
Magnetic Stimulation as a Therapeutic Approach for Brain Modulation and Repair: Underlying Molecular and Cellular Mechanisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
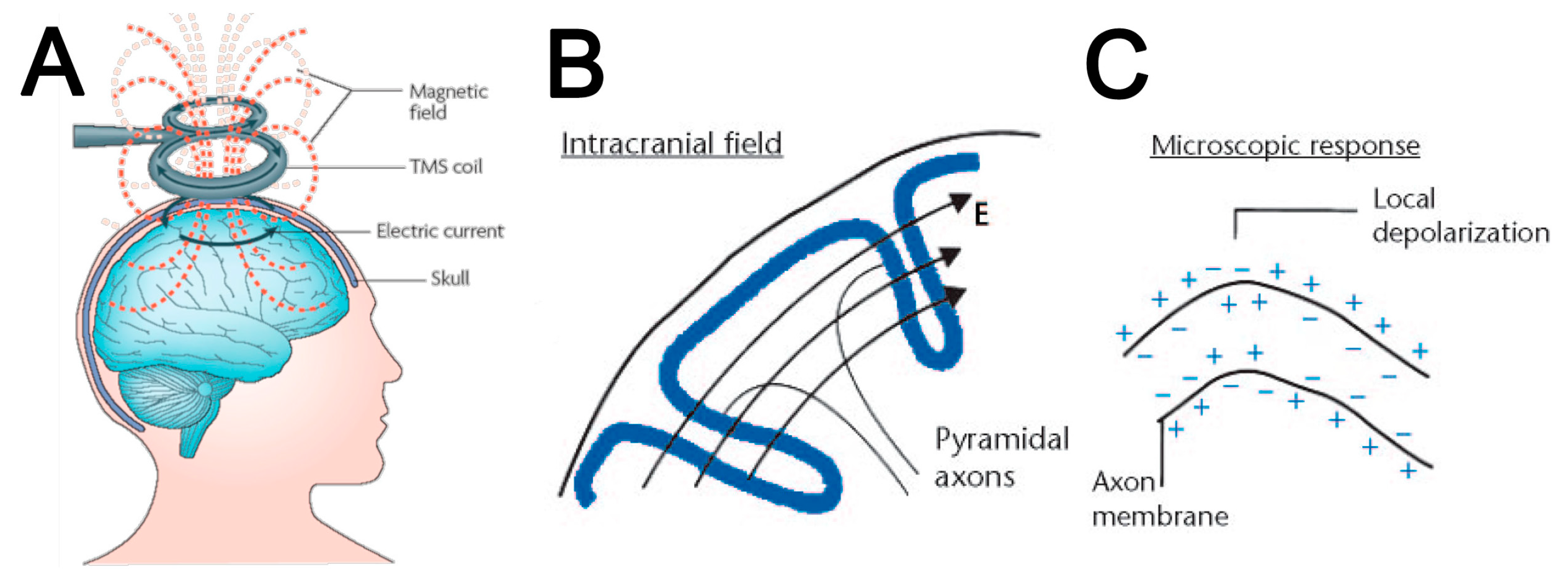
:1. Introduction: Why Use Magnetic Fields to Stimulate the Brain?
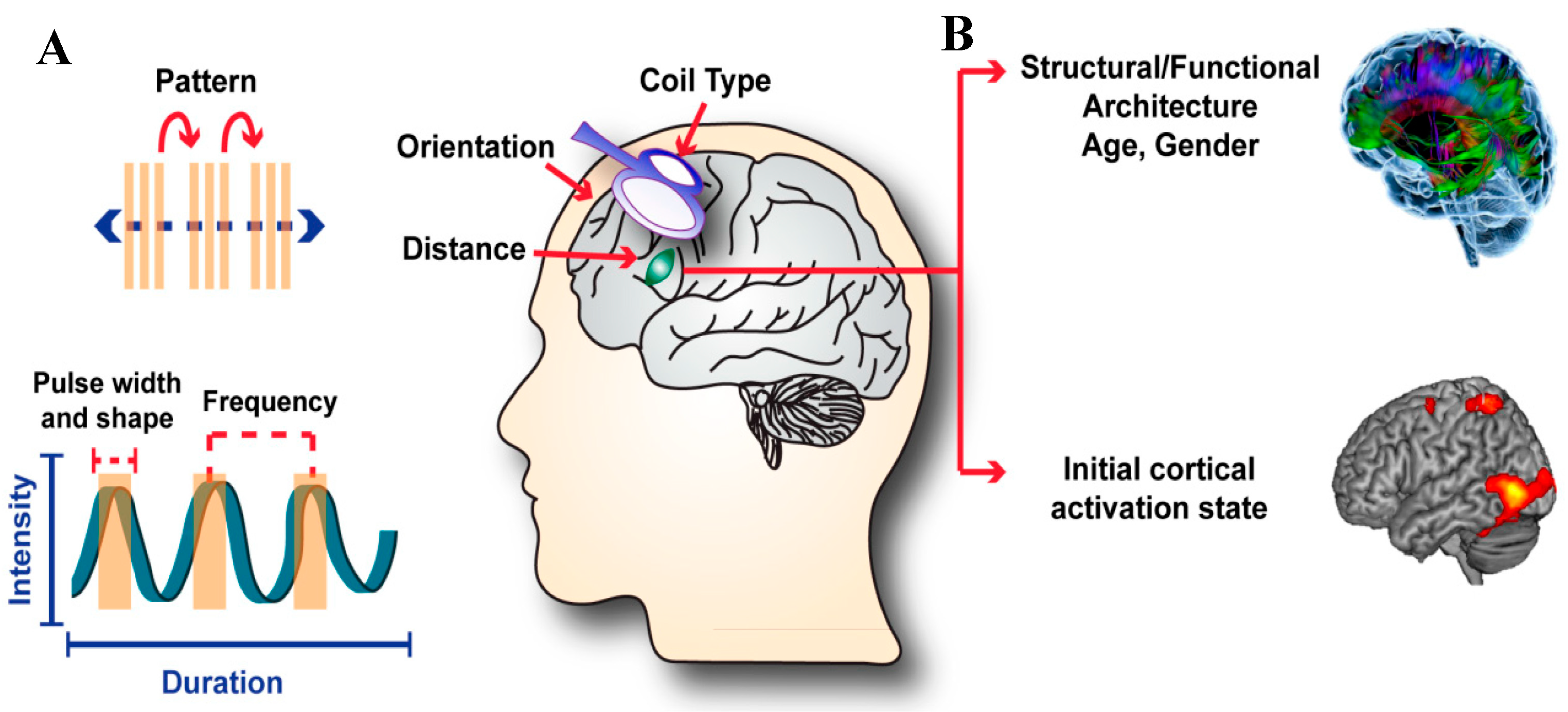
2. Magnetic Brain Stimulation Parameters Determine Outcomes
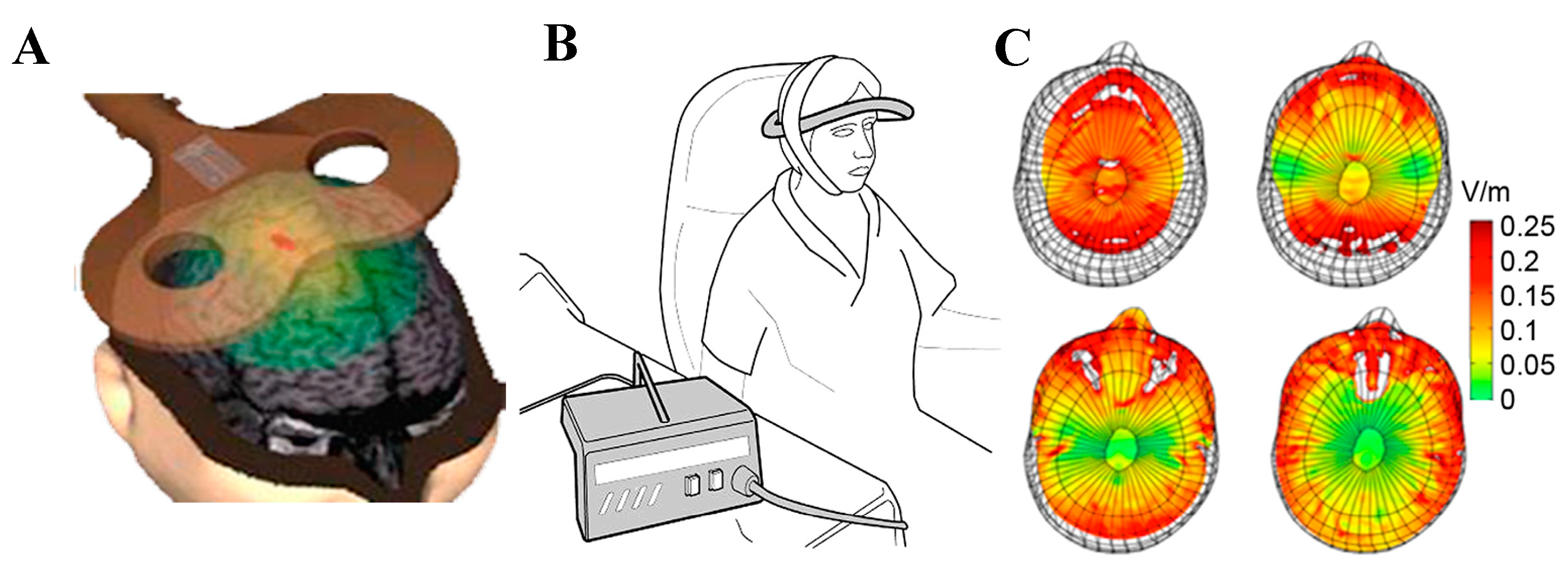
2.1. Types of Coil
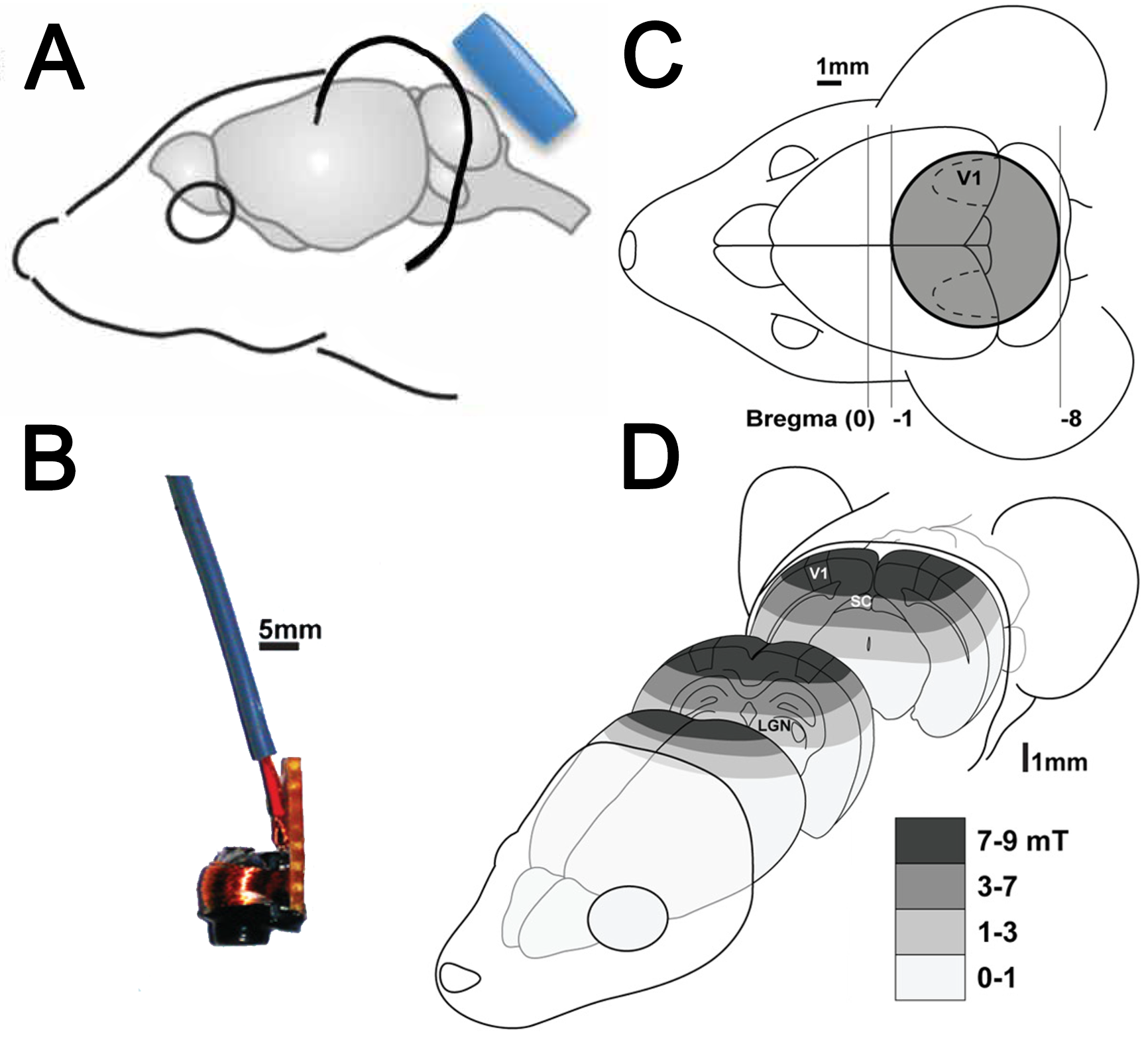
2.2. Stimulation Intensity
2.3. Frequency and Pattern of Magnetic Stimulation Pulses
3. Potential Cellular Mechanisms Mediating the Effects of Magnetic Stimulation
3.1. Synaptic Plasticity
3.2. Acute Effects on Neuronal Properties
3.2.1. Membrane Potential and Spontaneous Activity
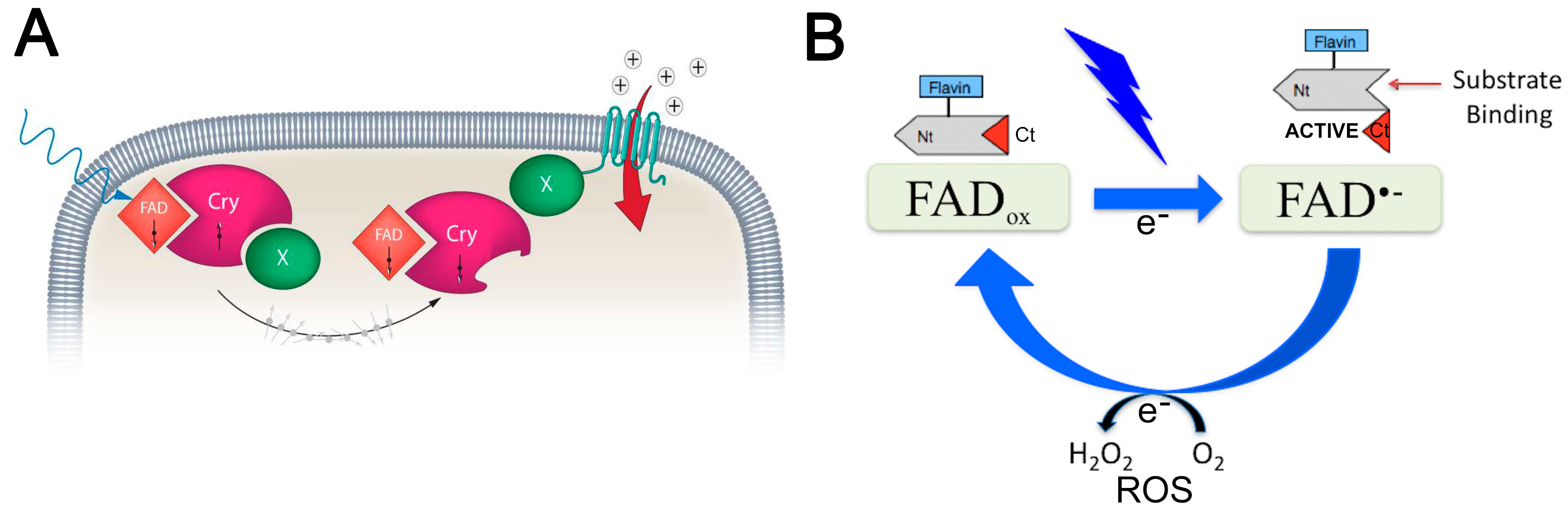
3.2.2. Activation of Cryptochrome Magnetoreceptors
3.3. Intracellular Cascades Modified by Magnetic Fields
3.3.1. ROS Production and Regulation of Oxidative Stress
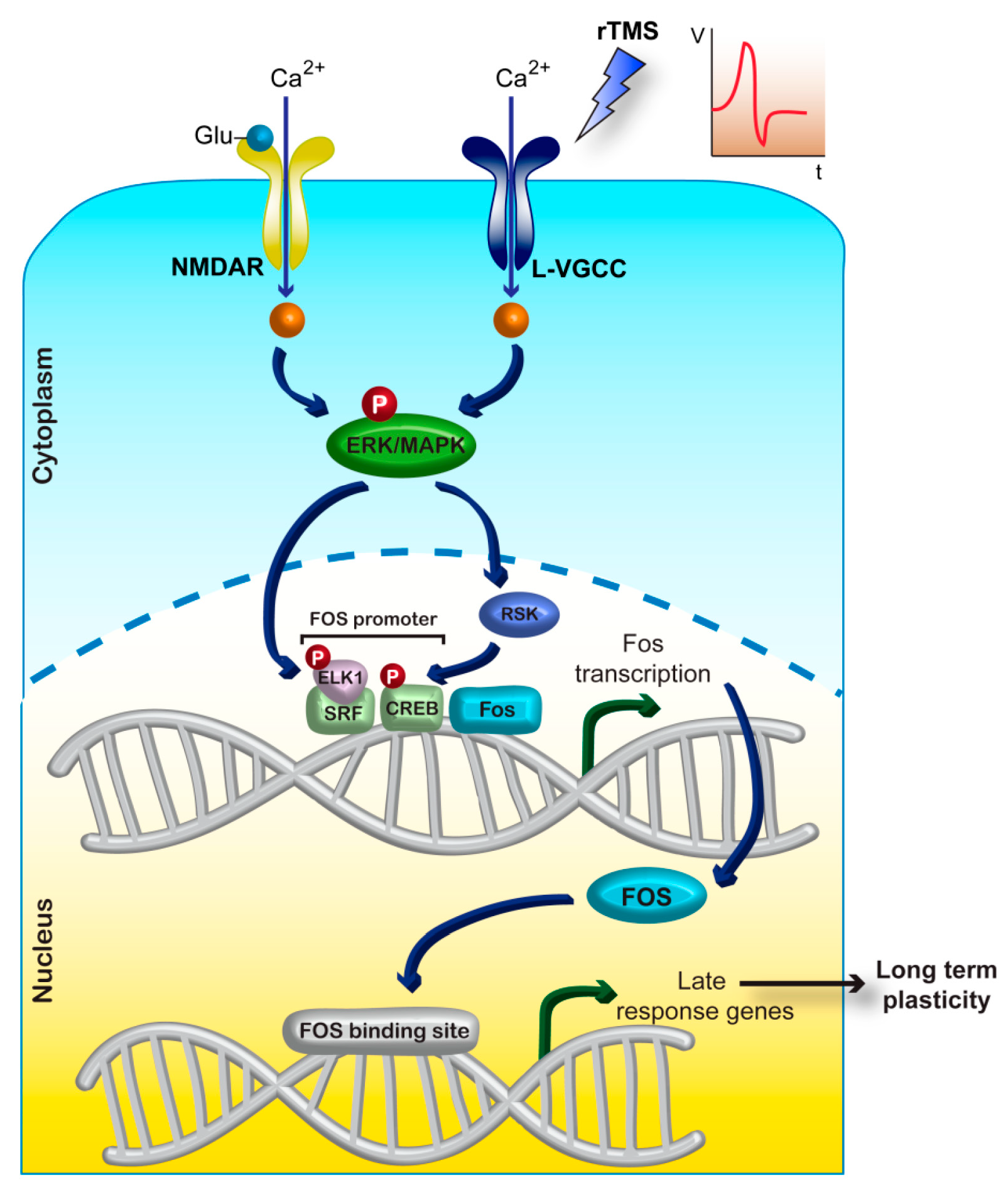
3.3.2. Intracellular Ca2+ Concentration and Downstream Signalling
3.3.3. Immediate-Early Gene Expression
3.4. Long-Lasting Brain Plasticity
3.4.1. Brain-Derived Neurotrophic Factor and Other Gene-Expression Changes
3.4.2. Dendritic Spine Plasticity
3.4.3. Neurogenesis
4. Relevance of Magnetic Stimulation to Neuropsychiatric Treatment
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BDNF | Brain-derived neurotrophic factor |
| BHFS | Biomimetic high-frequency stimulation |
| CAMKII | Calmodulin-dependent kinase II |
| CB | Calbindin: a calcium-binding protein |
| CREB | cAMP response element-binding protein |
| CRY | Cryptochrome |
| EEG | Electroencephalography |
| FAD | Flavin adenosine dinucleotide |
| fMRI | Functional magnetic resonance imaging |
| FSI | Fast-spiking interneurons |
| GABA | Gamma amino butyric acid |
| GAD | Glutamic acid decarboxylase; |
| IEG | Immediate-early gene |
| LFMS | Low-field magnetic stimulation |
| LI-rTMS | Low-intensity repetitive magnetic stimulation |
| LTP | Long-term potentiation |
| M1 | Primary motor cortex |
| MEP | Motor evoked potential |
| NMDA receptors | N-methyl-D-aspartate receptors are a subtype of glutamate receptors |
| PV | Parvalbumin: a calcium-binding protein |
| RMT | Resting-motor threshold |
| ROS | Reactive oxygen species |
| TMS | Transcranial magnetic stimulation |
| TBS | Theta-burst stimulation; continuous (cTBS) or intermittent (iTBS) |
| VGCC | Voltage-gated calcium channel |
References
- Barker, A.T.; Jalinous, R.; Freeston, I.L. Non-invasive magnetic stimulation of human motor cortex. Lancet 1985, 325, 1106–1107. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wu, Q.; Gao, L.; Zheng, Z.; Xiang, H.; Yang, K.; Yu, B.; Yao, J. Advancements in Transcranial Magnetic Stimulation Research and the Path to Precision. NDT 2023, 19, 1841–1851. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Capone, F.; Apollonio, F.; Borea, P.A.; Cadossi, R.; Fassina, L.; Grassi, C.; Liberti, M.; Paffi, A.; Parazzini, M.; et al. A Consensus Panel Review of Central Nervous System Effects of the Exposure to Low-Intensity Extremely Low-Frequency Magnetic Fields. Brain Stimul. 2013, 6, 469–476. [Google Scholar] [CrossRef] [PubMed]
- Pell, G.S.; Roth, Y.; Zangen, A. Modulation of Cortical Excitability Induced by Repetitive Transcranial Magnetic Stimulation: Influence of Timing and Geometrical Parameters and Underlying Mechanisms. Prog. Neurobiol. 2011, 93, 59–98. [Google Scholar] [CrossRef]
- Sack, A.T.; Paneva, J.; Küthe, T.; Dijkstra, E.; Zwienenberg, L.; Arns, M.; Schuhmann, T. Target Engagement and Brain State Dependence of Transcranial Magnetic Stimulation: Implications for Clinical Practice. Biol. Psychiatry, 2023; in press. [Google Scholar] [CrossRef]
- Valero-Cabré, A.; Amengual, J.L.; Stengel, C.; Pascual-Leone, A.; Coubard, O.A. Transcranial Magnetic Stimulation in Basic and Clinical Neuroscience: A Comprehensive Review of Fundamental Principles and Novel Insights. Neurosci. Biobehav. Rev. 2017, 83, 381–404. [Google Scholar] [CrossRef]
- Huang, W.; Chen, Q.; Liu, J.; Liu, L.; Tang, J.; Zou, M.; Zeng, T.; Li, H.; Jiang, Q.; Jiang, Q. Transcranial Magnetic Stimulation in Disorders of Consciousness: An Update and Perspectives. Aging Dis. 2023, 14, 1171–1183. [Google Scholar] [CrossRef] [PubMed]
- Oberman, L.M.; Benussi, A. Transcranial Magnetic Stimulation Across the Lifespan: Impact of Developmental and Degenerative Processes. Biol. Psychiatry 2023. [CrossRef] [PubMed]
- Ridding, M.C.; Rothwell, J.C. Is There a Future for Therapeutic Use of Transcranial Magnetic Stimulation? Nat. Rev. Neurosci. 2007, 8, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Butler, A.J.; Wolf, S.L. Putting the Brain on the Map: Use of Transcranial Magnetic Stimulation to Assess and Induce Cortical Plasticity of Upper-Extremity Movement. Phys. Ther. 2007, 87, 719–736. [Google Scholar] [CrossRef]
- Thut, G.; Veniero, D.; Romei, V.; Miniussi, C.; Schyns, P.; Gross, J. Rhythmic TMS Causes Local Entrainment of Natural Oscillatory Signatures. Curr. Biol. 2011, 21, 1176–1185. [Google Scholar] [CrossRef]
- Fröhlich, F.; McCormick, D.A. Endogenous Electric Fields May Guide Neocortical Network Activity. Neuron 2010, 67, 129–143. [Google Scholar] [CrossRef]
- Dedoncker, J.; Brunoni, A.R.; Baeken, C.; Vanderhasselt, M.-A. A Systematic Review and Meta-Analysis of the Effects of Transcranial Direct Current Stimulation (tDCS) Over the Dorsolateral Prefrontal Cortex in Healthy and Neuropsychiatric Samples: Influence of Stimulation Parameters. Brain Stimul. 2016, 9, 501–517. [Google Scholar] [CrossRef]
- Rohan, M.L.; Yamamoto, R.T.; Ravichandran, C.T.; Cayetano, K.R.; Morales, O.G.; Olson, D.P.; Vitaliano, G.; Paul, S.M.; Cohen, B.M. Rapid Mood-Elevating Effects of Low Field Magnetic Stimulation in Depression. Biol. Psychiatry 2014, 76, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Rodger, J.; Mo, C.; Wilks, T.; Dunlop, S.A.; Sherrard, R.M. Transcranial Pulsed Magnetic Field Stimulation Facilitates Reorganization of Abnormal Neural Circuits and Corrects Behavioral Deficits without Disrupting Normal Connectivity. FASEB J. 2012, 26, 1593–1606. [Google Scholar] [CrossRef]
- Seewoo, B.J.; Feindel, K.W.; Etherington, S.J.; Rodger, J. Resting-State fMRI Study of Brain Activation Using Low-Intensity Repetitive Transcranial Magnetic Stimulation in Rats. Sci. Rep. 2018, 8, 6706. [Google Scholar] [CrossRef] [PubMed]
- Makowiecki, K.; Garrett, A.; Harvey, A.R.; Rodger, J. Low-Intensity Repetitive Transcranial Magnetic Stimulation Requires Concurrent Visual System Activity to Modulate Visual Evoked Potentials in Adult Mice. Sci. Rep. 2018, 8, 5792. [Google Scholar] [CrossRef]
- Dufor, T.; Grehl, S.; Tang, A.D.; Doulazmi, M.; Traoré, M.; Debray, N.; Dubacq, C.; Deng, Z.-D.; Mariani, J.; Lohof, A.M.; et al. Neural Circuit Repair by Low-Intensity Magnetic Stimulation Requires Cellular Magnetoreceptors and Specific Stimulation Patterns. Sci. Adv. 2019, 5, eaav9847. [Google Scholar] [CrossRef] [PubMed]
- Haustead, D.J.; Lukehurst, S.S.; Clutton, G.T.; Bartlett, C.A.; Dunlop, S.A.; Arrese, C.A.; Sherrard, R.M.; Rodger, J. Functional Topography and Integration of the Contralateral and Ipsilateral Retinocollicular Projections of Ephrin-A-/- Mice. J. Neurosci. 2008, 28, 7376–7386. [Google Scholar] [CrossRef] [PubMed]
- Makowiecki, K.; Harvey, A.R.; Sherrard, R.M.; Rodger, J. Low-Intensity Repetitive Transcranial Magnetic Stimulation Improves Abnormal Visual Cortical Circuit Topography and Upregulates BDNF in Mice. J. Neurosci. 2014, 34, 10780–10792. [Google Scholar] [CrossRef]
- Poh, E.Z.; Harvey, A.R.; Makowiecki, K.; Rodger, J. Online LI-rTMS during a Visual Learning Task: Differential Impacts on Visual Circuit and Behavioral Plasticity in Adult Ephrin-A2A5-/- Mice. eNeuro 2018, 5. [Google Scholar] [CrossRef]
- Grehl, S.; Viola, H.M.; Fuller-Carter, P.I.; Carter, K.W.; Dunlop, S.A.; Hool, L.C.; Sherrard, R.M.; Rodger, J. Cellular and Molecular Changes to Cortical Neurons Following Low Intensity Repetitive Magnetic Stimulation at Different Frequencies. Brain Stimul. 2015, 8, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Hallett, M.; Rossini, P.M.; Pascual-Leone, A. Safety, Ethical Considerations, and Application Guidelines for the Use of Transcranial Magnetic Stimulation in Clinical Practice and Research. Clin. Neurophysiol. 2009, 120, 2008–2039. [Google Scholar] [CrossRef] [PubMed]
- Shafi, M.; Stern, A.P.; Pascual-Leone, A. Adding Low-Field Magnetic Stimulation to Noninvasive Electromagnetic Neuromodulatory Therapies. Biol. Psychiatry 2014, 76, 170–171. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Leone, A.; Walsh, V.; Rothwell, J. Transcranial Magnetic Stimulation in Cognitive Neuroscience—Virtual Lesion, Chronometry, and Functional Connectivity. Curr. Opin. Neurobiol. 2000, 10, 232–237. [Google Scholar] [CrossRef]
- Cohen, L.G.; Roth, B.J.; Nilsson, J.; Dang, N.; Panizza, M.; Bandinelli, S.; Friauf, W.; Hallett, M. Effects of Coil Design on Delivery of Focal Magnetic Stimulation. Technical Considerations. Electroencephalogr. Clin. Neurophysiol. 1990, 75, 350–357. [Google Scholar] [CrossRef]
- Hallett, M. Transcranial Magnetic Stimulation: A Primer. Neuron 2007, 55, 187–199. [Google Scholar] [CrossRef]
- Deng, Z.-D.; Lisanby, S.H.; Peterchev, A.V. Electric Field Depth–Focality Tradeoff in Transcranial Magnetic Stimulation: Simulation Comparison of 50 Coil Designs. Brain Stimul. 2013, 6, 1–13. [Google Scholar] [CrossRef]
- Trillenberg, P.; Bremer, S.; Oung, S.; Erdmann, C.; Schweikard, A.; Richter, L. Variation of Stimulation Intensity in Transcranial Magnetic Stimulation with Depth. J. Neurosci. Methods 2012, 211, 185–190. [Google Scholar] [CrossRef]
- Westin, G.G.; Bassi, B.D.; Lisanby, S.H.; Luber, B. Determination of Motor Threshold Using Visual Observation Overestimates Transcranial Magnetic Stimulation Dosage: Safety Implications. Clin. Neurophysiol. 2014, 125, 142–147. [Google Scholar] [CrossRef]
- Rothwell, J.C.; Thompson, P.D.; Day, B.L.; Dick, J.P.; Kachi, T.; Cowan, J.M.; Marsden, C.D. Motor Cortex Stimulation in Intact Man. 1. General Characteristics of EMG Responses in Different Muscles. Brain 1987, 110 Pt 5, 1173–1190. [Google Scholar]
- Capone, F.; Dileone, M.; Profice, P.; Pilato, F.; Musumeci, G.; Minicuci, G.; Ranieri, F.; Cadossi, R.; Setti, S.; Tonali, P.A.; et al. Does Exposure to Extremely Low Frequency Magnetic Fields Produce Functional Changes in Human Brain? J. Neural. Transm. 2009, 116, 257–265. [Google Scholar] [CrossRef]
- Bell, G.B.; Marino, A.A.; Chesson, A.L. Frequency-Specific Responses in the Human Brain Caused by Electromagnetic Fields. J. Neurol. Sci. 1994, 123, 26–32. [Google Scholar] [CrossRef]
- Cvetkovic, D.; Cosic, I. Alterations of Human Electroencephalographic Activity Caused by Multiple Extremely Low Frequency Magnetic Field Exposures. Med. Biol. Eng. Comput. 2009, 47, 1063. [Google Scholar] [CrossRef] [PubMed]
- Romei, V.; Bauer, M.; Brooks, J.L.; Economides, M.; Penny, W.; Thut, G.; Driver, J.; Bestmann, S. Causal Evidence That Intrinsic Beta-Frequency Is Relevant for Enhanced Signal Propagation in the Motor System as Shown through Rhythmic TMS. NeuroImage 2016, 126, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Rossini, P.M.; Burke, D.; Chen, R.; Cohen, L.G.; Daskalakis, Z.; Di Iorio, R.; Di Lazzaro, V.; Ferreri, F.; Fitzgerald, P.B.; George, M.S.; et al. Non-Invasive Electrical and Magnetic Stimulation of the Brain, Spinal Cord, Roots and Peripheral Nerves: Basic Principles and Procedures for Routine Clinical and Research Application. An Updated Report from an I.F.C.N. Committee. Clin. Neurophysiol. 2015, 126, 1071–1107. [Google Scholar] [CrossRef]
- Fitzgerald, P.B.; Fountain, S.; Daskalakis, Z.J. A Comprehensive Review of the Effects of rTMS on Motor Cortical Excitability and Inhibition. Clin. Neurophysiol. 2006, 117, 2584–2596. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Edwards, M.J.; Rounis, E.; Bhatia, K.P.; Rothwell, J.C. Theta Burst Stimulation of the Human Motor Cortex. Neuron 2005, 45, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Di Lazzaro, V.; Dileone, M.; Pilato, F.; Capone, F.; Musumeci, G.; Ranieri, F.; Ricci, V.; Bria, P.; Di Iorio, R.; de Waure, C.; et al. Modulation of Motor Cortex Neuronal Networks by rTMS: Comparison of Local and Remote Effects of Six Different Protocols of Stimulation. J. Neurophysiol. 2011, 105, 2150–2156. [Google Scholar] [CrossRef]
- Hoogendam, J.M.; Ramakers, G.M.J.; Di Lazzaro, V. Physiology of Repetitive Transcranial Magnetic Stimulation of the Human Brain. Brain Stimul. 2010, 3, 95–118. [Google Scholar] [CrossRef]
- Di Lazzaro, V.; Profice, P.; Pilato, F.; Dileone, M.; Oliviero, A.; Ziemann, U. The Effects of Motor Cortex rTMS on Corticospinal Descending Activity. Clin. Neurophysiol. 2010, 121, 464–473. [Google Scholar] [CrossRef]
- Matheson, N.A.; Shemmell, J.B.H.; De Ridder, D.; Reynolds, J.N.J. Understanding the Effects of Repetitive Transcranial Magnetic Stimulation on Neuronal Circuits. Front. Neural Circuits 2016, 10, 67. [Google Scholar] [CrossRef]
- Hamada, M.; Murase, N.; Hasan, A.; Balaratnam, M.; Rothwell, J.C. The Role of Interneuron Networks in Driving Human Motor Cortical Plasticity. Cereb. Cortex 2013, 23, 1593–1605. [Google Scholar] [CrossRef]
- Gamboa, O.L.; Antal, A.; Laczo, B.; Moliadze, V.; Nitsche, M.A.; Paulus, W. Impact of Repetitive Theta Burst Stimulation on Motor Cortex Excitability. Brain Stimul. 2011, 4, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Gentner, R.; Wankerl, K.; Reinsberger, C.; Zeller, D.; Classen, J. Depression of Human Corticospinal Excitability Induced by Magnetic Theta-Burst Stimulation: Evidence of Rapid Polarity-Reversing Metaplasticity. Cereb. Cortex 2008, 18, 2046–2053. [Google Scholar] [CrossRef] [PubMed]
- Funke, K.; Benali, A. Modulation of Cortical Inhibition by rTMS—Findings Obtained from Animal Models. J. Physiol. 2011, 589, 4423–4435. [Google Scholar] [CrossRef] [PubMed]
- Esser, S.K.; Huber, R.; Massimini, M.; Peterson, M.J.; Ferrarelli, F.; Tononi, G. A Direct Demonstration of Cortical LTP in Humans: A Combined TMS/EEG Study. Brain Res. Bull. 2006, 69, 86–94. [Google Scholar] [CrossRef]
- Litvak, V.; Zeller, D.; Oostenveld, R.; Maris, E.; Cohen, A.; Schramm, A.; Gentner, R.; Zaaroor, M.; Pratt, H.; Classen, J. LTP-like Changes Induced by Paired Associative Stimulation of the Primary Somatosensory Cortex in Humans: Source Analysis and Associated Changes in Behaviour. Eur. J. Neurosci. 2007, 25, 2862–2874. [Google Scholar] [CrossRef]
- Lee, L.; Siebner, H.R.; Rowe, J.B.; Rizzo, V.; Rothwell, J.C.; Frackowiak, R.S.J.; Friston, K.J. Acute Remapping within the Motor System Induced by Low-Frequency Repetitive Transcranial Magnetic Stimulation. J. Neurosci. 2003, 23, 5308–5318. [Google Scholar] [CrossRef] [PubMed]
- Rounis, E.; Lee, L.; Siebner, H.R.; Rowe, J.B.; Friston, K.J.; Rothwell, J.C.; Frackowiak, R.S.J. Frequency Specific Changes in Regional Cerebral Blood Flow and Motor System Connectivity Following rTMS to the Primary Motor Cortex. Neuroimage 2005, 26, 164–176. [Google Scholar] [CrossRef]
- Hubl, D.; Nyffeler, T.; Wurtz, P.; Chaves, S.; Pflugshaupt, T.; Lüthi, M.; von Wartburg, R.; Wiest, R.; Dierks, T.; Strik, W.K.; et al. Time Course of Blood Oxygenation Level-Dependent Signal Response after Theta Burst Transcranial Magnetic Stimulation of the Frontal Eye Field. Neuroscience 2008, 151, 921–928. [Google Scholar] [CrossRef]
- Abraham, W.C. Metaplasticity: Tuning Synapses and Networks for Plasticity. Nat. Rev. Neurosci. 2008, 9, 387. [Google Scholar] [CrossRef]
- Valero-Cabré, A.; Pascual-Leone, A.; Rushmore, R.J. Cumulative Sessions of Repetitive Transcranial Magnetic Stimulation (rTMS) Build up Facilitation to Subsequent TMS-Mediated Behavioural Disruptions. Eur. J. Neurosci. 2008, 27, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Karabanov, A.; Ziemann, U.; Hamada, M.; George, M.S.; Quartarone, A.; Classen, J.; Massimini, M.; Rothwell, J.; Siebner, H.R. Consensus Paper: Probing Homeostatic Plasticity of Human Cortex With Non-Invasive Transcranial Brain Stimulation. Brain Stimul. 2015, 8, 993–1006. [Google Scholar] [CrossRef] [PubMed]
- Todd, G.; Flavel, S.C.; Ridding, M.C. Priming Theta-Burst Repetitive Transcranial Magnetic Stimulation with Low- and High-Frequency Stimulation. Exp. Brain Res. 2009, 195, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Lenz, M.; Platschek, S.; Priesemann, V.; Becker, D.; Willems, L.M.; Ziemann, U.; Deller, T.; Müller-Dahlhaus, F.; Jedlicka, P.; Vlachos, A. Repetitive Magnetic Stimulation Induces Plasticity of Excitatory Postsynapses on Proximal Dendrites of Cultured Mouse CA1 Pyramidal Neurons. Brain Struct. Funct. 2015, 220, 3323–3337. [Google Scholar] [CrossRef] [PubMed]
- Tokay, T.; Holl, N.; Kirschstein, T.; Zschorlich, V.; Köhling, R. High-Frequency Magnetic Stimulation Induces Long-Term Potentiation in Rat Hippocampal Slices. Neurosci. Lett. 2009, 461, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Vlachos, A.; Müller-Dahlhaus, F.; Rosskopp, J.; Lenz, M.; Ziemann, U.; Deller, T. Repetitive Magnetic Stimulation Induces Functional and Structural Plasticity of Excitatory Postsynapses in Mouse Organotypic Hippocampal Slice Cultures. J. Neurosci. 2012, 32, 17514–17523. [Google Scholar] [CrossRef]
- Labedi, A.; Benali, A.; Mix, A.; Neubacher, U.; Funke, K. Modulation of Inhibitory Activity Markers by Intermittent Theta-Burst Stimulation in Rat Cortex Is NMDA-Receptor Dependent. Brain Stimul. 2014, 7, 394–400. [Google Scholar] [CrossRef] [PubMed]
- Trippe, J.; Mix, A.; Aydin-Abidin, S.; Funke, K.; Benali, A. θ Burst and Conventional Low-Frequency rTMS Differentially Affect GABAergic Neurotransmission in the Rat Cortex. Exp. Brain Res. 2009, 199, 411–421. [Google Scholar] [CrossRef]
- Aydin-Abidin, S.; Trippe, J.; Funke, K.; Eysel, U.T.; Benali, A. High- and Low-Frequency Repetitive Transcranial Magnetic Stimulation Differentially Activates c-Fos and Zif268 Protein Expression in the Rat Brain. Exp. Brain Res. 2008, 188, 249–261. [Google Scholar] [CrossRef]
- Moretti, J.; Terstege, D.J.; Poh, E.Z.; Epp, J.R.; Rodger, J. Low Intensity Repetitive Transcranial Magnetic Stimulation Modulates Brain-Wide Functional Connectivity to Promote Anti-Correlated c-Fos Expression. Sci. Rep. 2022, 12, 20571. [Google Scholar] [CrossRef]
- Volz, L.J.; Benali, A.; Mix, A.; Neubacher, U.; Funke, K. Dose-Dependence of Changes in Cortical Protein Expression Induced with Repeated Transcranial Magnetic Theta-Burst Stimulation in the Rat. Brain Stimul. 2013, 6, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Mix, A.; Hoppenrath, K.; Funke, K. Reduction in Cortical Parvalbumin Expression Due to Intermittent Theta-Burst Stimulation Correlates with Maturation of the Perineuronal Nets in Young Rats. Dev. Neurobiol. 2015, 75, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-Z.; Rothwell, J.C.; Edwards, M.J.; Chen, R.-S. Effect of Physiological Activity on an NMDA-Dependent Form of Cortical Plasticity in Human. Cereb. Cortex 2008, 18, 563–570. [Google Scholar] [CrossRef]
- Lenz, M.; Galanis, C.; Müller-Dahlhaus, F.; Opitz, A.; Wierenga, C.J.; Szabó, G.; Ziemann, U.; Deller, T.; Funke, K.; Vlachos, A. Repetitive Magnetic Stimulation Induces Plasticity of Inhibitory Synapses. Nat. Commun. 2016, 7, 10020. [Google Scholar] [CrossRef]
- Benali, A.; Trippe, J.; Weiler, E.; Mix, A.; Petrasch-Parwez, E.; Girzalsky, W.; Eysel, U.T.; Erdmann, R.; Funke, K. Theta-Burst Transcranial Magnetic Stimulation Alters Cortical Inhibition. J. Neurosci. 2011, 31, 1193–1203. [Google Scholar] [CrossRef]
- Thimm, A.; Funke, K. Multiple Blocks of Intermittent and Continuous Theta-Burst Stimulation Applied via Transcranial Magnetic Stimulation Differently Affect Sensory Responses in Rat Barrel Cortex. J. Physiol. 2015, 593, 967–985. [Google Scholar] [CrossRef]
- Tomar, M.; Rodger, J.; Moretti, J. Dorsal Striatum C-Fos Activity in Perseverative Ephrin-A2A5−/− Mice and the Cellular Effect of Low-Intensity rTMS. Front. Neural Circuits 2023, 17, 1179096. [Google Scholar] [CrossRef] [PubMed]
- Hoppenrath, K.; Härtig, W.; Funke, K. Intermittent Theta-Burst Transcranial Magnetic Stimulation Alters Electrical Properties of Fast-Spiking Neocortical Interneurons in an Age-Dependent Fashion. Front. Neural Circuits 2016, 10, 22. [Google Scholar] [CrossRef]
- Boyer, M.; Baudin, P.; Stengel, C.; Valero-Cabré, A.; Lohof, A.M.; Charpier, S.; Sherrard, R.M.; Mahon, S. In Vivo Low-Intensity Magnetic Pulses Durably Alter Neocortical Neuron Excitability and Spontaneous Activity. J. Physiol. 2022, 600, 4019–4037. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, J.; Sorrell, M.E.; Celnik, P.A.; Pelled, G. Immediate Effects of Repetitive Magnetic Stimulation on Single Cortical Pyramidal Neurons. PLoS ONE 2017, 12, e0170528. [Google Scholar] [CrossRef] [PubMed]
- Kozyrev, V.; Eysel, U.T.; Jancke, D. Voltage-Sensitive Dye Imaging of Transcranial Magnetic Stimulation-Induced Intracortical Dynamics. Proc. Natl. Acad. Sci. USA 2014, 111, 13553–13558. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Zhang, G.; Weng, X.; Han, Y.; Lang, Y.; Zhao, Y.; Zhao, X.; Wang, K.; Lin, Q.; Wang, C. In Vitro Assessment Reveals Parameters-Dependent Modulation on Excitability and Functional Connectivity of Cerebellar Slice by Repetitive Transcranial Magnetic Stimulation. Sci. Rep. 2016, 6, 23420. [Google Scholar] [CrossRef]
- Wang, H.-L.; Xian, X.-H.; Wang, Y.-Y.; Geng, Y.; Han, B.; Wang, M.-W.; Li, W.-B. Chronic High-Frequency Repetitive Transcranial Magnetic Stimulation Improves Age-Related Cognitive Impairment in Parallel with Alterations in Neuronal Excitability and the Voltage-Dependent Ca2+ Current in Female Mice. Neurobiol. Learn. Mem. 2015, 118, 1–7. [Google Scholar] [CrossRef]
- Power, J.M.; Wu, W.W.; Sametsky, E.; Oh, M.M.; Disterhoft, J.F. Age-Related Enhancement of the Slow Outward Calcium-Activated Potassium Current in Hippocampal CA1 Pyramidal Neurons in Vitro. J. Neurosci. 2002, 22, 7234–7243. [Google Scholar] [CrossRef]
- Burgdorf, J.; Kroes, R.A.; Weiss, C.; Oh, M.M.; Disterhoft, J.F.; Brudzynski, S.M.; Panksepp, J.; Moskal, J.R. Positive Emotional Learning Is Regulated in the Medial Prefrontal Cortex by GluN2B-Containing NMDA Receptors. Neuroscience 2011, 192, 515–523. [Google Scholar] [CrossRef]
- Disterhoft, J.F.; Thompson, L.T.; Moyer, J.R.; Mogul, D.J. Calcium-Dependent Afterhyperpolarization and Learning in Young and Aging Hippocampus. Life Sci. 1996, 59, 413–420. [Google Scholar] [CrossRef]
- Rose, G.M.; Ong, V.S.; Woodruff-Pak, D.S. Efficacy of MEM 1003, a Novel Calcium Channel Blocker, in Delay and Trace Eyeblink Conditioning in Older Rabbits. Neurobiol. Aging 2007, 28, 766–773. [Google Scholar] [CrossRef] [PubMed]
- Dodson, C.A.; Hore, P.J.; Wallace, M.I. A Radical Sense of Direction: Signalling and Mechanism in Cryptochrome Magnetoreception. Trends Biochem. Sci. 2013, 38, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Nießner, C.; Denzau, S.; Peichl, L.; Wiltschko, W.; Wiltschko, R. Magnetoreception in Birds: I. Immunohistochemical Studies Concerning the Cryptochrome Cycle. J. Exp. Biol. 2014, 217, 4221–4224. [Google Scholar] [CrossRef] [PubMed]
- Rodgers, C.T.; Hore, P.J. Chemical Magnetoreception in Birds: The Radical Pair Mechanism. Proc. Natl. Acad. Sci. USA 2009, 106, 353–360. [Google Scholar] [CrossRef] [PubMed]
- van der Horst, G.T.; Muijtjens, M.; Kobayashi, K.; Takano, R.; Kanno, S.; Takao, M.; de Wit, J.; Verkerk, A.; Eker, A.P.; van Leenen, D.; et al. Mammalian Cry1 and Cry2 Are Essential for Maintenance of Circadian Rhythms. Nature 1999, 398, 627–630. [Google Scholar] [CrossRef] [PubMed]
- Chaves, I.; Pokorny, R.; Byrdin, M.; Hoang, N.; Ritz, T.; Brettel, K.; Essen, L.-O.; van der Horst, G.T.J.; Batschauer, A.; Ahmad, M. The Cryptochromes: Blue Light Photoreceptors in Plants and Animals. Annu. Rev. Plant Biol. 2011, 62, 335–364. [Google Scholar] [CrossRef] [PubMed]
- Wiltschko, R.; Wiltschko, W. Sensing Magnetic Directions in Birds: Radical Pair Processes Involving Cryptochrome. Biosensors 2014, 4, 221–242. [Google Scholar] [CrossRef]
- Maeda, K.; Robinson, A.J.; Henbest, K.B.; Hogben, H.J.; Biskup, T.; Ahmad, M.; Schleicher, E.; Weber, S.; Timmel, C.R.; Hore, P.J. Magnetically Sensitive Light-Induced Reactions in Cryptochrome Are Consistent with Its Proposed Role as a Magnetoreceptor. Proc. Natl. Acad. Sci. USA 2012, 109, 4774–4779. [Google Scholar] [CrossRef]
- Solov’yov, I.A.; Chandler, D.E.; Schulten, K. Magnetic Field Effects in Arabidopsis Thaliana Cryptochrome-1. Biophys. J. 2007, 92, 2711–2726. [Google Scholar] [CrossRef]
- Zaporozhan, V.; Ponomarenko, A. Mechanisms of Geomagnetic Field Influence on Gene Expression Using Influenza as a Model System: Basics of Physical Epidemiology. Int. J. Environ. Res. Public Health 2010, 7, 938–965. [Google Scholar] [CrossRef]
- Kattnig, D.R.; Solov’yov, I.A.; Hore, P.J. Electron Spin Relaxation in Cryptochrome-Based Magnetoreception. Phys. Chem. Chem. Phys. 2016, 18, 12443–12456. [Google Scholar] [CrossRef]
- Consentino, L.; Lambert, S.; Martino, C.; Jourdan, N.; Bouchet, P.-E.; Witczak, J.; Castello, P.; El-Esawi, M.; Corbineau, F.; d’Harlingue, A.; et al. Blue-Light Dependent Reactive Oxygen Species Formation by Arabidopsis Cryptochrome May Define a Novel Evolutionarily Conserved Signaling Mechanism. New Phytol. 2015, 206, 1450–1462. [Google Scholar] [CrossRef]
- Pooam, M.; Arthaut, L.-D.; Burdick, D.; Link, J.; Martino, C.F.; Ahmad, M. Magnetic Sensitivity Mediated by the Arabidopsis Blue-Light Receptor Cryptochrome Occurs during Flavin Reoxidation in the Dark. Planta 2019, 249, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, M.-O.; Simkó, M. Grouping of Experimental Conditions as an Approach to Evaluate Effects of Extremely Low-Frequency Magnetic Fields on Oxidative Response in in Vitro Studies. Front. Public Health 2014, 2, 132. [Google Scholar] [CrossRef]
- Sherrard, R.M.; Morellini, N.; Jourdan, N.; El-Esawi, M.; Arthaut, L.-D.; Niessner, C.; Rouyer, F.; Klarsfeld, A.; Doulazmi, M.; Witczak, J.; et al. Low-Intensity Electromagnetic Fields Induce Human Cryptochrome to Modulate Intracellular Reactive Oxygen Species. PLoS Biol. 2018, 16, e2006229. [Google Scholar] [CrossRef]
- Nordmann, G.C.; Hochstoeger, T.; Keays, D.A. Magnetoreception-A Sense without a Receptor. PLoS Biol. 2017, 15, e2003234. [Google Scholar] [CrossRef]
- Usselman, R.J.; Hill, I.; Singel, D.J.; Martino, C.F. Spin Biochemistry Modulates Reactive Oxygen Species (ROS) Production by Radio Frequency Magnetic Fields. PLoS ONE 2014, 9, e93065. [Google Scholar] [CrossRef]
- El-Esawi, M.; Arthaut, L.-D.; Jourdan, N.; d’Harlingue, A.; Link, J.; Martino, C.F.; Ahmad, M. Blue-Light Induced Biosynthesis of ROS Contributes to the Signaling Mechanism of Arabidopsis Cryptochrome. Sci. Rep. 2017, 7, 13875. [Google Scholar] [CrossRef] [PubMed]
- Di Loreto, S.; Falone, S.; Caracciolo, V.; Sebastiani, P.; D’Alessandro, A.; Mirabilio, A.; Zimmitti, V.; Amicarelli, F. Fifty Hertz Extremely Low-Frequency Magnetic Field Exposure Elicits Redox and Trophic Response in Rat-Cortical Neurons. J. Cell. Physiol. 2009, 219, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Selaković, V.; Rauš Balind, S.; Radenović, L.; Prolić, Z.; Janać, B. Age-Dependent Effects of ELF-MF on Oxidative Stress in the Brain of Mongolian Gerbils. Cell Biochem. Biophys. 2013, 66, 513–521. [Google Scholar] [CrossRef] [PubMed]
- Görlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A Mutual Interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef]
- Rauš Balind, S.; Selaković, V.; Radenović, L.; Prolić, Z.; Janać, B. Extremely Low Frequency Magnetic Field (50 Hz, 0.5 mT) Reduces Oxidative Stress in the Brain of Gerbils Submitted to Global Cerebral Ischemia. PLoS ONE 2014, 9, e88921. [Google Scholar] [CrossRef]
- Morabito, C.; Rovetta, F.; Bizzarri, M.; Mazzoleni, G.; Fanò, G.; Mariggiò, M.A. Modulation of Redox Status and Calcium Handling by Extremely Low Frequency Electromagnetic Fields in C2C12 Muscle Cells: A Real-Time, Single-Cell Approach. Free Radic. Biol. Med. 2010, 48, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Pilla, A.; Fitzsimmons, R.; Muehsam, D.; Wu, J.; Rohde, C.; Casper, D. Electromagnetic Fields as First Messenger in Biological Signaling: Application to Calmodulin-Dependent Signaling in Tissue Repair. Biochim. Biophys. Acta 2011, 1810, 1236–1245. [Google Scholar] [CrossRef] [PubMed]
- Grassi, C.; D’Ascenzo, M.; Torsello, A.; Martinotti, G.; Wolf, F.; Cittadini, A.; Azzena, G.B. Effects of 50 Hz Electromagnetic Fields on Voltage-Gated Ca2+ Channels and Their Role in Modulation of Neuroendocrine Cell Proliferation and Death. Cell Calcium 2004, 35, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Morgado-Valle, C.; Verdugo-Díaz, L.; García, D.E.; Morales-Orozco, C.; Drucker-Colín, R. The Role of Voltage-Gated Ca2+ Channels in Neurite Growth of Cultured Chromaffin Cells Induced by Extremely Low Frequency (ELF) Magnetic Field Stimulation. Cell Tissue Res. 1998, 291, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Piacentini, R.; Ripoli, C.; Mezzogori, D.; Azzena, G.B.; Grassi, C. Extremely Low-Frequency Electromagnetic Fields Promote in Vitro Neurogenesis via Upregulation of Ca(v)1-Channel Activity. J. Cell. Physiol. 2008, 215, 129–139. [Google Scholar] [CrossRef]
- Cooper, L.L.; Li, W.; Lu, Y.; Centracchio, J.; Terentyeva, R.; Koren, G.; Terentyev, D. Redox Modification of Ryanodine Receptors by Mitochondria-Derived Reactive Oxygen Species Contributes to Aberrant Ca2+ Handling in Ageing Rabbit Hearts. J Physiol 2013, 591, 5895–5911. [Google Scholar] [CrossRef]
- Brini, M.; Calì, T.; Ottolini, D.; Carafoli, E. Neuronal Calcium Signaling: Function and Dysfunction. Cell. Mol. Life Sci. 2014, 71, 2787–2814. [Google Scholar] [CrossRef]
- Solà, C.; Barrón, S.; Tusell, J.M.; Serratosa, J. The Ca2+/Calmodulin Signaling System in the Neural Response to Excitability. Involvement of Neuronal and Glial Cells. Prog. Neurobiol. 1999, 58, 207–232. [Google Scholar] [CrossRef]
- Cornelisse, L.N.; van Elburg, R.A.J.; Meredith, R.M.; Yuste, R.; Mansvelder, H.D. High Speed Two-Photon Imaging of Calcium Dynamics in Dendritic Spines: Consequences for Spine Calcium Kinetics and Buffer Capacity. PLoS ONE 2007, 2, e1073. [Google Scholar] [CrossRef]
- Baek, A.; Park, E.J.; Kim, S.Y.; Nam, B.-G.; Kim, J.H.; Jun, S.W.; Kim, S.H.; Cho, S.-R. High-Frequency Repetitive Magnetic Stimulation Enhances the Expression of Brain-Derived Neurotrophic Factor Through Activation of Ca2+-Calmodulin-Dependent Protein Kinase II-cAMP-Response Element-Binding Protein Pathway. Front. Neurol. 2018, 9, 285. [Google Scholar] [CrossRef]
- Cho, S.I.; Nam, Y.S.; Chu, L.Y.; Lee, J.H.; Bang, J.S.; Kim, H.R.; Kim, H.-C.; Lee, Y.J.; Kim, H.-D.; Sul, J.D.; et al. Extremely Low-Frequency Magnetic Fields Modulate Nitric Oxide Signaling in Rat Brain. Bioelectromagnetics 2012, 33, 568–574. [Google Scholar] [CrossRef]
- Brookes, P.S.; Salinas, E.P.; Darley-Usmar, K.; Eiserich, J.P.; Freeman, B.A.; Darley-Usmar, V.M.; Anderson, P.G. Concentration-Dependent Effects of Nitric Oxide on Mitochondrial Permeability Transition and Cytochrome c Release. J. Biol. Chem. 2000, 275, 20474–20479. [Google Scholar] [CrossRef] [PubMed]
- Demirkazik, A.; Ozdemir, E.; Arslan, G.; Taskiran, A.S.; Pelit, A. The Effects of Extremely Low-Frequency Pulsed Electromagnetic Fields on Analgesia in the Nitric Oxide Pathway. Nitric Oxide 2019, 92, 49–54. [Google Scholar] [CrossRef]
- Jones, M.W.; Errington, M.L.; French, P.J.; Fine, A.; Bliss, T.V.P.; Garel, S.; Charnay, P.; Bozon, B.; Laroche, S.; Davis, S. A Requirement for the Immediate Early Gene Zif268 in the Expression of Late LTP and Long-Term Memories. Nat. Neurosci. 2001, 4, 289–296. [Google Scholar] [CrossRef] [PubMed]
- Sng, J.C.G.; Taniura, H.; Yoneda, Y. A Tale of Early Response Genes. Biol. Pharm. Bull. 2004, 27, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.B.; Toschi, N.; Kresse, A.E.; Post, A.; Keck, M.E. Long-Term Repetitive Transcranial Magnetic Stimulation Increases the Expression of Brain-Derived Neurotrophic Factor and Cholecystokinin mRNA, but Not Neuropeptide Tyrosine mRNA in Specific Areas of Rat Brain. Neuropsychopharmacology 2000, 23, 205. [Google Scholar] [CrossRef]
- Hellmann, J.; Jüttner, R.; Roth, C.; Bajbouj, M.; Kirste, I.; Heuser, I.; Gertz, K.; Endres, M.; Kronenberg, G. Repetitive Magnetic Stimulation of Human-Derived Neuron-like Cells Activates cAMP-CREB Pathway. Eur. Arch. Psychiatry Clin. Neurosci. 2012, 262, 87–91. [Google Scholar] [CrossRef]
- Hoppenrath, K.; Funke, K. Time-Course of Changes in Neuronal Activity Markers Following iTBS-TMS of the Rat Neocortex. Neurosci. Lett. 2013, 536, 19–23. [Google Scholar] [CrossRef]
- Lu, B.; Nagappan, G.; Guan, X.; Nathan, P.J.; Wren, P. BDNF-Based Synaptic Repair as a Disease-Modifying Strategy for Neurodegenerative Diseases. Nat. Rev. Neurosci. 2013, 14, 401–416. [Google Scholar] [CrossRef] [PubMed]
- Binder, D.K.; Scharfman, H.E. Brain-Derived Neurotrophic Factor. Growth Factors 2004, 22, 123–131. [Google Scholar] [CrossRef]
- Kovalchuk, Y.; Hanse, E.; Kafitz, K.W.; Konnerth, A. Postsynaptic Induction of BDNF-Mediated Long-Term Potentiation. Science 2002, 295, 1729–1734. [Google Scholar] [CrossRef] [PubMed]
- Savalli, G.; Diao, W.; Berger, S.; Ronovsky, M.; Partonen, T.; Pollak, D.D. Anhedonic Behavior in Cryptochrome 2-Deficient Mice Is Paralleled by Altered Diurnal Patterns of Amygdala Gene Expression. Amino Acids 2015, 47, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Cheeran, B.; Talelli, P.; Mori, F.; Koch, G.; Suppa, A.; Edwards, M.; Houlden, H.; Bhatia, K.; Greenwood, R.; Rothwell, J.C. A Common Polymorphism in the Brain-Derived Neurotrophic Factor Gene (BDNF) Modulates Human Cortical Plasticity and the Response to rTMS. J. Physiol. 2008, 586, 5717–5725. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, G.; Di Pino, G.; Capone, F.; Ranieri, F.; Florio, L.; Todisco, V.; Tedeschi, G.; Funke, K.; Di Lazzaro, V. Neurobiological After-Effects of Non-Invasive Brain Stimulation. Brain Stimul. 2017, 10, 1–18. [Google Scholar] [CrossRef]
- Gedge, L.; Beaudoin, A.; Lazowski, L.; du Toit, R.; Jokic, R.; Milev, R. Effects of Electroconvulsive Therapy and Repetitive Transcranial Magnetic Stimulation on Serum Brain-Derived Neurotrophic Factor Levels in Patients with Depression. Front. Psychiatry 2012, 3, 12. [Google Scholar] [CrossRef]
- Yukimasa, T.; Yoshimura, R.; Tamagawa, A.; Uozumi, T.; Shinkai, K.; Ueda, N.; Tsuji, S.; Nakamura, J. High-Frequency Repetitive Transcranial Magnetic Stimulation Improves Refractory Depression by Influencing Catecholamine and Brain-Derived Neurotrophic Factors. Pharmacopsychiatry 2006, 39, 52–59. [Google Scholar] [CrossRef]
- Zanardini, R.; Gazzoli, A.; Ventriglia, M.; Perez, J.; Bignotti, S.; Rossini, P.M.; Gennarelli, M.; Bocchio-Chiavetto, L. Effect of Repetitive Transcranial Magnetic Stimulation on Serum Brain Derived Neurotrophic Factor in Drug Resistant Depressed Patients. J. Affect. Disord. 2006, 91, 83–86. [Google Scholar] [CrossRef]
- Gersner, R.; Kravetz, E.; Feil, J.; Pell, G.; Zangen, A. Long-Term Effects of Repetitive Transcranial Magnetic Stimulation on Markers for Neuroplasticity: Differential Outcomes in Anesthetized and Awake Animals. J. Neurosci. 2011, 31, 7521. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, Z.; Su, Y.; Kang, L.; Geng, D.; Wang, Y.; Luan, F.; Wang, M.; Cui, H. Magnetic Stimulation Modulates Structural Synaptic Plasticity and Regulates BDNF-TrkB Signal Pathway in Cultured Hippocampal Neurons. Neurochem. Int. 2013, 62, 84–91. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, Z.; Kang, L.; Geng, D.; Wang, Y.; Wang, M.; Cui, H. Repetitive Transcranial Magnetic Stimulation (rTMS) Influences Spatial Cognition and Modulates Hippocampal Structural Synaptic Plasticity in Aging Mice. Exp. Gerontol. 2014, 58, 256–268. [Google Scholar] [CrossRef]
- Wang, F.; Geng, X.; Tao, H.-Y.; Cheng, Y. The Restoration After Repetitive Transcranial Magnetic Stimulation Treatment on Cognitive Ability of Vascular Dementia Rats and Its Impacts on Synaptic Plasticity in Hippocampal CA1 Area. J. Mol. Neurosci. 2010, 41, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Mei, Y.; Liu, C.; Yu, S. Effect of Transcranial Magnetic Stimulation on the Expression of C-Fos and Brain-Derived Neurotrophic Factor of the Cerebral Cortex in Rats with Cerebral Infarct. J. Huazhong Univ. Sci. Technol. 2007, 27, 415–418. [Google Scholar] [CrossRef]
- Morellini, N.; Grehl, S.; Tang, A.; Rodger, J.; Mariani, J.; Lohof, A.M.; Sherrard, R.M. What Does Low-Intensity rTMS Do to the Cerebellum? Cerebellum 2015, 14, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Ming, G.; Song, H. Adult Neurogenesis in the Mammalian Brain: Significant Answers and Significant Questions. Neuron 2011, 70, 687–702. [Google Scholar] [CrossRef]
- Abbasnia, K.; Ghanbari, A.; Abedian, M.; Ghanbari, A.; Sharififar, S.; Azari, H. The Effects of Repetitive Transcranial Magnetic Stimulation on Proliferation and Differentiation of Neural Stem Cells. Anat. Cell Biol. 2015, 48, 104–113. [Google Scholar] [CrossRef] [PubMed]
- Czéh, B.; Welt, T.; Fischer, A.K.; Erhardt, A.; Schmitt, W.; Müller, M.B.; Toschi, N.; Fuchs, E.; Keck, M.E. Chronic Psychosocial Stress and Concomitant Repetitive Transcranial Magnetic Stimulation: Effects on Stress Hormone Levels and Adult Hippocampal Neurogenesis. Biol. Psychiatry 2002, 52, 1057–1065. [Google Scholar] [CrossRef]
- Ueyama, E.; Ukai, S.; Ogawa, A.; Yamamoto, M.; Kawaguchi, S.; Ishii, R.; Shinosaki, K. Chronic Repetitive Transcranial Magnetic Stimulation Increases Hippocampal Neurogenesis in Rats. Psychiatry Clin. Neurosci. 2011, 65, 77–81. [Google Scholar] [CrossRef]
- Cuccurazzu, B.; Leone, L.; Podda, M.V.; Piacentini, R.; Riccardi, E.; Ripoli, C.; Azzena, G.B.; Grassi, C. Exposure to Extremely Low-Frequency (50 Hz) Electromagnetic Fields Enhances Adult Hippocampal Neurogenesis in C57BL/6 Mice. Exp. Neurol. 2010, 226, 173–182. [Google Scholar] [CrossRef]
- Fitzgerald, P. T008 rTMS in Depression—Is More Always Better? Clin. Neurophysiol. 2017, 128, e3. [Google Scholar] [CrossRef]
- Hedendahl, L.; Carlberg, M.; Hardell, L. Electromagnetic Hypersensitivity—An Increasing Challenge to the Medical Profession. Rev. Environ. Health 2015, 30, 209–215. [Google Scholar] [CrossRef]
- Jalilian, H.; Teshnizi, S.H.; Röösli, M.; Neghab, M. Occupational Exposure to Extremely Low Frequency Magnetic Fields and Risk of Alzheimer Disease: A Systematic Review and Meta-Analysis. Neurotoxicology 2018, 69, 242–252. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Phillips, B. A Pilot Study of the Use of EEG-Based Synchronized Transcranial Magnetic Stimulation (sTMS) for Treatment of Major Depression. BMC Psychiatry 2014, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Martiny, K.; Lunde, M.; Bech, P. Transcranial Low Voltage Pulsed Electromagnetic Fields in Patients with Treatment-Resistant Depression. Biol. Psychiatry 2010, 68, 163–169. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dufor, T.; Lohof, A.M.; Sherrard, R.M. Magnetic Stimulation as a Therapeutic Approach for Brain Modulation and Repair: Underlying Molecular and Cellular Mechanisms. Int. J. Mol. Sci. 2023, 24, 16456. https://doi.org/10.3390/ijms242216456
Dufor T, Lohof AM, Sherrard RM. Magnetic Stimulation as a Therapeutic Approach for Brain Modulation and Repair: Underlying Molecular and Cellular Mechanisms. International Journal of Molecular Sciences. 2023; 24(22):16456. https://doi.org/10.3390/ijms242216456
Chicago/Turabian StyleDufor, Tom, Ann M. Lohof, and Rachel M. Sherrard. 2023. "Magnetic Stimulation as a Therapeutic Approach for Brain Modulation and Repair: Underlying Molecular and Cellular Mechanisms" International Journal of Molecular Sciences 24, no. 22: 16456. https://doi.org/10.3390/ijms242216456