

CRISPR/Cas9-Mediated Knock-Out of the MtCLE35 Gene Highlights Its Key Role in the Control of Symbiotic Nodule Numbers under High-Nitrate Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
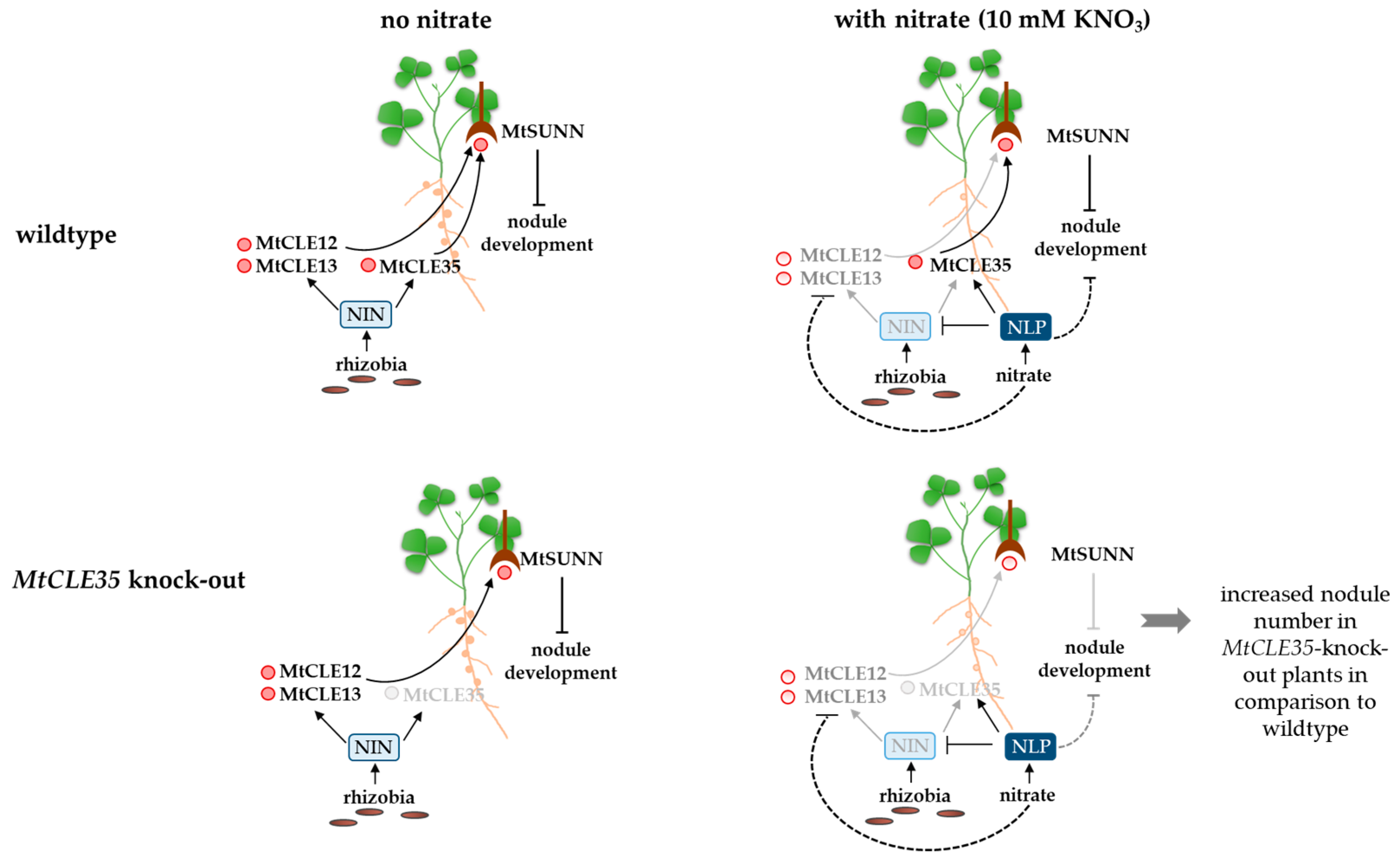
:1. Introduction
2. Results
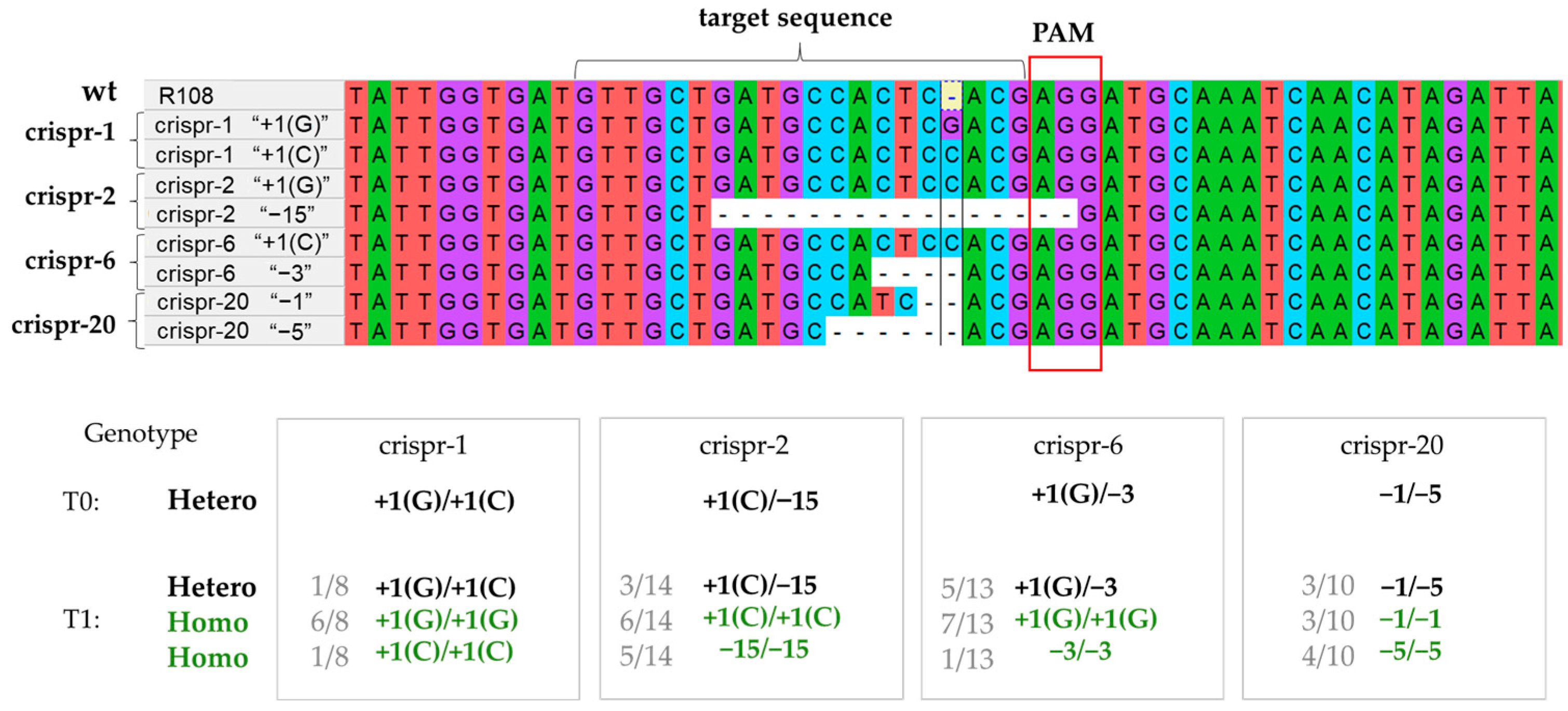
2.1. Generation of Medicago Truncatula Lines through the CRISPR/Cas9-Mediated Knock-Out of the MtCLE35 Gene
2.1.1. Construction of the Vector for the CRISPR/Cas9-Mediated Editing of the MtCLE35 Gene and Stable Transformation of Medicago truncatula Plants
2.1.2. Genotyping of T0–T1 Plants
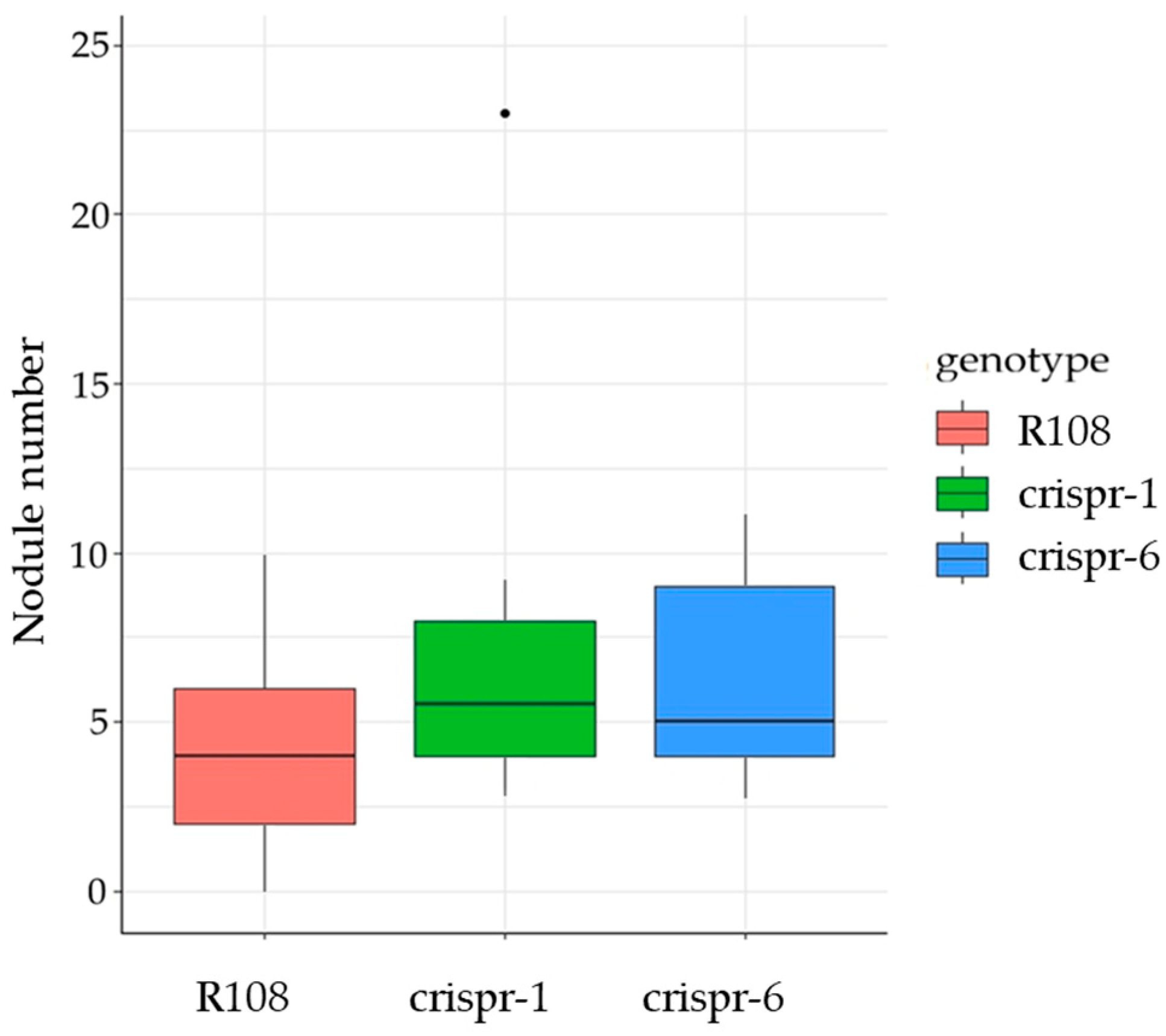
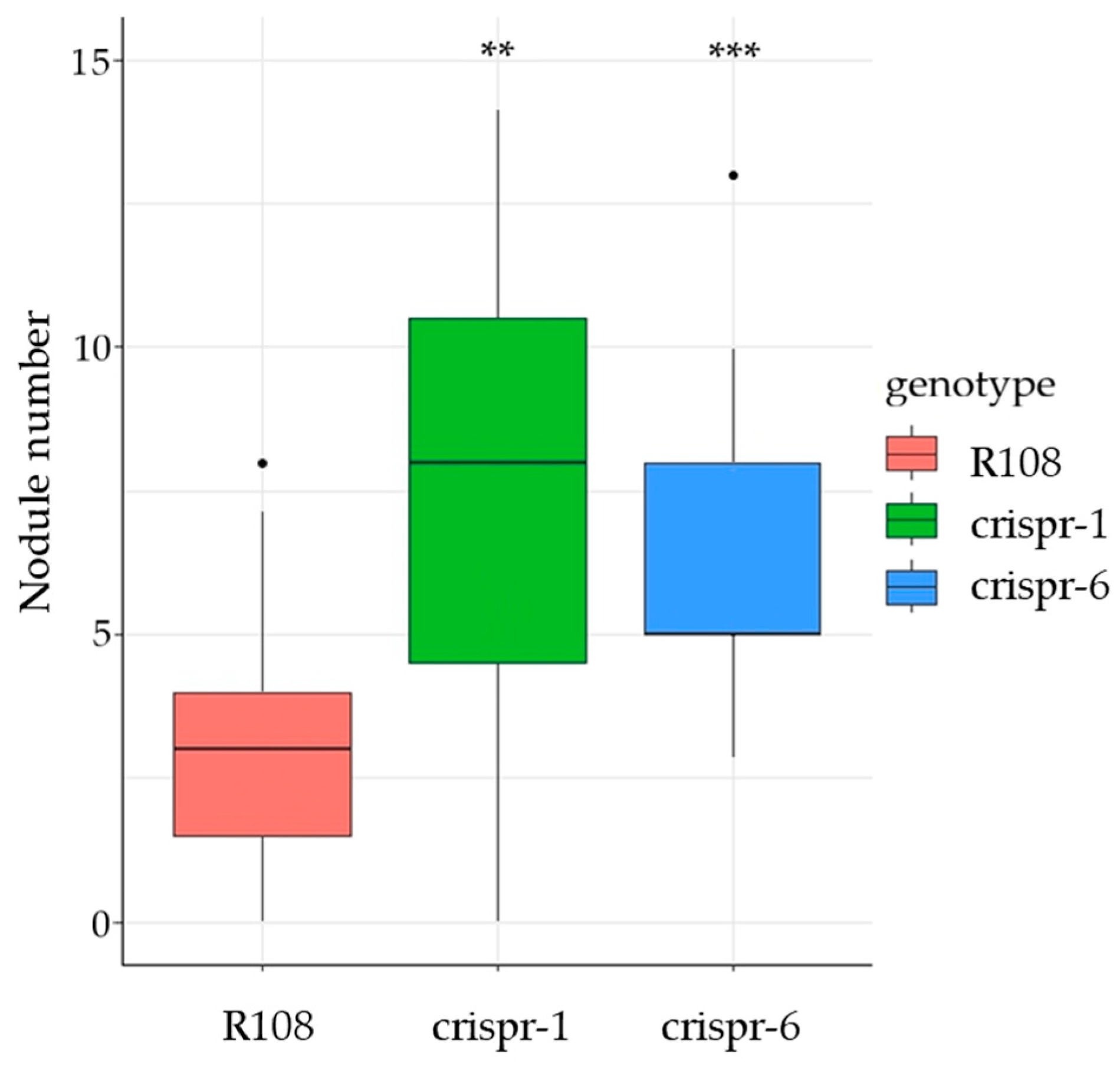
2.2. Effect of MtCLE35 Knock-Out on Nodule Number under Nitrogen-Free and Nitrate Treatment Conditions
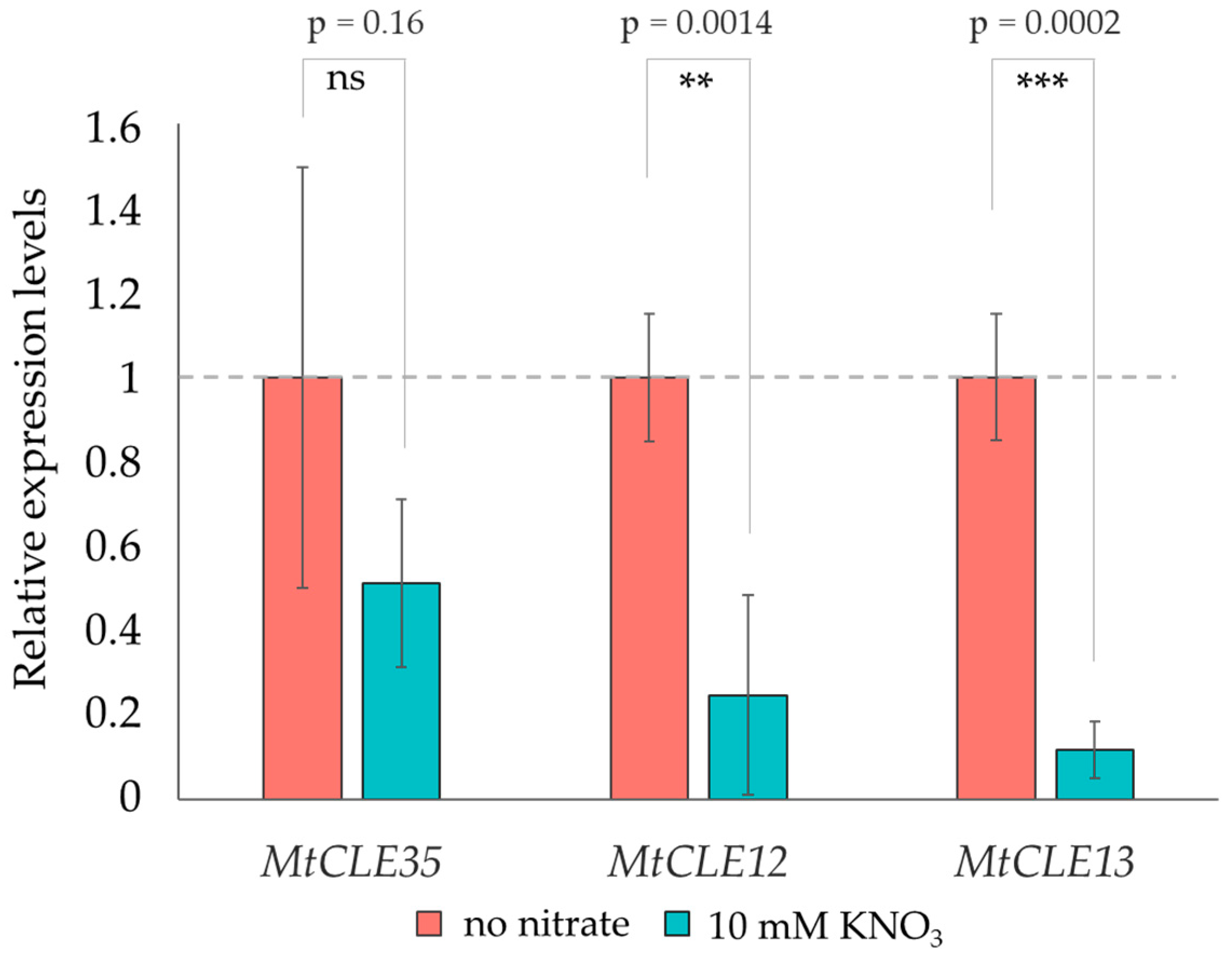
2.3. Effect of Nitrate on the Expression Levels of MtCLE Genes in Rhizobia-Inoculated Roots
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Construction of the Vector for the CRISPR/Cas9-Mediated Knock-out of the MtCLE35 Gene
4.3. Agrobacterium Tumefaciens-Mediated Transformation
4.4. Genotyping of T0–T1 Plants
4.5. RNA Extraction and Quantitative Reverse Transcription PCR (qRT-PCR) Analysis
4.6. Statistical Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Pei, Y.; Shen, Y.; Zhang, R.; Kang, M.; Ma, Y.; Li, D.; Chen, Y. Progress in the Self-Regulation System in Legume Nodule Development-AON (Autoregulation of Nodulation). Int. J. Mol. Sci. 2022, 23, 6676. [Google Scholar] [CrossRef] [PubMed]
- Searle, I.R.; Men, A.E.; Laniya, T.S.; Buzas, D.M.; Iturbe-Ormaetxe, I.; Carroll, B.J.; Gresshoff, P.M. Long-Distance Signaling in Nodulation Directed by a CLAVATA1-like Receptor Kinase. Science 2003, 299, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, R.; Hayashi, M.; Wu, G.-J.; Kouchi, H.; Imaizumi-Anraku, H.; Murakami, Y.; Kawasaki, S.; Akao, S.; Ohmori, M.; Nagasawa, M.; et al. HAR1 Mediates Systemic Regulation of Symbiotic Organ Development. Nature 2002, 420, 426–429. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, S.; Ohnishi, E.; Sato, S.; Takahashi, H.; Nakazono, M.; Tabata, S.; Kawaguchi, M. Nod Factor/Nitrate-Induced CLE Genes that Drive HAR1-Mediated Systemic Regulation of Nodulation. Plant Cell Physiol. 2009, 50, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Reid, D.E.; Ferguson, B.J.; Hayashi, S.; Lin, Y.-H.; Gresshoff, P.M. Molecular Mechanisms Controlling Legume Autoregulation of Nodulation. Ann. Bot. 2011, 108, 789–795. [Google Scholar] [CrossRef] [PubMed]
- Mortier, V.; Den Herder, G.; Whitford, R.; Van de Velde, W.; Rombauts, S.; D’haeseleer, K.; Holsters, M.; Goormachtig, S. CLE Peptides Control Medicago Truncatula Nodulation Locally and Systemically. Plant Physiol. 2010, 153, 222–237. [Google Scholar] [CrossRef]
- Lebedeva, M.; Azarakhsh, M.; Yashenkova, Y.; Lutova, L. Nitrate-Induced CLE Peptide Systemically Inhibits Nodulation in Medicago Truncatula. Plants 2020, 9, 1456. [Google Scholar] [CrossRef] [PubMed]
- Mens, C.; Hastwell, A.H.; Su, H.; Gresshoff, P.M.; Mathesius, U.; Ferguson, B.J. Characterisation of Medicago Truncatula CLE34 and CLE35 in Nitrate and Rhizobia Regulation of Nodulation. New Phytol. 2021, 229, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Moreau, C.; Gautrat, P.; Frugier, F. Nitrate-Induced CLE35 Signaling Peptides Inhibit Nodulation through the SUNN Receptor and miR2111 Repression. Plant Physiol. 2021, 185, 1216–1228. [Google Scholar] [CrossRef] [PubMed]
- Soyano, T.; Hirakawa, H.; Sato, S.; Hayashi, M.; Kawaguchi, M. NODULE INCEPTION Creates a Long-Distance Negative Feedback Loop Involved in Homeostatic Regulation of Nodule Organ Production. Proc. Natl. Acad. Sci. USA 2014, 111, 14607–14612. [Google Scholar] [CrossRef] [PubMed]
- Laffont, C.; Ivanovici, A.; Gautrat, P.; Brault, M.; Djordjevic, M.A.; Frugier, F. The NIN Transcription Factor Coordinates CEP and CLE Signaling Peptides That Regulate Nodulation Antagonistically. Nat. Commun. 2020, 11, 3167. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Lin, J.; Zhu, Y.; Fu, M.; Li, X.; Xie, F. NLP1 Reciprocally Regulates Nitrate Inhibition of Nodulation through SUNN-CRA2 Signaling in Medicago Truncatula. Plant Commun. 2021, 2, 100183. [Google Scholar] [CrossRef] [PubMed]
- Cosson, V.; Durand, P.; d’Erfurth, I.; Kondorosi, A.; Ratet, P. Medicago Truncatula Transformation Using Leaf Explants. In Agrobacterium Protocols; Wang, K., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2006; pp. 115–128. ISBN 978-1-59745-130-7. [Google Scholar]
- Lin, J.; Li, X.; Luo, Z.; Mysore, K.S.; Wen, J.; Xie, F. NIN Interacts with NLPs to Mediate Nitrate Inhibition of Nodulation in Medicago Truncatula. Nat. Plants 2018, 4, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Fahraeus, G. The Infection of Clover Root Hairs by Nodule Bacteria Studied by a Simple Glass Slide Technique. J. Gen. Microbiol. 1957, 16, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Lullien, V.; Barker, D.G.; de Lajudie, P.; Huguet, T. Plant Gene Expression in Effective and Ineffective Root Nodules of Alfalfa (Medicago Sativa). Plant Mol. Biol. 1987, 9, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Concordet, J.-P.; Haeussler, M. CRISPOR: Intuitive Guide Selection for CRISPR/Cas9 Genome Editing Experiments and Screens. Nucleic Acids Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Cas-OFFinder: A Fast and Versatile Algorithm that Searches for Potential Off-Target Sites of Cas9 RNA-Guided Endonucleases|Bioinformatics|Oxford Academic. Available online: https://academic.oup.com/bioinformatics/article/30/10/1473/267560 (accessed on 13 October 2023).
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 Toolkit for Multiplex Genome Editing in Plants. BMC Plant Biol. 2014, 14, 327. [Google Scholar] [CrossRef] [PubMed]
- Tvorogova, V.E.; Fedorova, Y.A.; Potsenkovskaya, E.A.; Kudriashov, A.A.; Efremova, E.P.; Kvitkovskaya, V.A.; Wolabu, T.W.; Zhang, F.; Tadege, M.; Lutova, L.A. The WUSCHEL-Related Homeobox Transcription Factor MtWOX9-1 Stimulates Somatic Embryogenesis in Medicago Truncatula. Plant Cell Tissue Organ. Cult. 2019, 138, 517–527. [Google Scholar] [CrossRef]
- Edwards, A.; Civitello, A.; Hammond, H.A.; Caskey, C.T. DNA Typing and Genetic Mapping with Trimeric and Tetrameric Tandem Repeats. Am. J. Hum. Genet. 1991, 49, 746–756. [Google Scholar] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lebedeva, M.A.; Dobychkina, D.A.; Lutova, L.A. CRISPR/Cas9-Mediated Knock-Out of the MtCLE35 Gene Highlights Its Key Role in the Control of Symbiotic Nodule Numbers under High-Nitrate Conditions. Int. J. Mol. Sci. 2023, 24, 16816. https://doi.org/10.3390/ijms242316816
Lebedeva MA, Dobychkina DA, Lutova LA. CRISPR/Cas9-Mediated Knock-Out of the MtCLE35 Gene Highlights Its Key Role in the Control of Symbiotic Nodule Numbers under High-Nitrate Conditions. International Journal of Molecular Sciences. 2023; 24(23):16816. https://doi.org/10.3390/ijms242316816
Chicago/Turabian StyleLebedeva, Maria A., Daria A. Dobychkina, and Lyudmila A. Lutova. 2023. "CRISPR/Cas9-Mediated Knock-Out of the MtCLE35 Gene Highlights Its Key Role in the Control of Symbiotic Nodule Numbers under High-Nitrate Conditions" International Journal of Molecular Sciences 24, no. 23: 16816. https://doi.org/10.3390/ijms242316816