
Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars
Abstract
:1. Introduction
2. Results
2.1. Robust Hot Pepper Protoplasts Isolated from Six Commercial Cultivars
2.2. The Conserved CaMLO2 Gene among Six Commercial Hot Peppers
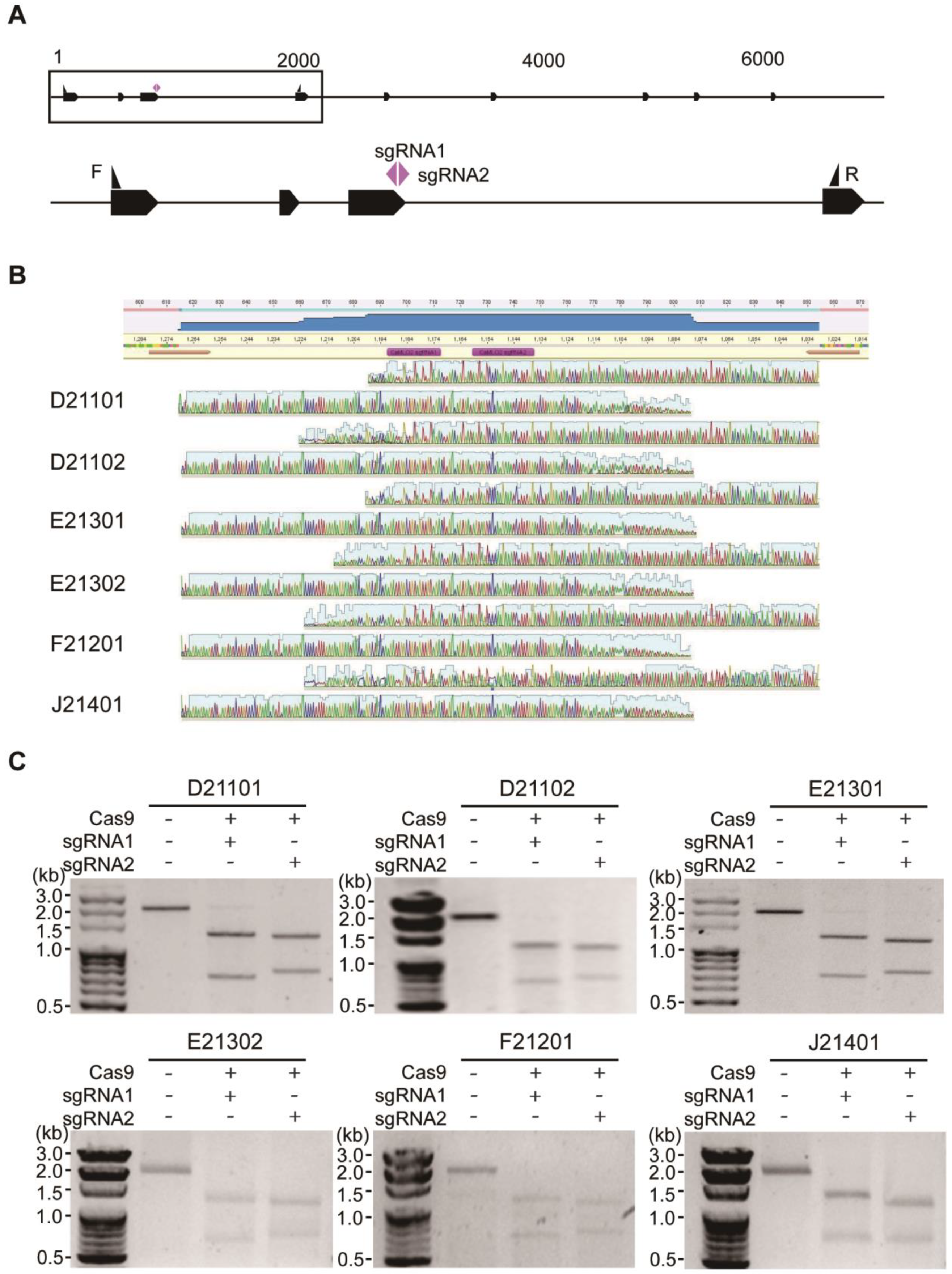
2.3. Comparison of Indel Frequencies of CaMLO2sgRNA1 or sgRNA2 among the Six Commercial Hot Peppers
2.4. Comparison of the Indel Patterns by CaMLO2sgRNA1 or sgRNA2 among the Six Commercial Hot Peppers
2.5. In Vivo Off-Taget Validation of the CaMLO2sgRNA1 Delivered across the Six Cultivars
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. CaMLO2 Genomic Loci Analyses by Sanger Sequencing
4.3. Preparation of the Single-Guide RNA and Cas9 Protein
4.4. In Vitro Cleavage Analysis Using the Designed CRISPR/Cas9 RNP
4.5. Protoplast Isolation and PEG-Mediated Transfection
4.6. Targeted Deep Sequencing
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pickersgill, B. Genetic resources and breeding of Capsicum spp. Euphytica. Euphytica 1997, 96, 129–133. [Google Scholar] [CrossRef]
- Juroszek, P.; Von Tiedemann, A. Plant pathogens, insect pests and weeds in a changing global climate: A review of approaches, challenges, research gaps, key studies and concepts. J. Agric. Sci. 2013, 151, 163–188. [Google Scholar] [CrossRef]
- Ashtaputre, S. Assessment of yield loss due to powdery mildew of chilli. Trends Biosci. 2014, 7, 1138–1141. [Google Scholar]
- Pietrusińska, A.; Tratwal, A. Characteristics of powdery mildew and its importance for wheat grown in Poland. Plant Prot. Sci. 2020, 56, 141–153. [Google Scholar] [CrossRef]
- Kim, H.; Choi, J.; Won, K.-H. A stable DNA-free screening system for CRISPR/RNPs-mediated gene editing in hot and sweet cultivars of Capsicum annuum. BMC Plant Biol. 2020, 20, 449. [Google Scholar] [CrossRef]
- Park, S.; Kim, H.-B.; Jeon, H.-J.; Kim, H. Agrobacterium-mediated Capsicum annuum gene editing in two cultivars, hot pepper CM334 and bell pepper Dempsey. Int. J. Mol. Sci. 2021, 22, 3921. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Jain, M. The CRISPR–Cas system for plant genome editing: Advances and opportunities. J. Exp. Bot. 2015, 66, 47–57. [Google Scholar] [CrossRef]
- Weeks, D.P.; Spalding, M.H.; Yang, B. Use of designer nucleases for targeted gene and genome editing in plants. Plant Biotechnol. J. 2016, 14, 483–495. [Google Scholar] [CrossRef]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.-S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232. [Google Scholar] [CrossRef]
- Gratz, S.J.; Cummings, A.M.; Nguyen, J.N.; Hamm, D.C.; Donohue, L.K.; Harrison, M.M.; Wildonger, J.; O’Connor-Giles, K.M. Genome engineering of Drosophila with the CRISPR RNA-guided Cas9 nuclease. Genetics 2013, 194, 1029–1035. [Google Scholar] [CrossRef]
- Hruscha, A.; Krawitz, P.; Rechenberg, A.; Heinrich, V.; Hecht, J.; Haass, C.; Schmid, B. Efficient CRISPR/Cas9 genome editing with low off-target effects in Zebrafish. Development 2013, 140, 4982–4987. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, H.; Bi, H.; Fromm, M.; Yang, B.; Weeks, D.P. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, Tobacco, Sorghum and Rice. Nucleic Acids Res. 2013, 41, e188. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-F.; Norville, J.E.; Aach, J.; McCormack, M.; Zhang, D.; Bush, J.; Church, G.M.; Sheen, J. Multiplex and homologous recombination–mediated genome editing in Arabidopsis and Nicotiana Benthamiana using guide RNA and Cas9. Nat. Biotechnol. 2013, 31, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yang, H.; Shivalila, C.S.; Dawlaty, M.M.; Cheng, A.W.; Zhang, F.; Jaenisch, R. One-step generation of mice carrying mutations in multiple genes by CRISPR/Cas-mediated genome engineering. Cell 2013, 153, 910–918. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for crop improvement: An update review. Front. Plant. Sci. 2018, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Oliva, R.; Ji, C.; Atienza-Grande, G.; Huguet-Tapia, J.C.; Perez-Quintero, A.; Li, T.; Eom, J.-S.; Li, C.; Nguyen, H.; Liu, B.; et al. Broad-spectrum resistance to bacterial blight in rice using genome editing. Nat. Biotechnol. 2019, 37, 1344–1350. [Google Scholar] [CrossRef] [PubMed]
- Kieu, N.P.; Lenman, M.; Wang, E.S.; Petersen, B.L.; Andreasson, E. Mutations introduced in susceptibility genes through CRISPR/Cas9 genome editing confer increased late blight resistance in potatoes. Sci. Rep. 2021, 11, 4487. [Google Scholar] [CrossRef]
- Chaudhary, R.; Atamian, H.S. Resistance-gene-mediated defense responses against biotic stresses in the crop model plant tomato. J. Plant Pathol. Microbiol. 2017, 8, 404. [Google Scholar] [CrossRef]
- Marx, V. Plants: A tool box of cell-based assays. Nat. Methods 2016, 13, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Reed, K.M.; Bargmann, B.O.R. Protoplast regeneration and its use in new plant breeding technologies. Front. Genome Ed. 2021, 3, 734951. [Google Scholar] [CrossRef] [PubMed]
- Galun, E. Plant protoplasts as physiological tools. Annu. Rev. Plant Physiol. 1981, 32, 237–266. [Google Scholar] [CrossRef]
- Slocum, R.D.; Kaur-Sawhney, R.; Galston, A.W. The physiology and biochemistry of polyamines in plants. Arch. Biochem. Biophys. 1984, 235, 283–303. [Google Scholar] [CrossRef] [PubMed]
- Ueno, K.; Kinoshita, T.; Inoue, S.; Emi, T.; Shimazaki, K. Biochemical characterization of plasma membrane H+-ATPase activation in guard cell protoplasts of Arabidopsis thaliana in response to blue light. Plant Cell Physiol. 2005, 46, 955–963. [Google Scholar] [CrossRef]
- Eeckhaut, T.; Lakshmanan, P.S.; Deryckere, D.; Van Bockstaele, E.; Van Huylenbroeck, J. Progress in plant protoplast research. Planta 2013, 238, 991–1003. [Google Scholar] [CrossRef]
- Woo, J.W.; Kim, J.; Kwon, I.S.; Corvalán, C.; Cho, S.W.; Kim, H.; Kim, S.-G.; Kim, S.-T.; Choe, S.; Kim, J.-S. DNA-free genome editing in plants with preassembled CRISPR-Cas9 ribonucleoproteins. Nat. Biotechnol. 2015, 33, 1162–1164. [Google Scholar] [CrossRef]
- Power, J.B.; Cummins, S.E.; Cocking, E.C. Fusion of isolated plant protoplasts. Nature 1970, 225, 1016–1018. [Google Scholar] [CrossRef]
- Svitashev, S.; Schwartz, C.; Lenderts, B.; Young, J.K.; Mark Cigan, A. Genome editing in maize directed by CRISPR–Cas9 ribonucleoprotein complexes. Nat. Commun. 2016, 7, 13274. [Google Scholar] [CrossRef]
- Lee, M.H.; Lee, J.; Choi, S.A.; Kim, Y.-S.; Koo, O.; Choi, S.H.; Ahn, W.S.; Jie, E.Y.; Kim, S.W. Efficient genome editing using CRISPR–Cas9 RNP delivery into cabbage protoplasts via electro-transfection. Plant Biotechnol. Rep. 2020, 14, 695–702. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.-T.; Ryu, J.; Kang, B.-C.; Kim, J.-S.; Kim, S.-G. CRISPR/Cpf1-mediated DNA-free plant genome editing. Nat. Commun. 2017, 8, 14406. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Tu, L.; Subburaj, S.; Bae, S.; Lee, G.-J. Simultaneous targeting of duplicated genes in Petunia protoplasts for flower color modification via CRISPR-Cas9 ribonucleoproteins. Plant Cell Rep. 2021, 40, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Bush, J.; Sheen, J. A versatile and efficient plant protoplast platform for genome editing by Cas9 RNPs. Front. Genome Ed. 2021, 3, 719190. [Google Scholar] [CrossRef] [PubMed]
- Madhavi Reddy, K.; Shivashankara, K.S.; Geetha, G.A.; Pavithra, K.C. Capsicum (Hot Pepper and Bell Pepper). In Abiotic Stress Physiology of Horticultural Crops; Springer: New Delhi, India, 2016; pp. 151–166. [Google Scholar] [CrossRef]
- Devoto, A.; Piffanelli, P.; Nilsson, I.; Wallin, E.; Panstruga, R.; von Heijne, G.; Schulze-Lefert, P. Topology, subcellular localization, and sequence diversity of the MLO family in plants. J. Biol. Chem. 1999, 274, 34993–35004. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.-I.; Kim, Y.-M.; Lee, J.M.; Lee, H.-A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.-T.; et al. Genome sequence of the hot pepper provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Lee, J.-H.; Venkatesh, J.; Jo, J.; Jang, S.; Kim, G.W.; Kim, J.-M.; Han, K.; Ro, N.; Lee, H.-Y.; Kwon, J.-K.; et al. High-quality chromosome-scale genomes facilitate effective identification of large structural variations in hot and sweet peppers. Hortic. Res. 2022, 9, uhac210. [Google Scholar] [CrossRef]
- Büschges, R.; Hollricher, K.; Panstruga, R.; Simons, G.; Wolter, M.; Frijters, A.; van Daelen, R.; van der Lee, T.; Diergaarde, P.; Groenendijk, J.; et al. The barley MLO gene: A novel control element of plant pathogen resistance. Cell 1997, 88, 695–705. [Google Scholar] [CrossRef]
- Consonni, C.; Humphry, M.E.; Hartmann, H.A.; Livaja, M.; Durner, J.; Westphal, L.; Vogel, J.; Lipka, V.; Kemmerling, B.; Schulze-Lefert, P.; et al. Conserved requirement for a plant host cell protein in powdery mildew pathogenesis. Nat. Genet. 2006, 38, 716–720. [Google Scholar] [CrossRef]
- Elliott, C.; Zhou, F.; Spielmeyer, W.; Panstruga, R.; Schulze-Lefert, P. Functional conservation of wheat and rice MLO orthologs in defense modulation to the powdery mildew fungus. Mol. Plant Microbe Interact. 2002, 15, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Hwang, B.K. The pepper MLO Gene, CaMLO2, is involved in the susceptibility cell-death response and bacterial and oomycete proliferation. Plant J. 2012, 72, 843–855. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cheng, X.; Shan, Q.; Zhang, Y.; Liu, J.; Gao, C.; Qiu, J.-L. Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew. Nat. Biotechnol. 2014, 32, 947–951. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lin, D.; Zhang, Y.; Deng, M.; Chen, Y.; Lv, B.; Li, B.; Lei, Y.; Wang, Y.; Zhao, L.; et al. Genome-edited powdery mildew resistance in wheat without growth penalties. Nature 2022, 602, 455–460. [Google Scholar] [CrossRef]
- Díaz, I.; Moreno, R.; Power, J.B. Plant regeneration from protoplasts of Capsicum annuum. Plant Cell Rep. 1988, 7, 210–212. [Google Scholar] [CrossRef]
- Prakash, A.H.; Rao, K.S.; Kumar, M.U. Plant regeneration from protoplasts of Capsicum annuum L. cv. California Wonder. J. Biosci. 1997, 22, 339–344. [Google Scholar] [CrossRef]
- Won, K.-H.; Park, S.; Choi, J.; Kim, H.H.; Kang, B.-C.; Kim, H. A reliable regeneration method in genome-editable bell pepper ‘Dempsey’. Horticulturae 2021, 7, 317. [Google Scholar] [CrossRef]
- Jie, E.Y.; Kim, S.W.; Jang, H.-R.; In, D.-S.; Liu, J.-R. Myo-inositol increases the plating efficiency of protoplast derived from cotyledon of cabbage (Brassica Oleracea var. Capitata). J. Plant Biotechnol. 2011, 38, 69–76. [Google Scholar] [CrossRef]
- Park, J.; Bae, S.; Kim, J.-S. Cas-Designer: A web-based tool for choice of CRISPR-Cas9 target sites. Bioinformatics 2015, 31, 4014–4016. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar Name | Agronomic Trait |
|---|---|
| D21101 | Korean cultivar (KC) for pepper powder, highly pungent, resistant to Phytophthora blight |
| D21102 | KC for pepper powder, highly pungent, resistant to Phytophthora blight, tomato spotted wilt virus (TSWV), and anthracnose |
| E21301 | India cultivar for pepper powder |
| E21302 | Southeast Asian cultivar, upright type, single-podded |
| F21201 | KC for green pepper, highly pungent, resistant to Phytophthora blight |
| J21401 | South American cultivar, jalapeño, resistant to bacterial leaf spot (BLS) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.-H.; Kim, H. Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars. Int. J. Mol. Sci. 2023, 24, 16775. https://doi.org/10.3390/ijms242316775
Park J-H, Kim H. Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars. International Journal of Molecular Sciences. 2023; 24(23):16775. https://doi.org/10.3390/ijms242316775
Chicago/Turabian StylePark, Jae-Hyeong, and Hyeran Kim. 2023. "Harnessing CRISPR/Cas9 for Enhanced Disease Resistance in Hot Peppers: A Comparative Study on CaMLO2-Gene-Editing Efficiency across Six Cultivars" International Journal of Molecular Sciences 24, no. 23: 16775. https://doi.org/10.3390/ijms242316775