
Modulation of the Gut Microbiota by the Plantaricin-Producing Lactiplantibacillus plantarum D13, Analysed in the DSS-Induced Colitis Mouse Model
 , , , , ,
, , , , ,
Abstract
:1. Introduction
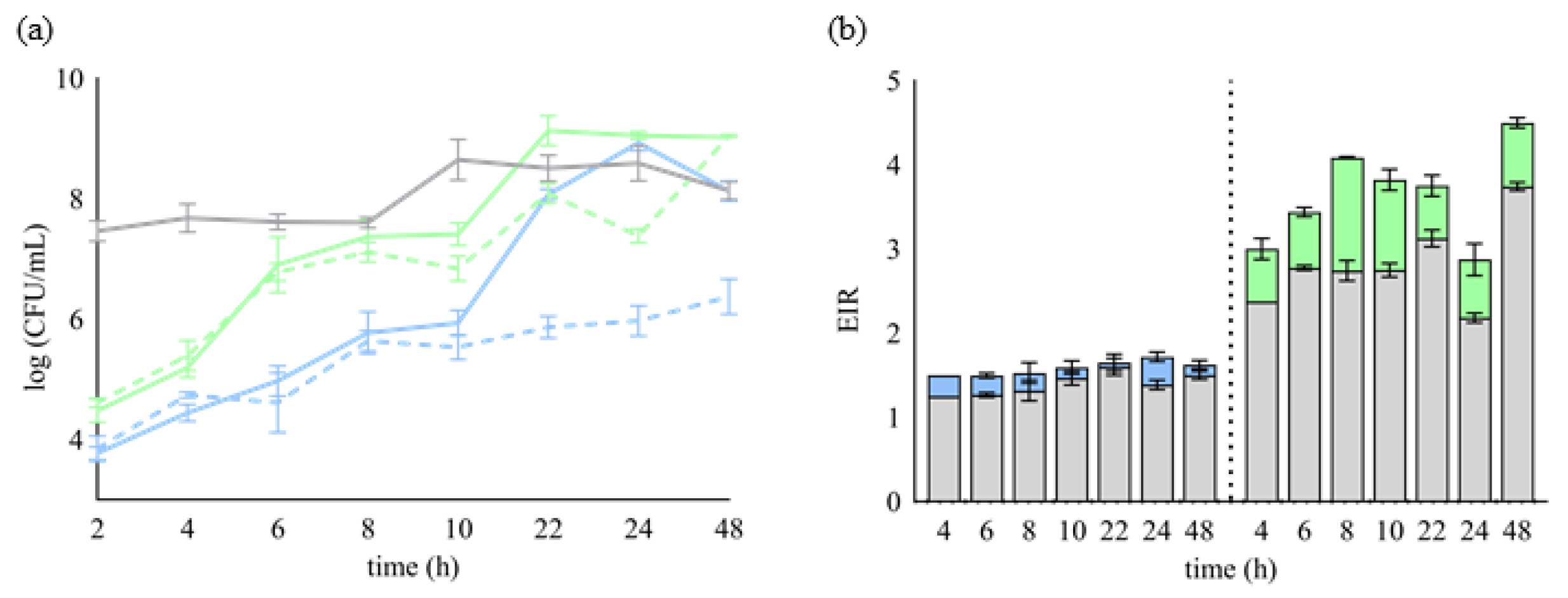
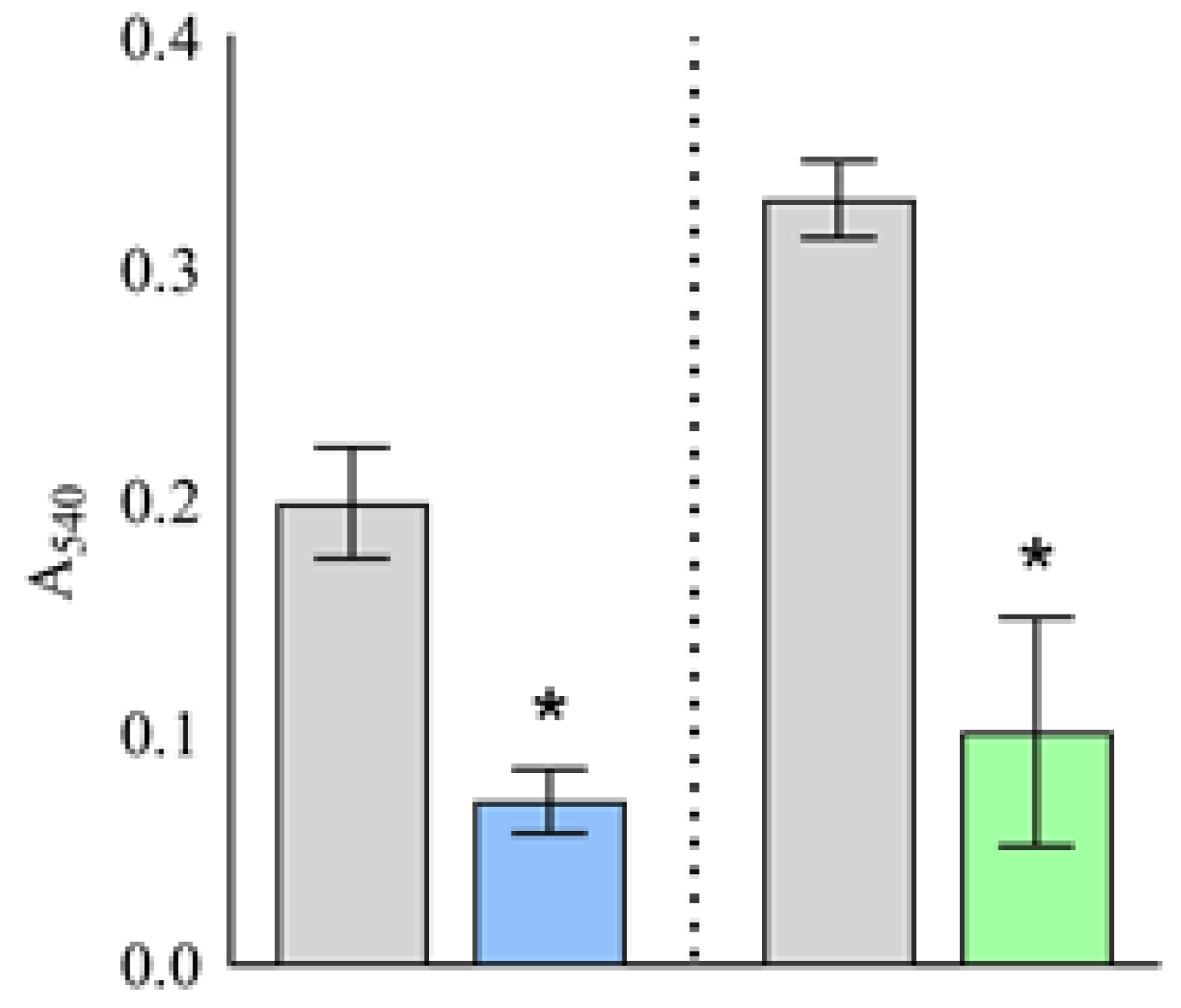
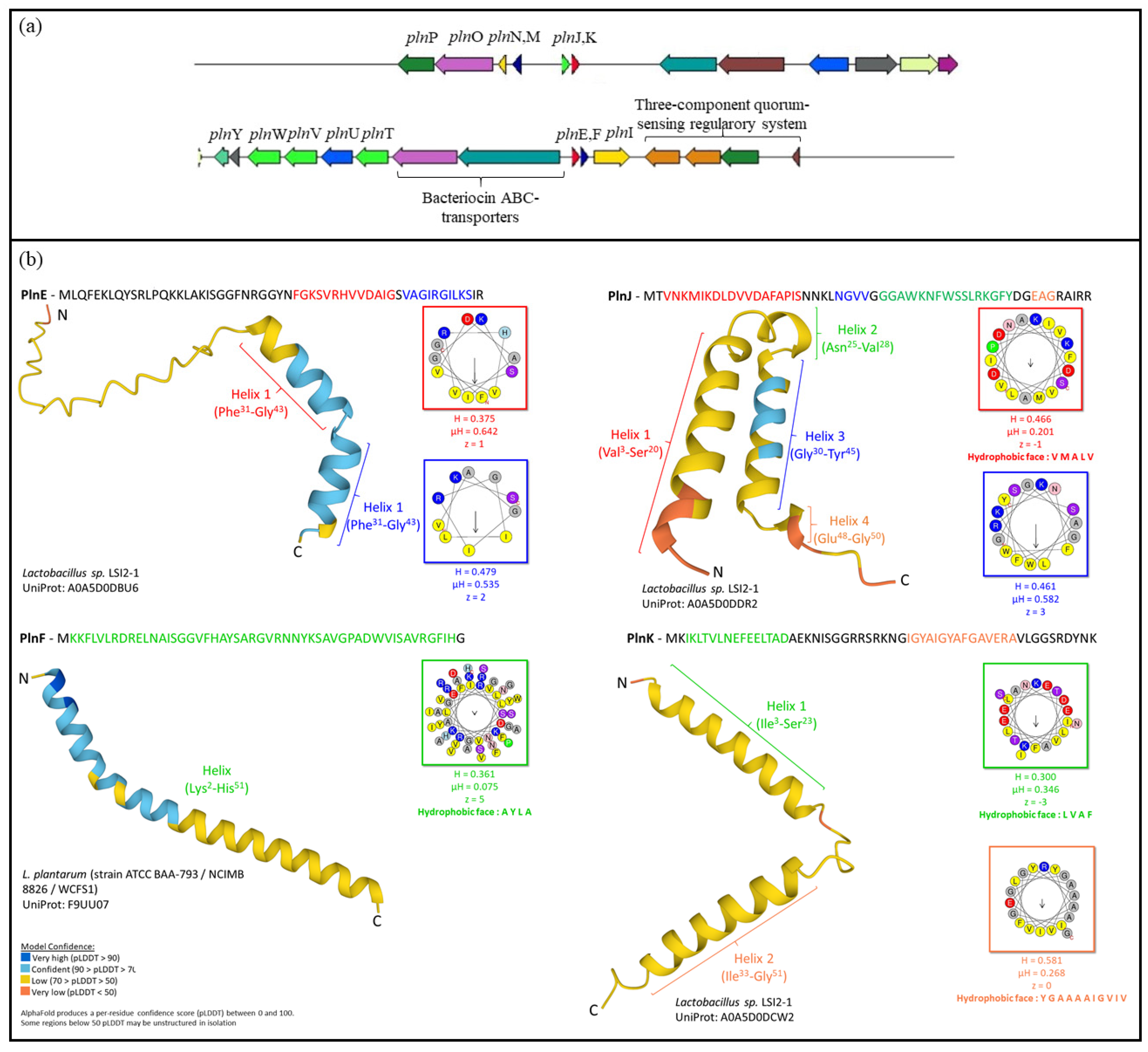
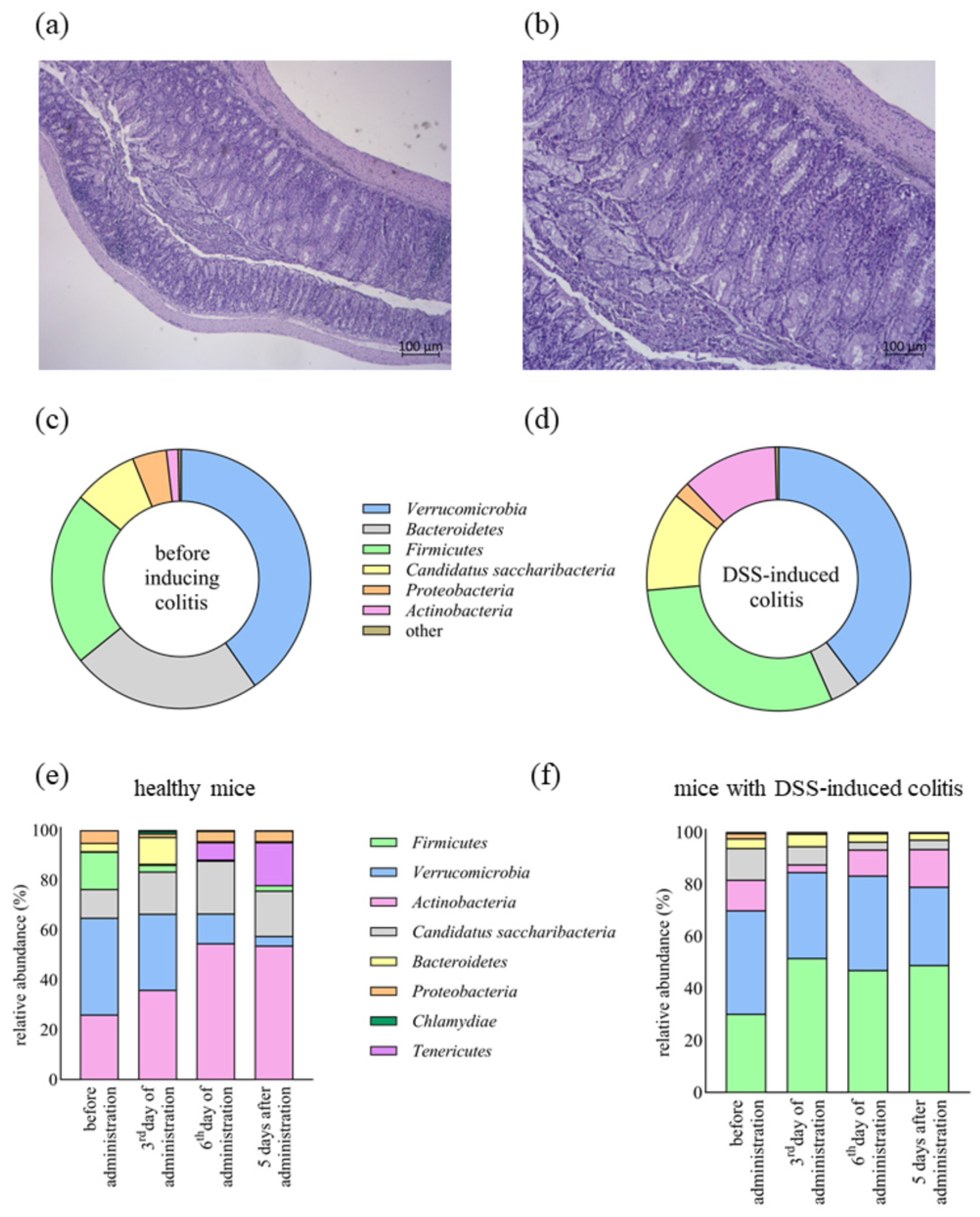
2. Results
2.1. Selection of Lb. plantarum Strains with Potential Plantaricin Production
2.2. Putative Plantaricins as Agents against Biofilm Formation
2.3. In Silico Characterisation of Plantaricin-Related Genes
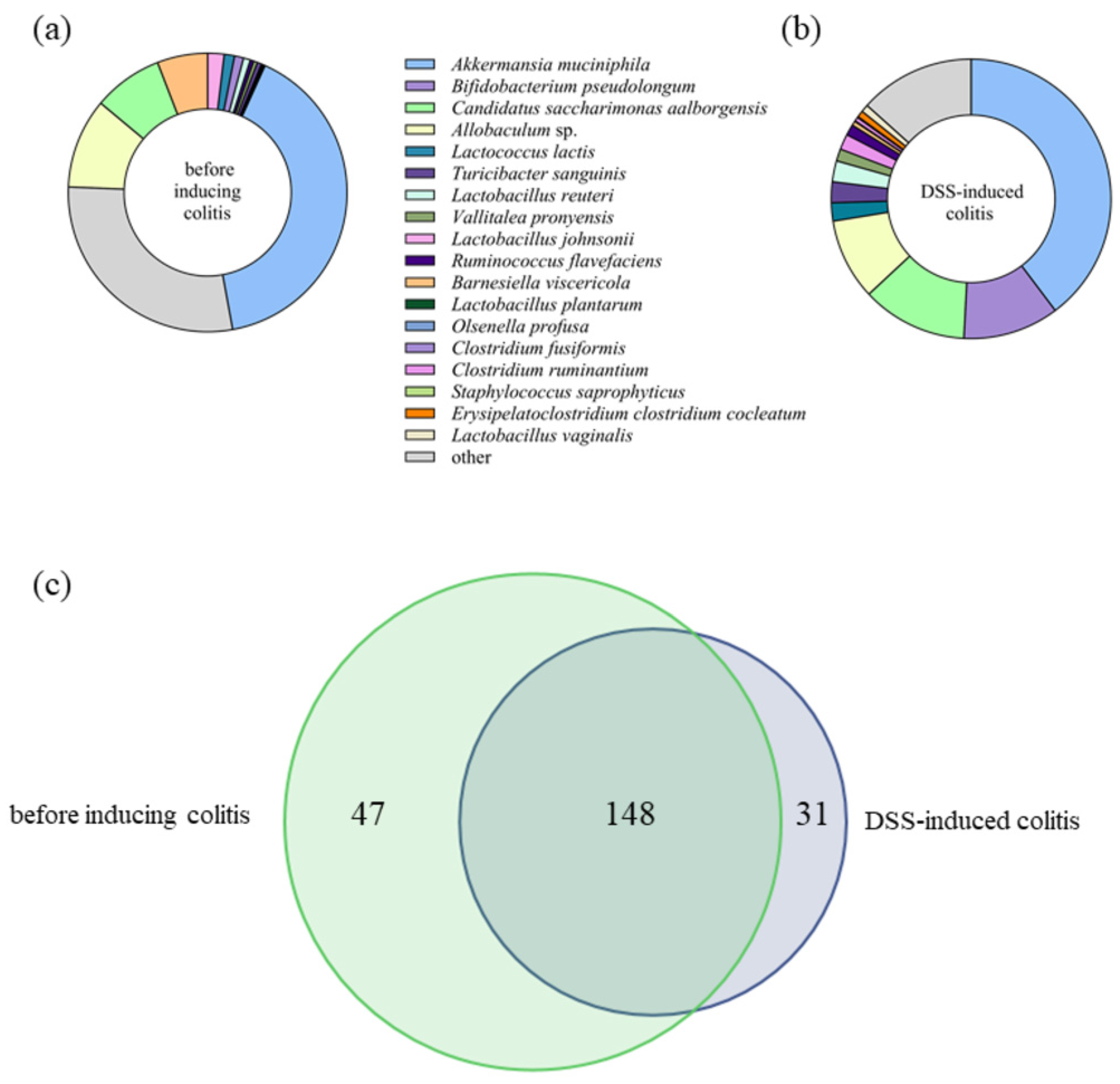
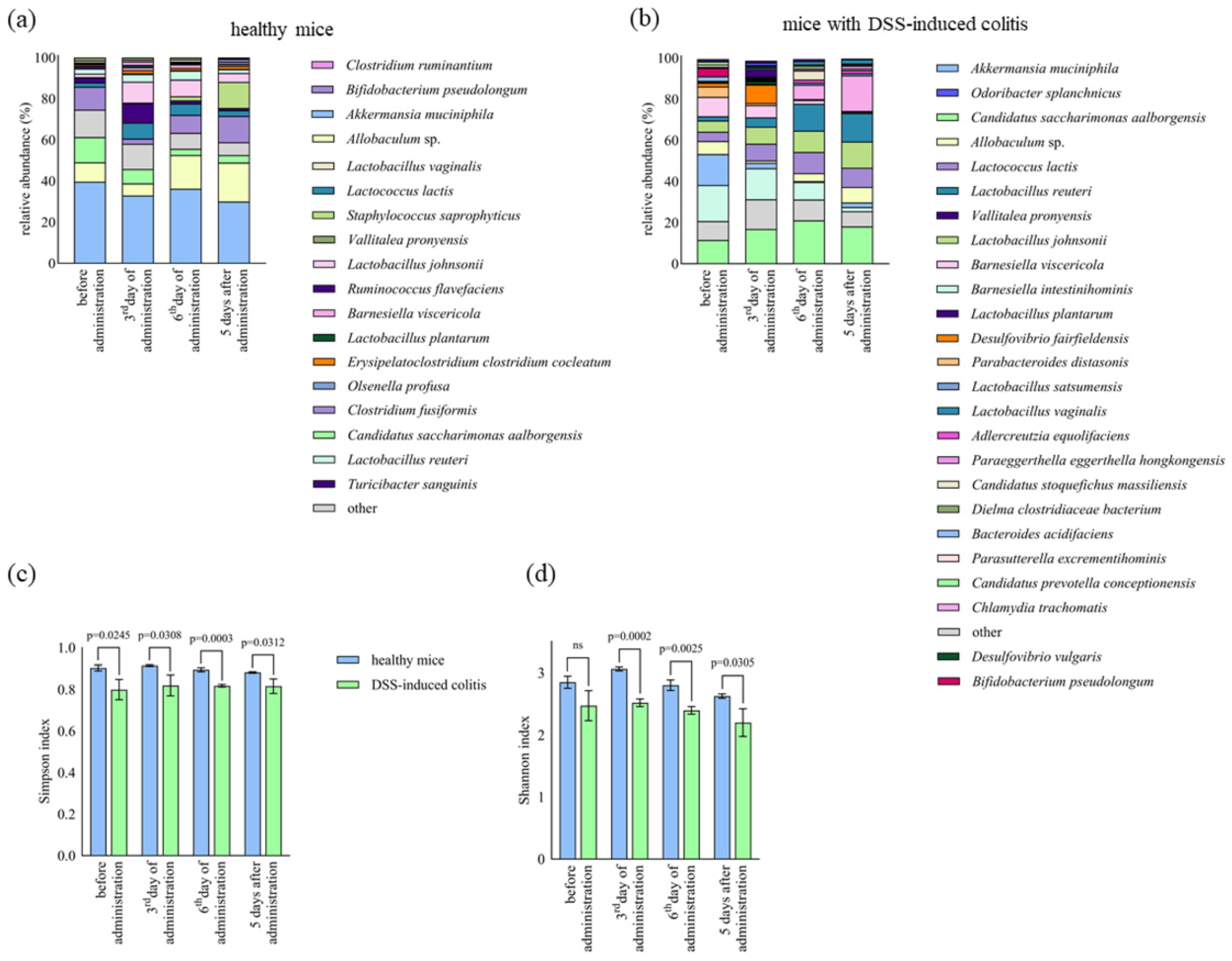
2.4. The Impact of Lb. plantarum D13 on the Composition of the Gut Microbiome of Mice with DSS-Induced Colitis
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Cultivation Conditions
4.2. Characterisation of the Antimicrobial Activity and the pln Loci
4.3. Inhibition of Biofilm Formation
4.4. Genome Sequencing and Analysis of the pln Loci
4.5. In Vivo Animal Trial
4.5.1. Experimental Animals
4.5.2. DSS-Induced Colitis and Administration of Lb. plantarum D13 Strain
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Quaglio, A.E.V.; Grillo, T.G.; De Oliveira, E.C.S.; Di Stasi, L.C.; Sassaki, L.Y. Gut microbiota, inflammatory bowel disease and colorectal cancer. World J. Gastroenterol. 2022, 28, 4053–4060. [Google Scholar] [CrossRef] [PubMed]
- Bouter, K.E.; van Raalte, D.H.; Groen, A.K.; Nieuwdorp, M. Role of the gutmicrobiome in the pathogenesis of obesity and obesity-related met-abolic dysfunction. Gastroenterology 2017, 152, 1671–1678. [Google Scholar] [CrossRef]
- Martyniak, A.; Medyńska-Przęczek, A.; Wędrychowicz, A.; Skoczeń, S.; Tomasik, P.J. Prebiotics, Probiotics, Synbiotics, Paraprobiotics and Postbiotic Compounds in IBD. Biomolecules 2021, 11, 1903. [Google Scholar] [CrossRef] [PubMed]
- Banić, M.; Butorac, K.; Čuljak, N.; Leboš Pavunc, A.; Novak, J.; Bellich, B.; Kazazić, S.; Kazazić, S.; Cescutti, P.; Šušković, J.; et al. The Human Milk Microbiota Produces Potential Therapeutic Biomolecules and Shapes the Intestinal Microbiota of Infants. Int. J. Mol. Sci. 2022, 23, 14382. [Google Scholar] [CrossRef] [PubMed]
- Fuhren, J.; Rösch, C.; ten Napel, M.; Schols, H.A.; Kleerebezem, M. Synbiotic Matchmaking in Lactobacillus plantarum: Substrate Screening and Gene-Trait Matching to Characterize Strain-Specific Carbohydrate Utilization. Appl. Environ. Microb. 2020, 86, e01081-20. [Google Scholar] [CrossRef]
- Kumar, V.; Sheoran, P.; Gupta, A.; Yadav, J.; Kumar Tiwari, S. Antibacterial property of bacteriocin produced by Lactobacillus plantarum LD4 isolated from a fermented food. Ann. Microbiol. 2016, 66, 1431–1440. [Google Scholar] [CrossRef]
- van den Nieuwboer, M.; van Hemert, S.; Claassen, E.; de Vos, W.M. Lactobacillus plantarum WCFS1 and its host interaction: A dozen years after the genome. Microb. Biotechnol. 2016, 9, 452–465. [Google Scholar] [CrossRef]
- Dobson, A.; Cotter, P.D.; Ross, R.P.; Hill, C. Bacteriocin Production: A Probiotic Trait? Appl. Environ. Microbiol. 2021, 78, 1–6. [Google Scholar] [CrossRef]
- Trubenová, B.; Roizman, D.; Moter, A.; Rolff, J.; Regoes, R.R. Population genetics, biofilm recalcitrance, and antibiotic resistance evolution. Trends Microbiol. 2022, 30, 841–852. [Google Scholar] [CrossRef]
- Duraisamy, S.; Balakrishnan, S.; Ranjith, S.; Husain, F.; Sathyan, A.; Peter, A.S.; Prahalathan, C.; Kumarasamy, A. Bacteriocin—A potential antimicrobial peptide towards disrupting and preventing biofilm formation in the clinical and environmental locales. Environ. Sci. Pollut. Res. 2020, 27, 44922–44936. [Google Scholar] [CrossRef]
- Soltani, S.; Hammami, R.; Cotter, P.D.; Rebuffat, S.; Said, L.B.; Gaudreau, H.; Bédard, F.; Biron, E.; Drider, D.; Fliss, I. Bacteriocins as a new generation of antimicrobials: Toxicity aspects and regulations. FEMS Microbiol. Rev. 2021, 45, 039. [Google Scholar] [CrossRef] [PubMed]
- Uroić, K.; Novak, J.; Hynönen, U.; Pietilä, T.E.; Leboš Pavunc, A.; Kant, R.; Kos, B.; Palva, A.; Šušković, J. The role of S-layer in adhesive and immunomodulating properties of probiotic starter culture Lactobacillus brevis D6 isolated from artisanal smoked fresh cheese. LWT-Food Sci. Technol. 2016, 69, 623–632. [Google Scholar] [CrossRef]
- Martín, R.; Langella, P. Emerging Health Concepts in the Probiotics Field: Streamlining the Definitions. Front. Microbiol. 2019, 10, 1047. [Google Scholar] [CrossRef]
- Vinderola, G.; Sanders, M.E.; Salminen, S. The Concept of Postbiotics. Foods 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Banić, M.; Uroić, K.; Leboš Pavunc, A.; Novak, J.; Zorić, K.; Durgo, K.; Petković, H.; Jamnik, P.; Kazazić, S.; Kazazić, S.; et al. Characterization of S-layer proteins of potential probiotic starter culture Lactobacillus brevis SF9B isolated from sauerkraut. LWT-Food Sci. Technol. 2018, 93, 257–267. [Google Scholar] [CrossRef]
- Novak, J.; Butorac, K.; Leboš Pavunc, A.; Banić, M.; Butorac, A.; Lepur, A.; Oršolić, N.; Tonković, K.; Bendelja, K.; Čuljak, N.; et al. A lactic acid bacteria consortium impacted the content of casein-derived biopeptides in dried fresh cheese. Molecules 2022, 27, 160. [Google Scholar] [CrossRef]
- Butorac, K.; Banić, M.; Novak, J.; Leboš Pavunc, A.; Uroić, K.; Durgo, K.; Oršolić, N.; Kukolj, M.; Radović, S.; Scalabrin, S.; et al. The functional capacity of plantaricin-producing Lactobacillus plantarum SF9C and S-layer-carrying Lactobacillus brevis SF9B to withstand gastrointestinal transit. Microb. Cell Fact. 2020, 19, 106. [Google Scholar] [CrossRef]
- Liu, F.; Rajabi, S.; Shi, C.; Afifirad, G.; Omidi, N.; Kouhsari, E.; Azizian, K. Antibacterial activity of recently approved antibiotics against methicillin-resistant Staphylococcus aureus (MRSA) strains: A systematic review and meta-analysis. Ann. Clin. Microbiol. Antimicrob. 2022, 21, 37. [Google Scholar] [CrossRef]
- Wu, A.; Fu, Y.; Kong, L.; Shen, Q.; Liu, M.; Zeng, X.; Wu, Z.; Guo, Y.; Pan, D. Production of a class IIb bacteriocin with broad-spectrum antimicrobial activity in Lactiplantibacillus plantarum RUB1. Probiotics Antimicrob. Prot. 2021, 13, 1820–1832. [Google Scholar] [CrossRef]
- Zhao, D.; Wang, Q.; Lu, F.; Bie, X.; Zhao, H.; Lu, Z.; Lu, Y. A novel plantaricin 827 effectively inhibits Staphylococcus aureus and extends shelf life of skim milk. LWT-Food Sci. Technol. 2022, 154, 112849. [Google Scholar] [CrossRef]
- Choi, S.; Baek, M.-G.; Chung, M.-J.; Lim, S.; Yi, H. Distribution of bacteriocin genes in the lineages of Lactiplantibacillus plantarum. Sci. Rep. 2021, 11, 20063. [Google Scholar] [CrossRef] [PubMed]
- Gholamzadeh, M.; Gharajeh, N.H.; Hejazi, M.A. Genetic and in silico analysis of plantaricin EFI locus in indigenous isolates of lactobacillus plantarum. Biotechnol. Prog. 2019, 35, e2773. [Google Scholar] [CrossRef] [PubMed]
- Heeney, D.D.; Yarov-Yarovoy, V.; Marco, M.L. Sensitivity to the two peptide bacteriocin plantaricin EF is dependent on CorC, a membrane-bound, magnesium/cobalt efflux protein. Microbiologyopen 2019, 8, e827. [Google Scholar] [CrossRef] [PubMed]
- Tenea, G.N. Decoding the gene variants of two native probiotic Lactiplantibacillus plantarum strains through whole-genome resequencing: Insights into bacterial adaptability to stressors and antimicrobial strength. Genes 2022, 13, 443. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Mei, C.; Huang, X.; Gu, Q.; Song, D. Antibacterial activity and mechanism of a bacteriocin derived from the valine-cecropin A(1–8)-plantaricin ZJ5(1–18) hybrid peptide against Escherichia coli O104. Food Biophys. 2020, 15, 442–451. [Google Scholar] [CrossRef]
- Pu, J.; Hang, S.; Liu, M.; Chen, Z.; Xiong, J.; Li, Y.; Wu, H.; Zhao, X.; Liu, S.; Gu, Q.; et al. A class IIb bacteriocin plantaricin NC8 modulates gut microbiota of different enterotypes in vitro. Front. Nutr. 2022, 9, 877948. [Google Scholar] [CrossRef]
- de Moreno de LeBlanc, A.; Levit, R.; Savoy de Giori, G.; LeBlanc, J.G. Vitamin producing lactic acid bacteria as complementary treatments for intestinal infammation. Anti-Inflamm. Anti-Allergy Agents Med. Chem. 2018, 17, 50–56. [Google Scholar] [CrossRef]
- Zibin, Z.; Wentao, L.; Ying, R.; Xiaoqiong, L.; Shenjun, Z.; Hua, Y.; Yingping, Y. Allobaculum involves in the modulation of intestinal ANGPTLT4 expression in mice treated by high-fat diet. Front. Nutr. 2021, 8, 690138. [Google Scholar]
- Li, W.; Zhang, K.; Yang, H. Pectin alleviates high fat (lard) diet-induced nonalcoholic fatty liver disease in mice: Possible role of short-chain fatty acids and gut microbiota regulated by pectin. J. Agric. Food Chem. 2018, 66, 8015–8025. [Google Scholar] [CrossRef]
- Gryaznova, M.; Dvoretskaya, Y.; Burakova, I.; Syromyatnikov, M.; Popov, E.; Kokina, A.; Mikhaylov, E.; Popov, V. Dynamics of changes in the gut microbiota of healthy mice fed with lactic acid bacteria and Bifidobacteria. Microorganisms 2022, 10, 1020. [Google Scholar] [CrossRef]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Bazargani, M.M.; Rohloff, J. Antibiofilm activity of essential oils and plant extracts against Staphylococcus aureus and Escherichia coli biofilms. Food Control 2016, 61, 156–164. [Google Scholar] [CrossRef]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Dis, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The seed and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 2014, 42, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Jumper, J.; Evans, R.; Pritzel, A.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Varadi, M.; Anyango, S.; Deshpande, M.; Nair, S.; Natassia, C.; Yordanova, G.; Yuan, D.; Stroe, O.; Wood, G.; Laydon, A.; et al. AlphaFold Protein Structure Database: Massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res. 2022, 50, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Gautier, R.; Douguet, D.; Antonny, B.; Drin, G. HELIQUEST: A web server to screen sequences with specific a-helical properties. Bioinformatics 2008, 24, 2101–2102. [Google Scholar] [CrossRef]
- Lan, D.; Niu, J.; Miao, J.; Dong, X.; Wang, H.; Yang, G.; Wang, K.; Miao, Y. Expression of guanylate cyclase-C, guanylin and uroguanylin is downregulated proportionally to the ulcerative colitis disease activity index. Sci. Rep. 2016, 6, 25034. [Google Scholar] [CrossRef]
- Koelink, P.J.; Wildenberg, M.E.; Stitt, L.W.; Feagan, B.G.; Koldijk, M.; van‘t Wout, A.B.; Atreya, R.; Vieth, M.; Brandse, J.F.; Duijst, S.; et al. Development of reliable, valid and responsive scoring systems for endoscopy and histology in animal models for inflammatory bowel disease. J. Crohn’s Colitis 2018, 12, 794–803. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Cultivation Conditions | Source |
|---|---|---|
| Lb. plantarum D4 | MRS, 37 °C, microaerophilic | Smoked fresh cheese |
| Lb. plantarum D5 | MRS, 37 °C, microaerophilic | Smoked fresh cheese |
| Lb. plantarum D7 | MRS, 37 °C, microaerophilic | Smoked fresh cheese |
| Lb. plantarum D13 | MRS, 37 °C, microaerophilic | Smoked fresh cheese |
| Lb. plantarum M4 | MRS, 37 °C, microaerophilic | Dried fresh cheese |
| Lb. plantarum M5 | MRS, 37 °C, microaerophilic | Dried fresh cheese |
| Lb. plantarum MA2 | MRS, 37 °C, microaerophilic | Dried fresh cheese |
| Lb. plantarum MA3 | MRS, 37 °C, microaerophilic | Dried fresh cheese |
| Lb. plantarum SF15C | MRS, 37 °C, microaerophilic | Sauerkraut |
| Lb. plantarum ZG1C | MRS, 37 °C, microaerophilic | Sauerkraut |
| Lb. plantarum M92C | MRS, 37 °C, microaerophilic | Fermented milk |
| Lb. plantarum L4 | MRS, 37 °C, microaerophilic | Silage |
| S. aureus ATCC® 25923™ | BHI, 37 °C, aerobic | ATCC 1 |
| L. monocytogenes ATCC® 19111™ | BHI, 37 °C, aerobic | ATCC 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Butorac, K.; Novak, J.; Banić, M.; Leboš Pavunc, A.; Čuljak, N.; Oršolić, N.; Odeh, D.; Perica, J.; Šušković, J.; Kos, B. Modulation of the Gut Microbiota by the Plantaricin-Producing Lactiplantibacillus plantarum D13, Analysed in the DSS-Induced Colitis Mouse Model. Int. J. Mol. Sci. 2023, 24, 15322. https://doi.org/10.3390/ijms242015322
Butorac K, Novak J, Banić M, Leboš Pavunc A, Čuljak N, Oršolić N, Odeh D, Perica J, Šušković J, Kos B. Modulation of the Gut Microbiota by the Plantaricin-Producing Lactiplantibacillus plantarum D13, Analysed in the DSS-Induced Colitis Mouse Model. International Journal of Molecular Sciences. 2023; 24(20):15322. https://doi.org/10.3390/ijms242015322
Chicago/Turabian StyleButorac, Katarina, Jasna Novak, Martina Banić, Andreja Leboš Pavunc, Nina Čuljak, Nada Oršolić, Dyana Odeh, Jana Perica, Jagoda Šušković, and Blaženka Kos. 2023. "Modulation of the Gut Microbiota by the Plantaricin-Producing Lactiplantibacillus plantarum D13, Analysed in the DSS-Induced Colitis Mouse Model" International Journal of Molecular Sciences 24, no. 20: 15322. https://doi.org/10.3390/ijms242015322