
The Molecular Role of Polyamines in Age-Related Diseases: An Update
 and
and
Abstract
:1. Introduction
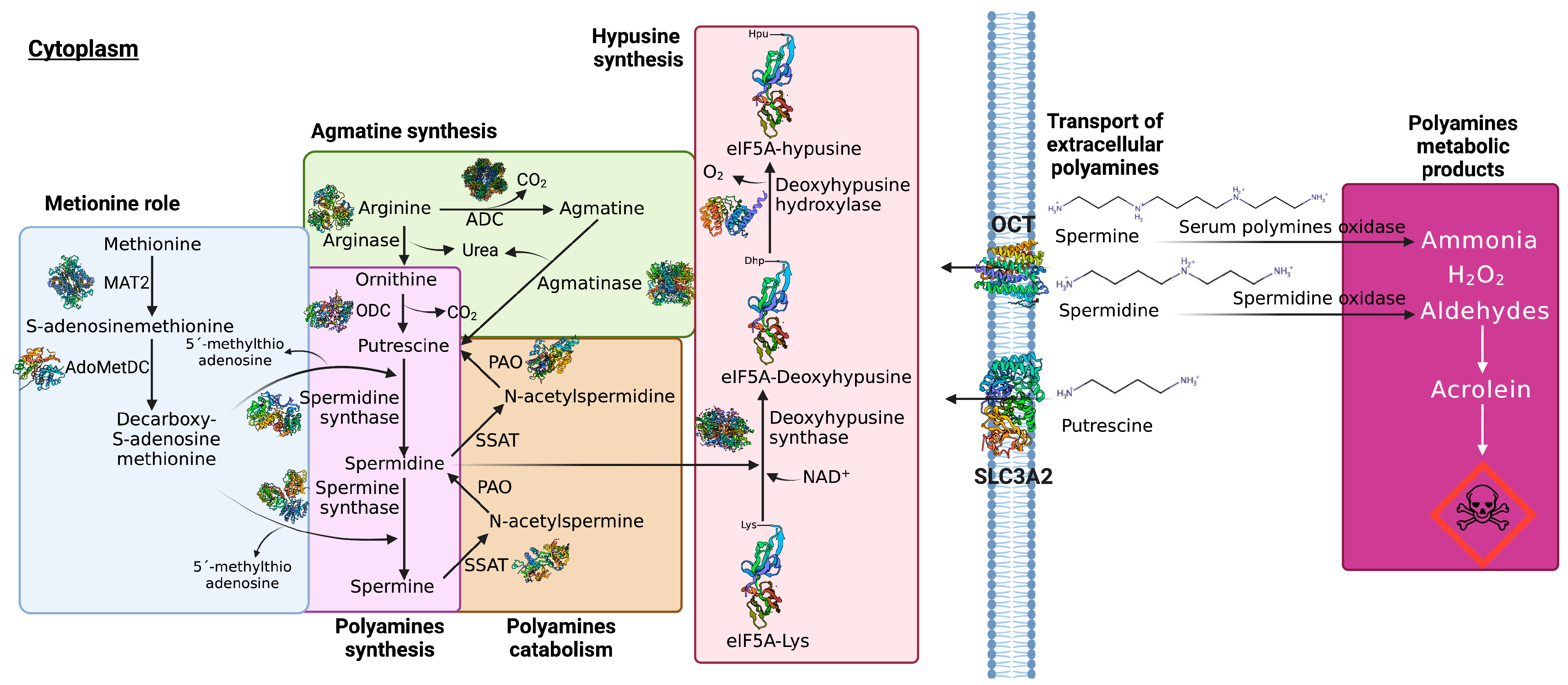
2. Biosynthesis of Pas
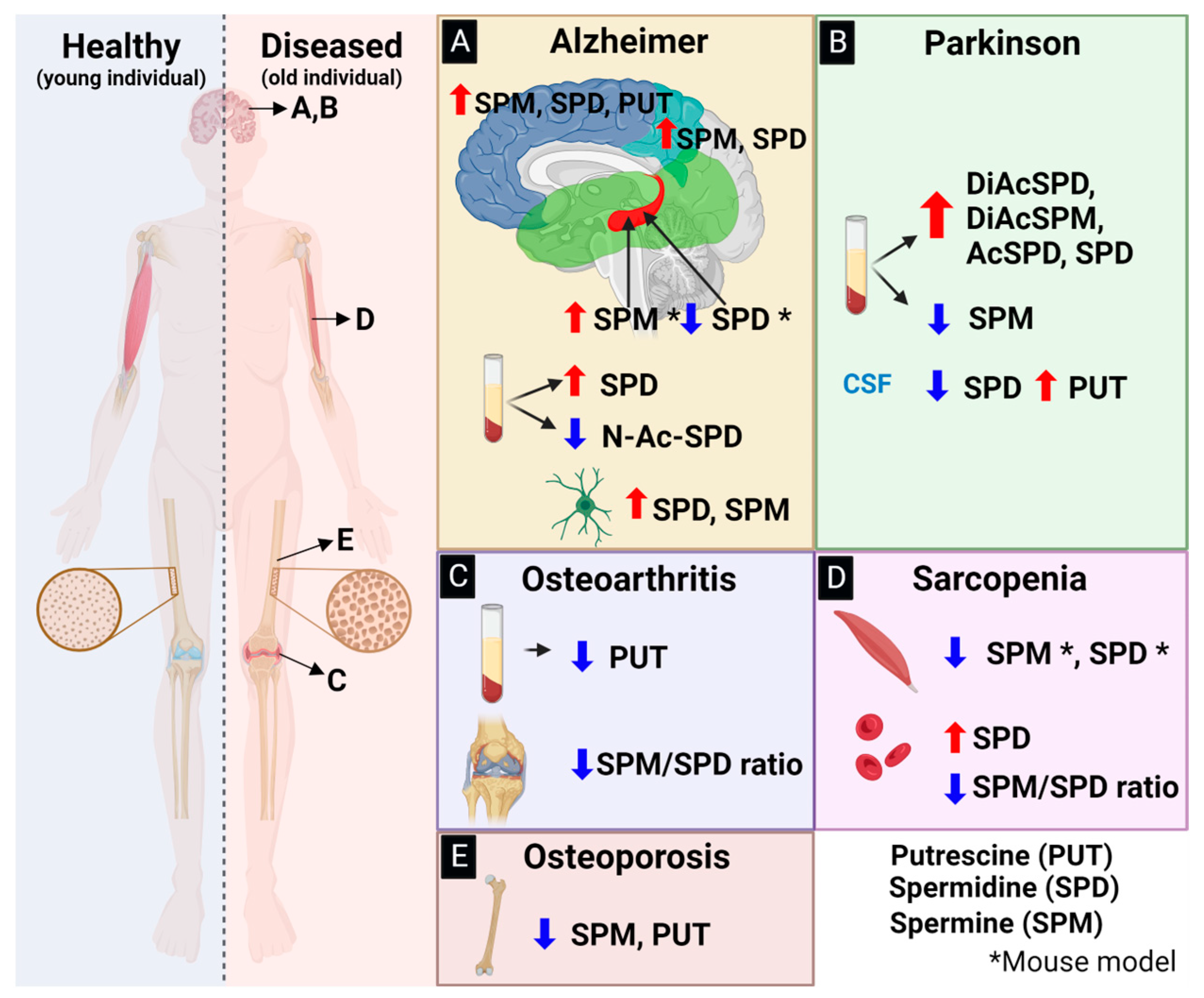
3. Polyamines as Novel Biomarkers for Age-Related Diseases
4. Polyamines and Neurodegenerative Diseases
4.1. The Role of PAs in Alzheimer’s Disease
4.2. The Role of PAs in Parkinson’s Disease
5. Polyamines and the Burden of Chronic-Disease Disability
5.1. The Role of PAs in Osteoporosis
5.2. The Role of PAs in Osteoarthritis
5.3. The Role of PAs in Sarcopenia and Frailty
6. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| α-syn | Alpha-synuclein |
| AD | Alzheimer’s disease |
| ADC | Arginine decarboxylase |
| ATP13A2 | ATPase cation transporting 13A2 |
| BMI | Bone mass index |
| CNS | Central nervous system |
| CSF | Cerebrospinal fluid |
| COL2A1 | Collagen type II alpha 1 chain |
| DHS | Deoxyhypusine synthase |
| DOHH | Deoxyhypusine hydrolase |
| EP300 | E1A-binding protein P300 |
| elF5A | Eukaryotic translation initiation factor 5A |
| GPC1 | Glypican 1 |
| MCI | Mild cognitive impairment |
| MTA | 5ߣ-methylthioadenosin |
| NMDA | N-Methyl-D-aspartate |
| NLRP3 | NLR family pyrin domain containing 3 |
| OCT-1-3 | Organic cation transporters 1, 2, and 3 |
| ODC | Ornithine decarboxylase |
| OA | Osteoarthritis |
| OVX | Ovariectomy |
| PD | Parkinson’s disease |
| PAO | Polyamine oxidase |
| PA | Polyamines |
| PUT | Putrescine |
| PSP | Progressive supranuclear palsy |
| PSR | Polyamine stress response |
| dcAdoMet | S-adenosylmethionine |
| SRS | Snyder–Robinson Syndrome |
| SLC3A2 | Solute Carrier Family 3 (Amino Acid Transporter Heavy Chain) |
| SPD | Spermidine |
| SMS | Spermidine synthase |
| SPM | Spermine |
| SSAT | Spermine/spermidine N1-acetyltransferase |
| SOX9 | SRY-box transcription 9 |
| WHO | World Health Organization |
References
- World Health Organization: Ageing and Health. Available online: https://www.who.int/news-room/fact-sheets/detail/ageing-and-health (accessed on 22 September 2023).
- Tang, B.; Li, Z.; Hu, S.; Xiong, J. Economic Implications of Health Care Burden for Elderly Population. Inq. J. Health Care Organ. Provis. Financ. 2022, 59, 469580221121511. [Google Scholar] [CrossRef] [PubMed]
- Rockwood, K.; Howlett, S.E. Age-related deficit accumulation and the diseases of ageing. Mech. Ageing Dev. 2019, 180, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhang, Z.; Ren, Y.; Wang, Y.; Fang, J.; Yue, H.; Ma, S.; Guan, F. Aging and age-related diseases: From mechanisms to therapeutic strategies. Biogerontology 2021, 22, 165–187. [Google Scholar] [CrossRef] [PubMed]
- GBD 2017 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 354 diseases and injuries for 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, M.; Xie, K.; Schaaf, K.; Bano, D.; Ehninger, D. Targeting the “hallmarks of aging” to slow aging and treat age-related disease: Fact or fiction? Mol. Psychiatry 2023, 28, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Liu, H.; Hu, Q.; Wang, L.; Liu, J.; Zheng, Z.; Zhang, W.; Ren, J.; Zhu, F.; Liu, G.-H. Epigenetic regulation of aging: Implications for interventions of aging and diseases. Signal Transduct. Target. Ther. 2022, 7, 1–22. [Google Scholar] [CrossRef]
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 1–40. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef]
- Bachrach, U. The early history of polyamine research. Plant Physiol. Biochem. 2010, 48, 490–495. [Google Scholar] [CrossRef]
- Schuster, I.; Bernhardt, R. Interactions of natural polyamines with mammalian proteins. Biomol. Concepts 2011, 2, 79–94. [Google Scholar] [CrossRef]
- Muñoz-Esparza, N.C.; Latorre-Moratalla, M.L.; Comas-Basté, O.; Toro-Funes, N.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Polyamines in Food. Front. Nutr. 2019, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Cocchi, S.; Suzzi, G. Polyamines and Gut Microbiota. Front. Nutr. 2019, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Persson, L.; Holm, I.; Stjernborg, L.; Heby, O. Regulation of polyamine synthesis in mammalian cells. Adv. Exp. Med. Biol. 1988, 250, 261–271. [Google Scholar] [PubMed]
- Uemura, T.; Gerner, E.W. Polyamine transport systems in mammalian cells and tissues. Methods Mol. Biol. 2011, 720, 339–348. [Google Scholar]
- Sala-Rabanal, M.; Li, D.C.; Dake, G.R.; Kurata, H.T.; Inyushin, M.; Skatchkov, S.N.; Nichols, C.G. Polyamine Transport by the Polyspecific Organic Cation Transporters OCT1, OCT2, and OCT3. Mol. Pharm. 2013, 10, 1450–1458. [Google Scholar] [CrossRef]
- Li, R.; Wu, X.; Zhu, Z.; Lv, Y.; Zheng, Y.; Lu, H.; Zhou, K.; Wu, D.; Zeng, W.; Dong, W.; et al. Polyamines protect boar sperm from oxidative stress in vitro. J. Anim. Sci. 2022, 100, 9. [Google Scholar] [CrossRef] [PubMed]
- Piletz, J.E.; Aricioglu, F.; Cheng, J.-T.; Fairbanks, C.A.; Gilad, V.H.; Haenisch, B.; Halaris, A.; Hong, S.; Lee, J.E.; Li, J.; et al. Agmatine: Clinical applications after 100 years in translation. Drug Discov. Today 2013, 18, 880–893. [Google Scholar] [CrossRef]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in aging and disease. Aging 2011, 3, 716–732. [Google Scholar] [CrossRef]
- Kim, H.I.; Schultz, C.R.; Chandramouli, G.V.R.; Geerts, D.; Risinger, J.I.; Bachmann, A.S. Pharmacological targeting of polyamine and hypusine biosynthesis reduces tumour activity of endometrial cancer. J. Drug Target. 2022, 30, 623–633. [Google Scholar] [CrossRef]
- Chin, A.; Bieberich, C.J.; Stewart, T.M.; Casero, R.A. Polyamine Depletion Strategies in Cancer: Remodeling the Tumor Immune Microenvironment to Enhance Anti-Tumor Responses. Med. Sci. 2022, 10, 31. [Google Scholar] [CrossRef]
- Malpica-Nieves, C.J.; Rivera-Aponte, D.E.; Tejeda-Bayron, F.A.; Mayor, A.M.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. The involvement of polyamine uptake and synthesis pathways in the proliferation of neonatal astrocytes. Amino Acids 2020, 52, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Cooper, H.L.; Folk, J.E. Identification of hypusine, an unusual amino acid, in a protein from human lymphocytes and of spermidine as its biosynthetic precursor. Proc. Natl. Acad. Sci. USA 1981, 78, 2869–2873. [Google Scholar] [CrossRef]
- Park, M.H.; Wolff, E.C. Hypusine, a polyamine-derived amino acid critical for eukaryotic translation. J. Biol. Chem. 2018, 293, 18710–18718. [Google Scholar] [CrossRef]
- Coleman, C.S.; Stanley, B.A.; Jones, A.D.; Pegg, A.E. Spermidine/spermine-N1-acetyltransferase-2 (SSAT2) acetylates thialysine and is not involved in polyamine metabolism. Biochem. J. 2004, 384, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Salvi, D.; Tavladoraki, P. The tree of life of polyamine oxidases. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Pegg, A.E. Toxicity of Polyamines and Their Metabolic Products. Chem. Res. Toxicol. 2013, 26, 1782–1800. [Google Scholar] [CrossRef]
- Buergel, T.; Steinfeldt, J.; Ruyoga, G.; Pietzner, M.; Bizzarri, D.; Vojinovic, D.; zu Belzen, J.U.; Loock, L.; Kittner, P.; Christmann, L.; et al. Metabolomic profiles predict individual multidisease outcomes. Nat. Med. 2022, 28, 2309–2320. [Google Scholar] [CrossRef]
- Sharma, R.; Ramanathan, A. The Aging Metabolome—Biomarkers to Hub Metabolites. Proteomics 2020, 20, e1800407. [Google Scholar] [CrossRef]
- Tang, J.; Wu, X.; Cheng, B.; Lu, Y. Identification of a polyamine-related signature and six novel prognostic biomarkers in oral squamous cell carcinoma. Front. Mol. Biosci. 2023, 10, 1073770. [Google Scholar] [CrossRef]
- Liu, R.; Lin, X.; Li, Z.; Li, Q.; Bi, K. Quantitative metabolomics for investigating the value of polyamines in the early diagnosis and therapy of colorectal cancer. Oncotarget 2017, 9, 4583–4592. [Google Scholar] [CrossRef]
- Sari, I.N.; Setiawan, T.; Kim, K.S.; Wijaya, Y.T.; Cho, K.W.; Kwon, H.Y. Metabolism and function of polyamines in cancer progression. Cancer Lett. 2021, 519, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Casero, R.A., Jr.; Stewart, T.M.; Pegg, A.E. Polyamine metabolism and cancer: Treatments, challenges and opportunities. Nat. Rev. Cancer 2018, 18, 681–695. [Google Scholar] [CrossRef] [PubMed]
- Sieckmann, T.; Kirschner, K.M. Polyamines, metabolites and metabolomics. Acta Physiol. 2020, 229, 13480. [Google Scholar] [CrossRef]
- Graham, S.F.; Chevallier, O.P.; Elliott, C.T.; Hölscher, C.; Johnston, J.; McGuinness, B.; Kehoe, P.G.; Passmore, A.P.; Green, B.D. Untargeted Metabolomic Analysis of Human Plasma Indicates Differentially Affected Polyamine and L-Arginine Metabolism in Mild Cognitive Impairment Subjects Converting to Alzheimer’s Disease. PLoS ONE 2015, 10, e0119452. [Google Scholar] [CrossRef]
- Sternberg, Z.; Podolsky, R.; Nir, A.; Yu, J.; Nir, R.; Halvorsen, S.W.; Quinn, J.F.; Kaye, J.; Kolb, C. Elevated spermidine serum levels in mild cognitive impairment, a potential biomarker of progression to Alzheimer dementia, a pilot study. J. Clin. Neurosci. 2022, 100, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Saiki, S.; Sasazawa, Y.; Fujimaki, M.; Kamagata, K.; Kaga, N.; Taka, H.; Li, Y.; Souma, S.; Hatano, T.; Imamichi, Y.; et al. A metabolic profile of polyamines in parkinson disease: A promising biomarker. Ann. Neurol. 2019, 86, 251–263. [Google Scholar] [CrossRef]
- Tootsi, K.; Vilba, K.; Märtson, A.; Kals, J.; Paapstel, K.; Zilmer, M. Metabolomic signature of amino acids and polyamines in the serum of osteoarthritis patients. Osteoarthr. Cartil. 2021, 29, S372–S373. [Google Scholar] [CrossRef]
- Tootsi, K.; Vilba, K.; Märtson, A.; Kals, J.; Paapstel, K.; Zilmer, M. Metabolomic Signature of Amino Acids, Biogenic Amines and Lipids in Blood Serum of Patients with Severe Osteoarthritis. Metabolites 2020, 10, 323. [Google Scholar] [CrossRef]
- Sanayama, H.; Ito, K.; Ookawara, S.; Uemura, T.; Sakiyama, Y.; Sugawara, H.; Tabei, K.; Igarashi, K.; Soda, K. Whole Blood Spermine/Spermidine Ratio as a New Indicator of Sarcopenia Status in Older Adults. Biomedicines 2023, 11, 1403. [Google Scholar] [CrossRef]
- Sanayama, H.; Ito, K.; Ookawara, S.; Uemura, T.; Imai, S.; Kiryu, S.; Iguchi, M.; Sakiyama, Y.; Sugawara, H.; Morishita, Y.; et al. Positive Correlation between Relative Concentration of Spermine to Spermidine in Whole Blood and Skeletal Muscle Mass Index: A Possible Indicator of Sarcopenia and Prognosis of Hemodialysis Patients. Biomedicines 2023, 11, 746. [Google Scholar] [CrossRef]
- Lv, H.; Jiang, F.; Guan, D.; Lu, C.; Guo, B.; Chan, C.; Peng, S.; Liu, B.; Guo, W.; Zhu, H.; et al. Metabolomics and Its Application in the Development of Discovering Biomarkers for Osteoporosis Research. Int. J. Mol. Sci. 2016, 17, 2018. [Google Scholar] [CrossRef]
- Qi, H.; Bao, J.; An, G.; Ouyang, G.; Zhang, P.; Wang, C.; Ying, H.; Ouyang, P.; Ma, B.; Zhang, Q. Association between the metabolome and bone mineral density in pre- and post-menopausal Chinese women using GC-MS. Mol. Biosyst. 2016, 12, 2265–2275. [Google Scholar] [CrossRef]
- Kong, S.H.; Kim, J.H.; Shin, C.S. Serum Spermidine as a Novel Potential Predictor for Fragility Fractures. J. Clin. Endocrinol. Metab. 2020, 106, e582–e591. [Google Scholar] [CrossRef]
- Morrison, L.D.; Kish, S.J. Brain polyamine levels are altered in Alzheimer’s disease. Neurosci. Lett. 1995, 197, 5–8. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, U.V.; Varma, V.R.; Griswold, M.E.; Blackshear, C.T.; An, Y.; Oommen, A.M.; Varma, S.; Troncoso, J.C.; Pletnikova, O.; O’Brien, R.; et al. Dysregulation of multiple metabolic networks related to brain transmethylation and polyamine pathways in Alzheimer disease: A targeted metabolomic and transcriptomic study. PLoS Med. 2020, 17, e1003012. [Google Scholar]
- Morrison, L.D.; Cao, X.-C.; Kish, S.J. Ornithine decarboxylase in human brain: Influence of aging, regional distribution, and Alzheimer’s disease. J. Neurochem. 2002, 71, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, S.M.; Atanasova, M.; Dimitrov, I.; Doytchinova, I.A. Cellular polyamines condense hyperphosphorylated Tau, triggering Alzheimer’s disease. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Tsutsui, H.; Akatsu, H.; Hashizume, Y.; Matsukawa, N.; Yamamoto, T.; Toyooka, T. Metabolic profiling of Alzheimer’s disease brains. Sci. Rep. 2013, 3, 2364. [Google Scholar] [CrossRef]
- Sandusky-Beltran, L.A.; Kovalenko, A.; Placides, D.S.; Ratnasamy, K.; Ma, C.; Hunt, J.B.; Liang, H.; Calahatian, J.I.T.; Michalski, C.; Fahnestock, M.; et al. Aberrant AZIN2 and polyamine metabolism precipitates tau neuropathology. J. Clin. Investig. 2021, 131, 26299. [Google Scholar] [CrossRef]
- Trushina, E.; Dutta, T.; Persson, X.-M.T.; Mielke, M.M.; Petersen, R.C. Identification of Altered Metabolic Pathways in Plasma and CSF in Mild Cognitive Impairment and Alzheimer’s Disease Using Metabolomics. PLoS ONE 2013, 8, e63644. [Google Scholar] [CrossRef]
- Lewandowski, N.M.; Ju, S.; Verbitsky, M.; Ross, B.; Geddie, M.L.; Rockenstein, E.; Adame, A.; Muhammad, A.; Vonsattel, J.P.; Ringe, D.; et al. Polyamine pathway contributes to the pathogenesis of Parkinson disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16970–16975. [Google Scholar] [CrossRef] [PubMed]
- Kalecký, K.; Bottiglieri, T. Targeted metabolomic analysis in Parkinson’s disease brain frontal cortex and putamen with relation to cognitive impairment. NPJ Park. Dis. 2023, 9, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Paik, M.J.; Ahn, Y.H.; Lee, P.H.; Kang, H.; Park, C.B.; Choi, S.; Lee, G. Polyamine patterns in the cerebrospinal fluid of patients with Parkinson’s disease and multiple system atrophy. Clin. Chim. Acta 2010, 411, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- LeWitt, P.A.; Li, J.; Wu, K.H.; Lu, M. Diagnostic metabolomic profiling of Parkinson’s disease biospecimens. Neurobiol. Dis. 2023, 177, 105962. [Google Scholar] [CrossRef]
- Gomes-Trolin, C.; Nygren, I.; Aquilonius, S.M.; Askmark, H. Increased red blood cell polyamines in ALS and Parkinson’s disease. Exp. Neurol. 2002, 177, 515–520. [Google Scholar] [CrossRef]
- Roede, J.R.; Uppal, K.; Park, Y.; Lee, K.; Tran, V.; Walker, D.; Strobel, F.H.; Rhodes, S.L.; Ritz, B.; Jones, D.P. Serum metabolomics of slow vs. rapid motor progression Parkinson’s disease: A pilot study. PLoS ONE 2013, 8, e77629. [Google Scholar] [CrossRef]
- Klatt, S.; Doecke, J.D.; Roberts, A.; Boughton, B.A.; Masters, C.L.; Horne, M.; Roberts, B.R. A six-metabolite panel as potential blood-based biomarkers for Parkinson’s disease. NPJ Park. Dis. 2021, 7, 1–14. [Google Scholar] [CrossRef]
- Lin, I.-H.; Chang, J.-L.; Hua, K.; Huang, W.-C.; Hsu, M.-T.; Chen, Y.-F. Skeletal muscle in aged mice reveals extensive transformation of muscle gene expression. BMC Genet. 2018, 19, 55. [Google Scholar] [CrossRef]
- Uchitomi, R.; Hatazawa, Y.; Senoo, N.; Yoshioka, K.; Fujita, M.; Shimizu, T.; Miura, S.; Ono, Y.; Kamei, Y. Metabolomic Analysis of Skeletal Muscle in Aged Mice. Sci. Rep. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Rhee, H.J.; Kim, E.; Lee, J.K. Physiological polyamines: Simple primordial stress molecules. J. Cell. Mol. Med. 2007, 11, 685–703. [Google Scholar] [CrossRef]
- Polis, B.; Karasik, D.; Samson, A.O. Alzheimer’s disease as a chronic maladaptive polyamine stress response. Aging 2021, 13, 10770–10795. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.; Romano, C.; Dichter, M.A.; Molinoff, P.B. Modulation of the NMDA receptor by polyamines. Life Sci. 1991, 48, 469–498. [Google Scholar] [CrossRef]
- Bowie, D. Polyamine-mediated channel block of ionotropic glutamate receptors and its regulation by auxiliary proteins. J. Biol. Chem. 2018, 293, 18789–18802. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, M. Chapter 26—Molecular basis of chronic neurodegeneration. In Clinical Molecular Medicine; Kumar, D., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 447–460. [Google Scholar]
- Cervelli, M.; Averna, M.; Vergani, L.; Pedrazzi, M.; Amato, S.; Fiorucci, C.; Rossi, M.N.; Maura, G.; Mariottini, P.; Cervetto, C.; et al. The Involvement of Polyamines Catabolism in the Crosstalk between Neurons and Astrocytes in Neurodegeneration. Biomedicines 2022, 10, 1756. [Google Scholar] [CrossRef] [PubMed]
- Perl, D.P. Neuropathology of Alzheimer’s disease. Mt. Sinai J. Med. 2010, 77, 32–42. [Google Scholar] [CrossRef]
- Xu, J.; Sun, Z.; Zhang, R.; Li, R.; Yu, Z.; Zhang, Q.; Ma, Y.; Xing, F.; Zheng, L. Non-linear association between serum spermidine and mild cognitive impairment: Results from a cross-sectional and longitudinal study. Front. Aging Neurosci. 2022, 14, 924984. [Google Scholar] [CrossRef]
- Pietrocola, F.; Lachkar, S.; Enot, D.P.; Niso-Santano, M.; Pedro, J.M.B.-S.; Sica, V.; Izzo, V.; Maiuri, M.C.; Madeo, F.; Mariño, G.; et al. Spermidine induces autophagy by inhibiting the acetyltransferase EP300. Cell Death Differ. 2015, 22, 509–516. [Google Scholar] [CrossRef]
- Frake, R.A.; Ricketts, T.; Menzies, F.M.; Rubinsztein, D.C. Autophagy and neurodegeneration. J. Clin. Investig. 2015, 125, 65–74. [Google Scholar] [CrossRef]
- Choi, A.M.; Ryter, S.W.; Levine, B. Autophagy in human health and disease. N. Engl. J. Med. 2013, 368, 651–662. [Google Scholar] [CrossRef]
- Freitag, K.; Sterczyk, N.; Wendlinger, S.; Obermayer, B.; Schulz, J.; Farztdinov, V.; Mülleder, M.; Ralser, M.; Houtman, J.; Fleck, L.; et al. Spermidine reduces neuroinflammation and soluble amyloid beta in an Alzheimer’s disease mouse model. J. Neuroinflamm. 2022, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Aarsland, D.; Batzu, L.; Halliday, G.M.; Geurtsen, G.J.; Ballard, C.; Ray Chaudhuri, K.; Weintraub, D. Parkinson disease—Associated cognitive impairment. Nat. Rev. Dis. Primers 2021, 7, 47. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.; Cao, W.J.; Zhao, R.; Lu, M.; Hu, G.; Qiao, C. ATP13A2 protects dopaminergic neurons in Parkinson’s disease: From biology to pathology. J. Biomed. Res. 2022, 36, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Haute, C.V.D.; Lobbestael, E.; Martin, S.; van Veen, S.; Vangheluwe, P.; Baekelandt, V. ATP13A2 Regulates Cellular α-Synuclein Multimerization, Membrane Association, and Externalization. Int. J. Mol. Sci. 2021, 22, 2689. [Google Scholar] [CrossRef] [PubMed]
- Usenovic, M.; Tresse, E.; Mazzulli, J.R.; Taylor, J.P.; Krainc, D. Deficiency of ATP13A2 Leads to Lysosomal Dysfunction, α-Synuclein Accumulation, and Neurotoxicity. J. Neurosci. 2012, 32, 4240–4246. [Google Scholar] [CrossRef]
- Fernández-Espejo, E.; Gavito, A.L.; Suárez, J.; Tolosa, E.; Vilas, D.; Aldecoa, I.; Berenguer, J.; Córdoba-Fernández, A.; Damas-Hermoso, F.; Rodríguez de Fonseca, F. Salivary ATP13A2 is a potential marker of therapy-induced motor complications and is expressed by inclusions in submandibulary glands in Parkinsonߣs disease. Clin. Park. Relat. Disord. 2022, 7, 100163. [Google Scholar] [CrossRef]
- Fernández-Espejo, E.; Rodriguez de Fonseca, F.; Suárez, J.; González-Aparicio, R.; Santurtún, A. ATP13A2 levels in serum and cerebrospinal fluid in patients with idiopathic Parkinson’s disease. Park. Relat Disord 2021, 88, 3–9. [Google Scholar] [CrossRef]
- NIH Maintaining Mobility and Preventing Disability Are Key to Living Independently as We Age. Available online: https://www.nia.nih.gov/news/maintaining-mobility-and-preventing-disability-are-key-living-independently-we-age (accessed on 15 December 2022).
- Salari, N.; Darvishi, N.; Bartina, Y.; Larti, M.; Kiaei, A.; Hemmati, M.; Shohaimi, S.; Mohammadi, M. Global prevalence of osteoporosis among the world older adults: A comprehensive systematic review and meta-analysis. J. Orthop. Surg. Res. 2021, 16, 1–13. [Google Scholar] [CrossRef]
- National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS). Osteoporosis. Available online: https://www.niams.nih.gov/health-topics/osteoporosis (accessed on 6 May 2023).
- Sözen, T.; Özışık, L.; Başaran, N. An overview and management of osteoporosis. Eur. J. Rheumatol. 2017, 4, 46–56. [Google Scholar] [CrossRef]
- Mu, K.; Akr, R. Osteoporosis In Females. Available online: https://www.ncbi.nlm.nih.gov/books/NBK559156/ (accessed on 7 July 2023).
- Brown, J.P. Long-Term Treatment of Postmenopausal Osteoporosis. Endocrinol. Metab. 2021, 36, 544–552. [Google Scholar] [CrossRef]
- Murray-Stewart, T.; Dunworth, M.; Foley, J.R.; Schwartz, C.E.; Casero, R.A. Polyamine Homeostasis in Snyder-Robinson Syndrome. Med. Sci. 2018, 6, 112. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chen, Y.; Huang, Y.; Hsu, Y.; Chiang, L.; Chen, T.; Wang, G. Exogenous polyamines promote osteogenic differentiation by reciprocally regulating osteogenic and adipogenic gene expression. J. Cell. Biochem. 2013, 114, 2718–2728. [Google Scholar] [CrossRef] [PubMed]
- Facchini, A.; Borzì, R.M.; Olivotto, E.; Platano, D.; Pagani, S.; Cetrullo, S.; Flamigni, F. Role of polyamines in hypertrophy and terminal differentiation of osteoarthritic chondrocytes. Amino Acids 2012, 42, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Park, G.; Node, J.; Ozaki, K.; Hiraiwa, M.; Kitaguchi, Y.; Kaneda, K.; Hiramoto, S.; Hinoi, E. Daily intake of polyamine-rich Saccharomyces cerevisiae S631 prevents osteoclastic activation and bone loss in ovariectomized mice. Food Sci. Biotechnol. 2019, 28, 1241–1245. [Google Scholar] [CrossRef] [PubMed]
- Sacitharan, P.K. Ageing and Osteoarthritis. In Biochemistry and Cell Biology of Ageing: Part II Clinical Science; Harris, J.R., Korolchuk, V.I., Eds.; Springer: Singapore, 2019; pp. 123–159. [Google Scholar]
- Madeo, F.; Bauer, M.A.; Carmona-Gutierrez, D.; Kroemer, G. Spermidine: A physiological autophagy inducer acting as an anti-aging vitamin in humans? Autophagy 2019, 15, 165–168. [Google Scholar] [CrossRef] [PubMed]
- D’Adamo, S.; Cetrullo, S.; Guidotti, S.; Silvestri, Y.; Minguzzi, M.; Santi, S.; Cattini, L.; Filardo, G.; Flamigni, F.; Borzì, R.M. Spermidine rescues the deregulated autophagic response to oxidative stress of osteoarthritic chondrocytes. Free Radic. Biol. Med. 2020, 153, 159–172. [Google Scholar] [CrossRef]
- Chen, Z.; Lin, C.X.; Song, B.; Li, C.C.; Qiu, J.X.; Li, S.X.; Lin, S.P.; Luo, W.Q.; Fu, Y.; Fang, G.B.; et al. Spermidine activates RIP1 deubiquitination to inhibit TNF-α-induced NF-κB/p65 signaling pathway in osteoarthritis. Cell Death Dis. 2020, 11, 503. [Google Scholar] [CrossRef]
- Minguzzi, M.; Guidotti, S.; Platano, D.; D’adamo, S.; Cetrullo, S.; Assirelli, E.; Santi, S.; Mariani, E.; Trisolino, G.; Filardo, G.; et al. Polyamine supplementation reduces DNA damage in adipose stem cells cultured in 3-D. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Tabbaa, M.; Gomez, T.R.; Campelj, D.G.; Gregorevic, P.; Hayes, A.; Goodman, C.A. The regulation of polyamine pathway proteins in models of skeletal muscle hypertrophy and atrophy: A potential role for mTORC1. Am. J. Physiol. Physiol. 2021, 320, C987–C999. [Google Scholar] [CrossRef]
- Jimenez-Gutierrez, G.E.; Martínez-Gómez, L.E.; Martínez-Armenta, C.; Pineda, C.; Martínez-Nava, G.A.; Lopez-Reyes, A. Molecular Mechanisms of Inflammation in Sarcopenia: Diagnosis and Therapeutic Update. Cells 2022, 11, 2359. [Google Scholar] [CrossRef]
- Hosoi, T.; Yakabe, M.; Sasakawa, H.; Sasako, T.; Ueki, K.; Kato, S.; Tokuoka, S.M.; Oda, Y.; Abe, M.; Matsumoto, T.; et al. Sarcopenia phenotype and impaired muscle function in male mice with fast-twitch muscle-specific knockout of the androgen receptor. Proc. Natl. Acad. Sci. USA 2023, 120, e2218032120. [Google Scholar] [CrossRef] [PubMed]
- Kanou, M.; Nakamura, K.; Horie, K.; Sakai, H.; Yanagihara, Y.; Sakakibara, I.; Yamana, K.; Imai, Y. Polyamine pathway is associated with muscle anabolic effects by androgen receptor ligand. JCSM Rapid. Commun. 2020, 4, 57–74. [Google Scholar] [CrossRef]
- Sagar, N.A.; Tarafdar, S.; Agarwal, S.; Tarafdar, A.; Sharma, S. Polyamines: Functions, Metabolism, and Role in Human Disease Management. Med. Sci. 2021, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Makletsova, M.G.; Rikhireva, G.T.; Kirichenko, E.Y.; Trinitatsky, I.Y.; Vakulenko, M.Y.; Ermakov, A.M. The Role of Polyamines in the Mechanisms of Cognitive Impairment. Neurochem. J. 2022, 16, 283–294. [Google Scholar] [CrossRef]
- Sánchez, M.; Suárez, L.; Banda, G.; Barreiro-Alonso, E.; Rodríguez-Uña, I.; Rubín, J.M.; Cantabrana, B. Age-associated polyamines in peripheral blood cells and plasma in 20 to 70 years of age subjects. Amino Acids 2023, 55, 789–798. [Google Scholar] [CrossRef]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6, 10. [Google Scholar] [CrossRef]
- Hofer, S.J.; Simon, A.K.; Bergmann, M.; Eisenberg, T.; Kroemer, G.; Madeo, F. Mechanisms of spermidine-induced autophagy and geroprotection. Nat. Aging 2022, 2, 1112–1129. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Disease | Type of Sample | Polyamine Impairment Effect | Ref. |
|---|---|---|---|
| Alzheimer’s Disease | Temporal cortex and occipital cortex tissue |
| [45] |
| Cortical surface tissue |
| [46] | |
| Brain tissue |
| [47] | |
| [48] | ||
| [49] | ||
| [50] | ||
| Plasma and CSF |
| [51] | |
| Serum |
| [36] | |
| Parkinson’s Disease | Brain tissue |
| [52] |
| [53] | ||
| CSF |
| [54] | |
| CSF and serum |
| [55] | |
| CSF and red blood cells |
| [56] | |
| Serum |
| [57] | |
| [58] | ||
| Plasma |
| [37] | |
| Osteoporosis |
| [44] | |
| Osteoarthritis | Serum |
| [39] |
| Sarcopenia | Whole blood |
| [40] |
| * Mouse skeletal muscle |
| [59] | |
| [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez Gutierrez, G.E.; Borbolla Jiménez, F.V.; Muñoz, L.G.; Tapia Guerrero, Y.S.; Murillo Melo, N.M.; Cristóbal-Luna, J.M.; Leyva Garcia, N.; Cordero-Martínez, J.; Magaña, J.J. The Molecular Role of Polyamines in Age-Related Diseases: An Update. Int. J. Mol. Sci. 2023, 24, 16469. https://doi.org/10.3390/ijms242216469
Jimenez Gutierrez GE, Borbolla Jiménez FV, Muñoz LG, Tapia Guerrero YS, Murillo Melo NM, Cristóbal-Luna JM, Leyva Garcia N, Cordero-Martínez J, Magaña JJ. The Molecular Role of Polyamines in Age-Related Diseases: An Update. International Journal of Molecular Sciences. 2023; 24(22):16469. https://doi.org/10.3390/ijms242216469
Chicago/Turabian StyleJimenez Gutierrez, Guadalupe Elizabeth, Fabiola V. Borbolla Jiménez, Luis G. Muñoz, Yessica Sarai Tapia Guerrero, Nadia Mireya Murillo Melo, José Melesio Cristóbal-Luna, Norberto Leyva Garcia, Joaquín Cordero-Martínez, and Jonathan J. Magaña. 2023. "The Molecular Role of Polyamines in Age-Related Diseases: An Update" International Journal of Molecular Sciences 24, no. 22: 16469. https://doi.org/10.3390/ijms242216469