
Phytochemistry and Biological Studies of Endemic Hawaiian Plants
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Discussions
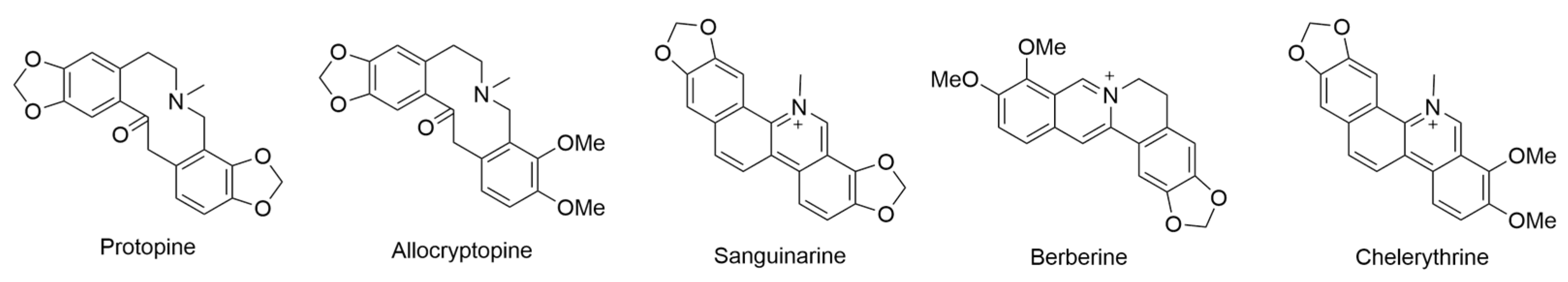
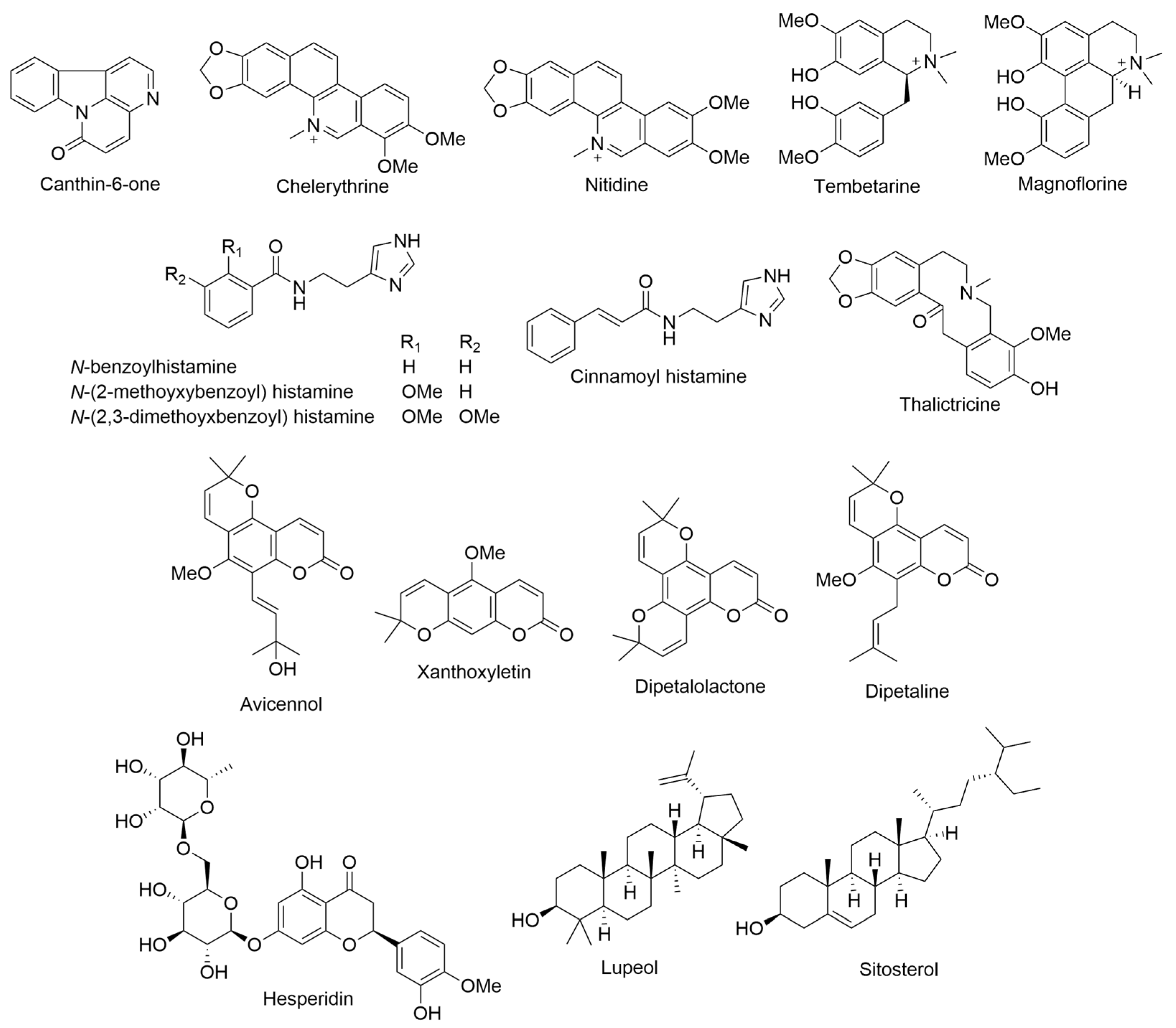
2.1. Argemone glauca L. ex Pope, var. glauca
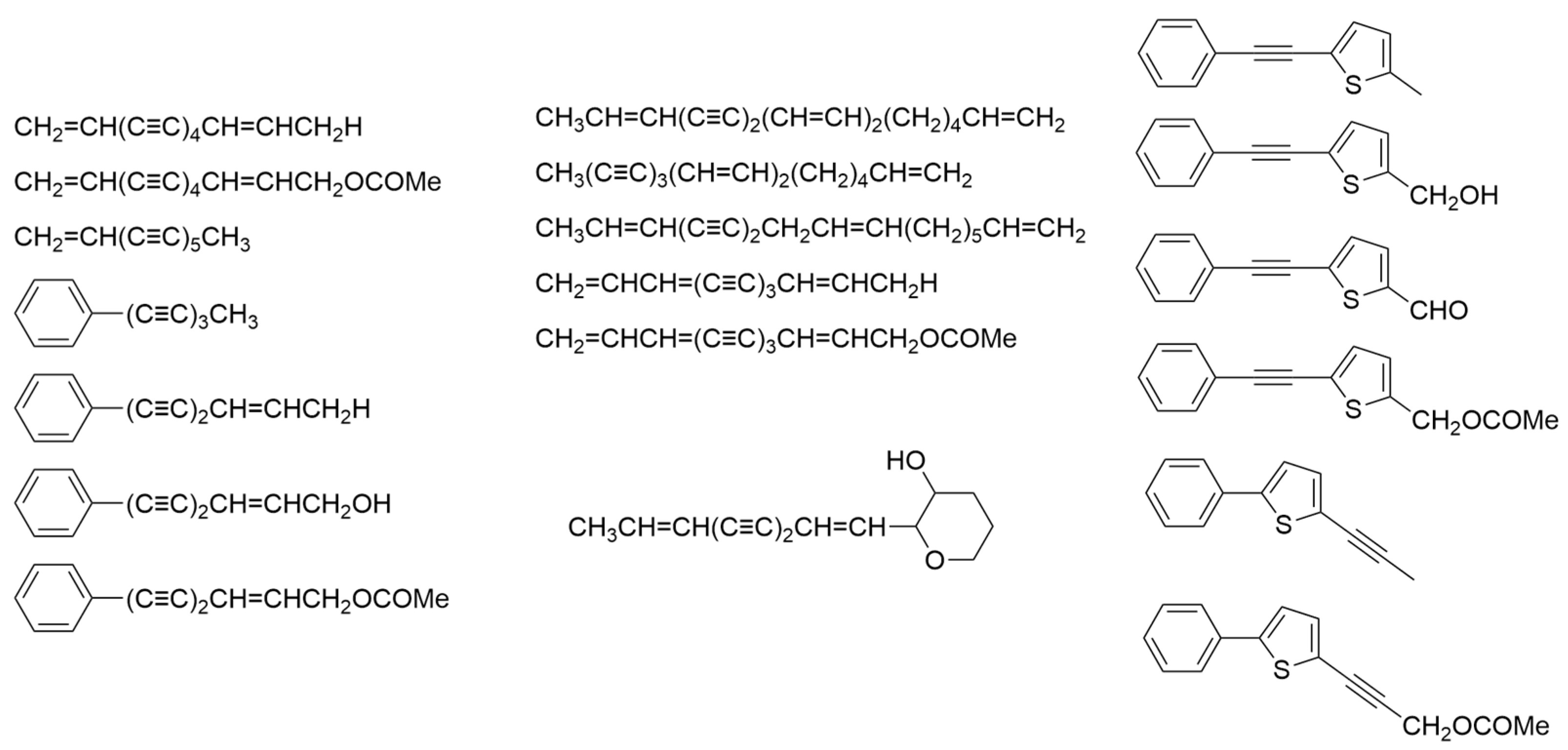
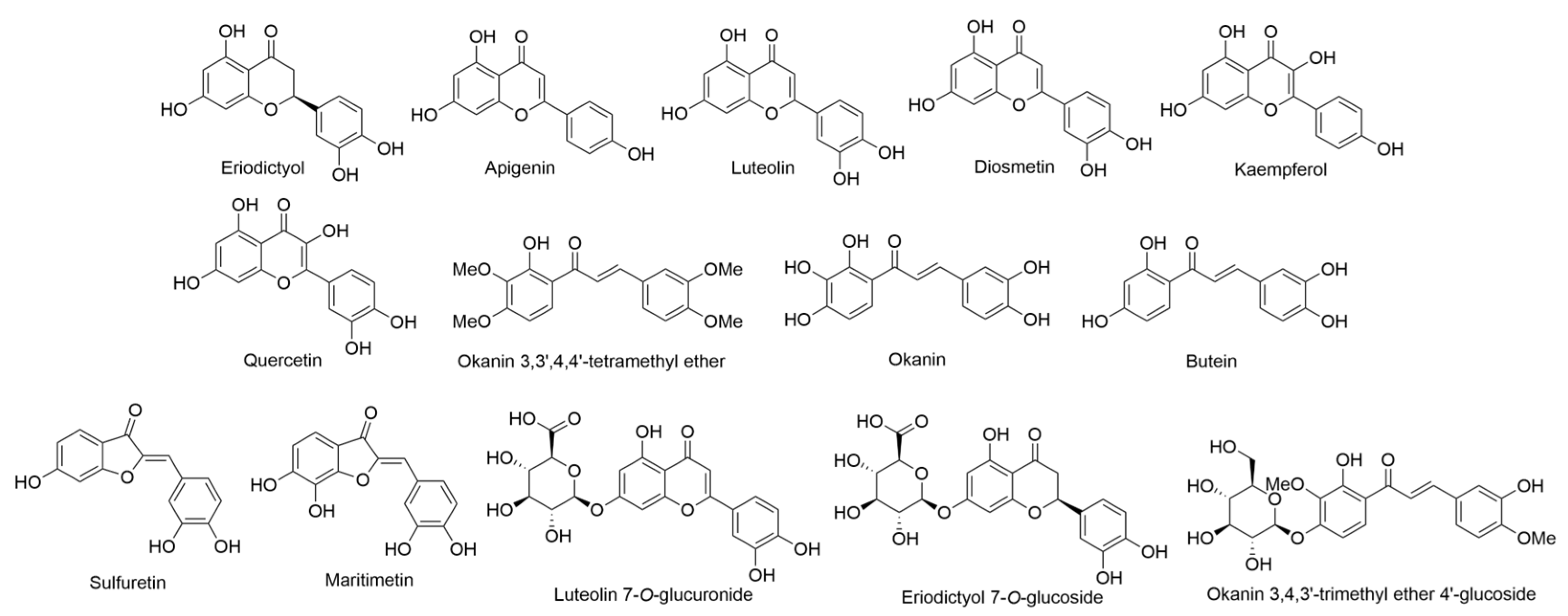
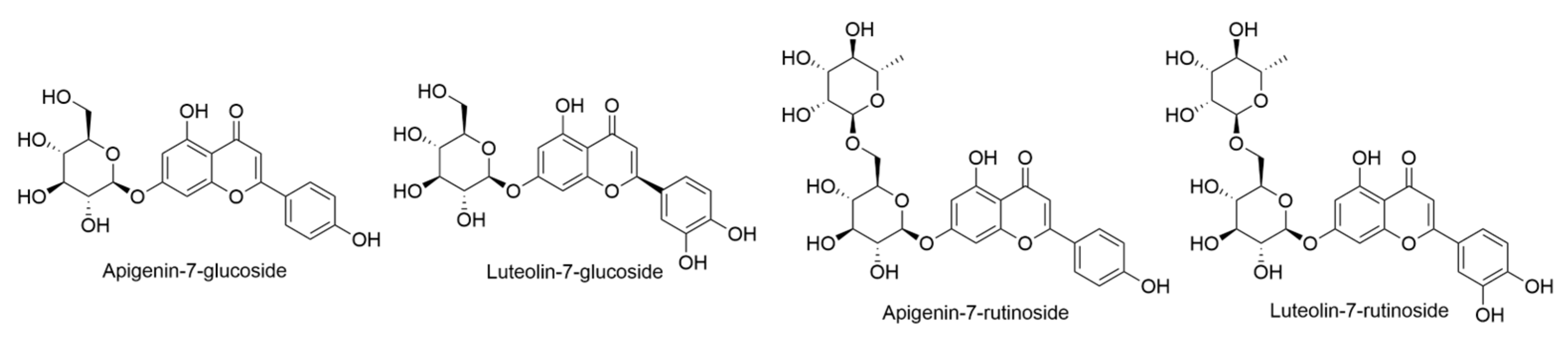
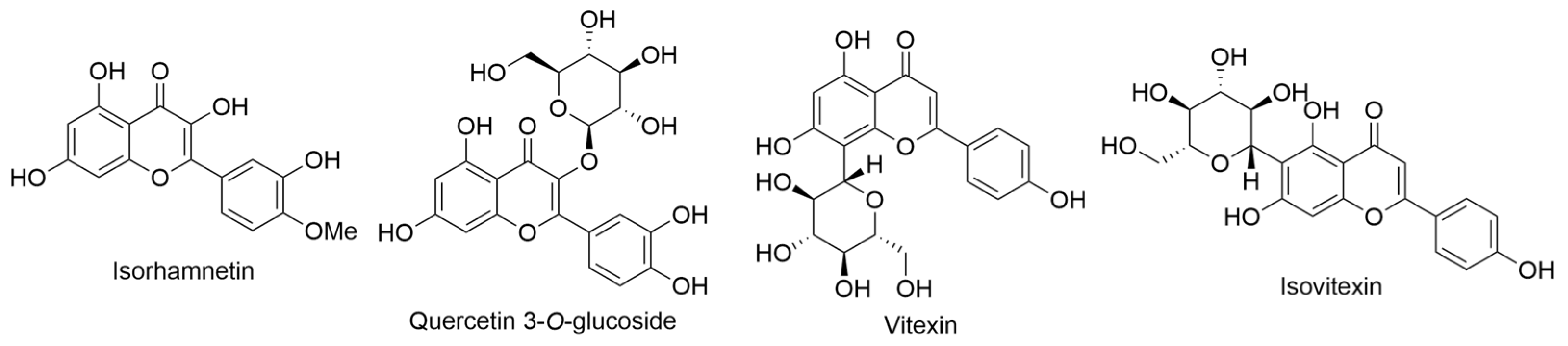
2.2. Bidens Genus
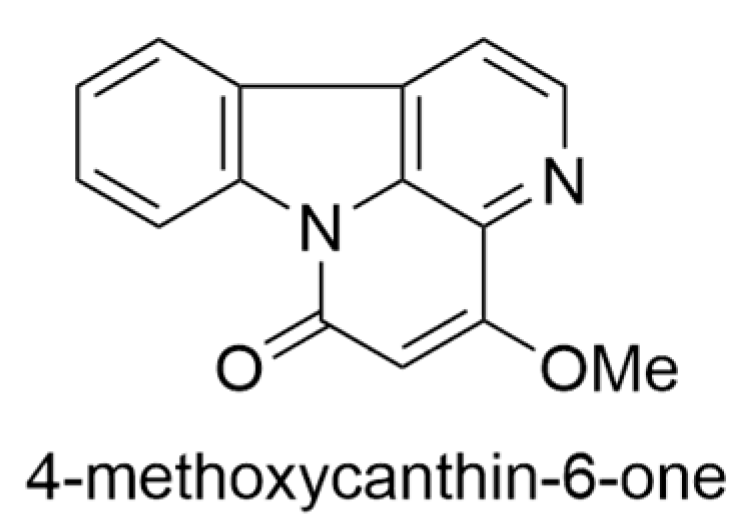
2.3. Charpentiera obovata Gaudich.
2.4. Clermontia persicifolia Gaudich.
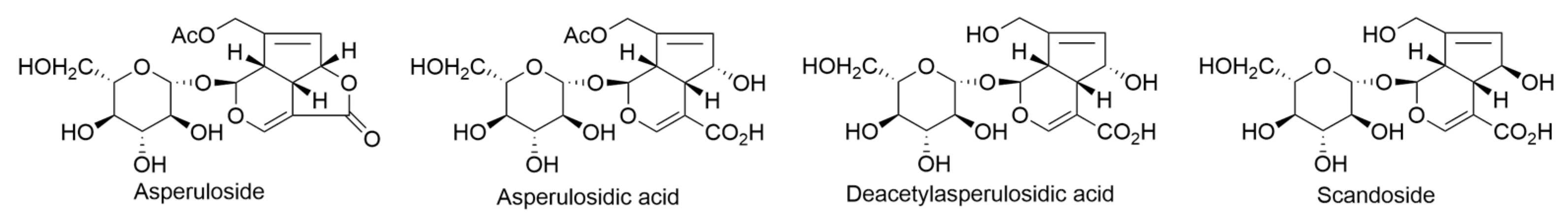
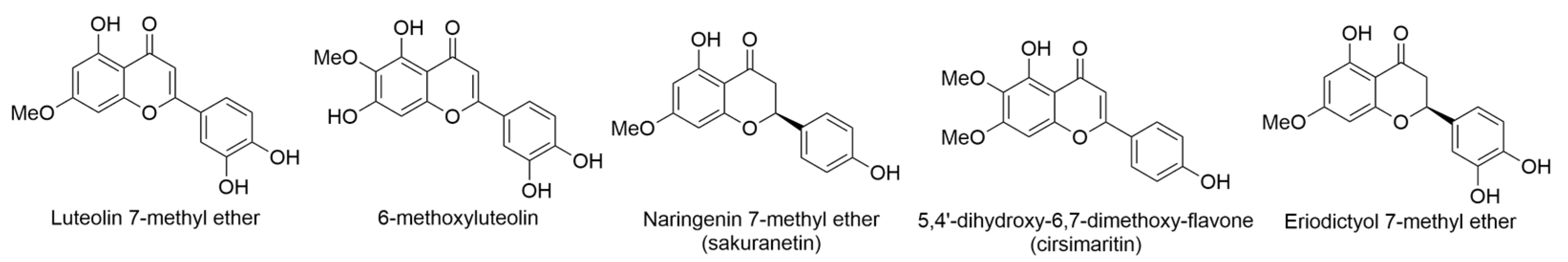
2.5. Coprosma ernodeoides A. Gray
2.6. Cuscuta sandwichiana Choisy
2.7. Dryopteris mauiensis C. Chr.
2.8. Dubautia arborea (A. Gray) D. D. Keck
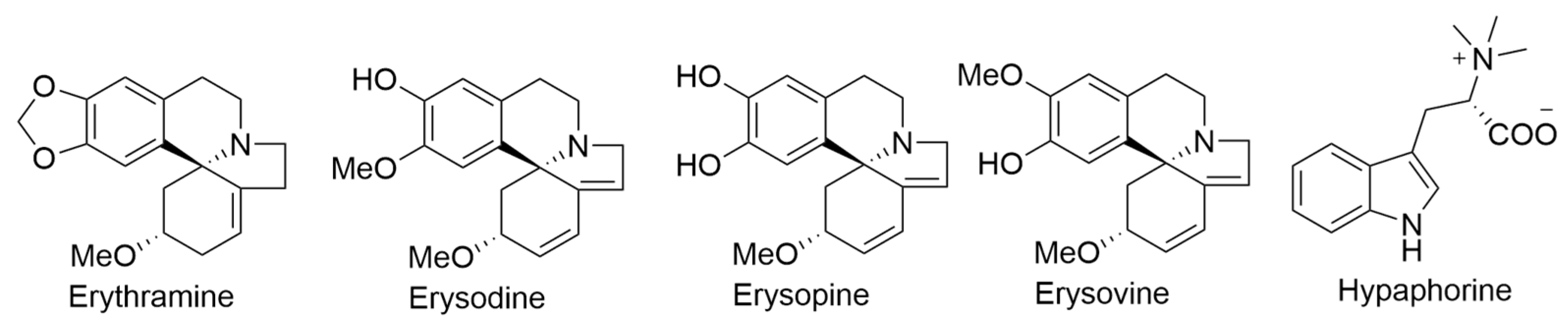
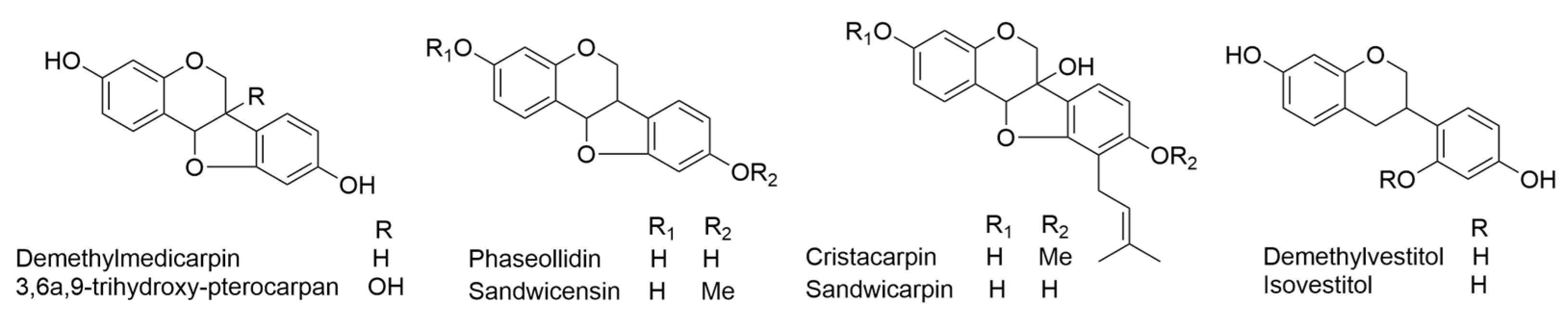
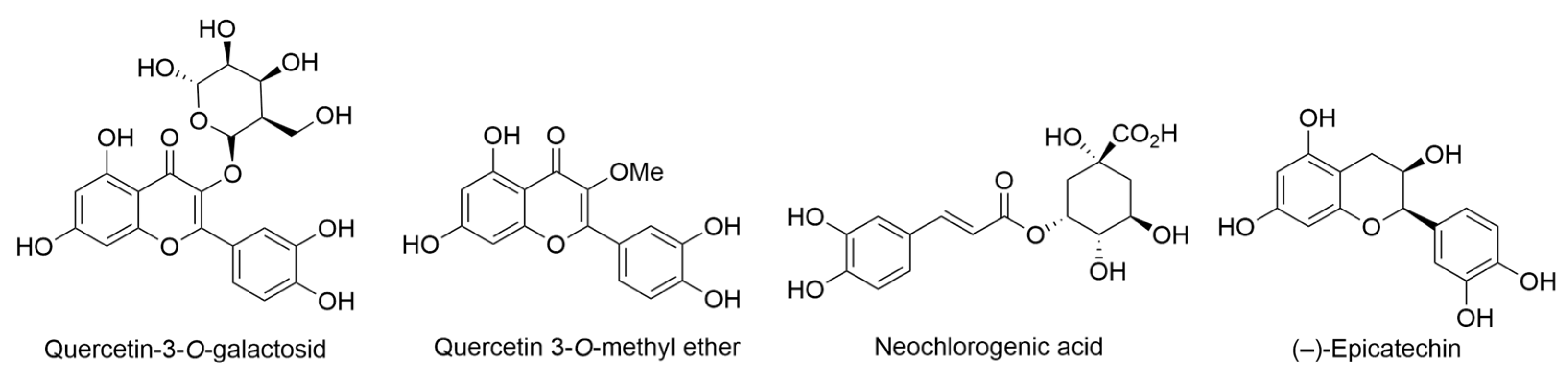
2.9. Erythrina sandwicensis O. Deg.
2.10. Gardenia brighamii H. Mann
2.11. Hesperomannia arborescens A. Gray
2.12. Hillebrandia sandwicensis Oliv.
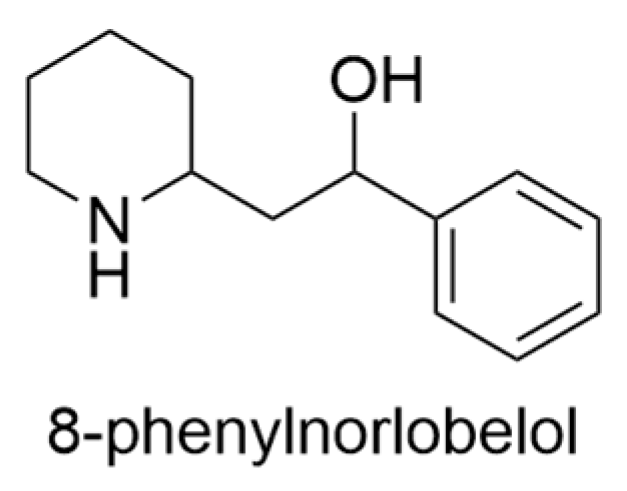
2.13. Lobelia yuccoides Hillebr.
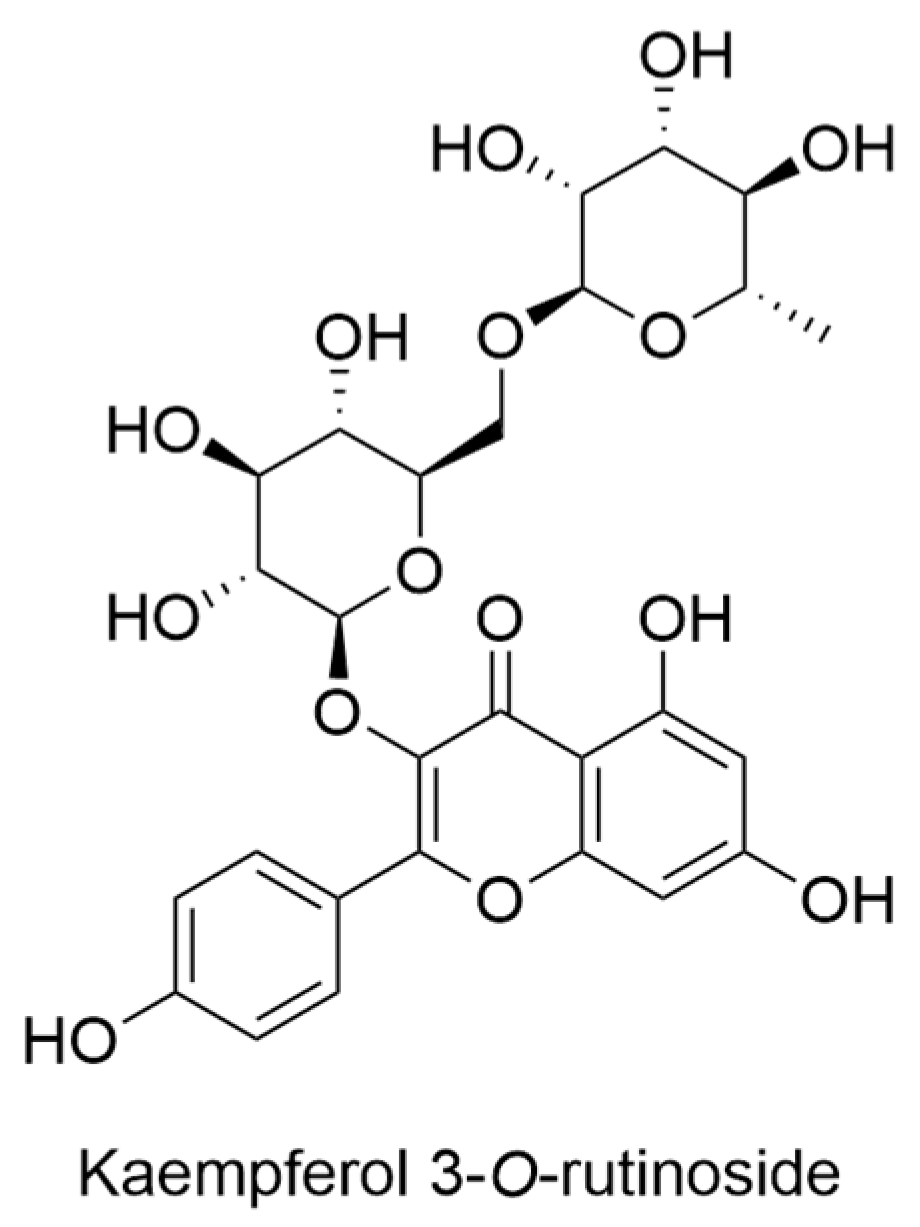
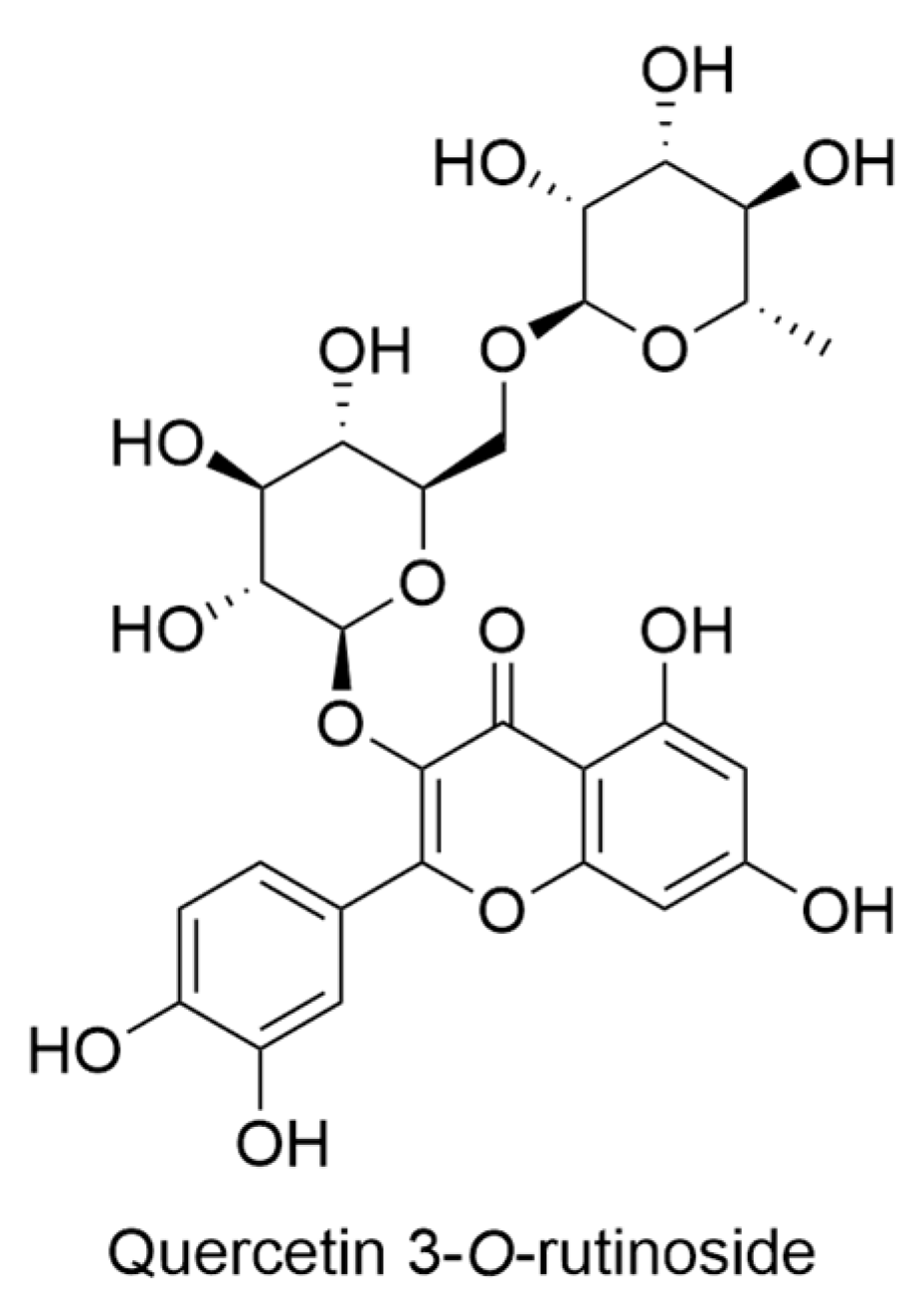
2.14. Lysimachia Genus
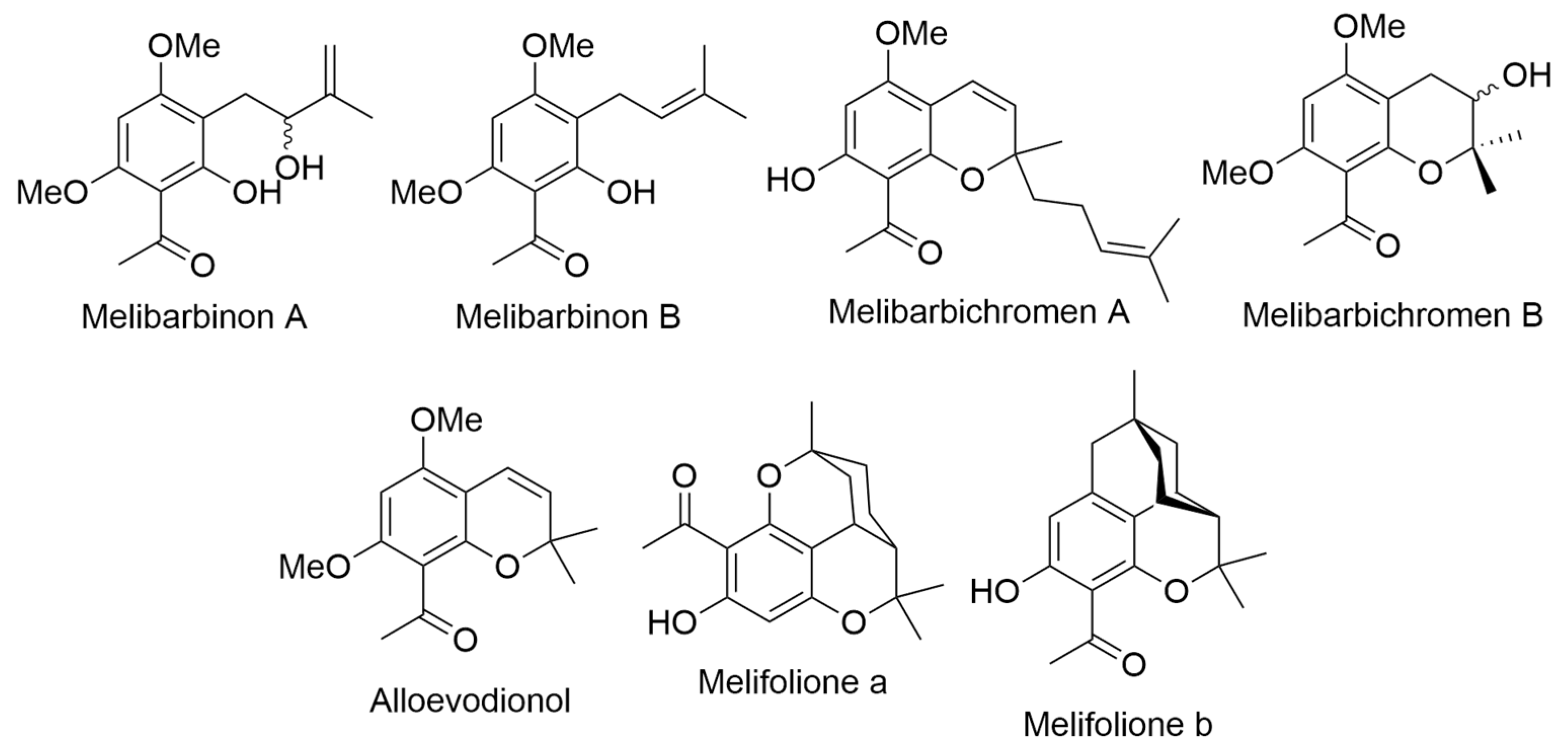
2.15. Melicope barbigera A. Gray (syn. Pelea barbigera Hillebr.)
2.16. Phyllanthus distichus Hook. & Arn.
2.17. Pipturus albidus A. Gray ex H. Mann
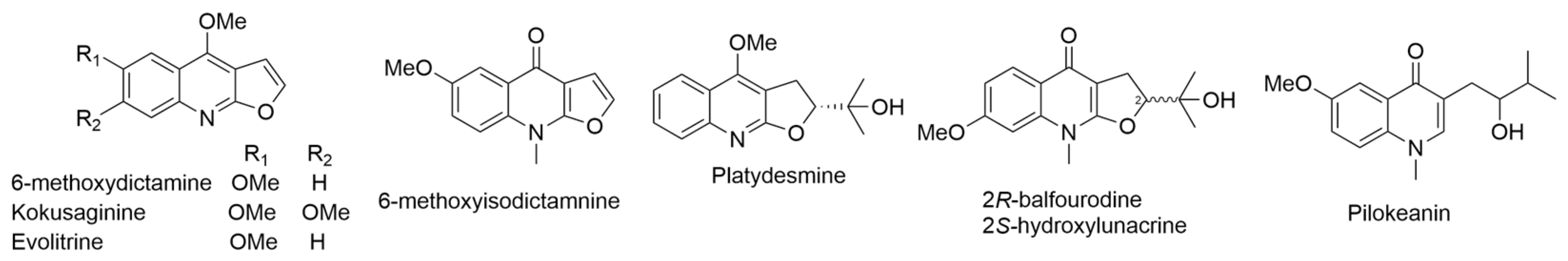
2.18. Platydesma campanulatum H. Mann or P. spathulatum (A. Gray) Skottsb. or P. campanulata H. Mann (syn. Melicope spathulata A. Gray)
2.19. Psychotria hawaiiensis (A. Gray) Fosberg
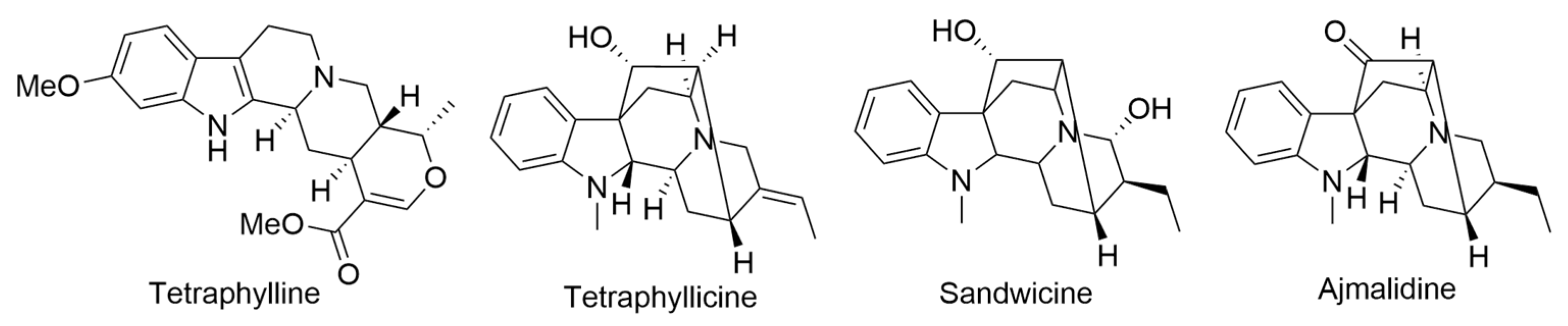
2.20. Rauvolfia sandwicensis A. DC.
2.21. Santalum paniculatum Hook. & Arn.
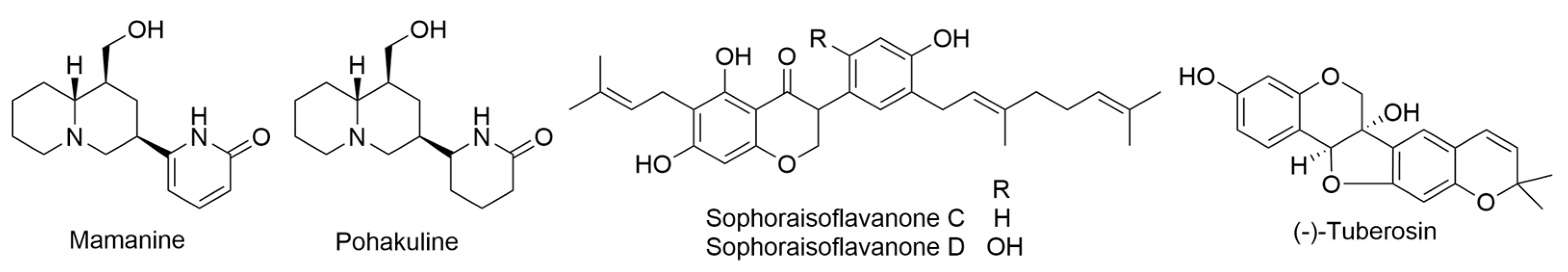
2.22. Sophora chrysophylla (Salisb.) Seem.
2.23. Vaccinium Genus
2.23.1. Vaccinium calycinum Sm.
2.23.2. Vaccinium reticulatum Sm.
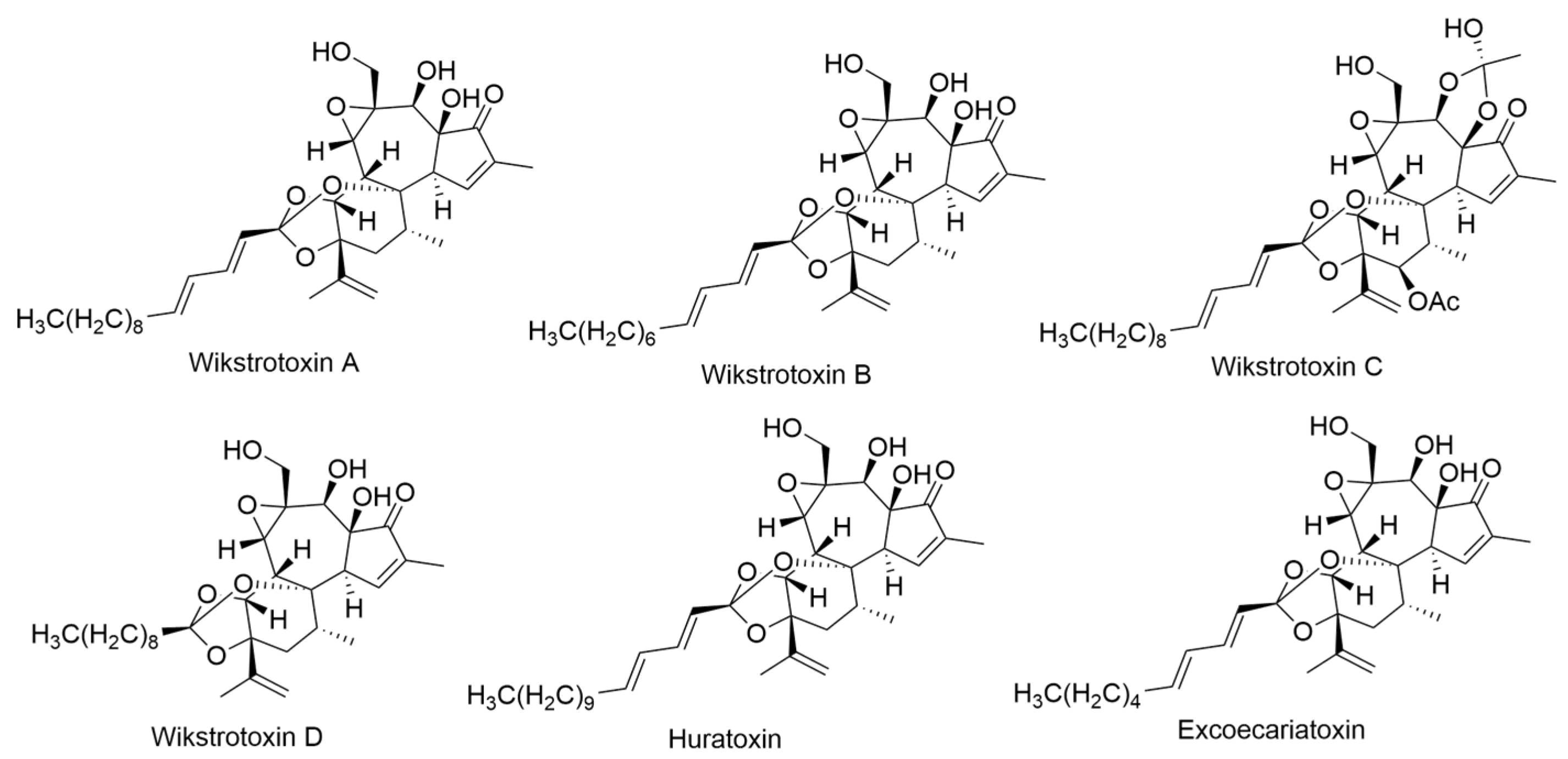
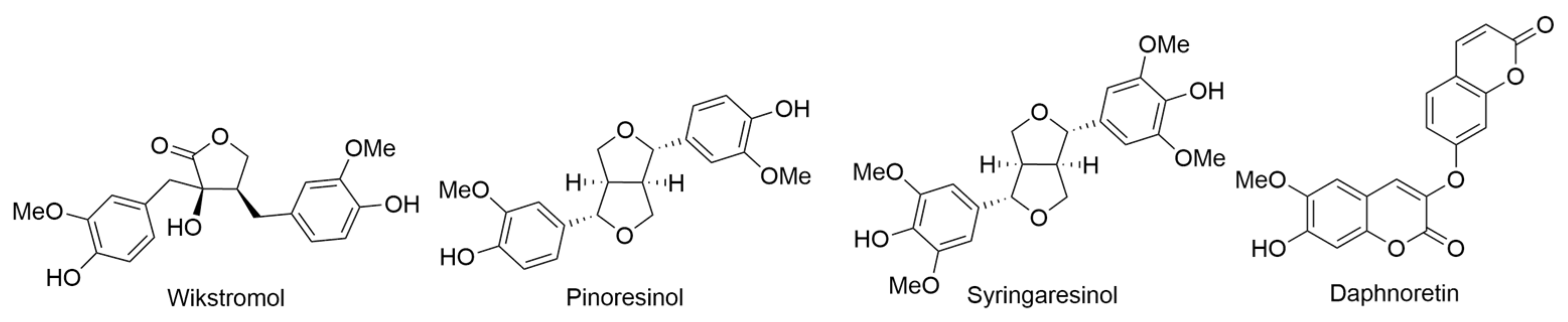
2.24. Wikstroemia Genus
2.24.1. Wikstroemia monticola Skottsb.
2.24.2. Wikstroemia uva-ursi A. Gray
2.25. Wilkesia Genus
2.25.1. Wilkesia gymnoxiphium A. Gray
2.25.2. Wilkesia hobdyi St. John
2.26. Zanthoxylum Genus
2.26.1. Zanthoxylum dipetalum H. Mann
2.26.2. Zanthoxylum hawaiiense Hillebr.
2.26.3. Zanthoxylum kauaense A. Gray
3. Conclusions
4. Perspectives
5. Methodology
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nagata, K.M. Hawaiian medicinal plants. Econ. Bot. 1971, 25, 245–254. [Google Scholar] [CrossRef]
- Kim, H.G.; Keeley, S.C.; Vroom, P.S.; Jansen, R.K. Molecular evidence for an African origin of the Hawaiian endemic Hesperomannia (Asteraceae). Proc. Natl. Acad. Sci. 1998, 95, 15440–15445. [Google Scholar] [CrossRef] [PubMed]
- Gemma, J.N.; Koske, R.E.; Habte, M. Mycorrhizal dependency of some endemic and endangered Hawaiian plant species. Am. J. Bot. 2002, 89, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Vernon, A.L.; Ranker, T.A. Current status of the ferns and lycophytes of the Hawaiian Islands. Am. Fern J. 2013, 103, 59–111. [Google Scholar] [CrossRef]
- Stone, B.C. A review of the endemic genera of Hawaiian plants. Bot. Rev. 1967, 33, 216–259. [Google Scholar] [CrossRef]
- Wagner, W.L.; Khan, N.R. Flora of the Hawaiian Islands Website. 2023. Available online: https://naturalhistory2.si.edu/botany/hawaiianflora/ (accessed on 11 September 2023).
- Bishop Museum. Ethnobotany Database Website. 2023. Available online: http://data.bishopmuseum.org/ethnobotanydb/ethnobotany.php?b=list&o=1 (accessed on 11 September 2023).
- Ownbey, G.B. The genus Argemone in South America and Hawaii. Brittonia 1961, 13, 91–109. [Google Scholar] [CrossRef]
- Stermitz, F.R.; Workman, S.M.; Klein, W.M. Alkaloids of Argemone glauca var. glauca. Phytochemistry 1971, 10, 675–677. [Google Scholar] [CrossRef]
- Knope, M.L.; Bellinger, M.R.; Datlof, E.M.; Gallaher, T.J.; Johnson, M.A. Insights into the evolutionary history of the Hawaiian Bidens (Asteraceae) adaptive radiation revealed through phylogenomics. J. Hered. 2020, 111, 119–137. [Google Scholar] [CrossRef]
- Sherff, E.E. Studies in the Genus Bidens. V. Bot. Gaz. 1920, 70, 89–109. [Google Scholar] [CrossRef]
- Marchant, Y.Y.; Ganders, F.R.; Wat, C.K.; Towers, G.N. Polyacetylenes in Hawaiian Bidens. Biochem. Syst. Ecol. 1984, 12, 167–178. [Google Scholar] [CrossRef]
- Ganders, F.R.; Bohm, B.A.; McCormick, S.P. Flavonoid variation in Hawaiian Bidens. Syst. Bot. 1990, 1, 231–239. [Google Scholar] [CrossRef]
- Pattabhiraman, T.R. An Investigation for Alkaloids in Charpentiera Obovata, Gaud; University of Hawai’i at Manoa: Honolulu, HI, USA, 1965. [Google Scholar]
- Crespi-Perellino, N.; Guicciardi, A.; Malyszko, G.; Arlandini, E.; Ballabio, M.; Minghetti, A. Occurrence of indole alkaloids in Ailanthus altissima cell cultures. J. Nat. Prod. 1986, 49, 1010–1014. [Google Scholar] [CrossRef]
- Givnish, T.J.; Bean, G.J.; Ames, M.; Lyon, S.P.; Sytsma, K.J. Phylogeny, floral evolution, and inter-island dispersal in Hawaiian Clermontia (Campanulaceae) based on ISSR variation and plastid spacer sequences. PLoS ONE 2013, 8, e62566. [Google Scholar] [CrossRef]
- Lammers, T.G. Systematics of Clermontia (Campanulaceae-Lobelioideae). Syst. Bot. Monogr. 1991, 20, 1–97. [Google Scholar] [CrossRef]
- Saleh, N.A.; Towers, G.H. Flavonoids of Clermontia persicifolia leaves. Phytochemistry 1972, 11, 1500. [Google Scholar] [CrossRef]
- Lang, M.; Zelta-Pinet, D.; Hagiwara, K.; Wright, A.D. The first natural products investigation of the Hawaiian endemic plant Coprosma ernodeoides or Nene Bush (‘Aiakanene). Planta. Med. 2015, 81, PX6. [Google Scholar] [CrossRef]
- Baldwin, P.H. Foods of the Hawaiian goose. Condor 1947, 49, 108–120. [Google Scholar] [CrossRef]
- Jang, G.F. New Cytotoxic Macrocyclic Gylcoresins from the Hawaiian Medicinal Plant “Kauna’oa”, Cuscuta Sandwichiana Choisy, Convolvulaceae; University of Hawaii at Manoa: Honolulu, HI, USA, 1990. [Google Scholar]
- Whistler, W.A. Polynesian Herbal Medicine; National Tropical Botanical Garden: Kalaheo, HI, USA, 1992. [Google Scholar]
- Yuncker, T.G. The genus Cuscuta. Mem. Torrey Bot. Club 1932, 18, 109–331. [Google Scholar]
- Costea, M.; Nesom, G.L.; Stefanović, S. Taxonomy of the Cuscuta pentagona complex (Convolvulaceae) in North America. SIDA Contrib. Bot. 2006, 11, 151–175. [Google Scholar]
- Locher, C.P.; Witvrouw, M.; De Béthune, M.P.; Burch, M.T.; Mower, H.F.; Davis, H.; Lasure, A.; Pauwels, R.; De Clercq, E.; Vlietinck, A.J. Antiviral activity of Hawaiian medicinal plants against human immunodeficiency virus type-1 (HIV-1). Phytomedicine 1996, 2, 259–264. [Google Scholar] [CrossRef]
- Geiger, J.M.; Ranker, T.A. Molecular phylogenetics and historical biogeography of Hawaiian Dryopteris (Dryopteridaceae). Mol. Phylogenet. Evol. 2005, 34, 392–407. [Google Scholar] [CrossRef] [PubMed]
- Palmer, D.D. Hawaii’s Ferns and Fern Allies; University of Hawaii Press: Honolulu, HI, USA, 2003. [Google Scholar]
- Widén, C.J.; Fraser-Jenkins, C.R.; Roux, J.P. A survey of phenolic compounds in Dryopteris and related fern genera. Part IV. Phloroglucinol derivatives and morphology in the section Marginatae (Pteridophyta, Dryopteridaceae). Finnish Zoological and Botanical Publishing Board. Ann. Bot. Fenn. 2015, 52, 53–83. [Google Scholar] [CrossRef]
- Bohm, B.A. Major exudate flavonoids of Dubautia arborea (Asteraceae). Biochem. Syst. Ecol. 1999, 27, 755–757. [Google Scholar] [CrossRef]
- Friar, E.A.; Cruse-Sanders, J.M.; McGlaughlin, M.E. Gene flow in Dubautia arborea and D. ciliolata: The roles of ecology and isolation by distance in maintaining species boundaries despite ongoing hybridization. Mol. Ecol. 2007, 16, 4028–4038. [Google Scholar] [CrossRef] [PubMed]
- Couch, C.S.; Simonis, J.S.; Thomas, R.Q.; Manuel, M.; Faucette, D.; Sparks, J.P. Erythrina sandwicensis Demography and Spatial Patterns of Erythrina Gall Wasp, Quadratichus erythrinae Infestation in Waikoloa, Hawai‘i; The Cornell-Hawai’i Graduate Field Research Laboratory, The Kohala Center: Waimea, HI, USA, 2009. [Google Scholar]
- Doccola, J.J.; Smith, S.L.; Strom, B.L.; Medeiros, A.C.; von Allmen, E. Systemically applied insecticides for treatment of Erythrina gall wasp, Quadrastichus erythrinae Kim (Hymenoptera: Eulophidae). Arboric. Urban For. 2009, 35, 173–181. [Google Scholar] [CrossRef]
- Folkers, K.; Koniuszy, F. Erythrina alkaloids. III. Isolation and characterization of a new alkaloid, erythramine. J. Am. Chem. Soc. 1939, 61, 1232–1235. [Google Scholar] [CrossRef]
- Folkers, K.; Koniuszy, F. Erythrina alkaloids. VI. Studies on the constitution of erythramine. J. Am. Chem. Soc. 1939, 61, 3053–3055. [Google Scholar] [CrossRef]
- Folkers, K.; Koniuszy, F. Erythrina alkaloids. IX. Isolation and characterization of erysodine, erysopine, erysocine and erysovine. J. Am. Chem. Soc. 1940, 62, 1677–1683. [Google Scholar] [CrossRef]
- Ingham, J.L. Induced isoflavonoids of Erythrina sandwicensis. Z. Naturforschung C 1980, 35, 384–386. [Google Scholar] [CrossRef]
- James, S.A.; Okamoto, F.; Tsuneyoshi, A. Genetic Variation in the Endemic Hawaiian Gardenia brighamii: Conservation and Horticultural Implications; Bishop Museum: Honolulu, HI, USA, 2007. [Google Scholar]
- John, H.S.; Richard, K.J. Revision of the native Hawaiian species of Gardenia (Rubiaceae). Hawaiian plant studies—15. Brittonia 1949, 6, 431–449. [Google Scholar] [CrossRef]
- Kafua, L.; Kritzinger, Q.; Hussein, A. Antifungal activity of Gardenia brighamii leaf extracts. S. Afr. J. Bot. 2010, 76, 411. [Google Scholar] [CrossRef]
- Morden, C.W.; Harbin, S.C. Evolution and biogeographic origins of the endemic Hawaiian Genus Hesperomannia (Asteraceae) 1. Pac. Sci. 2013, 67, 219–235. [Google Scholar] [CrossRef]
- Bohm, B.A.; Stuessy, T.F. Flavonoid chemistry of Barnadesioideae (Asteraceae). Syst. Bot. 1995, 1, 22–27. [Google Scholar] [CrossRef]
- Iwashina, T.; Saito, Y.; Kokubugata, G.; Peng, C.I. Flavonoids in the leaves of Hillebrandia and Begonia species (Begoniaceae). Biochem. Syst. Ecol. 2020, 90, 104040. [Google Scholar] [CrossRef]
- Goldberg, A.S. The Alkaloids of Lobelia yuccoides, Hillebr; University of Hawai’i at Manoa: Honolulu, HI, USA, 1966. [Google Scholar]
- Marr, K.L.; Bohm, B.A. A taxonomic revision of the endemic Hawaiian Lysimachia (Primulaceae) including three new species. Pac. Sci. 1997, 51, 254–287. [Google Scholar]
- Marr, K.L.; Bohm, B.A.; Cooke, C.; Gunning, P. Flavonoids of Hawaiian endemic Lysimachia in honour of Professor GH Neil Towers 75th birthday. Phytochemistry 1998, 49, 553–557. [Google Scholar] [CrossRef]
- Paetzold, C.; Wood, K.R.; Eaton, D.A.; Wagner, W.L.; Appelhans, M.S. Phylogeny of Hawaiian Melicope (Rutaceae): RAD-seq resolves species relationships and reveals ancient introgression. Front. Plant Sci. 2019, 10, 1074–1089. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.R.; Appelhans, M.S.; Wagner, W.L. Melicope stonei, section Pelea (Rutaceae), a new species from Kaua‘i, Hawaiian Islands: With notes on its distribution, ecology, conservation status, and phylogenetic placement. PhytoKeys 2017, 83, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Le, K.T.; Bandolik, J.J.; Kassack, M.U.; Wood, K.R.; Paetzold, C.; Appelhans, M.S.; Passreiter, C.M. New Acetophenones and chromenes from the leaves of Melicope barbigera A. Gray. Molecules 2021, 26, 688. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.L.; Lorence, D.H. A nomenclator of Pacific oceanic island Phyllanthus (Phyllanthaceae), including Glochidion. PhytoKeys 2011, 4, 67–94. [Google Scholar] [CrossRef]
- Verwijs, J.I.; Bouman, R.W.; van Welzen, P.C. A taxonomic revision of Phyllanthus subgenus Macraea (Phyllanthaceae). Blumea-Biodiversity. Evol. Biogeogr. Plants 2019, 64, 231–252. [Google Scholar] [CrossRef]
- Win, H.H.; Moe, T.S.; Hlaing, T.T.; San, M.M.; Win, Z.K.; Mya, K.M. Indigenous Myanmar medicinal plants and comparison of their in vitro antioxidant, antiglycation, and antimicrobial activities. Cogent Biol. 2019, 5, 1589634. [Google Scholar] [CrossRef]
- Thu, M.M.; Khaing, T.T.; Tin, T. Physico-chemical properties, antibacterial activities and antioxidant activities of prepared wine from Phyllanthus distichus Hook. & Arn. Technol. Univ. Myitkyina 1st Univ. Res. J. Sci. Eng. Eng. Educ. URJSEE 2019, 1, 151. [Google Scholar]
- Mohapatra, D.; Mishra, S. Hypoglycemic and antidiabetic evaluation of leaves of Phyllanthus distichus in normal and alloxan induced diabetic rats. Indian J. Pharm. Pharmacol. 2020, 7, 189–220. [Google Scholar] [CrossRef]
- Locher, C.P.; Burch, M.T.; Mower, H.F.; Berestecky, J.; Davis, H.; Van Poel, B.; Lasure, A.; Berghe, D.V.; Vlietinck, A.J. Anti-microbial activity and anti-complement activity of extracts obtained from selected Hawaiian medicinal plants. J. Ethnopharmacol. 1995, 49, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Kartika, H. Nutrients, Polyphenols, and Total Antioxidant Activities in Mamaki, Pipturus albidus. Ph.D. Dissertation, University of Hawaii at Manoa, Honolulu, HI, USA, 2006. [Google Scholar]
- Kartika, H.; Li, Q.X.; Wall, M.M.; Nakamoto, S.T.; Iwaoka, W.T. Major phenolic acids and total antioxidant activity in Mamaki leaves, Pipturus Albidus. J. Food Sci. 2007, 72, S696–S701. [Google Scholar] [CrossRef] [PubMed]
- Koher, G.; Khan, A.; Suarez-Vega, G.; Meesakul, P.; Bacani, A.J.; Kohno, T.; Zhu, X.; Kim, K.H.; Cao, S.; Jia, Z. A Comprehensive insight into Māmaki (Pipturus albidus): Its ethnomedicinal heritage, human health research, and phytochemical properties. Plants 2023, 12, 2924. [Google Scholar] [CrossRef] [PubMed]
- Meesakul, P.; Shea, T.; Wong, S.X.; Kuroki, Y.; Cao, S. Hawaiian plants with beneficial effects on sleep, anxiety, and mood, etc. Pharmaceuticals 2023, 16, 1228. [Google Scholar] [CrossRef]
- Ko, W.H.; Ko, S.S. A new twig blight of Mamaki (Pipturus albidus) caused by Botryosphaeria ribis. Can. J. Plant Pathol. 2001, 23, 312–314. [Google Scholar] [CrossRef]
- Kim, Y. Factors Influencing Antioxidant Phytochemical Stability of Teas; Texas A&M University: College Station, TX, USA, 2008. [Google Scholar]
- Kartika, H.; Shido, J.; Nakamoto, S.T.; Li, Q.X.; Iwaoka, W.T. Nutrient and mineral composition of dried Mamaki leaves (Pipturus albidus) and infusions. J. Food Compos. Anal. 2011, 24, 44–48. [Google Scholar] [CrossRef]
- Sun, A.; Kondratyuk, T.; Wongwiwatthananukit, S.; Sun, D.; Chang, L.C. investigation of antioxidant, anticancer, and chemopreventive properties of Hawaiian grown Māmaki tea (Pipturus albidus). Nat. Prod. Commun. 2022, 17, 1934578X221080945. [Google Scholar] [CrossRef]
- Appelhans, M.S.; Wood, K.R.; Wagner, W.L. Reduction of the Hawaiian genus Platydesma into Melicope section Pelea (Rutaceae) and notes on the monophyly of the section. PhytoKeys 2017, 91, 125–137. [Google Scholar] [CrossRef] [PubMed]
- Werny, F.; Scheuer, P. Hawaiian plant studies—IX: The alkaloids of Platydesma campanulata mann. Tetrahedron 1963, 19, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Sohmer, S.H. Psychotria L. (Rubiaceae) in the Hawaiian Islands; Lyonia Occasional Papers; Harold L. Lyon Arboretum: Honolulu, HI, USA, 1977; Volume 1, pp. 103–186. [Google Scholar]
- Abbott, L.L.; Pratt, L.W. Rare Plants of Naulu Forest and Poliokeawe Pali; Hawaii Volcanoes National Park: Hawaii County, HI, USA, 1996. [Google Scholar]
- Forest, N. Rare Plants of Naulu Forest and Poliokeawe Pali, Hawaii Volcanoes National Park; Cooperative National Park Resources Studies Unit University of Hawai’i at Manoa: Honolulu, HI, USA, 1996. [Google Scholar]
- Gorman, M.; Neuss, N.; Djerassi, C.; Kutney, J.P.; Scheuer, P.J. Alkaloid studies–XIX: Alkaloids of some Hawaiian Rauwolfia species: The structure of sandwicine and its interconversion with ajmaline and ajmalidine. Tetrahedron 1957, 1, 328–337. [Google Scholar] [CrossRef]
- Martin, N.J.; Prado, S.; Lecellier, G.; Thomas, O.P.; Raharivelomanana, P. Nukuhivensiums, indolo [2, 3-a] quinoliziniums from the Marquesan plant Rauvolfia nukuhivensis. Molecules 2012, 17, 12015–12022. [Google Scholar] [CrossRef] [PubMed]
- Merlin, M.D.; Thomson, L.A.; Elevitch, C.R. Santalum ellipticum, S. freycinetianum, S. haleakalae, and S. paniculatum (Hawaiian sandalwood). Traditional Trees of the Pacific Islands: Their Culture, Environment and Use; Permanent Agricultural Resources: Honolulu, HI, USA, 2006; pp. 695–714. [Google Scholar]
- Speetjens, T.M. Regeneration of Hemiparasitic Hawaiian Sandalwood (Santalum paniculatum Hook. & Arn.): The Role of Seedling Nutrition and Plant Hosts. Ph.D. Dissertation, Purdue University, West Lafayette, IN, USA, 2022. [Google Scholar]
- Braun, N.A.; Sim, S.; Kohlenberg, B.; Lawrence, B.M. Hawaiian sandalwood: Oil composition of Santalum paniculatum and comparison with other sandal species. Nat. Prod. Commun. 2014, 9, 1934578X1400900936. [Google Scholar] [CrossRef]
- Chavan, P.A. Evaluation of antimicrobial activity of various medicinal plants extracts of Latur zone against pathogens. Int. J. Life. Sci. Scienti. Res. 2016, 2, 612–618. [Google Scholar] [CrossRef]
- Ofori, H.; Hettiarachchi, D.; Sostaric, T.; Pandohee, J.; Chudasama, H.D.; Boyce, M.C. Characterisation of sandalwood essential oils: The application of high performance thin-layer chromatography. J. Essent. Oil Res. 2021, 33, 32–43. [Google Scholar] [CrossRef]
- Wagan, T.A.; Wang, W.; Hua, H.; Rong-Hua, L.; Cai, W. The effects of three essential oils on adult repellency, larval fumigant toxicity, and egg hatch of Tribolium castaneum (Coleoptera: Tenebrionidae). Fla. Entomol. 2022, 105, 160–166. [Google Scholar] [CrossRef]
- Kadooka, M.M.; Chang, M.Y.; Fukami, H.; Scheuer, P.J.; Clardy, J.; Solheim, B.A.; Springer, J.P. Mamanine and pohakuline, two unprecedented quinolizidine alkaloids from Sophora chrysophylla. Tetrahedron 1976, 32, 919–924. [Google Scholar] [CrossRef]
- Murakoshi, I.; Ito, M.; Haginiwa, J.; Ohmiya, S.; Otomasu, H.; Hirano, R.T. Lupin alkaloids from Sophora chrysophylla. Phytochemistry 1984, 23, 887–891. [Google Scholar] [CrossRef]
- Shirataki, Y.; Tsuzuku, T.; Yokoe, I.; Hirano, R.; Komatsu, M. Studies on the constituents of Sophora species. XXIII.: Constituents of the root of Sphora chrysophylla SEEM. (1). Chem. Pharm. Bull. 1990, 38, 1712–1716. [Google Scholar] [CrossRef]
- Bassil, N.V.; Bunch, T.; Nyberg, A.; Hummer, K.; Zee, F.T. Microsatellite markers distinguish Hawaiian ohelo from other Vaccinium L. section Myrtillus species. Int. Symp. Mol. Markers Hortic. 2009, 859, 81–88. [Google Scholar] [CrossRef]
- Wagner, W.L.; Herbst, D.R.; Sohmer, S.H. Manual of the Flowering Plants of Hawai’i. In Manual of the Flowering Plants of Hawai’i; University of Hawaii Press: Honolulu, HI, USA, 1990. [Google Scholar]
- Bohm, B.A.; Koupai-Abyazani, M.R. Flavonoids and condensed tannins from leaves of Hawaiian Vaccinium reticulatum and V. calycinum (Ericaceae). Pac. Sci. 1994, 48, 458–463. [Google Scholar]
- Hummer, K.; Durst, R.; Zee, F.; Atnip, A.; Giusti, M.M. Phytochemicals in fruits of Hawaiian wild cranberry relatives. J. Sci. Food Agric. 2014, 94, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Liu, X.; Nakamoto, S.T.; Wall, M.; Li, Y. Antimicrobial activity of Ohelo berry (Vaccinium calycinum) juice against Listeria monocytogenes and its potential for milk preservation. Microorganisms 2022, 10, 548. [Google Scholar] [CrossRef]
- Liu, X.; Wu, B.; Nakamoto, S.T.; Imamura, J.L.; Li, Y. Inhibition of Listeria monocytogenes and Escherichia coli O157: H7 Growth by Ohelo berry (Vaccinium calycinum) Fractions: Anthocyanins, non-anthocyanin phenolics, and organic acids. Microorganisms 2022, 10, 2231. [Google Scholar] [CrossRef]
- Medeiros, A.C. Flowering Plants and Gymnosperms of Haleakala National Park; University of Hawaii at Manoa: Honolulu, HI, USA, 1998. [Google Scholar]
- Jolad, S.D.; Hoffmann, J.J.; Timmermann, B.N.; Schram, K.H.; Cole, J.R.; Bates, R.B.; Klenck, R.E.; Tempesta, M.S. Daphnane diterpenes from Wikstroemia monticola: Wikstrotoxins A–D, huratoxin, and excoecariatoxin. J. Nat. Prod. 1983, 46, 675–680. [Google Scholar] [CrossRef]
- Torrance, S.J.; Hoffmann, J.J.; Cole, J.R. Wikstromol, antitumor lignan from Wikstroemia foetida var. oahuensis Gray and Wikstroemia uva-ursi Gray (Thymelaeaceae). J. Pharm. Sci. 1979, 68, 664–665. [Google Scholar] [CrossRef]
- Wang, H.B.; Liu, L.P.; Wang, X.Y. 13C-NMR data of daphnane diterpenoids. Magn. Reson. Chem. 2013, 51, 580–592. [Google Scholar] [CrossRef]
- Bornhorst, H.L.; Rauch, F.D. Native Hawaiian Plants for Landscaping, Conservation, and Reforestation. Ornamentals and Flowers; University of Hawaii: Honolulu, HI, USA, 2003. [Google Scholar]
- Tanuwidjaja, C.; Webb, D.T.; Sagawa, Y. Micropropagation of ‘Ãkia (Wikstroemia uva-ursi A. Gray). Plant Cell Tissue Organ Cult. 1998, 53, 85–90. [Google Scholar] [CrossRef]
- Gupta, S.; Gillett, G.W. Observations on Hawaiian species of Wikstroemia (Angiospermae: Thymelaeaceae). Pac. Sci. 1969, 23, 83–88. [Google Scholar]
- Rauch, F.D.; David, H. Akia. Ornamentals and Flowers; University of Hawaii: Honolulu, HI, USA, 1997. [Google Scholar]
- Gupta, S.; Gillet, G.W. Chemotaxonomic studies of Hawaiian Wikstroemia. Econ. Bot. 1969, 23, 24–31. [Google Scholar] [CrossRef]
- Lee, K.H.; Tagahara, K.; Suzuki, H.; Wu, R.Y.; Haruna, M.; Hall, I.H.; Huang, H.C.; Ito, K.; Iida, T.; Lai, J.S. Antitumor agents. 49. Tricin, kaempferol-3-0-β-D-Glucopyranoside and (+)-nortrachelogenin, antileukemic principles from Wikstroemia indica. J. Nat. Prod. 1981, 44, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Robichaux, R.H.; Carr, G.D.; Liebman, M.; Pearcy, R.W. Adaptive radiation of the Hawaiian silversword alliance (Compositae-Madiinae): Ecological, morphological, and physiological diversity. Ann. Mo. Bot. Gard. 1990, 1, 64–72. [Google Scholar] [CrossRef]
- Rundel, P.W.; Smith, A.P.; Meinzer, F.C. Tropical Alpine Environments: Plant Form and Function; Cambridge University Press: Cambridge, UK, 1994. [Google Scholar]
- Koske, R.E.; Gemma, J.N. Mycorrhizal status of two Hawaiian plant species (Asteraceae) in a tropical alpine habitat: The threatened Haleakala silversword (Argyroxiphium sandwicense subsp. macrocephalum) and the endemic Dubautia menziesii. Pac. Sci. 2002, 56, 423–430. [Google Scholar] [CrossRef]
- Carlquist, S. Wood anatomy of Wilkesia (Asteraceae) with relation to systematics, organography, and habit. J. Torrey Bot. Soc. 1998, 1, 261–267. [Google Scholar] [CrossRef]
- Bohm, B.A.; Fong, C. Nonpolar flavonoids of Wilkesia and Argyroxiphium. Phytochemistry 1990, 29, 1175–1177. [Google Scholar] [CrossRef]
- Hillebrand, W. Flora of the Hawaiian Islands: A Description of Their Phanerogams and Vascular Cryptogams; Williams & Norgate: London, UK, 1888; p. 76. [Google Scholar]
- Fish, F.; Gray, A.I.; Waterman, P.G. Alkaloids, coumarins, triterpenes and a flavanone from the root of Zanthoxylum dipetalum. Phytochemistry 1975, 14, 2073–2076. [Google Scholar] [CrossRef]
- Fish, F.; Gray, A.I.; Waigh, R.D.; Waterman, P.G. Detalolactone: A novel pyranocoumarin from the root bark of Zanthoxylum dipetalum. Phytochemistry 1976, 15, 313–316. [Google Scholar] [CrossRef]
- Arslanian, R.L.; Mondragon, B.; Stermitz, F.R.; Marr, K.L. Acyl histamines and a rare protopine-type alkaloid from leaves of Zanthoxylum dipetalum. Biochem. Syst. Ecol. 1990, 18, 345–347. [Google Scholar] [CrossRef]
- Shaw, R.B. Colorado State University Fort Collins. Influence of Wildland Fire on the Recovery of Endangered Plant Species Study Project; Colorado State University: Fort Collins, CO, USA, 1995. [Google Scholar]
- Marr, K.L.; Tang, C.S. Volatile insecticidal compounds and chemical variability of Hawaiian Zanthoxylum (Rutaceae) species. Biochem. Syst. Ecol. 1992, 20, 209–217. [Google Scholar] [CrossRef]
- Marr, K.L. Insecticidal Activity of the Endemic Hawaiian Species of the Genus Zanthoxylum L. (Rutaceae); University of Hawai’i at Manoa: Honolulu, HI, USA, 1989. [Google Scholar]





















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meesakul, P.; Shea, T.; Fenstemacher, R.; Wong, S.X.; Kuroki, Y.; Wada, A.; Cao, S. Phytochemistry and Biological Studies of Endemic Hawaiian Plants. Int. J. Mol. Sci. 2023, 24, 16323. https://doi.org/10.3390/ijms242216323
Meesakul P, Shea T, Fenstemacher R, Wong SX, Kuroki Y, Wada A, Cao S. Phytochemistry and Biological Studies of Endemic Hawaiian Plants. International Journal of Molecular Sciences. 2023; 24(22):16323. https://doi.org/10.3390/ijms242216323
Chicago/Turabian StyleMeesakul, Pornphimon, Tyler Shea, Roland Fenstemacher, Shi Xuan Wong, Yutaka Kuroki, Aya Wada, and Shugeng Cao. 2023. "Phytochemistry and Biological Studies of Endemic Hawaiian Plants" International Journal of Molecular Sciences 24, no. 22: 16323. https://doi.org/10.3390/ijms242216323