

Expression of Cannabinoid Receptors in the Trigeminal Ganglion of the Horse

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
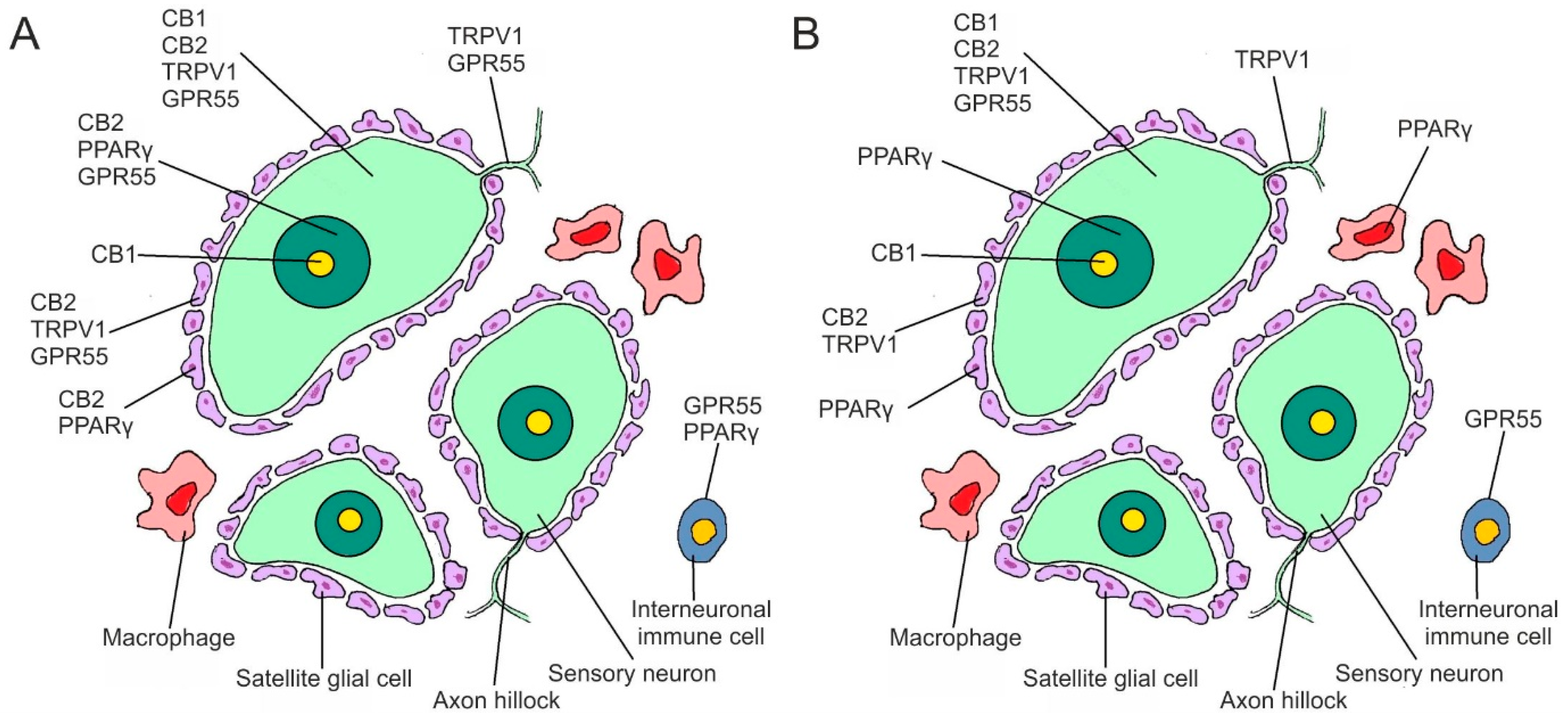
2. Results
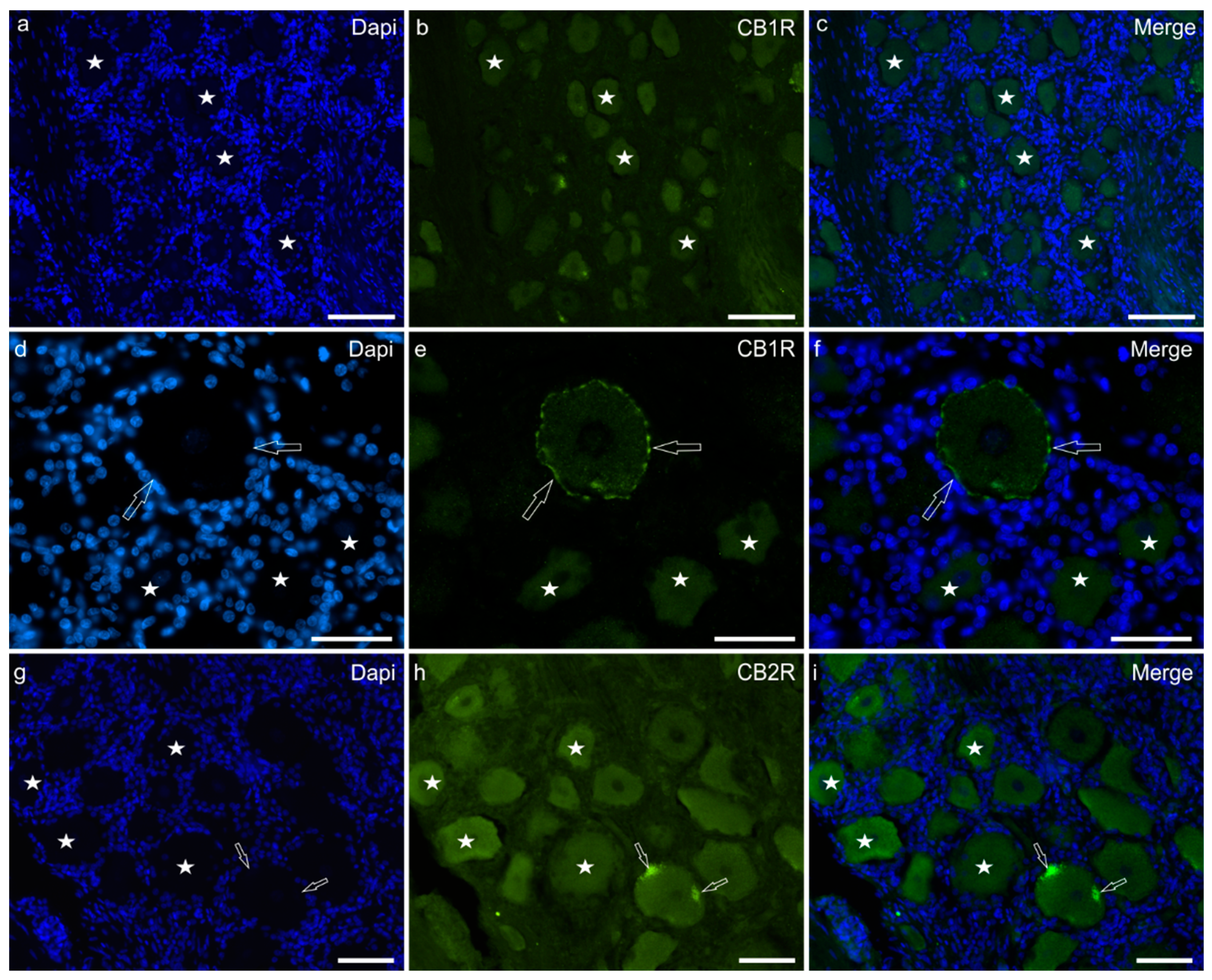
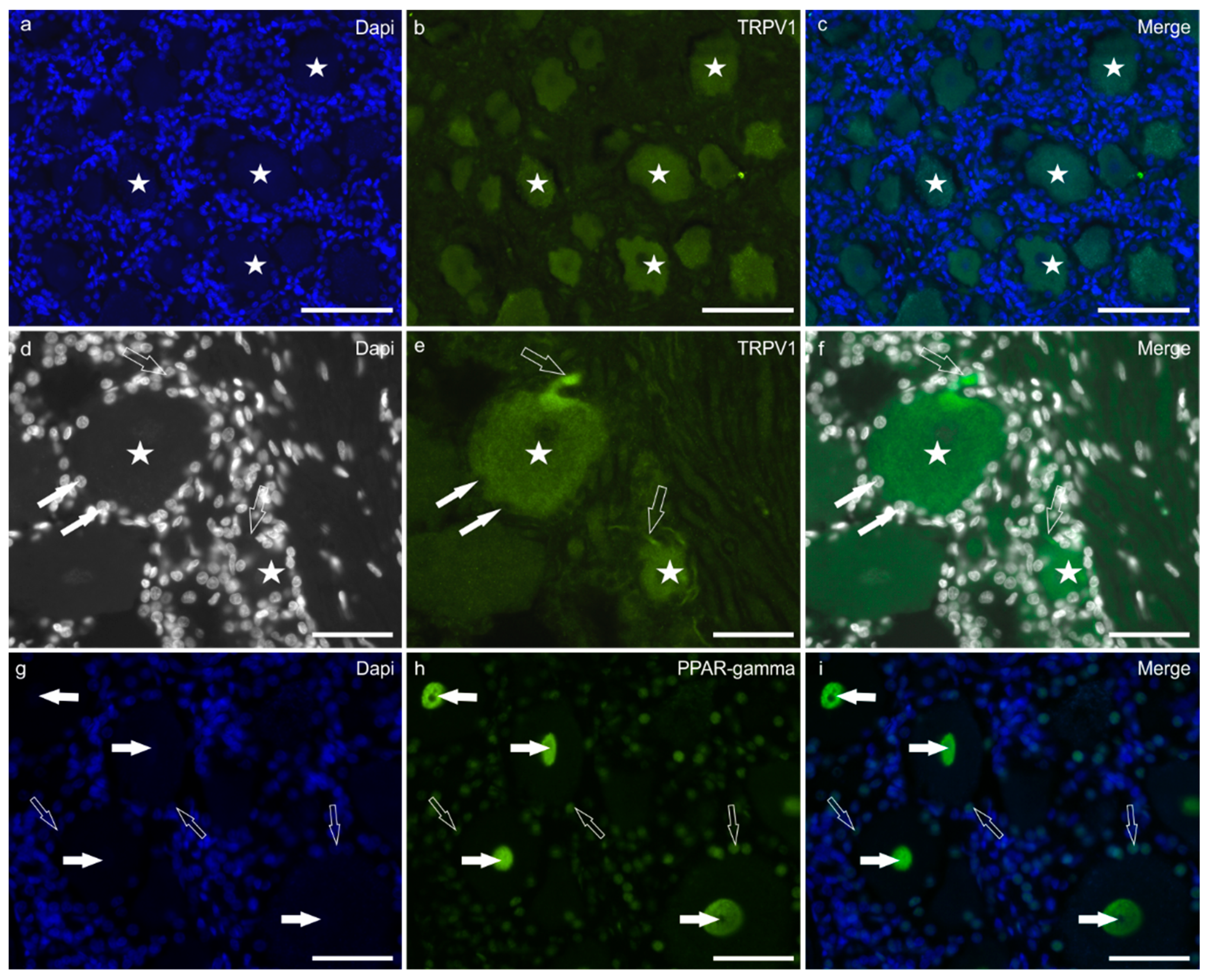
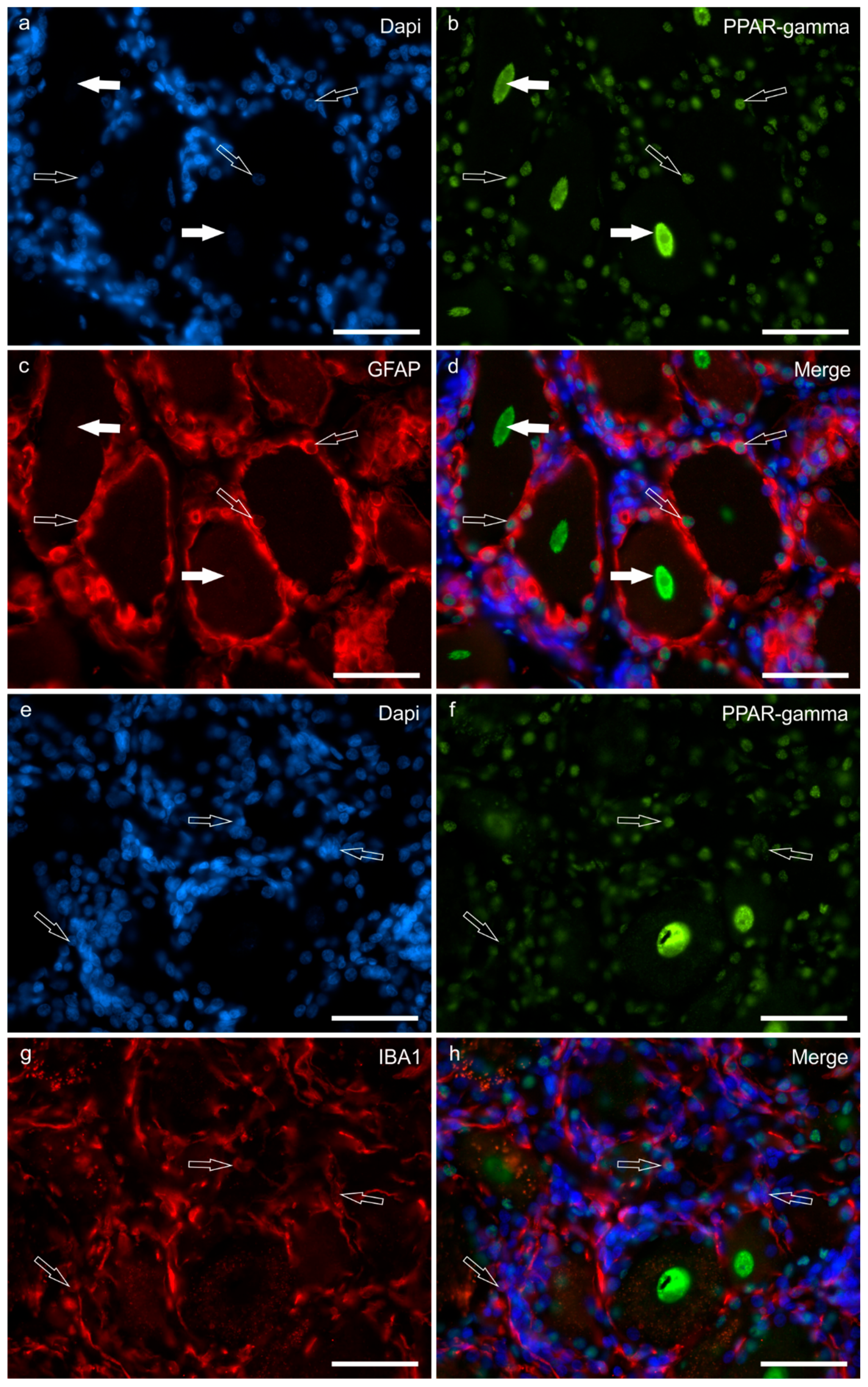
2.1. Immunoreactivity in Neurons—Cryosections
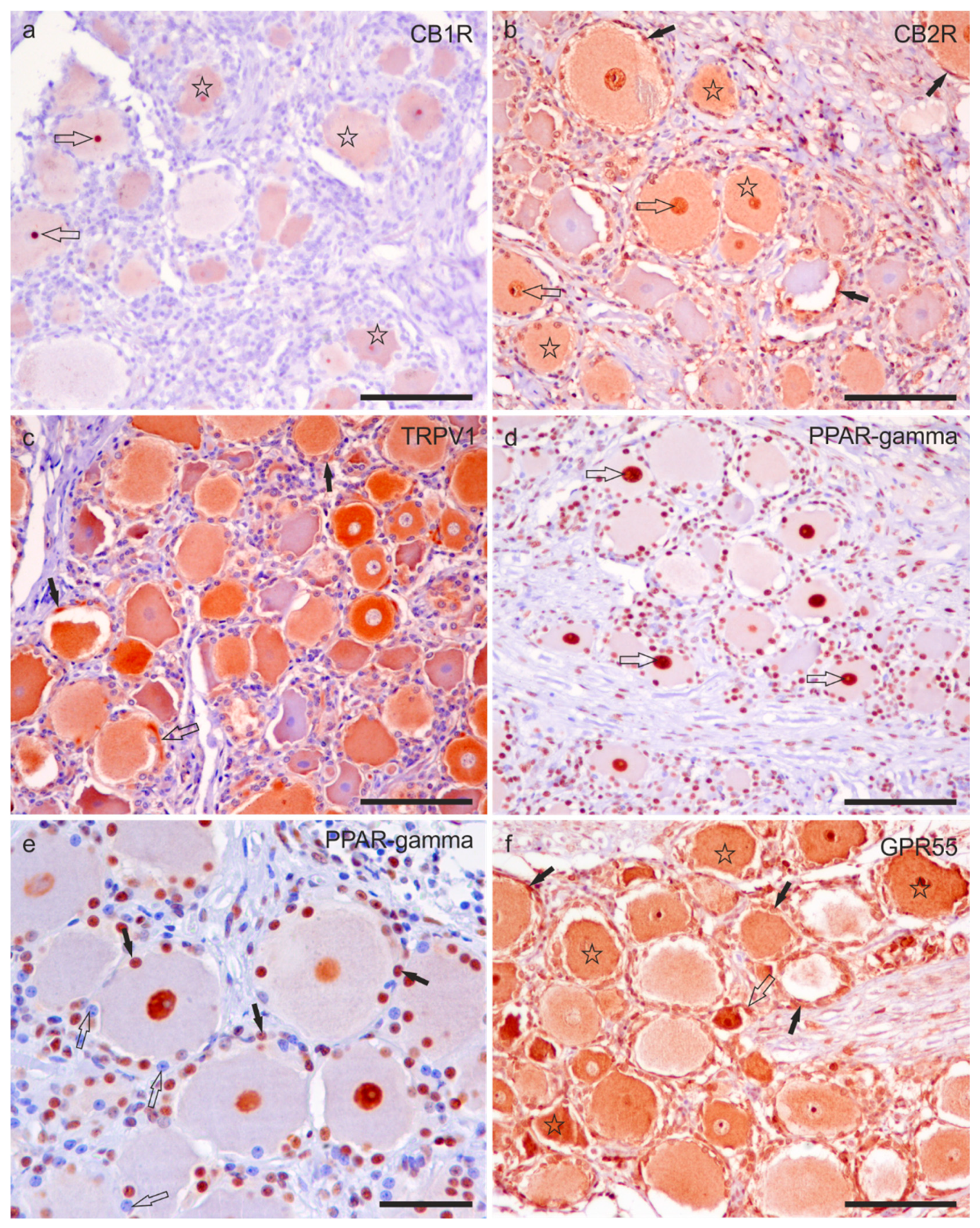
2.2. Immunoreactivity in Neurons—FFPE Sections
3. Discussion
Immunoreactivity in Trigeminal Neurons
4. Materials and Methods
4.1. Animals
4.2. Immunofluorescence on Cryosections
4.3. Immunohistochemistry on FFPE
4.4. Quantitative Analysis of the Immunoreactivity
4.5. Specificity of the Primary Antibodies
4.6. Specificity of the Secondary Antibody
4.7. Fluorescence Microscopy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ross, S.E.; Murray, J.K.; Roberts, V.L.H. Prevalence of Headshaking within the Equine Population in the UK. Equine Vet. J. 2018, 50, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Roberts, V.L.; Fews, D.; McNamara, J.M.; Love, S. Trigeminal Nerve Root Demyelination Not Seen in Six Horses Diagnosed with Trigeminal-Mediated Headshaking. Front. Vet. Sci. 2017, 4, 72. [Google Scholar] [CrossRef]
- Aleman, M.; Rhodes, D.; Williams, D.C.; Guedes, A.; Madigan, J.E. Sensory Evoked Potentials of the Trigeminal Nerve for the Diagnosis of Idiopathic Headshaking in a Horse. J. Vet. Int. Med. 2014, 28, 250–253. [Google Scholar] [CrossRef]
- Elliott, M.B.; Ward, S.J.; Abood, M.E.; Tuma, R.F.; Jallo, J.I. Understanding the Endocannabinoid System as a Modulator of the Trigeminal Pain Response to Concussion. Concussion 2017, 2, CNC49. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular Targets of the Phytocannabinoids: A Complex Picture. In Phytocannabinoids; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J., Eds.; Progress in the Chemistry of Organic Natural Products; Springer International Publishing: Cham, Switzerland, 2017; Volume 103, pp. 103–131. ISBN 978-3-319-45539-6. [Google Scholar]
- Ligresti, A.; De Petrocellis, L.; Di Marzo, V. From Phytocannabinoids to Cannabinoid Receptors and Endocannabinoids: Pleiotropic Physiological and Pathological Roles Through Complex Pharmacology. Physiol. Rev. 2016, 96, 1593–1659. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Huang, C.; Hsu, K. Therapeutic Potential of Cannabinoids in Trigeminal Neuralgia. Curr. Drug Targets CNS Neurol. Disord. 2004, 3, 507–514. [Google Scholar] [CrossRef]
- Lee, G.; Grovey, B.; Furnish, T.; Wallace, M. Medical Cannabis for Neuropathic Pain. Curr. Pain. Headache Rep. 2018, 22, 8. [Google Scholar] [CrossRef]
- Ellis, K.L.; Contino, E.K. Treatment Using Cannabidiol in a Horse with Mechanical Allodynia. Equine Vet. Educ. 2021, 33, e79–e82. [Google Scholar] [CrossRef]
- Sánchez-Aparicio, P.; Florán, B.; Rodríguez Velázquez, D.; Ibancovichi, J.A.; Varela Guerrero, J.A.; Recillas, S. Cannabinoids CB2 Receptors, One New Promising Drug Target for Chronic and Degenerative Pain Conditions in Equine Veterinary Patients. J. Equine Vet. Sci. 2020, 85, 102880. [Google Scholar] [CrossRef]
- Luedke, C.; Wilhelm, T. Cannabinoids in Equine Medicine. In Cannabis Therapy in Veterinary Medicine: A Complete Guide; Cital, S., Kramer, K., Hughston, L., Gaynor, J.S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 295–305. ISBN 978-3-030-68317-7. [Google Scholar]
- Zamith Cunha, R.; Zannoni, A.; Salamanca, G.; De Silva, M.; Rinnovati, R.; Gramenzi, A.; Forni, M.; Chiocchetti, R. Expression of Cannabinoid (CB1 and CB2) and Cannabinoid-Related Receptors (TRPV1, GPR55, and PPARα) in the Synovial Membrane of the Horse Metacarpophalangeal Joint. Front. Vet. Sci. 2023, 10, 1045030. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Rinnovati, R.; Tagliavia, C.; Stanzani, A.; Galiazzo, G.; Giancola, F.; Silva, M.D.; Capodanno, Y.; Spadari, A. Localisation of Cannabinoid and Cannabinoid-Related Receptors in the Equine Dorsal Root Ganglia. Equine Vet. J. 2021, 53, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Galiazzo, G.; Tagliavia, C.; Giancola, F.; Rinnovati, R.; Sadeghinezhad, J.; Bombardi, C.; Grandis, A.; Pietra, M.; Chiocchetti, R. Localisation of Cannabinoid and Cannabinoid-Related Receptors in the Horse Ileum. J. Equine Vet. Sci. 2021, 104, 103688. [Google Scholar] [CrossRef] [PubMed]
- Tajti, J.; Szok, D.; Csáti, A.; Szabó, Á.; Tanaka, M.; Vécsei, L. Exploring Novel Therapeutic Targets in the Common Pathogenic Factors in Migraine and Neuropathic Pain. Int. J. Mol. Sci. 2023, 24, 4114. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Demartini, C.; Zanaboni, A.M.; Francavilla, M.; De Icco, R.; Ahmad, L.; Tassorelli, C. The Endocannabinoid System and Related Lipids as Potential Targets for the Treatment of Migraine-related Pain. Headache 2022, 62, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Richardson, D.; Pearson, R.G.; Kurian, N.; Latif, M.L.; Garle, M.J.; Barrett, D.A.; Kendall, D.A.; Scammell, B.E.; Reeve, A.J.; Chapman, V. Characterisation of the Cannabinoid Receptor System in Synovial Tissue and Fluid in Patients with Osteoarthritis and Rheumatoid Arthritis. Arthritis Res. Ther. 2008, 10, R43. [Google Scholar] [CrossRef]
- O’Brien, M.; McDougall, J.J. Cannabis and Joints: Scientific Evidence for the Alleviation of Osteoarthritis Pain by Cannabinoids. Curr. Opin. Pharmacol. 2018, 40, 104–109. [Google Scholar] [CrossRef]
- Lowe, H.; Toyang, N.; Steele, B.; Bryant, J.; Ngwa, W. The Endocannabinoid System: A Potential Target for the Treatment of Various Diseases. Int. J. Mol. Sci. 2021, 22, 9472. [Google Scholar] [CrossRef]
- Fiani, B.; Sarhadi, K.J.; Soula, M.; Zafar, A.; Quadri, S.A. Current Application of Cannabidiol (CBD) in the Management and Treatment of Neurological Disorders. Neurol. Sci. 2020, 41, 3085–3098. [Google Scholar] [CrossRef]
- Longworth, J.; Szafron, M.; Gruza, A.; Da Silva, K. Cannabis and Cannabinoid Medications for the Treatment of Chronic Orofacial Pain: A Scoping Review. Dent. Rev. 2023, 3, 100063. [Google Scholar] [CrossRef]
- Price, T.J.; Helesic, G.; Parghi, D.; Hargreaves, K.M.; Flores, C.M. The Neuronal Distribution of Cannabinoid Receptor Type 1 in the Trigeminal Ganglion of the Rat. Neuroscience 2003, 120, 155–162. [Google Scholar] [CrossRef]
- Christiansen, I.M.; Edvinsson, J.C.A.; Reducha, P.V.; Edvinsson, L.; Haanes, K.A. Dual Action of the Cannabinoid Receptor 1 Ligand Arachidonyl-2′-Chloroethylamide on Calcitonin Gene-Related Peptide Release. J. Headache Pain. 2022, 23, 30. [Google Scholar] [CrossRef] [PubMed]
- Aleman, M.; Williams, D.C.; Brosnan, R.J.; Nieto, J.E.; Pickles, K.J.; Berger, J.; Lecouteur, R.A.; Holliday, T.A.; Madigan, J.E. Sensory Nerve Conduction and Somatosensory Evoked Potentials of the Trigeminal Nerve in Horses with Idiopathic Headshaking. J. Vet. Intern. Med. 2013, 27, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Greco, R.; Gasperi, V.; Maccarrone, M.; Tassorelli, C. The Endocannabinoid System and Migraine. Exp. Neurol. 2010, 224, 85–91. [Google Scholar] [CrossRef]
- Russo, D.; Bombardi, C.; Castellani, G.; Chiocchetti, R. Characterization of Spinal Ganglion Neurons in Horse (Equus caballus). A Morphometric, Neurochemical and Tracing Study. Neuroscience 2011, 176, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Ahluwalia, J.; Urban, L.; Capogna, M.; Bevan, S.; Nagy, I. Cannabinoid 1 Receptors Are Expressed in Nociceptive Primary Sensory Neurons. Neuroscience 2000, 100, 685–688. [Google Scholar] [CrossRef]
- Bridges, D.; Rice, A.S.C.; Egertová, M.; Elphick, M.R.; Winter, J.; Michael, G.J. Localisation of Cannabinoid Receptor 1 in Rat Dorsal Root Ganglion Using in Situ Hybridisation and Immunohistochemistry. Neuroscience 2003, 119, 803–812. [Google Scholar] [CrossRef]
- Agarwal, N.; Pacher, P.; Tegeder, I.; Amaya, F.; Constantin, C.E.; Brenner, G.J.; Rubino, T.; Michalski, C.W.; Marsicano, G.; Monory, K.; et al. Cannabinoids Mediate Analgesia Largely via Peripheral Type 1 Cannabinoid Receptors in Nociceptors. Nat. Neuro Sci. 2007, 10, 870–879. [Google Scholar] [CrossRef]
- Chiocchetti, R.; Galiazzo, G.; Tagliavia, C.; Stanzani, A.; Giancola, F.; Menchetti, M.; Militerno, G.; Bernardini, C.; Forni, M.; Mandrioli, L. Cellular Distribution of Canonical and Putative Cannabinoid Receptors in Canine Cervical Dorsal Root Ganglia. Front. Vet. Sci. 2019, 6, 313. [Google Scholar] [CrossRef]
- Khasabova, I.A.; Simone, D.A.; Seybold, V.S. Cannabinoids Attenuate Depolarization-Dependent Ca2+ Influx in Intermediate-Size Primary Afferent Neurons of Adult Rats. Neuroscience 2002, 115, 613–625. [Google Scholar] [CrossRef]
- Lalonde, M.R.; Jollimore, C.A.B.; Stevens, K.; Barnes, S.; Kelly, M.E.M. Cannabinoid Receptor-Mediated Inhibition of Calcium Signaling in Rat Retinal Ganglion Cells. Mol. Vis. 2006, 12, 1160–1166. [Google Scholar]
- Zhang, Y.; Xie, H.; Lei, G.; Li, F.; Pan, J.; Liu, C.; Liu, Z.; Liu, L.; Cao, X. Regulatory Effects of Anandamide on Intracellular Ca2+ Concentration Increase in Trigeminal Ganglion Neurons. Neural Regen. Res. 2014, 9, 878. [Google Scholar] [CrossRef]
- Price, T.J.; Patwardhan, A.; Akopian, A.N.; Hargreaves, K.M.; Flores, C.M. Modulation of Trigeminal Sensory Neuron Activity by the Dual Cannabinoid-Vanilloid Agonists Anandamide, N -Arachidonoyl-Dopamine and Arachidonyl-2-Chloroethylamide: Cannabinoid-Vanilloid Modulation of Trigeminal Neuron Activity. Br. J. Pharmacol. 2004, 141, 1118–1130. [Google Scholar] [CrossRef]
- Akerman, S.; Holland, P.R.; Goadsby, P.J. Cannabinoid (CB1) Receptor Activation Inhibits Trigeminovascular Neurons. J. Pharmacol. Exp. Ther. 2007, 320, 64–71. [Google Scholar] [CrossRef]
- Atwood, B.K.; Mackie, K. CB2: A Cannabinoid Receptor with an Identity Crisis. Br. J. Pharmacol. 2010, 160, 467–479. [Google Scholar] [CrossRef]
- Anand, U.; Otto, W.R.; Sanchez-Herrera, D.; Facer, P.; Yiangou, Y.; Korchev, Y.; Birch, R.; Benham, C.; Bountra, C.; Chessell, I.P.; et al. Cannabinoid Receptor CB2 Localisation and Agonist-Mediated Inhibition of Capsaicin Responses in Human Sensory Neurons. Pain 2008, 138, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, T.; Crystal, J.D.; Zvonok, A.M.; Makriyannis, A.; Hohmann, A.G. Self-Medication of a Cannabinoid CB2 Agonist in an Animal Model of Neuropathic Pain. Pain 2011, 152, 1976–1987. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N. Cannabinoid Receptor with an “identity Crisis” Gets a Second Look. Nat. Med. 2015, 21, 966–967. [Google Scholar] [CrossRef]
- Svízenská, I.H.; Brázda, V.; Klusáková, I.; Dubový, P. Bilateral Changes of Cannabinoid Receptor Type 2 Protein and mRNA in the Dorsal Root Ganglia of a Rat Neuropathic Pain Model. J. Histochem. Cytochem. 2013, 61, 529–547. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The Cloned Capsaicin Receptor Integrates Multiple Pain-Producing Stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Russo, D.; Clavenzani, P.; Sorteni, C.; Bo Minelli, L.; Botti, M.; Gazza, F.; Panu, R.; Ragionieri, L.; Chiocchetti, R. Neurochemical Features of Boar Lumbosacral Dorsal Root Ganglion Neurons and Characterization of Sensory Neurons Innervating the Urinary Bladder Trigone. J. Comp. Neurol. 2013, 521, 342–366. [Google Scholar] [CrossRef]
- Tanimoto, T.; Takeda, M.; Nasu, M.; Kadoi, J.; Matsumoto, S. Immunohistochemical Co-Expression of Carbonic Anhydrase II with Kv1.4 and TRPV1 in Rat Small-Diameter Trigeminal Ganglion Neurons. Brain Res. 2005, 1044, 262–265. [Google Scholar] [CrossRef]
- Ichikawa, H.; Sugimoto, T. VR1-Immunoreactive Primary Sensory Neurons in the Rat Trigeminal Ganglion. Brain Res. 2001, 890, 184–188. [Google Scholar] [CrossRef]
- Patapoutian, A.; Peier, A.M.; Story, G.M.; Viswanath, V. ThermoTRP Channels and beyond: Mechanisms of Temperature Sensation. Nat. Rev. Neurosci. 2003, 4, 529–539. [Google Scholar] [CrossRef] [PubMed]
- Culshaw, A.J.; Bevan, S.; Christiansen, M.; Copp, P.; Davis, A.; Davis, C.; Dyson, A.; Dziadulewicz, E.K.; Edwards, L.; Eggelte, H.; et al. Identification and Biological Characterization of 6-Aryl-7-Isopropylquinazolinones as Novel TRPV1 Antagonists That Are Effective in Models of Chronic Pain. J. Med. Chem. 2006, 49, 471–474. [Google Scholar] [CrossRef] [PubMed]
- Krause, J.E.; Chenard, B.L.; Cortright, D.N. Transient Receptor Potential Ion Channels as Targets for the Discovery of Pain Therapeutics. Curr. Opin. Investig. Drugs 2005, 6, 48–57. [Google Scholar] [PubMed]
- Marwaha, L.; Bansal, Y.; Singh, R.; Saroj, P.; Sodhi, R.K.; Kuhad, A. Niflumic Acid, a TRPV1 Channel Modulator, Ameliorates Stavudine-Induced Neuropathic Pain. Inflammopharmacol 2016, 24, 319–334. [Google Scholar] [CrossRef]
- Sagar, D.R.; Gaw, A.G.; Okine, B.N.; Woodhams, S.G.; Wong, A.; Kendall, D.A.; Chapman, V. Dynamic Regulation of the Endocannabinoid System: Implications for Analgesia. Mol. Pain. 2009, 5, 59. [Google Scholar] [CrossRef] [PubMed]
- Di Marzo, V.; Piscitelli, F. The Endocannabinoid System and Its Modulation by Phytocannabinoids. Neurotherapeutics 2015, 12, 692–698. [Google Scholar] [CrossRef]
- Mlost, J.; Bryk, M.; Starowicz, K. Cannabidiol for Pain Treatment: Focus on Pharmacology and Mechanism of Action. Int. J. Mol. Sci. 2020, 21, 8870. [Google Scholar] [CrossRef] [PubMed]
- Barak, Y.; Nelson, M.C.; Ong, E.S.; Jones, Y.Z.; Ruiz-Lozano, P.; Chien, K.R.; Koder, A.; Evans, R.M. PPARγ Is Required for Placental, Cardiac, and Adipose Tissue Development. Mol. Cell 1999, 4, 585–595. [Google Scholar] [CrossRef]
- Tontonoz, P.; Spiegelman, B.M. Fat and Beyond: The Diverse Biology of PPARγ. Annu. Rev. Biochem. 2008, 77, 289–312. [Google Scholar] [CrossRef]
- Zhao, X.; Ou, Z.; Grotta, J.C.; Waxham, N.; Aronowski, J. Peroxisome-Proliferator-Activated Receptor-Gamma (PPARγ) Activation Protects Neurons from NMDA Excitotoxicity. Brain Res. 2006, 1073–1074, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Quintanilla, R.A.; Utreras, E.; Cabezas-Opazo, F.A. Role of PPARγ in the Differentiation and Function of Neurons. PPAR Res. 2014, 2014, e768594. [Google Scholar] [CrossRef] [PubMed]
- Maeda, T.; Kishioka, S. Chapter 13 PPAR and Pain. In International Review of Neurobiology; Academic Press: Cambridge, MA, USA, 2009; Volume 85, pp. 165–177. [Google Scholar]
- Lyons, D.N.; Zhang, L.; Danaher, R.J.; Miller, C.S.; Westlund, K.N. PPARγ agonists attenuate trigeminal neuropathic pain. Clin. J. Pain. 2017, 33, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Churi, S.B.; Abdel-Aleem, O.S.; Tumber, K.K.; Scuderi-Porter, H.; Taylor, B.K. Intrathecal Rosiglitazone Acts at Peroxisome Proliferator-Activated Receptor-Gamma to Rapidly Inhibit Neuropathic Pain in Rats. J. Pain. 2008, 9, 639–649. [Google Scholar] [CrossRef]
- Vučković, S.; Srebro, D.; Vujović, K.S.; Vučetić, Č.; Prostran, M. Cannabinoids and Pain: New Insights From Old Molecules. Front. Pharmacol. 2018, 9, 1259. [Google Scholar] [CrossRef]
- Aviram, J.; Samuelly-Leichtag, G. Efficacy of Cannabis-Based Medicines for Pain Management: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Pain Physician 2017, 20, E755–E796. [Google Scholar] [CrossRef]
- Hill, K.P. Medical Marijuana for Treatment of Chronic Pain and Other Medical and Psychiatric Problems: A Clinical Review. JAMA 2015, 313, 2474. [Google Scholar] [CrossRef]
- Chu, Y.; Jia, S.; Xu, K.; Liu, Q.; Mai, L.; Liu, J.; Fan, W.; Huang, F. Single-Cell Transcriptomic Profile of Satellite Glial Cells in Trigeminal Ganglion. Front. Mol. Neurosci. 2023, 16, 1117065. [Google Scholar] [CrossRef]
- Takahashi, Y.; Hasegawa-Moriyama, M.; Sakurai, T.; Inada, E. The Macrophage-Mediated Effects of the Peroxisome Proliferator-Activated Receptor-Gamma Agonist Rosiglitazone Attenuate Tactile Allodynia in the Early Phase of Neuropathic Pain Development. Anesth. Analg. 2011, 113, 398–404. [Google Scholar] [CrossRef]
- Hasegawa-Moriyama, M.; Kurimoto, T.; Nakama, M.; Godai, K.; Kojima, M.; Kuwaki, T.; Kanmura, Y. Peroxisome Proliferator-Activated Receptor-Gamma Agonist Rosiglitazone Attenuates Inflammatory Pain through the Induction of Heme Oxygenase-1 in Macrophages. Pain 2013, 154, 1402–1412. [Google Scholar] [CrossRef] [PubMed]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.-O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The Orphan Receptor GPR55 Is a Novel Cannabinoid Receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar] [CrossRef] [PubMed]
- Console-Bram, L.; Marcu, J.; Abood, M.E. Cannabinoid Receptors: Nomenclature and Pharmacological Principles. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 38, 4–15. [Google Scholar] [CrossRef]
- Chiocchetti, R.; De Silva, M.; Aspidi, F.; Cunha, R.Z.; Gobbo, F.; Tagliavia, C.; Sarli, G.; Morini, M. Distribution of Cannabinoid Receptors in Keratinocytes of Healthy Dogs and Dogs with Atopic Dermatitis. Front. Vet. Sci. 2022, 9, 915896. [Google Scholar] [CrossRef]
- Ono, T.; Yamashita, T.; Kano, R.; Inoue, M.; Okada, S.; Kano, K.; Koizumi, S.; Iwabuchi, K.; Hirabayashi, Y.; Matsuo, I.; et al. GPR55 Contributes to Neutrophil Recruitment and Mechanical Pain Induction after Spinal Cord Compression in Mice. Brain Behav. Immun. 2023, 110, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Carey, L.M.; Gutierrez, T.; Deng, L.; Lee, W.-H.; Mackie, K.; Hohmann, A.G. Inflammatory and Neuropathic Nociception Is Preserved in GPR55 Knockout Mice. Sci. Rep. 2017, 7, 944. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Du, J.; Li, L.; Xu, X.; Liang, S. Central Role of Purinergic Receptors with Inflammation in Neuropathic Pain-Related Macrophage-SGC-Neuron Triad. Neuropharmacology 2023, 228, 109445. [Google Scholar] [CrossRef]
- Lu, X.; Richardson, P.M. Inflammation near the Nerve Cell Body Enhances Axonal Regeneration. J. Neurosci. 1991, 11, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.-Y.; Sun, Y.-N.; Wang, F.-T.; Li, Q.; Su, L.; Zhao, Z.-F.; Meng, X.-L.; Zhao, H.; Wu, X.; Sun, Q.; et al. Activation of Satellite Glial Cells in Lumbar Dorsal Root Ganglia Contributes to Neuropathic Pain after Spinal Nerve Ligation. Brain Res. 2012, 1427, 65–77. [Google Scholar] [CrossRef]
- Dubový, P.; Jančálek, R.; Klusáková, I.; Svíženská, I.; Pejchalová, K. Intra- and Extraneuronal Changes of Immunofluorescence Staining for TNF- and TNFR1 in the Dorsal Root Ganglia of Rat Peripheral Neuropathic Pain Models. Cell Mol. Neurobiol. 2006, 26, 1203–1215. [Google Scholar] [CrossRef]
- Takeda, M.; Tanimoto, T.; Kadoi, J.; Nasu, M.; Takahashi, M.; Kitagawa, J.; Matsumoto, S. Enhanced Excitability of Nociceptive Trigeminal Ganglion Neurons by Satellite Glial Cytokine Following Peripheral Inflammation. Pain 2007, 129, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Dubový, P.; Klusáková, I.; Svíženská, I.; Brázda, V. Satellite Glial Cells Express IL-6 and Corresponding Signal-Transducing Receptors in the Dorsal Root Ganglia of Rat Neuropathic Pain Model. Neuron Glia Biol. 2010, 6, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Hanani, M. Satellite Glial Cells in Sensory Ganglia: From Form to Function. Brain Res. Rev. 2005, 48, 457–476. [Google Scholar] [CrossRef] [PubMed]
- Ohara, P.T.; Vit, J.-P.; Bhargava, A.; Romero, M.; Sundberg, C.; Charles, A.C.; Jasmin, L. Gliopathic Pain: When Satellite Glial Cells Go Bad. Neuroscientist 2009, 15, 450–463. [Google Scholar] [CrossRef]
- Takeda, K.; Muramatsu, M.; Chikuma, T.; Kato, T. Effect of Memantine on the Levels of Neuropeptides and Microglial Cells in the Brain Regions of Rats with Neuropathic Pain. J. Mol. Neurosci. 2009, 39, 380–390. [Google Scholar] [CrossRef]
- Huang, T.-Y.; Belzer, V.; Hanani, M. Gap Junctions in Dorsal Root Ganglia: Possible Contribution to Visceral Pain. Eur. J. Pain 2010, 14, 49.e1–49.e11. [Google Scholar] [CrossRef]
- Jasmin, L.; Vit, J.-P.; Bhargava, A.; Ohara, P.T. Can Satellite Glial Cells Be Therapeutic Targets for Pain Control? Neuron Glia Biol. 2010, 6, 63–71. [Google Scholar] [CrossRef]
- Costa, F.A.L.; Neto, F.L.M. Satellite Glial Cells in Sensory Ganglia: Its Role in Pain. BJAN 2015, 65, 73–81. [Google Scholar] [CrossRef]
- Ye, Y.; Salvo, E.; Romero-Reyes, M.; Akerman, S.; Shimizu, E.; Kobayashi, Y.; Michot, B.; Gibbs, J. Glia and Orofacial Pain: Progress and Future Directions. Int. J. Mol. Sci. 2021, 22, 5345. [Google Scholar] [CrossRef]
- Kupczyk, P.; Rykala, M.; Serek, P.; Pawlak, A.; Slowikowski, B.; Holysz, M.; Chodaczek, G.; Madej, J.P.; Ziolkowski, P.; Niedzwiedz, A. The Cannabinoid Receptors System in Horses: Tissue Distribution and Cellular Identification in Skin. J. Vet. Intern. Med. 2022, 36, 1508–1524. [Google Scholar] [CrossRef]
- Galiazzo, G.; Giancola, F.; Stanzani, A.; Fracassi, F.; Bernardini, C.; Forni, M.; Pietra, M.; Chiocchetti, R. Localization of Cannabinoid Receptors CB1, CB2, GPR55, and PPARα in the Canine Gastrointestinal Tract. Histochem. Cell Biol. 2018, 150, 187–205. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Receptors | CB1R | CB2R | TRPV1 | GPR55 | PPAR-ƴ |
|---|---|---|---|---|---|
| Neurons | + IF Cryo + IHC iFFPE | ++ IF Cryo ++ IHC FFPE | +++ IF Cryo +++ IHC FFPE | +/− IF Cryo ++/+++ IHC FFPE | +++ IF Cryo +++ IHC FFPE |
| Satellite glial cells | − IF Cryo − IHC FFPE | ++ IF Cryo +++ IHC FFPE | − IF Cryo − IHC FFPE | − IF Cryo ++ IHC FFPE | +++ IF Cryo +++ IHC FFPE |
| Inflammatory or immune cells | − IF Cryo − IHC FFPE | − IF Cryo − IHC FFPE | − IF Cryo − IHC FFPE | − IF Cryo +++ IHC FFPE | +++ IF Cryo +++ IHC FFPE |
| Primary Antibody | Host | Code | Dilution | Source |
|---|---|---|---|---|
| CB1R | Rabbit | ab23703 | 1:100 (IF); 1:400 (IHC) | Abcam |
| CB2R | Rabbit | PA1-744 | 1:250 (IF); 1:200 (IHC) | Thermo Fisher |
| GPR55 | Rabbit | NB110-55498 | 1:200 (IF); 1:200 (IHC) | Novus Biol. |
| GFAP | Chicken | Ab4674 | 1:800 (IF) | Abcam |
| IBA1 | Goat | NB100-1028 | 1:80 (IF) | Novus Biol. |
| PPARɣ | Rabbit | Ab45036 | 1:300 (IF); 1:400 (IHC) | Abcam |
| TRPV1 | Rabbit | ACC-030 | 1:200 (IF); 1:300 (IHC) | Alomone |
| Double Labelling Experiment | Secondary Reagent Combination |
|---|---|
| Rabbit anti-PPARγ | Donkey anti-Rabbit 488 |
| Chicken anti-GFAP | Donkey anti-Chicken TRITC |
| Rabbit anti-PPARγ | Donkey anti-Rabbit 488 |
| Goat anti-IBA1 | Donkey anti-Goat 594 |
| Rabbit anti-GPR55 | Donkey anti-Rabbit 488 |
| Chicken anti-GFAP | Donkey anti-Chicken TRITC |
| Rabbit anti-GPR55 | Donkey anti-Rabbit 488 |
| Goat anti-IBA1 | Donkey anti-Goat 594 |
| Secondary Antibody | Host | Code | Dilution | Source |
|---|---|---|---|---|
| Anti-chicken TRITC | Donkey | 703-025-155 | 1:200 | Jackson |
| Anti-goat 594 | Donkey | ab150132 | 1:600 | Abcam |
| Anti-rabbit 488 | Donkey | A-21206 | 1:1000 | Thermo Fisher |
| Anti-rabbit biotinylated | Goat | BA-1000 | 1:200 | Vector laboratories |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zamith Cunha, R.; Semprini, A.; Salamanca, G.; Gobbo, F.; Morini, M.; Pickles, K.J.; Roberts, V.; Chiocchetti, R. Expression of Cannabinoid Receptors in the Trigeminal Ganglion of the Horse. Int. J. Mol. Sci. 2023, 24, 15949. https://doi.org/10.3390/ijms242115949
Zamith Cunha R, Semprini A, Salamanca G, Gobbo F, Morini M, Pickles KJ, Roberts V, Chiocchetti R. Expression of Cannabinoid Receptors in the Trigeminal Ganglion of the Horse. International Journal of Molecular Sciences. 2023; 24(21):15949. https://doi.org/10.3390/ijms242115949
Chicago/Turabian StyleZamith Cunha, Rodrigo, Alberto Semprini, Giulia Salamanca, Francesca Gobbo, Maria Morini, Kirstie J. Pickles, Veronica Roberts, and Roberto Chiocchetti. 2023. "Expression of Cannabinoid Receptors in the Trigeminal Ganglion of the Horse" International Journal of Molecular Sciences 24, no. 21: 15949. https://doi.org/10.3390/ijms242115949