
Complement C3-Deficiency-Induced Constipation in FVB/N-C3em1Hlee/Korl Knockout Mice Was Significantly Relieved by Uridine and Liriope platyphylla L. Extracts
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Effects of Urd and AEtLP on Excretion Parameters in C3-Deficiency-Induced Constipation
2.2. Effects of Urd and AEtLP on GI Transit in C3-Deficiency-Induced Constipation
2.3. Effects of Urd and AEtLP on the Histopathological Structure of the Mid Colon in C3-Deficiency-Induced Constipation
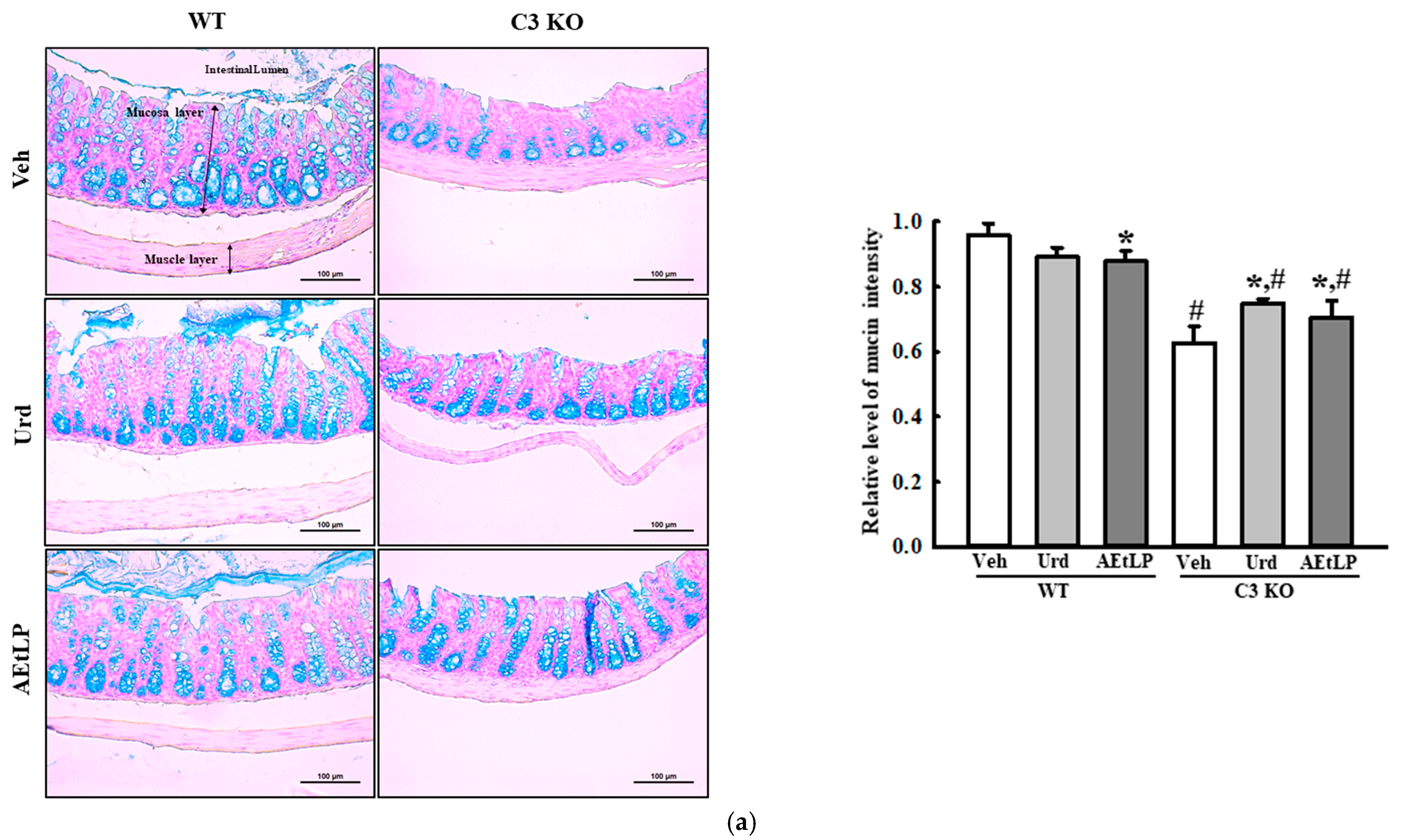
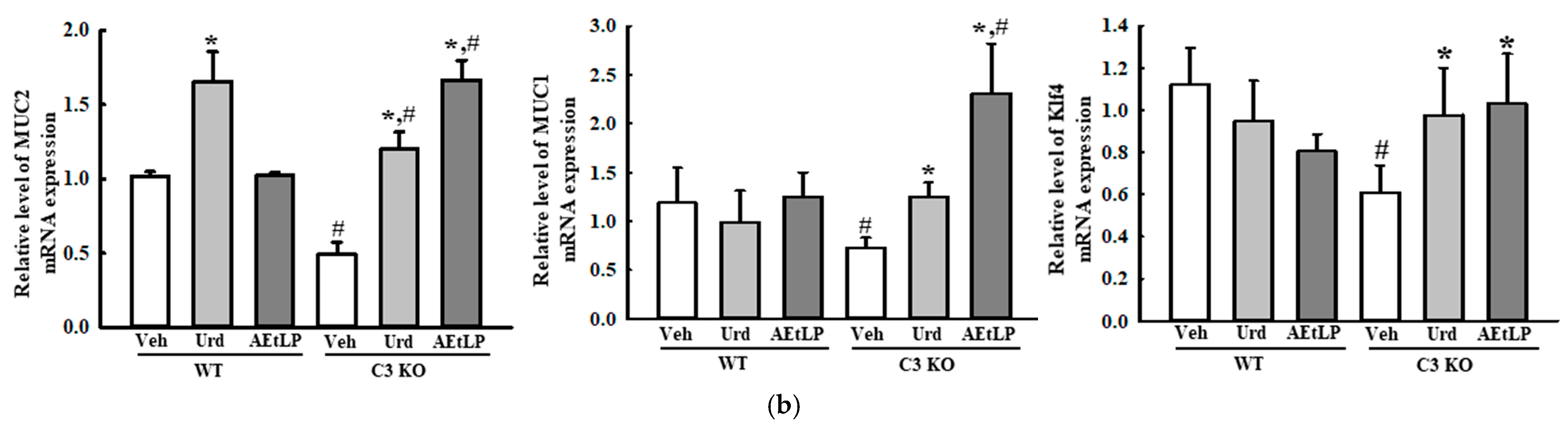
2.4. Effects of Urd and AEtLP on Mucin Secretion and Water Retention in C3-Deficiency-Induced Constipation
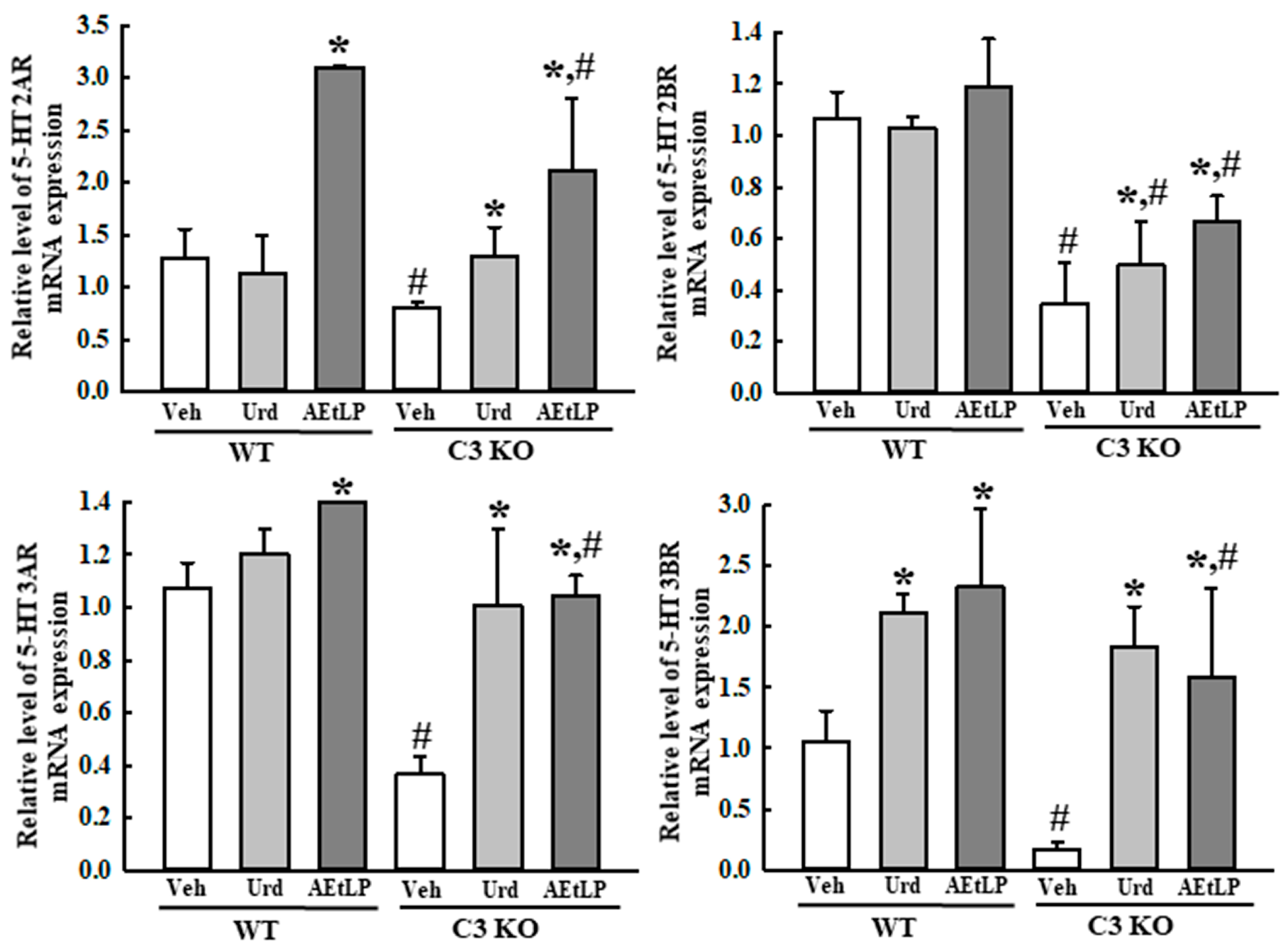
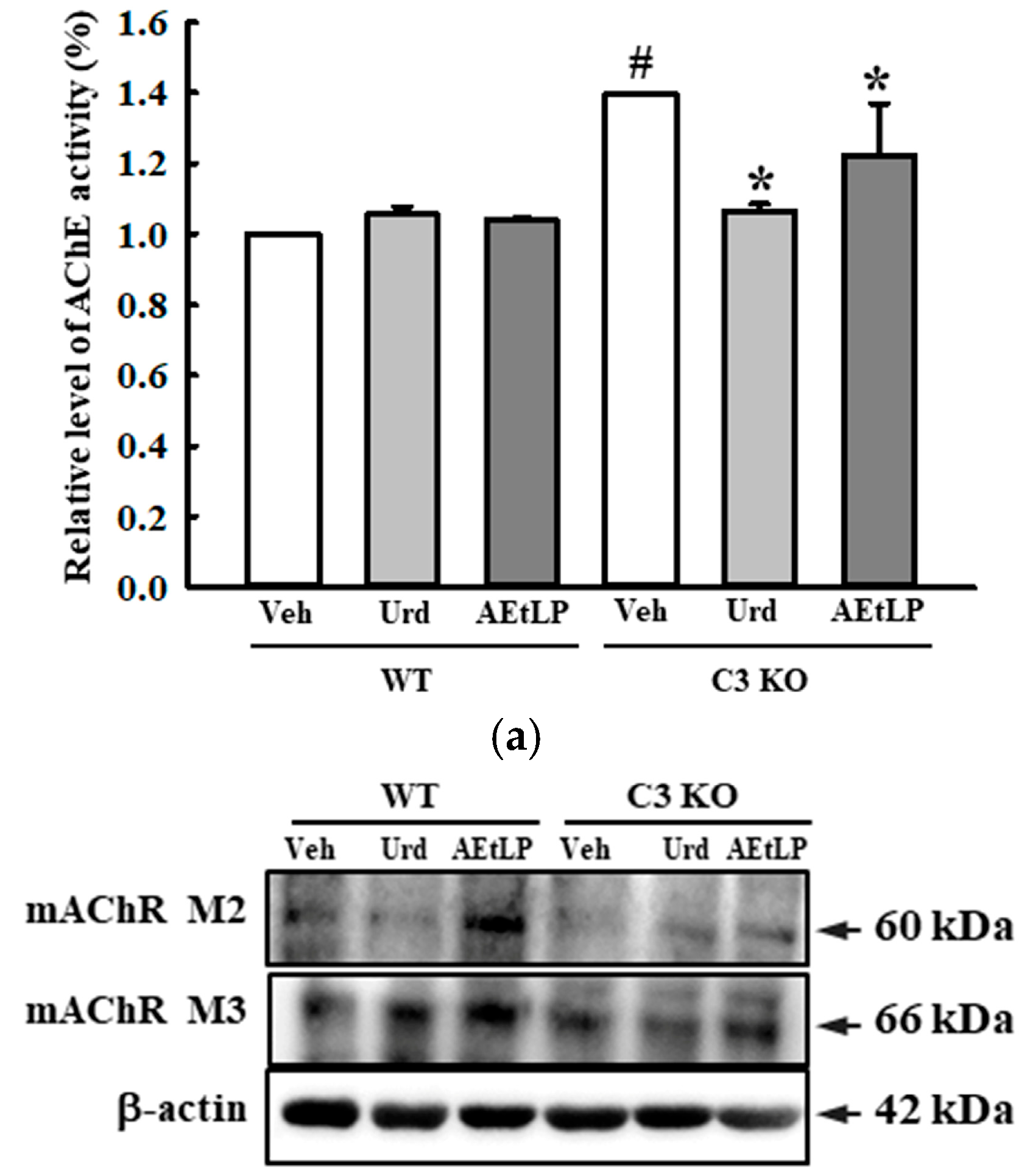
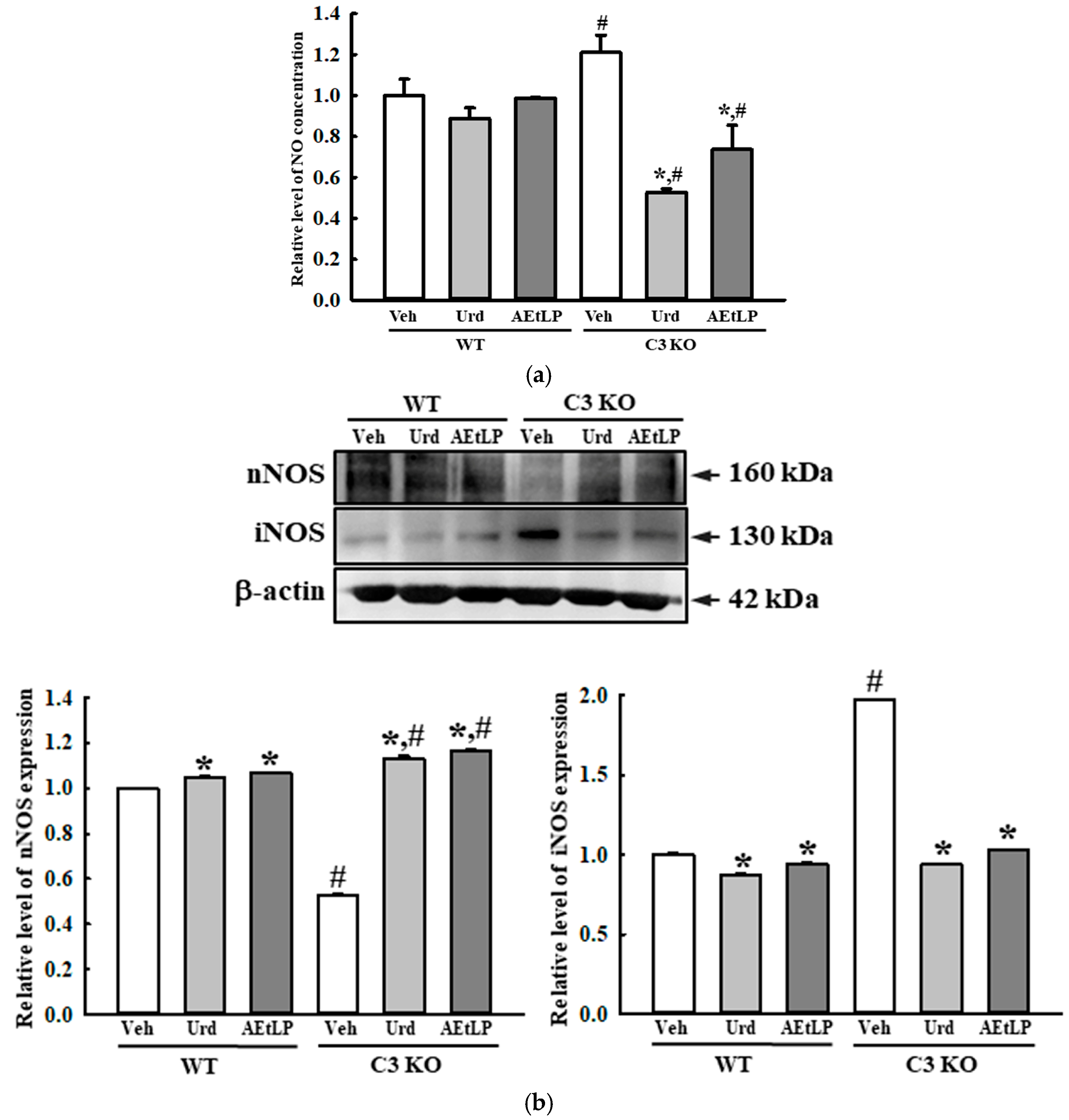
2.5. Effects of Urd and AEtLP on ENS in C3-Deficiency-Induced Constipation
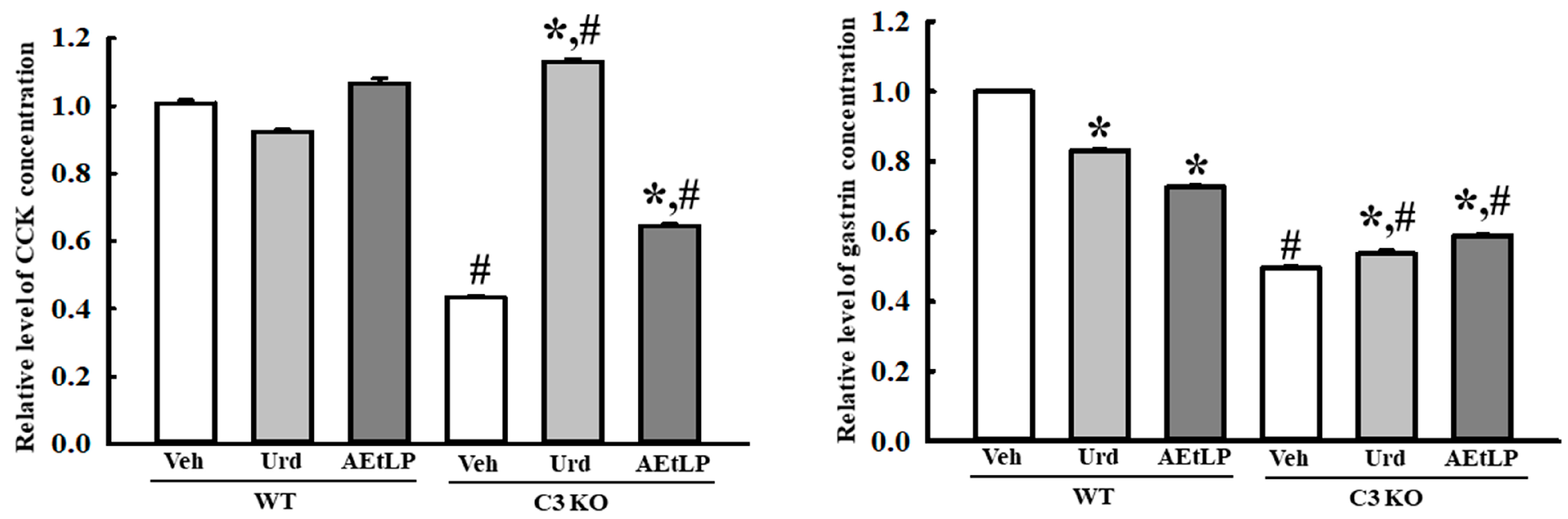
2.6. Effects of Urd and AEtLP on the Regulation of the GI Hormone in C3-Deficiency-Induced Constipation
3. Discussion
4. Materials and Methods
4.1. Preparation of Urd and AEtLP
4.2. Care and Use of Animals
4.3. Experimental Design for C3 KO Mice
4.4. Measurement of Food Intake and Water Consumption
4.5. Measurement of Excretion Parameter
4.6. Analysis of GI Motility and Length
4.7. Histopathological Analysis
4.8. Western Blot Analysis
4.9. RT-qPCR Analysis
4.10. AChE Activity Assay
4.11. Measurement of NO Concentrations
4.12. Measurement of GI Hormones Concentrations
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walia, R.; Mahajan, L.; Steffen, R. Recent advances in chronic constipation. Curr. Opin. Pediatr. 2009, 21, 661–666. [Google Scholar] [PubMed]
- McCallum, I.J.; Ong, S.; Mercer-Jones, M. Chronic constipation in adults. BMJ 2009, 338, b831. [Google Scholar] [PubMed]
- Diaz, S.; Bittar, K.; Mendez, M.D. Constiaption, 1st ed; StatPearls: Tresure Island, FL, USA, 2023. [Google Scholar]
- Hasler, W.L.; Wilson, L.A.; Nguyen, L.A.; Snape, W.J.; Abell, T.L.; Koch, K.L.; McCallum, R.W.; Pasricha, P.J.; Sarosiek, I.; Farrugia, G. Opioid use and potency are associated with clinical features, quality of life, and use of resources in patients with gastroparesis. Clin. Gastroenterol. Hepatol. 2019, 17, 1285–1294. [Google Scholar] [PubMed]
- De Andino, N.M. Current treatment paradigm and landscape for the management of chronic idiopathic constipation in adults: Focus on plecanatide. J. Am. Assoc. Nurse Pract. 2018, 30, 412–420. [Google Scholar]
- Andresen, V.; Layer, P. Medical therapy of constipation: Current standards and beyond. Visc. Med. 2018, 34, 123–127. [Google Scholar] [CrossRef]
- Zarate, N.; Spencer, N.J. Chronic constipation: Lessons from animal studies. Best. Pract. Res. Clin. Gastroenterol. 2011, 25, 59–71. [Google Scholar]
- Kim, J.E.; Seol, A.; Choi, Y.J.; Lee, S.J.; Jin, Y.J.; Roh, Y.J.; Song, H.J.; Hong, J.T.; Hwang, D.Y. Similarities and differences in constipation phenotypes between lep knockout mice and high fat diet-induced obesity mice. PLoS ONE 2022, 17, e0276445. [Google Scholar]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Jang, M.; Hong, J.T.; Hwang, D.Y. Loperamide-induced constipation activates inflammatory signaling pathways in the mid colon of SD Rats via complement C3 and its receptors. Curr. Mol. Med. 2022, 22, 458–469. [Google Scholar]
- Kim, J.E.; Song, H.J.; Choi, Y.J.; Jin, Y.J.; Roh, Y.J.; Seol, A.; Park, S.H.; Park, J.M.; Kang, H.G.; Hwang, D.Y. Improvement of the intestinal epithelial barrier during laxative effects of phlorotannin in loperamide-induced constipation of SD rats. Lab. Anim. Res. 2023, 39, 1. [Google Scholar] [CrossRef]
- Kakino, M.; Tazawa, S.; Maruyama, H.; Tsuruma, K.; Araki, Y.; Shimazawa, M.; Hara, H. Laxative effects of agarwood on low-fiber diet-induced constipation in rats. BMC Complement. Altern. Med. 2010, 10, 68. [Google Scholar] [CrossRef]
- Young, A.; Viswanath, A.; Kalladka, M.; Khan, J.; Eliav, E.; Diehl, S.R. Mouse model demonstrates strain differences in susceptibility to opioid side effects. Neurosci. Lett. 2018, 675, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zou, X.; Kuang, G.; Ren, Y.; Deng, C.; Lin, Q.; Zhao, X.; Xu, S.; Song, J.L. Preventative effects of fermented Chimonobambusa quadrangularis shoot on activated carbon-induced constipation. Exp. Ther. Med. 2017, 13, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Ahrenstedt, O.; Kuntson, L.; Nilsson, B.; Nilsson-Ekdahl, K.; Odlind, B.; Hallgren, R. Enhanced local production of complement components in the small intestines of patients with Crohn’s disease. N. Engl. J. Med. 1990, 322, 1345–1349. [Google Scholar] [CrossRef] [PubMed]
- Halstensen, T.S.; Mollnes, T.E.; Garred, P.; Fausa, O.; Brandtzaeg, P. Surface epithelium related activation of complement differs in Crohn’s disease and ulcerative colitis. Gut 1992, 33, 902–908. [Google Scholar] [CrossRef]
- Laufer, J.; Oren, R.; Goldberg, I.; Horwitz, A.; Kopolovic, J.; Chowers, Y.; Passwell, J.H. Cellular localization of complement C3 and C4 transcripts in intestinal specimens from patients with Crohn’s disease. Clin. Exp. Immunol. 2000, 120, 30–37. [Google Scholar] [CrossRef]
- Sugihara, T.; Kobori, A.; Imaeda, H.; Tsujikawa, T.; Amagase, K.; Takeuchi, K.; Fujuyama, Y.; Andoh, A. The increased mucosal mRNA expressions of complement C3 and interleukin-17 in inflammatory bowel disease. Clin. Exp. Immunol. 2010, 160, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Kim, J.E.; Choi, Y.J.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Hong, J.T.; Lee, H.; Hwang, D.Y. Deficiency of complement component 3 may be linked to the development of constipation in FVB/N-C3em1Hlee/Korl mice. FASEB J. 2021, 35, e21221. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Jin, Y.J.; Lee, H.; Hwang, D.Y. Promotion of the inflammatory response in mid colon of complement component 3 knockout mice. Sci. Rep. 2022, 12, 1700. [Google Scholar] [CrossRef]
- Choi, Y.J.; Song, H.J.; Kim, J.E.; Lee, S.J.; Jin, Y.J.; Roh, Y.J.; Seol, A.; Kim, H.S.; Hwang, D.Y. Dysregulation of the enteric nervous system in the mid colon of complement component 3 knockout mice with constipation phenotypes. Int. J. Mol. Sci. 2022, 23, 6862. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, J.E.; Lee, S.J.; Gong, J.E.; Son, H.J.; Hong, J.T.; Hwang, D.Y. Dysbiosis of fecal microbiota from complement 3 knockout mice with constipation phenotypes contributes to development of defecation delay. Front. Physiol. 2021, 12, 650789. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Ko, J.; Hong, J.T.; Hwang, D.Y. Aqueous extracts of Liriope platyphylla induced significant laxative effects on loperamide-induced constipation of SD rats. BMC Complement. Altern. Med. 2013, 13, 333. [Google Scholar] [CrossRef]
- Kim, J.E.; Go, J.; Sung, J.E.; Lee, H.A.; Yun, W.B.; Hong, J.T.; Hwang, D.Y. Uridine stimulate laxative effect in the loperamide-induced constipation of SD rats through regulation of the mAChRs signaling pathway and mucin secretion. BMC Gastroenterol. 2017, 17, 21. [Google Scholar] [CrossRef]
- Kim, J.E.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, H.G.; Choi, Y.H.; Hwang, D.Y. Effects of five candidate laxatives derived from Liriope platyphylla on the 5-HT receptor signaling pathway in three cell types present in the transverse colon. Mol. Med. Rep. 2017, 15, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Go, J.; Koh, E.K.; Song, S.H.; Sung, J.E.; Lee, H.A.; Lee, Y.H.; Hong, J.T.; Hwang, D.Y. Gallotannin-enriched extract isolated from Galla Rhois may be a functional candidate with laxative effects for treatment of loperamide-induced constipation of SD rats. PLoS ONE 2016, 11, e0161144. [Google Scholar]
- Kim, J.E.; Park, J.W.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Choi, Y.; Lee, Y.J.; Seo, S.; Hong, J.T.; Hwang, D.Y. Laxative effect of Spicatoside a by cholinergic regulation of enteric nerve in loperamide-induced constipation: ICR mice model. Molecules 2019, 24, 896. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Lee, Y.J.; Sung, J.E.; Jung, Y.S.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Antioxidant activity and laxative effects of tannin-enriched extract of Ecklonia cava in loperamide-induced constipation of SD rats. PLoS ONE 2021, 16, e0246363. [Google Scholar]
- Kim, J.E.; Yun, W.B.; Lee, M.L.; Choi, J.Y.; Park, J.J.; Kim, H.R.; Song, B.R.; Hong, J.T.; Song, H.K.; Hwang, D.Y. Synergic laxative effects of an herbal mixture of Liriope platyphylla, Glycyrrhiza uralensis, and Cinnamomum cassia in loperamide-induced constipation of Sprague Dawley rats. J. Med. Food. 2019, 22, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Park, J.W.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Choi, Y.S.; Lee, Y.J.; Lee, H.S.; Hong, J.T.; Hwang, D.Y. Anti-inflammatory response and muscarinic cholinergic regulation during the laxative effect of Asparagus cochinchinensis in loperamide-induced constipation of SD rats. Int. J. Mol. Sci. 2019, 20, 946. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, M.R.; Park, J.J.; Choi, J.Y.; Song, B.R.; Son, H.J.; Choi, Y.W.; Kim, K.M.; Hong, J.T.; Hwang, D.Y. Quercetin promotes gastrointestinal motility and mucin secretion in loperamide-induced constipation of SD rats through regulation of the mAChRs downstream signal. Pharm. Biol. 2018, 56, 309–317. [Google Scholar] [CrossRef]
- Dobolyi, A.; Juhász, G.; Kovács, Z.; Kardos, J. Uridine function in the central nervous system. Curr. Top. Med. Chem. 2011, 11, 1058–1067. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Glycogen Metabolism, 5th ed.; W.H. Freeman: New York, NY, USA, 2022; pp. 865–900. [Google Scholar]
- Van Groeningen, C.J.; Peters, G.J.; Pinedo, H.M. Modulation of fluorouracil toxicity with uridine. Semin. Oncol. 1992, 19, 148–154. [Google Scholar] [PubMed]
- Van Groeningen, C.J.; Peters, G.J.; Pinedo, H.M. Reversal of 5-fluorouracil-induced toxicity by oral administration of uridine. Ann. Oncol. 1993, 4, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, X.; Zhou, J.; Yuan, J.; Cui, B.; An, R.; Hu, Z. Traditional Chinese formula, lubricating gut pill, improves loperamide-induced rat constipation involved in enhance of Cl− secretion across distal colonic epithelium. J. Ethnopharmacol. 2010, 130, 347–353. [Google Scholar] [PubMed]
- MacNicol, M.; Schulman, H. Cross-talk between protein kinase C and multifunctional Ca2+/calmodulin-dependent protein kinase. J. Biol. Chem. 1992, 267, 12197–12201. [Google Scholar]
- Auernhammer, C.J.; Renner, U.; Müller, O.A.; Stalla, J.; Stalla, G.K. Loperamide inhibits corticotrophic cell function by a naloxone-lnsensitive mechanism in the rat in vitro. Neuroendocrinology 1993, 57, 1019–1027. [Google Scholar]
- Chen, W.; Chung, H.H.; Cheng, J.T. Opiate-induced constipation related to activation of small intestine opioid μ2-receptors. World J. Gastroenterol. 2012, 18, 1391–1396. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, J.W.; Cho, H.R.; Kim, K.Y.; Lee, J.K.; Sohn, J.H.; Ku, S.K. Laxative effects of fermented rice extract in rats with loperamide-induced constipation. Exp. Ther. Med. 2014, 8, 1847–1854. [Google Scholar]
- Kojima, R.; Nozawa, K.; Doihara, H.; Keto, Y.; Kaku, H.; Yokoyama, T.; Itou, H. Effects of novel TRPA1 receptor agonist ASP7663 in models of drug-induced constipation and visceral pain. Eur. J. Pharmacol. 2014, 723, 288–293. [Google Scholar] [CrossRef]
- Kim, J.E.; Yun, W.B.; Sung, J.E.; Lee, H.A.; Choi, J.Y.; Choi, Y.S.; Jung, Y.S.; Kim, K.S.; Hwang, D.Y. Characterization the response of Korl: ICR mice to loperamide induced constipation. Lab. Anim. Res. 2016, 32, 231–240. [Google Scholar] [CrossRef]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X.Y. Regulation, signaling and physiological functions of G-proteins. J. Mol. Biol. 2016, 19, 3850–3868. [Google Scholar]
- Koehl, A.; Hu, H.; Maeda, S.; Zhang, Y.; Qu, Q.; Paggi, J.M.; Latorraca, N.R.; Hilger, D.; Dawson, R.; Matile, H.; et al. Structure of the µ-opioid receptor-Gi protein complex. Nature 2018, 558, 547–552. [Google Scholar]
- Phieler, J.; Garcia-Martin, R.; Lambris, J.D. The role of the complement system in metabolic organs and metabolic diseases. In Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2013; Volume 25, pp. 47–53. [Google Scholar]
- Park, J.W.; Kim, J.E.; Kang, M.J.; Choi, H.J.; Bae, S.J.; Hwang, D.Y. Compensatory role of C3 convertase on the strain difference for C3 protein expression in FVB/N, C3H/HeN and C57BL/6N mice. Lab. Anim. Res. 2020, 36, 4. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Subramanian, H.; Gupta, K.; Ali, H. Regulation of C3a receptor signaling in human mast cells by G protein coupled receptor kinases. PLoS ONE 2011, 6, e22559. [Google Scholar]
- McCorvy, J.D.; Roth, B.L. Structure and function of serotonin G protein coupled receptors. Pharmacol. Ther. 2015, 150, 129–142. [Google Scholar] [PubMed]
- Kruse, A.C.; Kobilka, B.K.; Gautam, D.; Sexton, P.M.; Christopoulos, A.; Wess, J. Muscarinic acetylcholine receptors: Novel opportunities for drug development. Nat. Rev. Drug Discov. 2014, 13, 549–560. [Google Scholar] [PubMed]
- Lee, Y.C.; Lee, J.C.; Seo, Y.B.; Kook, Y.B. Liriopis tuber inhibit OVA-induced airway inflammation and bronchial hyperresponsiveness in murine model of asthma. J. Ethnopharmacol. 2005, 101, 144–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, Y.K.; Nam, S.H.; Choi, S.I.; Goo, J.S.; Jang, M.J.; Lee, H.S.; Son, H.J.; Lee, C.Y.; Hwang, D.Y. The symptoms of atopic dermatitis in NC/Nga mice were significantly relieved by the water extract of Liriope platyphylla. Lab. Anim. Res. 2010, 26, 377–384. [Google Scholar] [CrossRef]
- Choi, S.B.; Wha, J.D.; Park, S. The insulin sensitizing effect of homoisoflavone-enriched fraction in Liriope Platyphylla Wang et Tang via PI3-kinase pathway. Life Sci. 2004, 75, 2653–2664. [Google Scholar] [CrossRef]
- Jeong, S.; Chae, K.; Jung, Y.S.; Rho, Y.H.; Lee, J.; Ha, J.; Yoon, K.H.; Kim, G.C.; Oh, K.S.; Shin, S.S. The Korean traditional medicine Gyeongshingangjeehwan inhibits obesity through the regulation of leptin and PPAR alpha action in OLETF rats. J. Ethnopharmacol. 2008, 119, 245–251. [Google Scholar] [CrossRef]
- Choi, S.I.; Go, J.; Kim, J.E.; Lee, Y.J.; Kwak, M.H.; Jung, Y.J.; Hwang, D.Y. Precautionary effects of red Liriope platyphylla on NGF secretion and aβ42 deposition under the preclinical stage of Alzheimer’s disease in Tg2576 mice. Lab. Anim. Res. 2013, 29, 212–220. [Google Scholar] [CrossRef]
- Lee, H.R.; Kim, J.E.; Goo, J.S.; Choi, S.I.; Hwang, I.S.; Lee, Y.J.; Son, H.J.; Lee, H.S.; Lee, J.S.; Hwang, D.Y. Red Liriope platyphylla contains a large amount of polyphenolic compounds which stimulate insulin secretion and suppress fatty liver formation through the regulation of fatty acid oxidation in OLETF rats. Int. J. Mol. Med. 2012, 30, 905–913. [Google Scholar] [CrossRef]
- Wang, H.C.; Wu, C.C.; Cheng, T.S.; Kuo, C.Y.; Tsai, Y.C.; Chiang, S.Y.; Wong, T.S.; Wu, Y.C.; Chang, F.R. Active constituents from Liriope platyphylla root against cancer growth In Vitro. Evid. Based Complement. Alternat. Med. 2013, 2013, 857929. [Google Scholar] [PubMed]
- Jiang, T.; Tang, X.Y.; Wu, J.S.; Wang, L.Q.; Huang, M.; Han, T.; Qin, L.P. Study on chemical constituents of the root of Liriope platyphylla. Zhong Yao Cai 2011, 34, 1537–1539. [Google Scholar] [PubMed]
- Jiang, T.; Huang, B.K.; Zhang, Q.Y.; Han, T.; Zheng, H.C.; Qin, L.P. Studies on chemical constituents of Liriope platyphylla. Zhong Yao Cai 2007, 30, 1079–1081. [Google Scholar] [PubMed]
- Hwang, D.Y. Enormous Potential for Development Liriope platyphylla Wang et Tang as a Therapeutic Drug on the Human Chronic Disease. In Alternative Medicine; IntechOpen: London, UK, 2012; Chapter 10. [Google Scholar]
- Kim, H.J.; Park, S.Y.; Kim, D.G.; Park, S.H.; Lee, H.; Hwang, D.Y.; Jung, M.H.; Ha, K.T.; Kim, B.J. Effects of the roots of Liriope platyphylla Wang et Tang on gastrointestinal motility function. J. Ethnopharmacol. 2016, 184, 144–153. [Google Scholar] [CrossRef]
- Jones-Bolin, S. Guidelines for the care and use of laboratory animals in biomedical research. Curr. Protoc. Pharmacol. 2012, 59, A.4B.1–A.4B.9. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]
- Lee, S.J.; Roh, Y.J.; Kim, J.E.; Jin, Y.J.; Song, H.J.; Seol, A.; Park, S.H.; Douangdeuane, B.; Souliya, O.; Choi, S.I. Protective effects of Dipterocarpus tuberculatus in blue light-induced macular degeneration in A2E-laden ARPE19 cells and retina of BALB/c mice. Antioxidants 2023, 12, 329. [Google Scholar] [CrossRef]
- Park, J.; Kim, J.E.; Jin, Y.J.; Roh, Y.J.; Song, H.J.; Seol, A.; Park, S.H.; Seo, S.; Lee, H.; Hwang, D.Y. Anti-atopic dermatitis effects of abietic acid isolated from rosin under condition optimized by response surface methodology in DNCB-spread BALB/c mice. Pharmaceuticals 2023, 16, 407. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C (T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, J.E.; Choi, Y.J.; Gong, J.E.; Park, S.H.; Douangdeuane, B.; Souliya, O.; Park, J.M.; Lee, H.S.; Kim, B.H. Therapeutic effects of Dipterocarpus tuberculatus with high antioxidative activity against UV-induced photoaging of NHDF cells and nude mice. Antioxidants 2021, 10, 791. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, Y.; Zhang, X.Y.; Liu, X.; Liu, X.; Wu, X.; Jose, P.A.; Duan, S.; Xu, F.J.; Yang, Z. Intestinal gastrin/CCKBR (Cholecystokinin B Receptor) ameliorates salt-sensitive hypertension by inhibiting intestinal Na+/H+ exchanger 3 activity through a PKC (Protein Kinase C)-mediated NHERF1 and NHERF2 pathway. Hypertension 2022, 79, 1668–1679. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories | WT | C3 KO | ||||
|---|---|---|---|---|---|---|
| Veh | Urd | AEtLP | Veh | Urd | AEtLP | |
| Stool number (ea) | 105.3 ± 7.6 | 95.3 ± 1.2 | 131 ± 4.6 * | 53 ± 10 # | 121 ± 19.8 *,# | 91.7 ± 4.9 *,# |
| Stool weight (g) | 0.9 ± 0.3 | 0.9 ± 0.3 | 0.9 ± 0.2 | 0.5 ± 0.1 # | 0.9 ± 1.1 | 1.3 ± 0.2 *,# |
| Stool water contents (%) | 20 ± 2.2 | 22.9 ± 3 | 43.4 ± 7.3 * | 8 ± 1.2 # | 20.8 ± 1.6 * | 35.8 ± 4.8 *,# |
| Urine volume (mL) | 1 ± 0.09 | 1.3 ± 0.05 | 1.35 ± 0.05 | 0.95 ± 0.07 | 1.25 ± 0.07 | 0.8 ± 0.05 |
| Food intake (g) | 8 ± 1 | 12.4 ± 2.3 | 9.5 ± 0.5 | 14.1 ± 0.1 | 10.5 ± 0.7 | 16 ± 0.05 |
| Water consumption (mL) | 6 ± 1.4 | 5.8 ± 0.6 | 6.8 ± 1 | 6.7 ± 0.6 | 6.4 ± 0.6 | 7.7 ± 1.5 |
| Categories | Feature (Unit) | WT | C3 KO | ||||
|---|---|---|---|---|---|---|---|
| Veh | Urd | AEtLP | Veh | Urd | AEtLP | ||
| Crypt | Length (µm) | 140.2 ± 4.5 | 145.7 ± 8.9 | 147.8 ± 2.9 | 78.3 ± 2.9 * | 136.2 ± 11.2 # | 170.6 ± 6.7 *,# |
| Flat luminal surface | Thickness (µm) | 14.1 ± 1.7 | 14.3 ± 1.5 | 17.02 ± 1.4 * | 8.7 ± 1.0 * | 15 ± 1.5 # | 15 ± 0.7 # |
| Muscle layer | Thickness (µm) | 58.2 ± 1.7 | 56.2 ± 2.3 | 45.8 ± 2.4 * | 34.9 ± 0.9 * | 21.2 ± 0.9 *,# | 34.9 ± 1.8 * |
| Mucosa layer | Thickness (µm) | 192.8 ± 0.72 | 187.7 ± 9.8 | 172.2 ± 3.8 * | 107.7 ± 9.2 * | 115.2 ± 4.1 * | 149.5 ± 2.6 *,# |
| Epithelial cells | Enterocyte, goblet cells, and paneath cells | Regular arrangement | Regular arrangement | Regular arrangement | Irregular arrangement Cell-shaped destruction Decrease in cell number | Cell-shaped recovery Recovery of cell number | Cell-shaped recovery Recovery of cell number |
| Muscularis mucosae | Thickness (µm) | 8.18 ± 0.8 | 7.8 ± 0.5 | 7.8 ± 1.1 | 14.7 ± 0.8 * | 9.8 ± 1.0 # | 15.31 ± 3.1 * |
| Categories | Alteration Rate after Treatment | |||
|---|---|---|---|---|
| Urd | AEtLP | |||
| Lop- Induced Constipation | C3-Deficiency-Induced Constipation | Lop- Induced Constipation | C3-Deficiency-Induced Constipation | |
| Stool number (ea) | 85.2 ± 33% | 129.2 ± 7% | 67.8 ± 16% | 74.9 ± 26% |
| Stool weight (g) | 105.7 ± 49% | 82.2 ± 17% | 96.4 ± 0.4% | 161.7 ± 13% |
| Stool water contents (%) | 110.5 ± 12% | 159.5 ± 20% | 122 ± 73% | 339 ± 16% |
| Gastrointestinal motility (%) | ND | 36 ± 6% | ND | 27 ± 3% |
| Intestine length (µm) | ND | 2.7% | ND | 4.8 ± 2% |
| Mucosa layer (µm) | 275.3 ± 38% | 7.8 ± 5% | 10.3 ± 8.7% | 39.8 ± 10% |
| Muscle layer (µm) | 221.3 ± 25% | −39.8 ± 1% | 124 ± 24% | ND |
| Flat luminal surface thickness (μm) | 39.3 ± 4% | 72.6 ± 3% | 200 ± 0% | 73.3 ± 12% |
| AQP8 level (%) | 513.8 ± 28% | 118.8 ± 18% | ND | 126.8 ± 47% |
| MUC2 level (%) | 150 ± 0% | 140.2 ± 4% | ND | 220.8 ± 15% |
| mAChRM2 level (%) | 254.8 ± 43% | 28.6 ± 7% | −20 ± 4% | 50 ± 7% |
| mAChRM3 level (%) | 237.8 ± 40% | −3 ± 4.8% | −7.7 ± 0.3% | 60.7 ± 8% |
| PI3K level (%) | −43.6 ± 3% | −11.7 ± 2% | −47.7 ± 4% | −29.4 ± 1% |
| PKC level (%) | −92.8 ± 0.5% | −23 ± 1% | −37.9 ± 0.4% | −64 ± 0.8% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, H.-J.; Kim, J.-E.; Jin, Y.-J.; Roh, Y.-J.; Seol, A.; Kim, T.-R.; Park, K.-H.; Park, E.-S.; An, B.-S.; Yang, S.-Y.; et al. Complement C3-Deficiency-Induced Constipation in FVB/N-C3em1Hlee/Korl Knockout Mice Was Significantly Relieved by Uridine and Liriope platyphylla L. Extracts. Int. J. Mol. Sci. 2023, 24, 15757. https://doi.org/10.3390/ijms242115757
Song H-J, Kim J-E, Jin Y-J, Roh Y-J, Seol A, Kim T-R, Park K-H, Park E-S, An B-S, Yang S-Y, et al. Complement C3-Deficiency-Induced Constipation in FVB/N-C3em1Hlee/Korl Knockout Mice Was Significantly Relieved by Uridine and Liriope platyphylla L. Extracts. International Journal of Molecular Sciences. 2023; 24(21):15757. https://doi.org/10.3390/ijms242115757
Chicago/Turabian StyleSong, Hee-Jin, Ji-Eun Kim, You-Jeong Jin, Yu-Jeong Roh, Ayun Seol, Tae-Ryeol Kim, Ki-Ho Park, Eun-Seo Park, Beum-Soo An, Seung-Yun Yang, and et al. 2023. "Complement C3-Deficiency-Induced Constipation in FVB/N-C3em1Hlee/Korl Knockout Mice Was Significantly Relieved by Uridine and Liriope platyphylla L. Extracts" International Journal of Molecular Sciences 24, no. 21: 15757. https://doi.org/10.3390/ijms242115757