | Deficits in gene expression involved in carbohydrate digestion and transport |
| Williams et al., 2012 [70] | ASD: 15 CON: 7 | 3–5 | Biopsy of ilium and cecum | V2 16S rRNA GS FLX sequencer | High level of species from Sutterella genus | Sutterella 16S rRNA in ASD group and absent in control | |
| Adams et al., 2011 [66] | ASD: 58 CON: 39 | ASD: 6.91 ± 3.4 CON: 7.7 ± 4.4 | Stool | The Vitek®2 identification cards and Vitek 2 system | G: Lactobacillus, Bacillus spp. | G: Bifidobacterium, Enterococcus Species: Enterobacter cloacae | Decrease in SCFAs (lower SCFAs due to higher absortion/lower intake of fibers) |
| Tomova et al., 2015 [73] | ASD: 10 CON: 10 Siblings of ASD: 9 | ASD: 2–9 CON:2–11 Sib.: 5–17 | Stool | RT-PCR | Clostridia cluster l, Desulfovibrio | P: Bacteroidetes | Fecal TNFα increased in stool. Correlation between the amount of Desulfovibrio present and autism severity |
| Wang et al., [74] | ASD: 23 ASD siblings: 22 CON (unrelated): 9 | ASD: 10.2 ± 0.75 CON: 9.5± 1.25 Sib.: 12 ± 1 | Stool | RT-PCR | S: Clostridium difficile | S: Akkermansia muciniphila, Bifidobacterium spp. | Lower abundance of Akkermansia muciniphila is suggestive of changes in the mucosal barrier |
| David et al., 2021 [56] | ASD: 60 CON: 57 (siblings) | 2–11 | Stool | 16S rRNA V4 Illumina MiSeq | G: Bacteroides, Ruminococcus, Anaerococcus | F: Lachnospiraceae G: Desulfovibrio, Bifidobacterium | Unique crowdsourcing recruitment of subjects. |
| Kang et al., 2013 [75] | ASD: 20 CON: 20 | 2–16 | Stool | V2/V3 16S bTEFAP FLX Sequencer | G: Akkermansia present at very high level | P: Proteobacteria, Verrucomicrobi, G: Veillonellaceae, Prevotella, Coprococcus | Less diverse gut microbial composition in ASD |
| Finegold et al., 2017 [18] | ASD: 33 CON: 13 | 2–9 | Stool | Anerobic bacteria culture. ABI 3130 | Increase in Clostridium | Increase in C. perfringens beta2-toxin gene in ASD vs. control | |
| Song et al., 2004 [76] | ASD: 15 CON: 8 | Not specified | Stool | TaqMan RT-PCR 16S rRNA | Increases in Clostridium 46-fold: C. bolteae 9.0-fold: cluster I 3.5-fold: cluster XI | Study focused on Clostridium | |
| Zhang et al., 2018 [77] | ASD: 35 CON: 6 | ASD: 4.9 ± 1.5 CON: 4.6 ± 1.1 | Stool | 16S rRNA (V3–V4) Illumina HiSeq | P: Bacteroidetes G: Sutterella, Odoribacter, Butyricimona | P: Firmicutes Genus: Veillonella, Streptococcus | ASD group was characterized by increase in constipation |
| Son et al., 2015 [78] | ASD: 59 CON: 44 (siblings of ASD) | ASD:4–18 CON:7–14 | Stool | V1V2 and V1V3 of 16S rRNA Illumina HiSeq | No difference found | No difference found | ASD group was characterized by increase in constipation |
| Wang et al., 2013 [79] | ASD: 23 CON: 31 | Not specified | Stool | RT-PCR | G: Sutterella S: Ruminococcus torques | Focused on Sutterella | |
| Jendraszak et al., 2021 [67] | ASD: 33 CON: 16 Allergies: 24 | ASD: 4–6 CON: 3–9 ALG: 4–9 | Stool | Microbial culture and RT-PCR | G: Klebsiella, Bifidobacterium | Probiotic use helps stabilize microbial composition | |
| He et al., 2023 [40] | ASD: 40 CON: 40 | ASD: 5.3 ± 1.34 CON: 5.83 ± 1.28 | Stool | V3-V4 of the 16S rRNA Illumina HiSeq 2500 | Ruminococcaceae_UCG_002, Erysipelotrichaceae_UCG_003, Phascolarctobacterium, Megamonas, Ruminiclostridium_5, Parabacteroides, Prevotella_2, Fusobacterium, Prevotella 9 | Anaerostipes, Lactobacillus, Ruminococcus_gnavus_group, Lachnospiraceae_NK4A136_group, Ralstonia, Eubacterium_eligens_ group, and Ruminococcus_1 | Children enrolled in this study suffered from constipation. Significant increase in SCFAs in the ASD group |
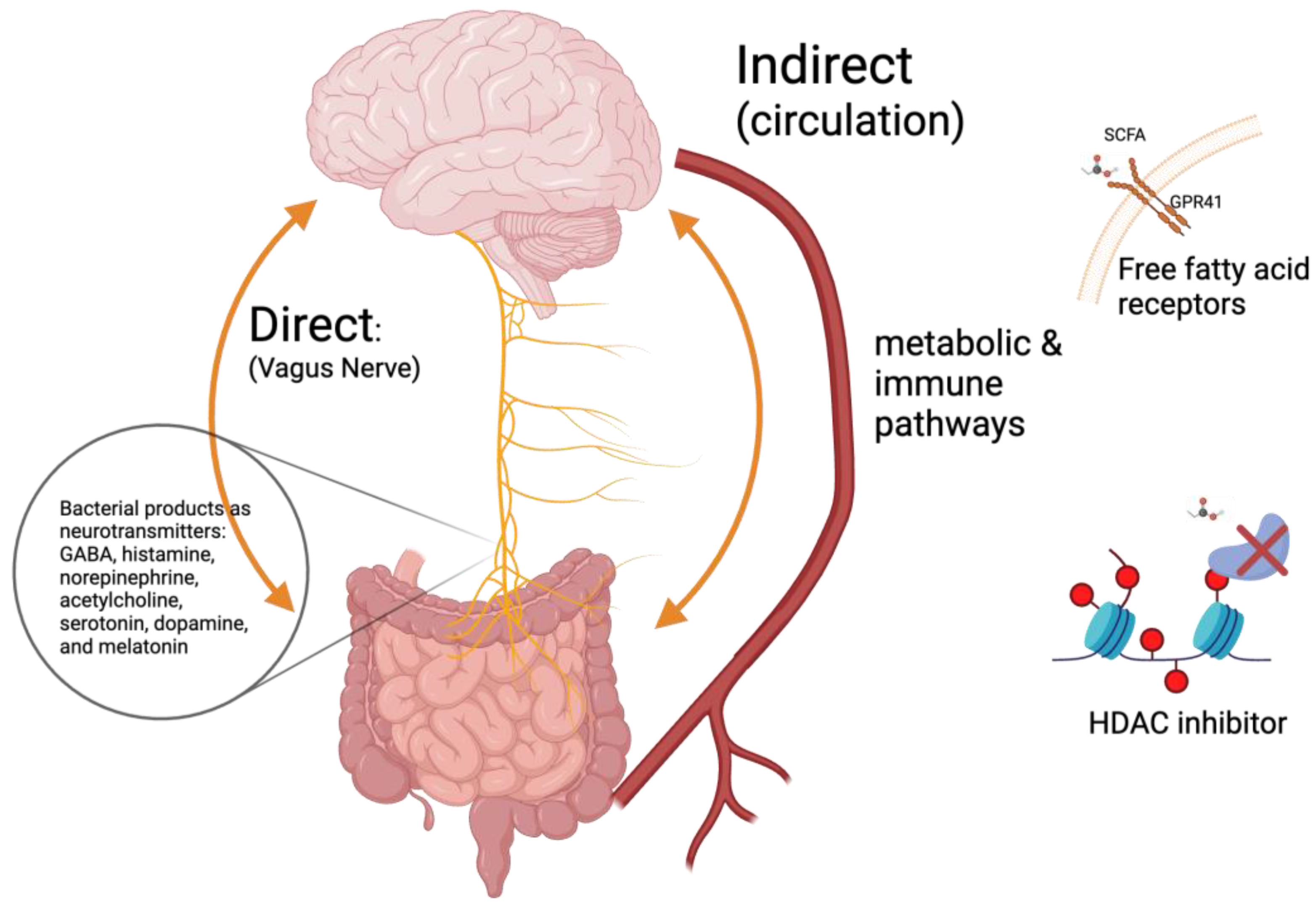
3.3. Gut–Brain Axis
3.4. In Vivo Effect of SCFAs in Adult ASD-Animal Model
| Author Year | Study Design | Outcomes | ||
|---|---|---|---|---|
| Animal | Sample Size | Treatment | ||
| Sharon et al., 2019 [102] | Mice: Germ-free C57BL/6J weanlings (3–4 weeks of age) | 16 donor fecal samples 9 animals colonized by bacteria from each donor sample | GF mice grafted with gut microbiota from ASD and TD control subjects | Microbiota from ASD altered the behavior of mice: increased repetitive behavior, decreased locomotion, and decreased communication. It also induced alternative splicing of genes in the mice brain in ASD vs. TD control. Differences in the metabolome profile. |
| MacFabe et al., 2007 [105] | Adult male Long–Evans rats (~75 days old) | Total of 74 rats across groups Group sizes 6–9 animals | Infusion with PPA. Low: 4.0 μL of a 0.052 M solution; high: PPA (4.0 μL of a 0.26 M solution. Controls: PBS or propanol | PPA treatment: increase in oxidative stress markers. Altered behavior (repetitive dystonic behaviors, hyperactivity, and turning behavior). Increased reactive astrogliosis (GFAP immunoreactivity) and activated microglia (CD68 immunoreactivity). |
| Meeking et al., 2020 [112] | Adult male Long–Evans | Total of 35 rats across groups | 7 days, twice a day, 4 h apart, infusion of buffered PPA (low dose 0.052 M or high dose 0.26 M, pH 7.5, 4 μL/infusion) control: phosphate buffered saline (PBS, 0.1 M) | PPA-treated rats exhibited more locomotive activity, stereotypic behavior, and nose pokes versus control, which are associated with a rat model of ASD. The symptoms were dose-dependent and increased with consecutive treatments. |
| De Theije et al., 2014 [113] | BALB/C mice from Charles River laboratories | 8 pups in treatment group and 11 in control | Dams treated at gestational day 11 with 600 mg/kg of valproic acid (VPA). Pups weaned at P21. Behavioral experiments performed at P28, after which they were sacrificed. VPA treatment during gestation is well established in the animal model of ASD. | An increase in cecal levels of BA in in utero VPA-treated pups vs. control. A decrease in Bacteroidales (order) and increase in Clostridiales (order) in VPA vs. control. Increased neutrophil infiltration in the intestine. |
| MacFabe et al., 2011 [106] | Adolescent (41 ± 4 days) Long–Evans male | 20 and 17 animals in PPA and control groups, respectively | Intracerebroventricular injection of 4 μl of 0.26 M buffered PPA prior to each test session | PPA vs. control group characterized by activation of microglia and astrocytes, lesser sociability, and a focus on particular objects in a group of objects. |
| El-Ansary et al., 2015 [107] | Male Western albino rats | 6 animals in each group | PPA: 250 mg/kg body weight/day (orally) Ampicillin: 50 mg/kg for three weeks | Treatment with PPA and ampicillin led to an increase in catalase activity and lipid peroxidation, while glutathione and potassium levels were decreased in comparison to the control group. |
| El-Ansary et al., 2018 [108] | Young male golden Syrian hamsters | 10 animals in each group | PPA: 250 mg per kg of body weight (BW) (oroigastric) Clindamycin: 30 mg single dose | An increase in Candida albicans and Clostridia in PPA and clindamycin groups. An increase in Na+/Mg2+ and glutamate/GABA ratios. |
| Lobzhanidze et al., 2019 [109] | Adolescent male Wistar rats (P30–35) | 15 animals in each group | Single injection of buffered PPA with a dose of 175 mg/kg | In the PPA vs. control groups, the number of neurons was decreased, while the number of glial cells was increased in the amygdala. Also, both microglia and astrocytes were activated, and neurons exhibited signs of apoptosis. The behavioral changes include decreased sociability (a decrease in the amount of time and number of encounters with unfamiliar rats). |
| Foley et al., 2014 [110] | Long–Evans rats, offspring treated in utero and postnatal | 8 to 11 animals in each group | Prenatal administration of PPA (500 mg/kg,) and LPS (50 μg/kg). Postnatal PPA administered at PPA (500 mg/kg) | Treatments (both prenatal and postnatal) altered the behavior of rodents to autism-like behavior. PPA-treated rats spend less time in the center of the open field and exhibited increased anxiety. Treatment induced delays in eye opening. |
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Vahia, V.N. Diagnostic and statistical manual of mental disorders 5: A quick glance. Indian. J. Psychiatry 2013, 55, 220–223. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, J.; Man, H.Y. Fundamental Elements in Autism: From Neurogenesis and Neurite Growth to Synaptic Plasticity. Front. Cell Neurosci. 2017, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Abdelli, L.S.; Samsam, A.; Naser, S.A. Propionic Acid Induces Gliosis and Neuro-inflammation through Modulation of PTEN/AKT Pathway in Autism Spectrum Disorder. Sci. Rep. 2019, 9, 8824. [Google Scholar] [CrossRef]
- Lord, C.; Risi, S.; DiLavore, P.S.; Shulman, C.; Thurm, A.; Pickles, A. Autism from 2 to 9 years of age. Arch. Gen. Psychiatry 2006, 63, 694–701. [Google Scholar] [CrossRef]
- Committee to Evaluate the Supplemental Security Income Disability Program for Children with Mental Disorders; Board on the Health of Select Populations; Board on Children, Youth, and Families; Institute of Medicine; Division of Behavioral and Social Sciences and Education; National Academies of Sciences, Engineering, and Medicine. Mental Disorders and Disabilities Among Low-Income Children; Boat, T.F., Wu, J.T., Eds.; National Academies Press: Washington, DC, USA, 2015. [Google Scholar]
- Diagnostic and Statistical Manual of Mental Disorders, DSM-5, 5th ed.; American Psychiatric Assiciation: Arlington, VA, USA, 2013.
- Maenner, M.J.; Shaw, K.A.; Bakian, A.V.; Bilder, D.A.; Durkin, M.S.; Esler, A.; Furnier, S.M.; Hallas, L.; Hall-Lande, J.; Hudson, A.; et al. Prevalence and Characteristics of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2018. MMWR Surveill. Summ. 2021, 70, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiao, L.; Chen, R.S.; Chen, C.; Xun, G.L.; Lu, X.Z.; Shen, Y.D.; Wu, R.R.; Xia, K.; Zhao, J.P.; et al. Social impairment of children with autism spectrum disorder affects parental quality of life in different ways. Psychiatry Res. 2018, 266, 168–174. [Google Scholar] [CrossRef]
- Modabbernia, A.; Velthorst, E.; Reichenberg, A. Environmental risk factors for autism: An evidence-based review of systematic reviews and meta-analyses. Mol. Autism 2017, 8, 13. [Google Scholar] [CrossRef]
- Bölte, S.; Girdler, S.; Marschik, P.B. The contribution of environmental exposure to the etiology of autism spectrum disorder. Cell Mol. Life Sci. 2019, 76, 1275–1297. [Google Scholar] [CrossRef]
- Estes, M.L.; McAllister, A.K. Immune mediators in the brain and peripheral tissues in autism spectrum disorder. Nat. Rev. Neurosci. 2015, 16, 469–486. [Google Scholar] [CrossRef]
- Rosenberg, R.E.; Law, J.K.; Yenokyan, G.; McGready, J.; Kaufmann, W.E.; Law, P.A. Characteristics and concordance of autism spectrum disorders among 277 twin pairs. Arch. Pediatr. Adolesc. Med. 2009, 163, 907–914. [Google Scholar] [CrossRef]
- Rossignol, D.A.; Frye, R.E. A review of research trends in physiological abnormalities in autism spectrum disorders: Immune dysregulation, inflammation, oxidative stress, mitochondrial dysfunction and environmental toxicant exposures. Mol. Psychiatry 2012, 17, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Silver, W.G.; Rapin, I. Neurobiological basis of autism. Pediatr. Clin. N. Am. 2012, 59, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Bauman, M.L. Medical comorbidities in autism: Challenges to diagnosis and treatment. Neurotherapeutics 2010, 7, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, E.Y. Gastrointestinal Issues in Autism Spectrum Disorder. Harv. Rev. Psychiatry 2014, 22, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Hughes, H.K.; Rose, D.; Ashwood, P. The Gut Microbiota and Dysbiosis in Autism Spectrum Disorders. Curr. Neurol. Neurosci. Rep. 2018, 18, 81. [Google Scholar] [CrossRef] [PubMed]
- Finegold, S.M.; Summanen, P.H.; Downes, J.; Corbett, K.; Komoriya, T. Detection of Clostridium perfringens toxin genes in the gut microbiota of autistic children. Anaerobe 2017, 45, 133–137. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The Gut Microbiota and Autism Spectrum Disorders. Front. Cell. Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef]
- Strati, F.; Cavalieri, D.; Albanese, D.; De Felice, C.; Donati, C.; Hayek, J.; Jousson, O.; Leoncini, S.; Renzi, D.; Calabrò, A.; et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome 2017, 5, 24. [Google Scholar] [CrossRef]
- Brody, T.O.M. 6—LIPIDS. In Nutritional Biochemistry, 2nd ed.; Brody, T.O.M., Ed.; Academic Press: San Diego, CA, USA, 1999; pp. 311–378. [Google Scholar]
- Schönfeld, P.; Wojtczak, L. Short- and medium-chain fatty acids in energy metabolism: The cellular perspective. J. Lipid Res. 2016, 57, 943–954. [Google Scholar] [CrossRef]
- Íñiguez-Gutiérrez, L.; Godínez-Méndez, L.A.; Fafutis-Morris, M.; Padilla-Arellano, J.R.; Corona-Rivera, A.; Bueno-Topete, M.R.; Rojas-Rejón Ó, A.; Delgado-Rizo, V. Physiological concentrations of short-chain fatty acids induce the formation of neutrophil extracellular traps in vitro. Int. J. Immunopathol. Pharmacol. 2020, 34, 2058738420958949. [Google Scholar] [CrossRef]
- Yang, L.L.; Millischer, V.; Rodin, S.; MacFabe, D.F.; Villaescusa, J.C.; Lavebratt, C. Enteric short-chain fatty acids promote proliferation of human neural progenitor cells. J. Neurochem. 2020, 154, 635–646. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of propionate and butyrate by the human colonic microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46 (Suppl. 2), S33–S50. [Google Scholar] [PubMed]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [PubMed]
- Macfarlane, S.; Macfarlane, G.T. Regulation of short-chain fatty acid production. Proc. Nutr. Soc. 2003, 62, 67–72. [Google Scholar] [CrossRef]
- Shimizu, H.; Masujima, Y.; Ushiroda, C.; Mizushima, R.; Taira, S.; Ohue-Kitano, R.; Kimura, I. Dietary short-chain fatty acid intake improves the hepatic metabolic condition via FFAR3. Sci. Rep. 2019, 9, 16574. [Google Scholar] [CrossRef]
- EFSA Panel on Food additives and Nutrient Sources added to Food (ANS). Scientific Opinion on the re-evaluation of propionic acid (E 280), sodium propionate (E 281), calcium propionate (E 282) and potassium propionate (E 283) as food additives. EFSA J. 2014, 12, 3779. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Jädert, C.; Holm, L.; Nyman, M.E. Propionic and butyric acids, formed in the caecum of rats fed highly fermentable dietary fibre, are reflected in portal and aortic serum. Br. J. Nutr. 2013, 110, 1565–1572. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Huo, Y.J.; Li, Y.; Han, Y.; Zhou, D. Gut-brain axis: Focus on gut metabolites short-chain fatty acids. World J. Clin. Cases 2022, 10, 1754–1763. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [PubMed]
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. (Lausanne) 2020, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Edmonson, C.; Ziats, M.N.; Rennert, O.M. Altered glial marker expression in autistic post-mortem prefrontal cortex and cerebellum. Mol. Autism 2014, 5, 3. [Google Scholar] [CrossRef]
- He, J.; Gong, X.; Hu, B.; Lin, L.; Lin, X.; Gong, W.; Zhang, B.; Cao, M.; Xu, Y.; Xia, R.; et al. Altered Gut Microbiota and Short-chain Fatty Acids in Chinese Children with Constipated Autism Spectrum Disorder. Sci. Rep. 2023, 13, 19103. [Google Scholar] [CrossRef]
- Coretti, L.; Paparo, L.; Riccio, M.P.; Amato, F.; Cuomo, M.; Natale, A.; Borrelli, L.; Corrado, G.; Comegna, M.; Buommino, E.; et al. Gut Microbiota Features in Young Children With Autism Spectrum Disorders. Front. Microbiol. 2018, 9, 3146. [Google Scholar] [CrossRef]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal microbiota and metabolome of children with autism and pervasive developmental disorder not otherwise specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef]
- de la Bâtie, C.D.; Barbier, V.; Roda, C.; Brassier, A.; Arnoux, J.B.; Valayannopoulos, V.; Guemann, A.S.; Pontoizeau, C.; Gobin, S.; Habarou, F.; et al. Autism spectrum disorders in propionic acidemia patients. J. Inherit. Metab. Dis. 2018, 41, 623–629. [Google Scholar] [CrossRef]
- Finegold, S.M.; Dowd, S.E.; Gontcharova, V.; Liu, C.; Henley, K.E.; Wolcott, R.D.; Youn, E.; Summanen, P.H.; Granpeesheh, D.; Dixon, D.; et al. Pyrosequencing study of fecal microflora of autistic and control children. Anaerobe 2010, 16, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [PubMed]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [PubMed]
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [PubMed]
- D’Argenio, V. The Prenatal Microbiome: A New Player for Human Health. High. Throughput 2018, 7, 38. [Google Scholar] [CrossRef]
- Dominguez-Bello, M.G.; Costello, E.K.; Contreras, M.; Magris, M.; Hidalgo, G.; Fierer, N.; Knight, R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc. Natl. Acad. Sci. USA 2010, 107, 11971–11975. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Barrett, E.; Kerr, C.; Murphy, K.; O’Sullivan, O.; Ryan, C.A.; Dempsey, E.M.; Murphy, B.P.; O’Toole, P.W.; Cotter, P.D.; Fitzgerald, G.F.; et al. The individual-specific and diverse nature of the preterm infant microbiota. Arch. Dis. Child. Fetal Neonatal Ed. 2013, 98, F334–F340. [Google Scholar] [CrossRef]
- Pereira, F.C.; Berry, D. Microbial nutrient niches in the gut. Environ. Microbiol. 2017, 19, 1366–1378. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Li, S.; Gan, R.Y.; Zhou, T.; Xu, D.P.; Li, H.B. Impacts of gut bacteria on human health and diseases. Int. J. Mol. Sci. 2015, 16, 7493–7519. [Google Scholar] [CrossRef]
- Fattorusso, A.; Di Genova, L.; Dell’Isola, G.B.; Mencaroni, E.; Esposito, S. Autism Spectrum Disorders and the Gut Microbiota. Nutrients 2019, 11, 521. [Google Scholar] [CrossRef]
- Liu, F.; Li, J.; Wu, F.; Zheng, H.; Peng, Q.; Zhou, H. Altered composition and function of intestinal microbiota in autism spectrum disorders: A systematic review. Transl. Psychiatry 2019, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- David, M.M.; Tataru, C.; Daniels, J.; Schwartz, J.; Keating, J.; Hampton-Marcell, J.; Gottel, N.; Gilbert, J.A.; Wall, D.P. Children with Autism and Their Typically Developing Siblings Differ in Amplicon Sequence Variants and Predicted Functions of Stool-Associated Microbes. mSystems 2021, 6, e00193-20. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Yang, J.; Zhang, J.; Liang, C.; Wang, Y.; Chen, B.; Zhao, C.; Wang, J.; Zhang, G.; Zhao, D.; et al. Correlation of Gut Microbiome Between ASD Children and Mothers and Potential Biomarkers for Risk Assessment. Genom. Proteom. Bioinform. 2019, 17, 26–38. [Google Scholar] [CrossRef]
- Parracho, H.M.; Bingham, M.O.; Gibson, G.R.; McCartney, A.L. Differences between the gut microflora of children with autistic spectrum disorders and that of healthy children. J. Med. Microbiol. 2005, 54, 987–991. [Google Scholar] [CrossRef]
- Hughes, R.; Magee, E.A.; Bingham, S. Protein degradation in the large intestine: Relevance to colorectal cancer. Curr. Issues Intest. Microbiol. 2000, 1, 51–58. [Google Scholar] [PubMed]
- Niehus, R.; Lord, C. Early medical history of children with autism spectrum disorders. J. Dev. Behav. Pediatr. 2006, 27, S120–S127. [Google Scholar] [CrossRef]
- Finegold, S.M. Therapy and epidemiology of autism--clostridial spores as key elements. Med. Hypotheses 2008, 70, 508–511. [Google Scholar] [CrossRef]
- Finegold, S.M. Desulfovibrio species are potentially important in regressive autism. Med. Hypotheses 2011, 77, 270–274. [Google Scholar] [CrossRef]
- Sandler, R.H.; Finegold, S.M.; Bolte, E.R.; Buchanan, C.P.; Maxwell, A.P.; Väisänen, M.L.; Nelson, M.N.; Wexler, H.M. Short-term benefit from oral vancomycin treatment of regressive-onset autism. J. Child. Neurol. 2000, 15, 429–435. [Google Scholar] [CrossRef]
- De Angelis, M.; Francavilla, R.; Piccolo, M.; De Giacomo, A.; Gobbetti, M. Autism spectrum disorders and intestinal microbiota. Gut Microbes 2015, 6, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, N.; Yang, J.J.; Zhao, D.M.; Chen, B.; Zhang, G.Q.; Chen, S.; Cao, R.F.; Yu, H.; Zhao, C.Y.; et al. Probiotics and fructo-oligosaccharide intervention modulate the microbiota-gut brain axis to improve autism spectrum reducing also the hyper-serotonergic state and the dopamine metabolism disorder. Pharmacol. Res. 2020, 157, 104784. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.B.; Audhya, T.; McDonough-Means, S.; Rubin, R.A.; Quig, D.; Geis, E.; Gehn, E.; Loresto, M.; Mitchell, J.; Atwood, S.; et al. Nutritional and metabolic status of children with autism vs. neurotypical children, and the association with autism severity. Nutr. Metab. (Lond.) 2011, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Jendraszak, M.; Gałęcka, M.; Kotwicka, M.; Regdos, A.; Pazgrat-Patan, M.; Andrusiewicz, M. Commercial microbiota test revealed differences in the composition of intestinal microorganisms between children with autism spectrum disorders and neurotypical peers. Sci. Rep. 2021, 11, 24274. [Google Scholar] [CrossRef]
- Yao, S.; Zhao, Z.; Wang, W.; Liu, X. Bifidobacterium Longum: Protection against Inflammatory Bowel Disease. J. Immunol. Res. 2021, 2021, 8030297. [Google Scholar] [CrossRef]
- Kushak, R.I.; Winter, H.S.; Buie, T.M.; Cox, S.B.; Phillips, C.D.; Ward, N.L. Analysis of the Duodenal Microbiome in Autistic Individuals: Association With Carbohydrate Digestion. J. Pediatr. Gastroenterol. Nutr. 2017, 64, e110–e116. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Hornig, M.; Parekh, T.; Lipkin, W.I. Application of novel PCR-based methods for detection, quantitation, and phylogenetic characterization of Sutterella species in intestinal biopsy samples from children with autism and gastrointestinal disturbances. mBio 2012, 3, e00261-11. [Google Scholar] [CrossRef]
- Williams, B.L.; Hornig, M.; Buie, T.; Bauman, M.L.; Cho Paik, M.; Wick, I.; Bennett, A.; Jabado, O.; Hirschberg, D.L.; Lipkin, W.I. Impaired carbohydrate digestion and transport and mucosal dysbiosis in the intestines of children with autism and gastrointestinal disturbances. PLoS ONE 2011, 6, e24585. [Google Scholar] [CrossRef]
- Liu, S.; Li, E.; Sun, Z.; Fu, D.; Duan, G.; Jiang, M.; Yu, Y.; Mei, L.; Yang, P.; Tang, Y.; et al. Altered gut microbiota and short chain fatty acids in Chinese children with autism spectrum disorder. Sci. Rep. 2019, 9, 287. [Google Scholar] [CrossRef]
- Tomova, A.; Husarova, V.; Lakatosova, S.; Bakos, J.; Vlkova, B.; Babinska, K.; Ostatnikova, D. Gastrointestinal microbiota in children with autism in Slovakia. Physiol. Behav. 2015, 138, 179–187. [Google Scholar] [CrossRef]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Low relative abundances of the mucolytic bacterium Akkermansia muciniphila and Bifidobacterium spp. in feces of children with autism. Appl. Environ. Microbiol. 2011, 77, 6718–6721. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.W.; Park, J.G.; Ilhan, Z.E.; Wallstrom, G.; Labaer, J.; Adams, J.B.; Krajmalnik-Brown, R. Reduced incidence of Prevotella and other fermenters in intestinal microflora of autistic children. PLoS ONE 2013, 8, e68322. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Liu, C.; Finegold, S.M. Real-time PCR quantitation of clostridia in feces of autistic children. Appl. Environ. Microbiol. 2004, 70, 6459–6465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Ma, W.; Zhang, J.; He, Y.; Wang, J. Analysis of gut microbiota profiles and microbe-disease associations in children with autism spectrum disorders in China. Sci. Rep. 2018, 8, 13981. [Google Scholar] [CrossRef] [PubMed]
- Son, J.S.; Zheng, L.J.; Rowehl, L.M.; Tian, X.; Zhang, Y.; Zhu, W.; Litcher-Kelly, L.; Gadow, K.D.; Gathungu, G.; Robertson, C.E.; et al. Comparison of Fecal Microbiota in Children with Autism Spectrum Disorders and Neurotypical Siblings in the Simons Simplex Collection. PLoS ONE 2015, 10, e0137725. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Christophersen, C.T.; Sorich, M.J.; Gerber, J.P.; Angley, M.T.; Conlon, M.A. Increased abundance of Sutterella spp. and Ruminococcus torques in feces of children with autism spectrum disorder. Mol. Autism. 2013, 4, 42. [Google Scholar] [CrossRef]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar]
- Appleton, J. The Gut-Brain Axis: Influence of Microbiota on Mood and Mental Health. Integr. Med. (Encinitas) 2018, 17, 28–32. [Google Scholar]
- Berthoud, H.R.; Neuhuber, W.L. Functional and chemical anatomy of the afferent vagal system. Auton. Neurosci. 2000, 85, 1–17. [Google Scholar] [CrossRef]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Goehler, L.E.; Gaykema, R.P.; Opitz, N.; Reddaway, R.; Badr, N.; Lyte, M. Activation in vagal afferents and central autonomic pathways: Early responses to intestinal infection with Campylobacter jejuni. Brain Behav. Immun. 2005, 19, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Barandouzi, Z.A.; Lee, J.; Del Carmen Rosas, M.; Chen, J.; Henderson, W.A.; Starkweather, A.R.; Cong, X.S. Associations of neurotransmitters and the gut microbiome with emotional distress in mixed type of irritable bowel syndrome. Sci. Rep. 2022, 12, 1648. [Google Scholar] [CrossRef] [PubMed]
- McVey Neufeld, K.A.; Mao, Y.K.; Bienenstock, J.; Foster, J.A.; Kunze, W.A. The microbiome is essential for normal gut intrinsic primary afferent neuron excitability in the mouse. Neurogastroenterol. Motil. 2013, 25, 183-e88. [Google Scholar] [CrossRef]
- Reigstad, C.S.; Salmonson, C.E.; Rainey, J.F., 3rd; Szurszewski, J.H.; Linden, D.R.; Sonnenburg, J.L.; Farrugia, G.; Kashyap, P.C. Gut microbes promote colonic serotonin production through an effect of short-chain fatty acids on enterochromaffin cells. Faseb. J. 2015, 29, 1395–1403. [Google Scholar] [CrossRef]
- Wehrwein, E.A.; Orer, H.S.; Barman, S.M. Overview of the Anatomy, Physiology, and Pharmacology of the Autonomic Nervous System. Compr. Physiol. 2016, 6, 1239–1278. [Google Scholar]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef]
- Falomir-Lockhart, L.J.; Cavazzutti, G.F.; Giménez, E.; Toscani, A.M. Fatty Acid Signaling Mechanisms in Neural Cells: Fatty Acid Receptors. Front. Cell Neurosci. 2019, 13, 162. [Google Scholar] [CrossRef]
- Hara, T.; Kimura, I.; Inoue, D.; Ichimura, A.; Hirasawa, A. Free fatty acid receptors and their role in regulation of energy metabolism. Rev. Physiol. Biochem. Pharmacol. 2013, 164, 77–116. [Google Scholar]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef]
- Stilling, R.M.; van de Wouw, M.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. The neuropharmacology of butyrate: The bread and butter of the microbiota-gut-brain axis? Neurochem. Int. 2016, 99, 110–132. [Google Scholar] [CrossRef]
- Ang, Z.; Ding, J.L. GPR41 and GPR43 in Obesity and Inflammation - Protective or Causative? Front. Immunol. 2016, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Qian, X.H.; Xie, R.Y.; Liu, X.L.; Chen, S.D.; Tang, H.D. Mechanisms of Short-Chain Fatty Acids Derived from Gut Microbiota in Alzheimer’s Disease. Aging Dis. 2022, 13, 1252–1266. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin A attenuate nuclear factor kappaB activation and tumor necrosis factor alpha secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef]
- Sharon, G.; Cruz, N.J.; Kang, D.W.; Gandal, M.J.; Wang, B.; Kim, Y.M.; Zink, E.M.; Casey, C.P.; Taylor, B.C.; Lane, C.J.; et al. Human Gut Microbiota from Autism Spectrum Disorder Promote Behavioral Symptoms in Mice. Cell 2019, 177, 1600–1618.e17. [Google Scholar] [CrossRef]
- Chang, Y.C.; Cole, T.B.; Costa, L.G. Behavioral Phenotyping for Autism Spectrum Disorders in Mice. Curr. Protoc. Toxicol. 2017, 72, 11.22.1–11.22.21. [Google Scholar] [CrossRef]
- Halladay, A.K.; Bishop, S.; Constantino, J.N.; Daniels, A.M.; Koenig, K.; Palmer, K.; Messinger, D.; Pelphrey, K.; Sanders, S.J.; Singer, A.T.; et al. Sex and gender differences in autism spectrum disorder: Summarizing evidence gaps and identifying emerging areas of priority. Mol. Autism 2015, 6, 36. [Google Scholar] [CrossRef]
- MacFabe, D.F.; Cain, D.P.; Rodriguez-Capote, K.; Franklin, A.E.; Hoffman, J.E.; Boon, F.; Taylor, A.R.; Kavaliers, M.; Ossenkopp, K.P. Neurobiological effects of intraventricular propionic acid in rats: Possible role of short chain fatty acids on the pathogenesis and characteristics of autism spectrum disorders. Behav. Brain Res. 2007, 176, 149–169. [Google Scholar] [CrossRef]
- MacFabe, D.F.; Cain, N.E.; Boon, F.; Ossenkopp, K.P.; Cain, D.P. Effects of the enteric bacterial metabolic product propionic acid on object-directed behavior, social behavior, cognition, and neuroinflammation in adolescent rats: Relevance to autism spectrum disorder. Behav. Brain Res. 2011, 217, 47–54. [Google Scholar] [CrossRef] [PubMed]
- El-Ansary, A.; Bhat, R.S.; Al-Daihan, S.; Al Dbass, A.M. The neurotoxic effects of ampicillin-associated gut bacterial imbalances compared to those of orally administered propionic acid in the etiology of persistent autistic features in rat pups: Effects of various dietary regimens. Gut Pathog. 2015, 7, 7. [Google Scholar] [CrossRef]
- El-Ansary, A.; Bacha, A.B.; Bjørklund, G.; Al-Orf, N.; Bhat, R.S.; Moubayed, N.; Abed, K. Probiotic treatment reduces the autistic-like excitation/inhibition imbalance in juvenile hamsters induced by orally administered propionic acid and clindamycin. Metab. Brain Dis. 2018, 33, 1155–1164. [Google Scholar] [CrossRef]
- Lobzhanidze, G.; Lordkipanidze, T.; Zhvania, M.; Japaridze, N.; MacFabe, D.F.; Pochkidze, N.; Gasimov, E.; Rzaev, F. Effect of propionic acid on the morphology of the amygdala in adolescent male rats and their behavior. Micron 2019, 125, 102732. [Google Scholar] [CrossRef]
- Foley, K.A.; Ossenkopp, K.P.; Kavaliers, M.; Macfabe, D.F. Pre- and neonatal exposure to lipopolysaccharide or the enteric metabolite, propionic acid, alters development and behavior in adolescent rats in a sexually dimorphic manner. PLoS ONE 2014, 9, e87072. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Jin, X.; Guo, P.; Sun, Y.; Zhou, L.; Qing, Y.; Shen, W.; Ji, G. Efficacy of Faecal Microbiota Transplantation for the Treatment of Autism in Children: Meta-Analysis of Randomised Controlled Trials. Evid. Based Complement. Alternat Med. 2023, 2023, 5993628. [Google Scholar] [CrossRef] [PubMed]
- Meeking, M.M.; MacFabe, D.F.; Mepham, J.R.; Foley, K.A.; Tichenoff, L.J.; Boon, F.H.; Kavaliers, M.; Ossenkopp, K.P. Propionic acid induced behavioural effects of relevance to autism spectrum disorder evaluated in the hole board test with rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 97, 109794. [Google Scholar] [CrossRef]
- de Theije, C.G.; Wopereis, H.; Ramadan, M.; van Eijndthoven, T.; Lambert, J.; Knol, J.; Garssen, J.; Kraneveld, A.D.; Oozeer, R. Altered gut microbiota and activity in a murine model of autism spectrum disorders. Brain Behav. Immun. 2014, 37, 197–206. [Google Scholar] [CrossRef]
- Morais, L.H.; Schreiber, H.L.t.; Mazmanian, S.K. The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 2021, 19, 241–255. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Zhu, H.; Li, X.; Shi, W.; Guo, Y.; Du, X.; Zhang, L.; Su, L.; Qin, C. Comparative Metagenomics and Metabolomes Reveals Abnormal Metabolism Activity Is Associated with Gut Microbiota in Alzheimer’s Disease Mice. Int. J. Mol. Sci. 2022, 23, 11560. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Huh, J.R.; Shah, K. Microbiota and the gut-brain-axis: Implications for new therapeutic design in the CNS. EBioMedicine 2022, 77, 103908. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lagod, P.P.; Naser, S.A. The Role of Short-Chain Fatty Acids and Altered Microbiota Composition in Autism Spectrum Disorder: A Comprehensive Literature Review. Int. J. Mol. Sci. 2023, 24, 17432. https://doi.org/10.3390/ijms242417432
Lagod PP, Naser SA. The Role of Short-Chain Fatty Acids and Altered Microbiota Composition in Autism Spectrum Disorder: A Comprehensive Literature Review. International Journal of Molecular Sciences. 2023; 24(24):17432. https://doi.org/10.3390/ijms242417432
Chicago/Turabian StyleLagod, Piotr P., and Saleh A. Naser. 2023. "The Role of Short-Chain Fatty Acids and Altered Microbiota Composition in Autism Spectrum Disorder: A Comprehensive Literature Review" International Journal of Molecular Sciences 24, no. 24: 17432. https://doi.org/10.3390/ijms242417432