
Multi-Omic Temporal Landscape of Plasma and Synovial Fluid-Derived Extracellular Vesicles Using an Experimental Model of Equine Osteoarthritis
 , , ,
, , ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Model Outcome
2.2. EV Characteristics
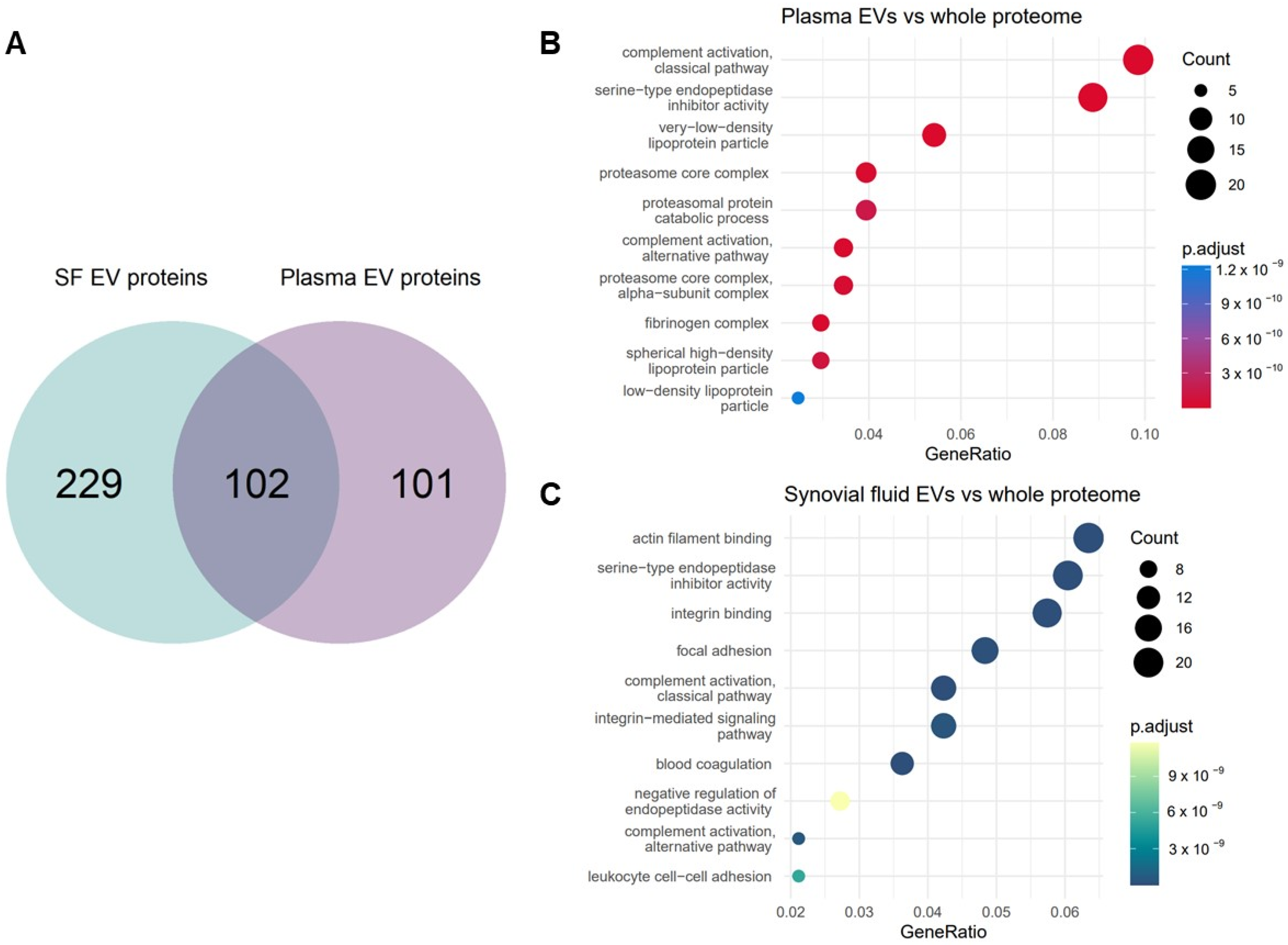
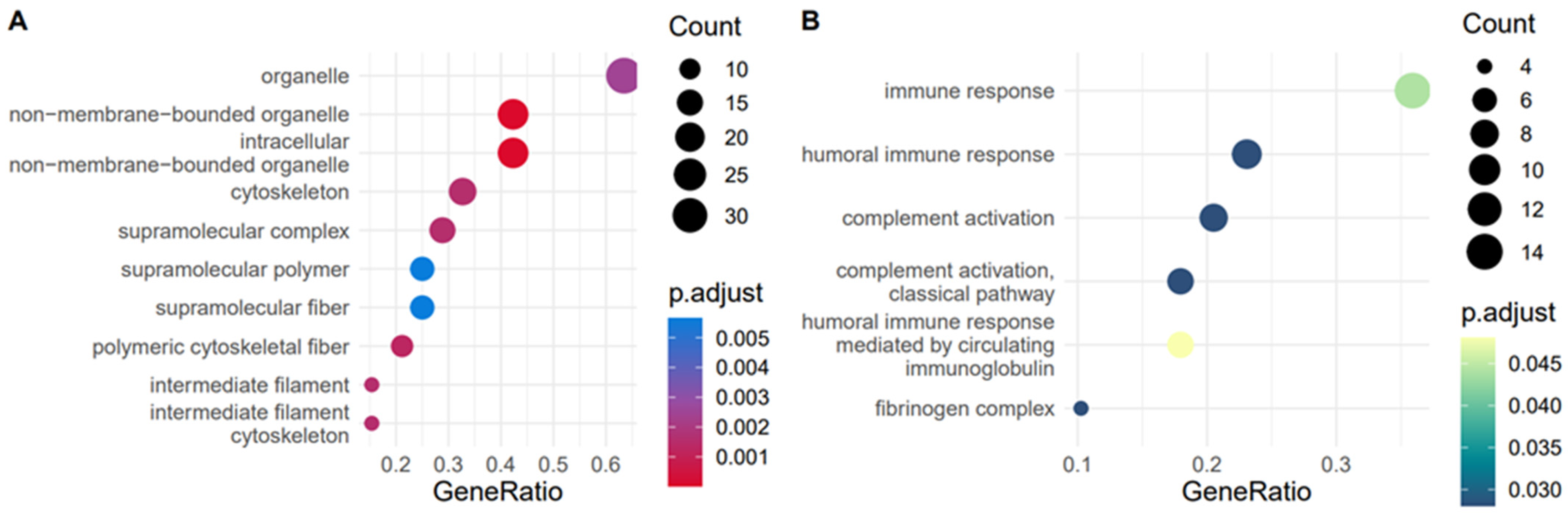
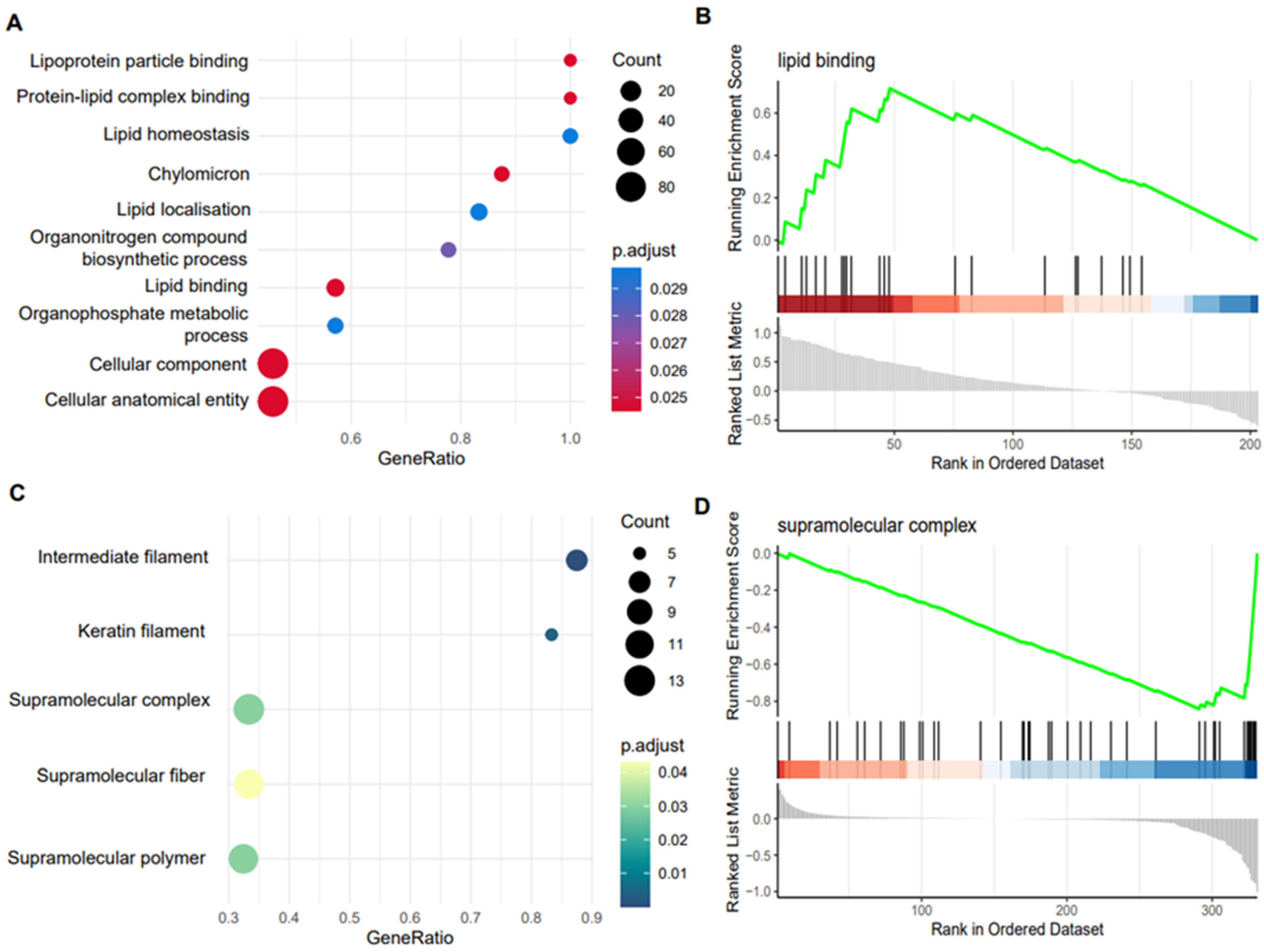
2.3. Characterisation of the Equine Plasma and Synovial Fluid EV Proteome
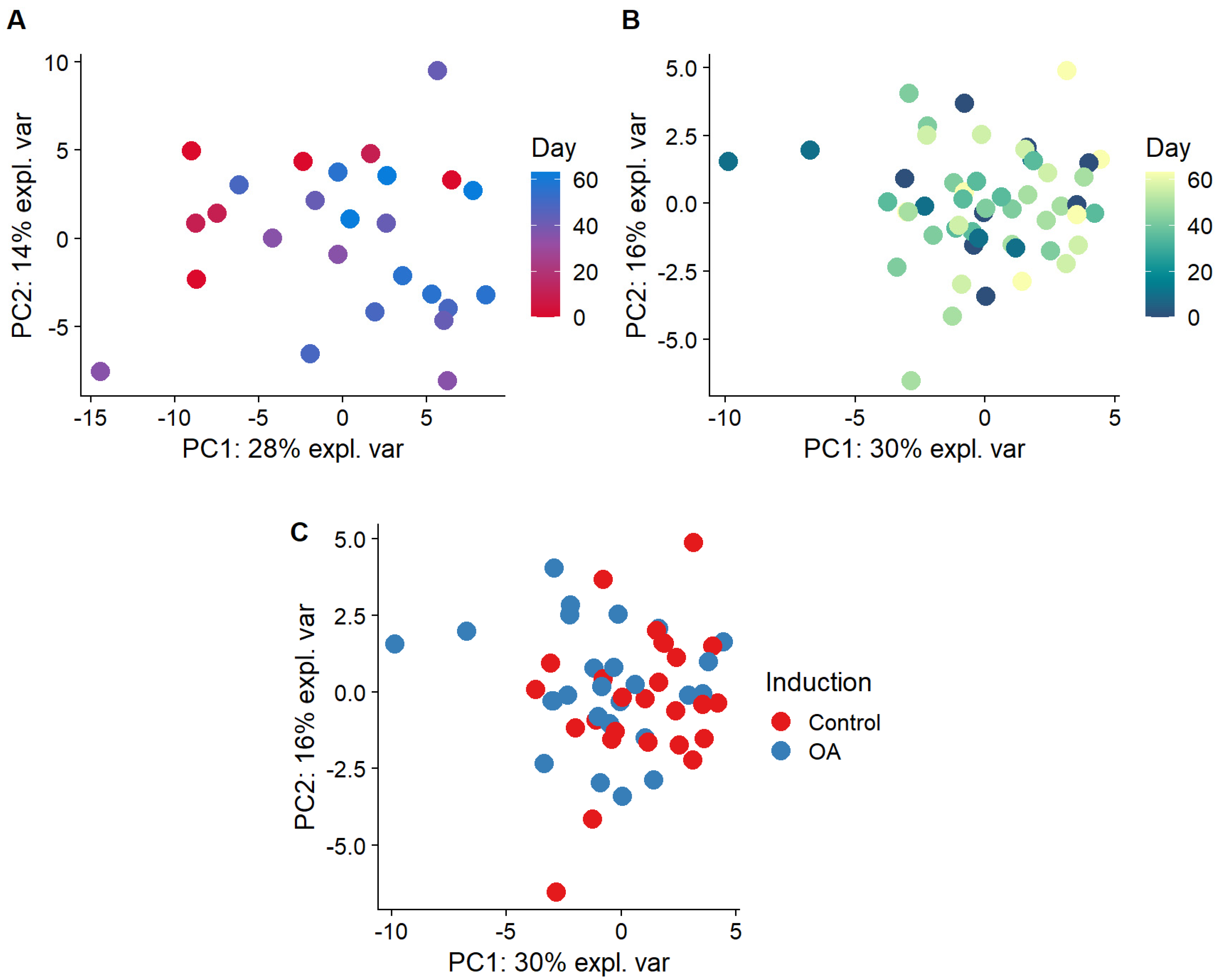
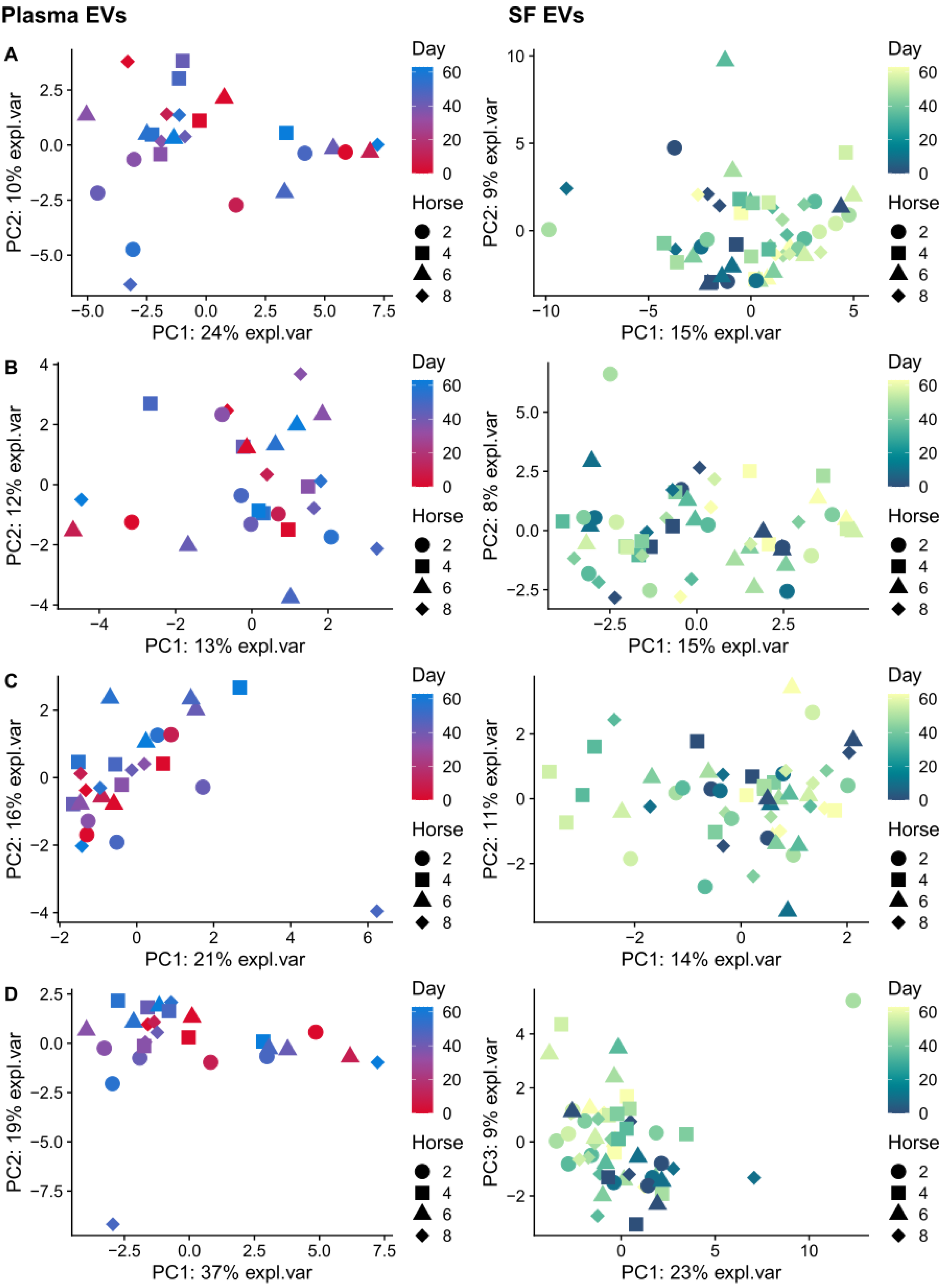
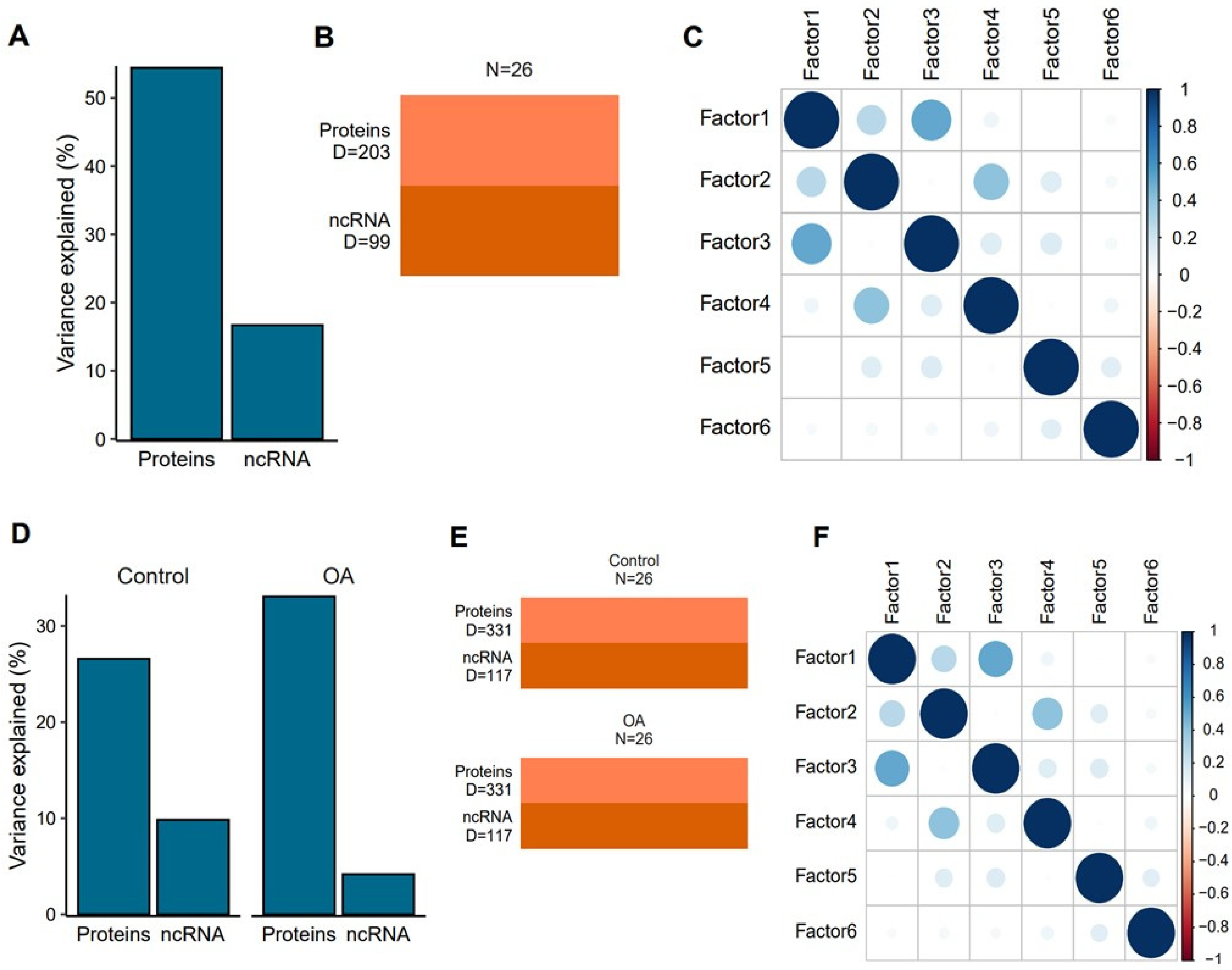
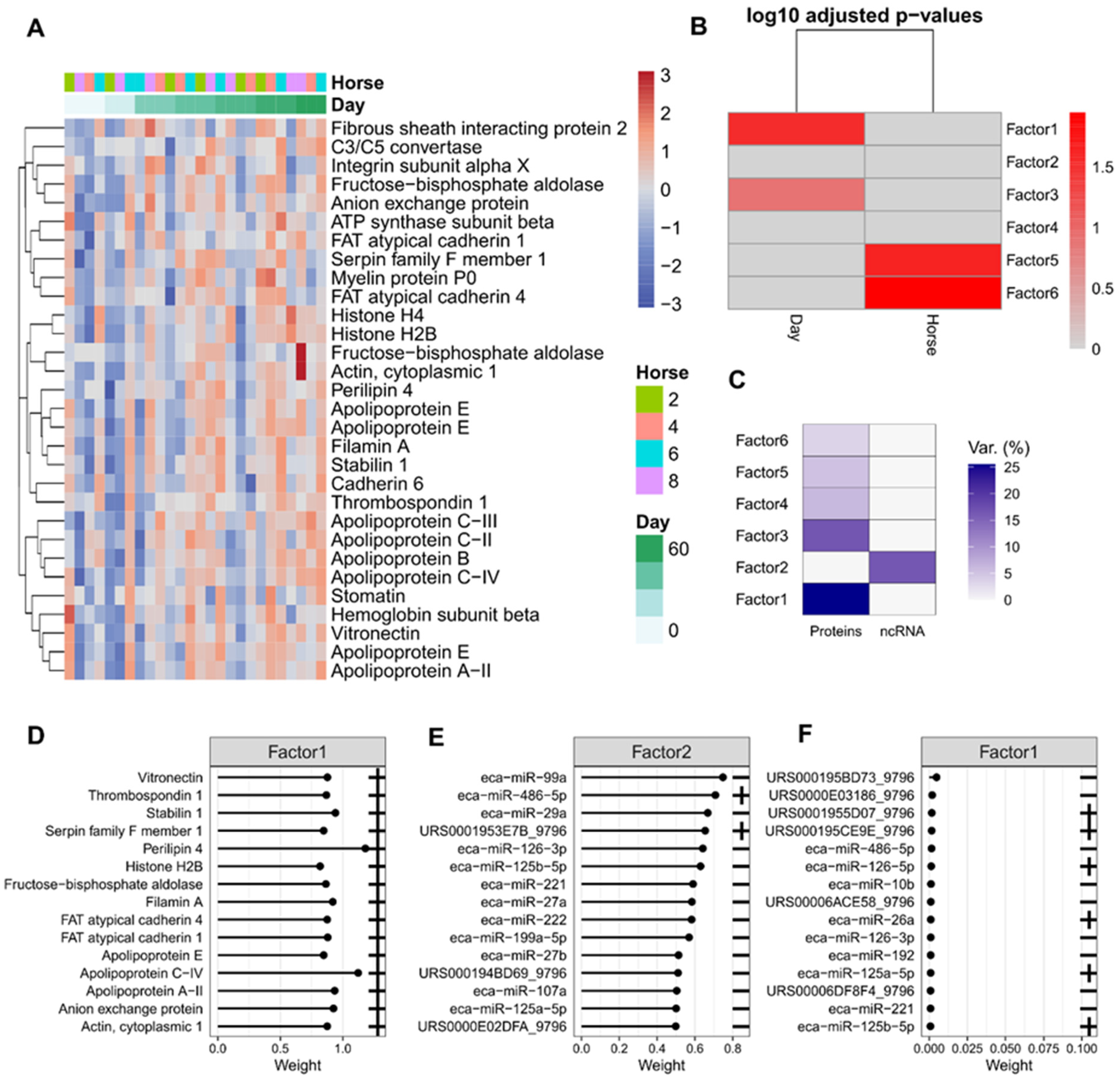
2.4. A Multivariate Approach Identified a Separation by Day for Proteins from Plasma-Derived EVs
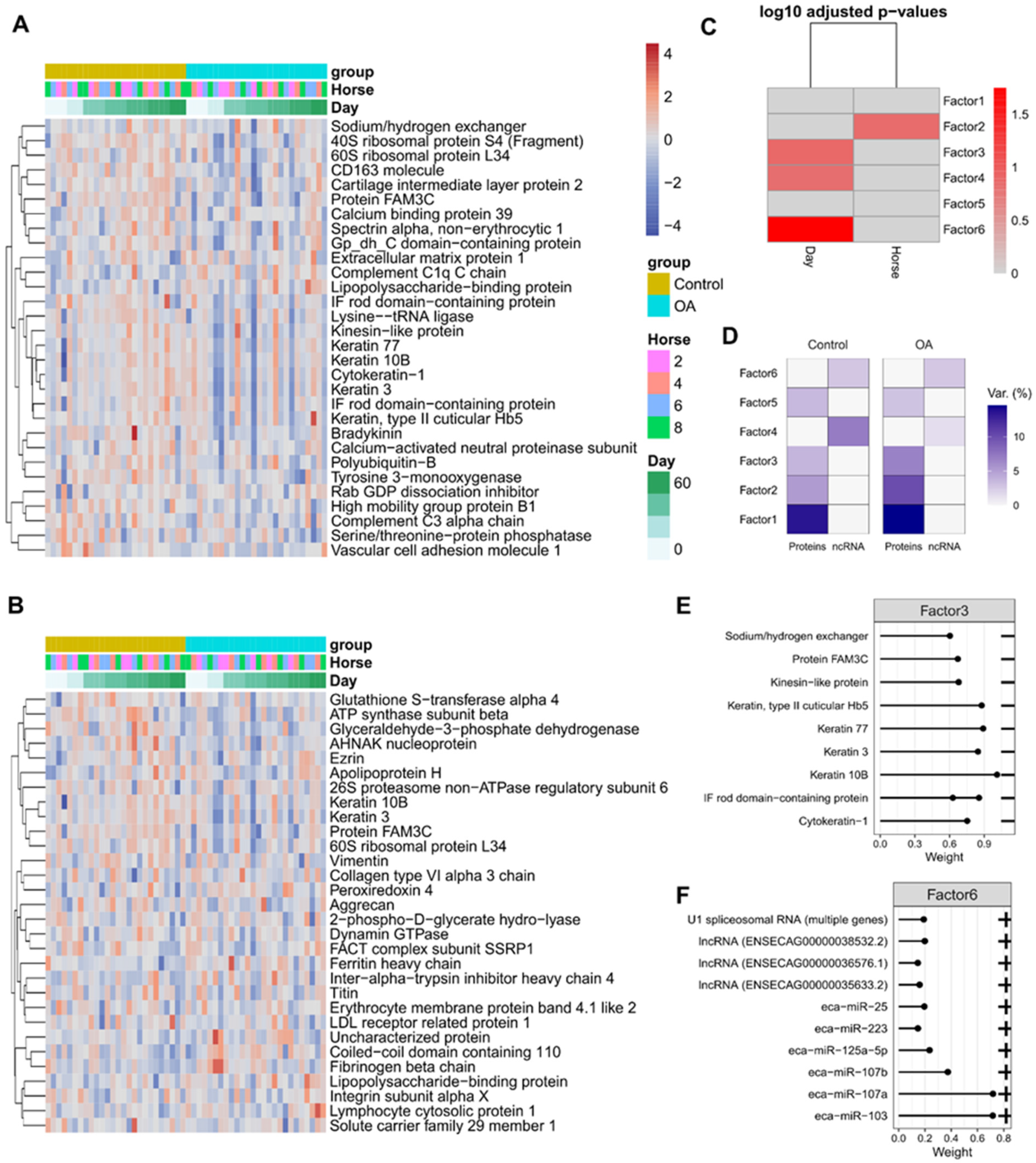
2.5. Differential Expression of Plasma and Synovial Fluid-Derived EVs
2.6. Small RNA Sequencing Analysis Quality Control Prior to Multi-Omics Analysis
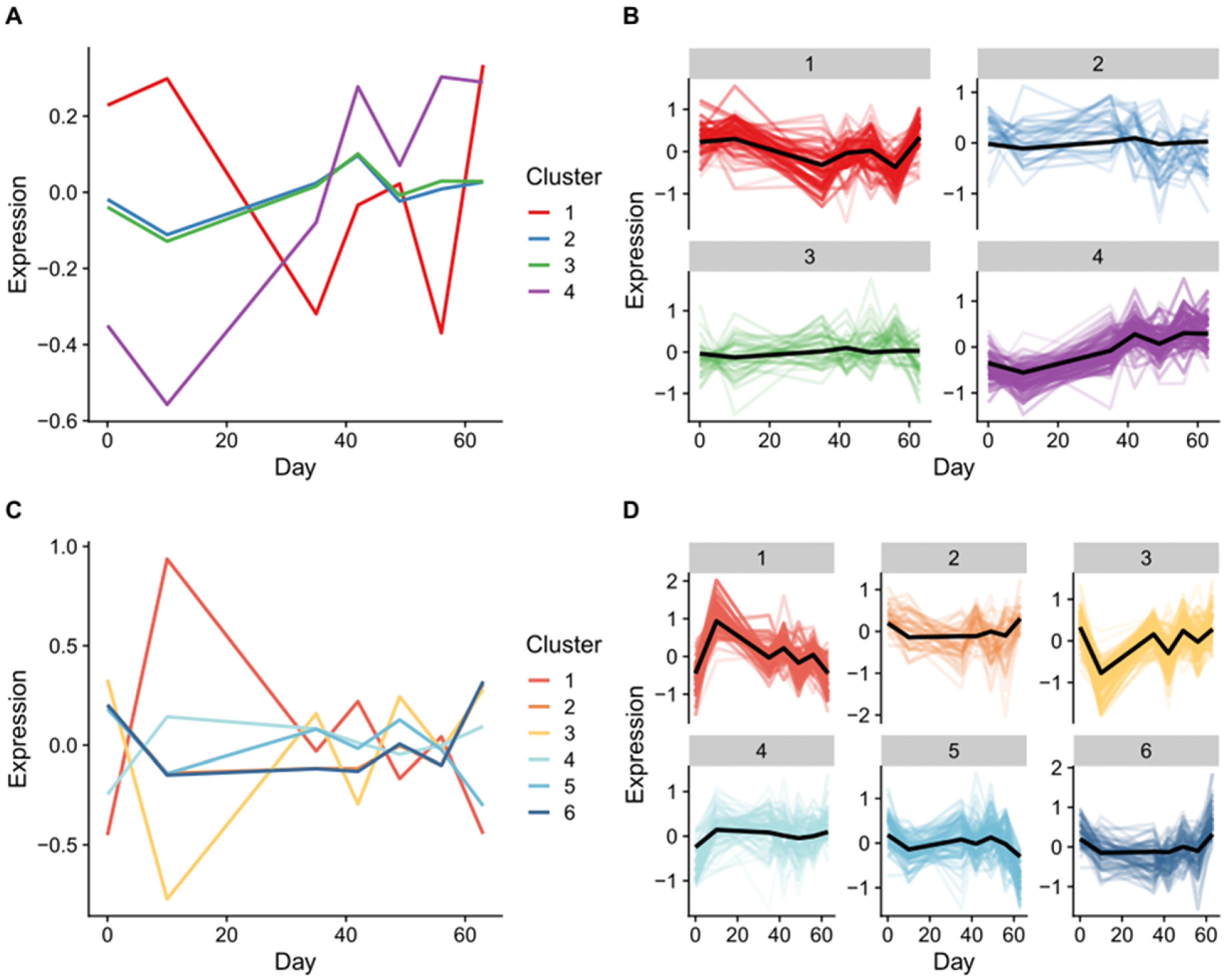
2.7. Multi-Omics Time Series Reveals Patterns of Expression during OA Progression
2.8. Multi-Omics Factor Analysis Demonstrates That Proteins Drive the Bulk of EV Cargo Variation during OA Progression
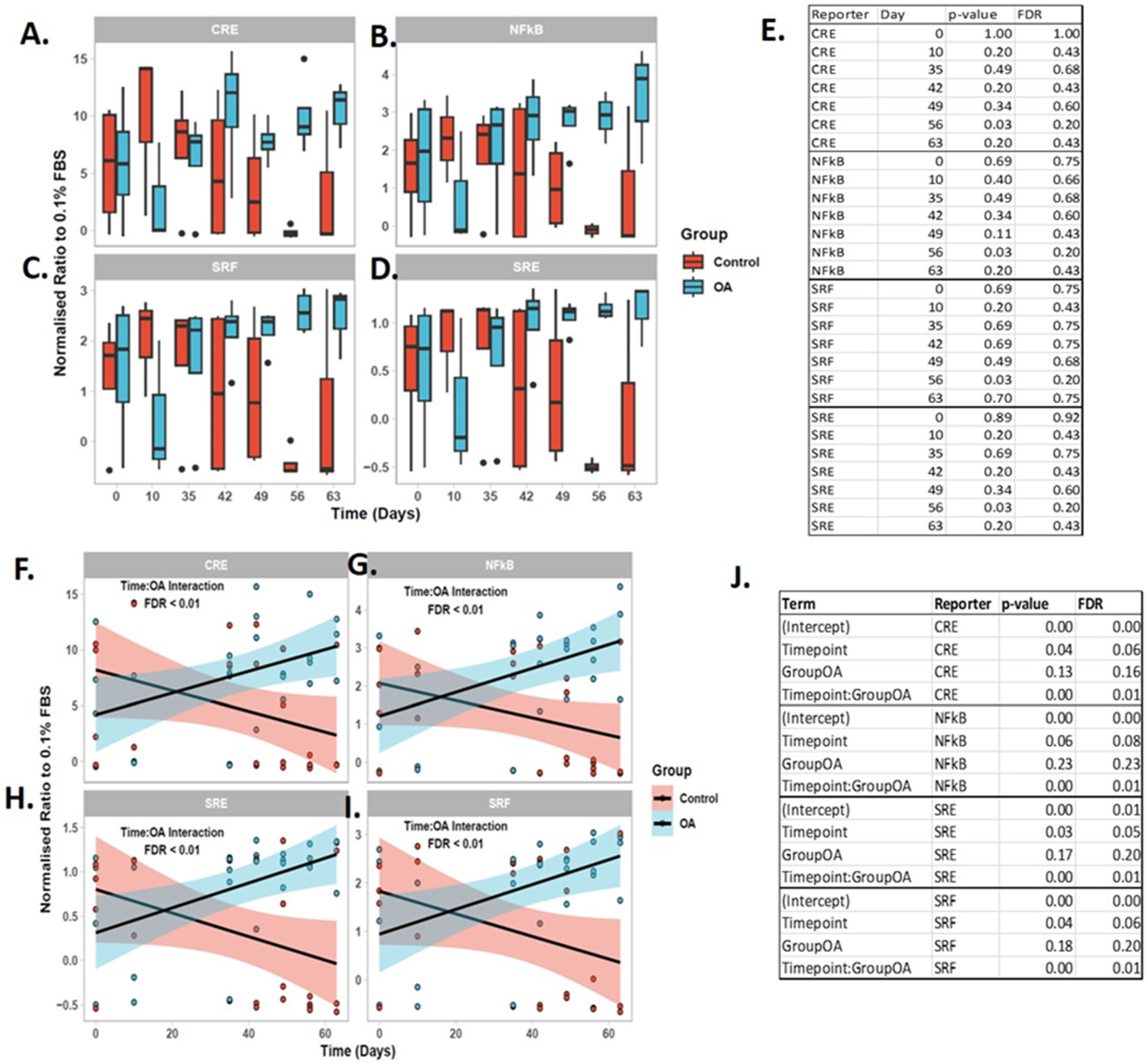
2.9. Synovial Fluid-Derived EVs Induced Intracellular Chondrocyte Signaling
3. Discussion
4. Materials and Methods
4.1. Horses and Study Design, Induction of Osteoarthritis
4.2. Synovial Fluid and Plasma Sampling
4.3. Post-Mortem Examination
4.4. EV Isolation
4.5. EV Characterisation
4.6. Plasma and Synovial Fluid-Derived EV Spectral Library Preparation for Proteomics
4.7. Plasma and Synovial Fluid-Derived EV Sample Preparation for SWATH-MS
4.8. Small RNA Sequencing of EVs
4.9. Reporter Gene Assay
4.10. Statistical Analysis
4.10.1. Single-Modality Approaches
4.10.2. Statistical Analysis of Reporter Data
4.10.3. Multi-Omics Time Series Clustering
4.10.4. Multi-Omics Factor Analysis
4.10.5. Functional Enrichment
4.10.6. Visualisation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Todhunter, P.G.; Kincaid, S.A.; Todhunter, R.J.; Kammermann, J.R.; Johnstone, B.; Baird, A.N.; Hanson, R.R.; Wright, J.M.; Lin, H.C.; Purohit, R.C. Immunohistochemical analysis of an equine model of synovitis-induced arthritis. Am. J. Vet. Res. 1996, 57, 1080–1093. [Google Scholar]
- Ireland, J.L.; Clegg, P.D.; McGowan, C.M.; McKane, S.A.; Chandler, K.J.; Pinchbeck, G.L. Comparison of owner-reported health problems with veterinary assessment of geriatric horses in the United Kingdom. Equine Vet. J. 2012, 44, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Little, C.B.; Ghosh, P.; Rose, R. The effect of strenuous versus moderate exercise on the metabolism of proteoglycans in articular cartilage from different weight-bearing regions of the equine third carpal bone. Osteoarthr. Cartil. 1997, 5, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Lopez, E.; Coras, R.; Torres, A.; Lane, N.E.; Guma, M. Synovial inflammation in osteoarthritis progression. Nat. Rev. Rheumatol. 2022, 18, 258–275. [Google Scholar] [CrossRef] [PubMed]
- Loeser, R.F. Molecular mechanisms of cartilage destruction: Mechanics, inflammatory mediators, and aging collide. Arthritis Rheum. 2006, 54, 1357–1360. [Google Scholar] [CrossRef]
- Junker, S.; Krumbholz, G.; Frommer, K.W.; Rehart, S.; Steinmeyer, J.; Rickert, M.; Schett, G.; Muller-Ladner, U.; Neumann, E. Differentiation of osteophyte types in osteoarthritis—Proposal of a histological classification. Jt. Bone Spine 2016, 83, 63–67. [Google Scholar] [CrossRef]
- Donell, S. Subchondral bone remodelling in osteoarthritis. EFORT Open Rev. 2019, 4, 221–229. [Google Scholar] [CrossRef]
- Caron, J.P. Understanding the pathogenesis of equine osteoarthritis. Br. Vet. J. 1992, 148, 369–371. [Google Scholar] [CrossRef]
- Goldring, M.B. Update on the biology of the chondrocyte and new approaches to treating cartilage diseases. Best. Pract. Res. Clin. Rheumatol. 2006, 20, 1003–1025. [Google Scholar] [CrossRef]
- Kaur, S.; Abu-Shahba, A.G.; Paananen, R.O.; Hongisto, H.; Hiidenmaa, H.; Skottman, H.; Seppanen-Kaijansinkko, R.; Mannerstrom, B. Small non-coding RNA landscape of extracellular vesicles from human stem cells. Sci. Rep. 2018, 8, 15503. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Zhou, S.; Li, S.; Kuang, L.; Chen, H.; Luo, X.; Ouyang, J.; He, M.; Du, X.; Chen, L. Exosomes: Roles and therapeutic potential in osteoarthritis. Bone Res. 2020, 8, 25. [Google Scholar] [CrossRef]
- Withrow, J.; Murphy, C.; Liu, Y.; Hunter, M.; Fulzele, S.; Hamrick, M.W. Extracellular vesicles in the pathogenesis of rheumatoid arthritis and osteoarthritis. Arthritis Res. Ther. 2016, 18, 286. [Google Scholar] [CrossRef] [PubMed]
- Ragni, E.; Perucca Orfei, C.; De Luca, P.; Lugano, G.; Vigano, M.; Colombini, A.; Valli, F.; Zacchetti, D.; Bollati, V.; de Girolamo, L. Interaction with hyaluronan matrix and miRNA cargo as contributors for in vitro potential of mesenchymal stem cell-derived extracellular vesicles in a model of human osteoarthritic synoviocytes. Stem Cell Res. Ther. 2019, 10, 109. [Google Scholar] [CrossRef] [PubMed]
- Clarke, E.J.; Johnson, E.; Caamano Gutierrez, E.; Andersen, C.; Berg, L.C.; Jenkins, R.E.; Lindegaard, C.; Uvebrant, K.; Lundgren-Akerlund, E.; Turlo, A.; et al. Temporal extracellular vesicle protein changes following intraarticular treatment with integrin α10β1-selected mesenchymal stem cells in equine osteoarthritis. Front. Vet. Sci. 2022, 9, 1057667. [Google Scholar] [CrossRef]
- Cai, J.; Wu, J.; Wang, J.; Li, Y.; Hu, X.; Luo, S.; Xiang, D. Extracellular vesicles derived from different sources of mesenchymal stem cells: Therapeutic effects and translational potential. Cell Biosci. 2020, 10, 69. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, Y.; Xiao, K.; Xiang, S.; Li, Z.; Weng, X. Emerging Role of Exosomes in the Joint Diseases. Cell. Physiol. Biochem. 2018, 47, 2008–2017. [Google Scholar] [CrossRef]
- Zhao, Y.; Xu, J. Synovial fluid-derived exosomal lncRNA PCGEM1 as biomarker for the different stages of osteoarthritis. Int. Orthop. 2018, 42, 2865–2872. [Google Scholar] [CrossRef]
- Anderson, J.R.; Jacobsen, S.; Walters, M.; Bundgaard, L.; Diendorfer, A.; Hackl, M.; Clarke, E.J.; James, V.; Peffers, M.J. Small non-coding RNA landscape of extracellular vesicles from a post-traumatic model of equine osteoarthritis. Front. Vet. Sci. 2022, 9, 901269. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, S.N.; Anderson, J.; Brown, T.J.; Nanus, D.E.; Housmans, B.; Green, J.A.; Hackl, M.; Choi, K.K.; Arkill, K.P.; Welting, T.; et al. The role of extracellular vesicle miRNAs and tRNAs in synovial fibroblast senescence. Front. Mol. Biosci. 2022, 9, 971621. [Google Scholar] [CrossRef]
- Foers, A.D.; Garnham, A.L.; Chatfield, S.; Smyth, G.K.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Extracellular Vesicles in Synovial Fluid from Rheumatoid Arthritis Patients Contain miRNAs with Capacity to Modulate Inflammation. Int. J. Mol. Sci. 2021, 22, 4910. [Google Scholar] [CrossRef]
- Clarke, E.J.; Lima, C.; Anderson, J.R.; Castanheira, C.; Beckett, A.; James, V.; Hyett, J.; Goodacre, R.; Peffers, M.J. Optical photothermal infrared spectroscopy can differentiate equine osteoarthritic plasma extracellular vesicles from healthy controls. Anal. Methods 2022, 14, 3661–3670. [Google Scholar] [CrossRef] [PubMed]
- Foers, A.D.; Dagley, L.F.; Chatfield, S.; Webb, A.I.; Cheng, L.; Hill, A.F.; Wicks, I.P.; Pang, K.C. Proteomic analysis of extracellular vesicles reveals an immunogenic cargo in rheumatoid arthritis synovial fluid. Clin. Transl. Immunol. 2020, 9, e1185. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Liu, Y.; Huang, Q.; Sun, S.; Ji, Z.; Huang, L.; Li, Z.; Huang, X.; Deng, W.; Li, T. TMT-Based Quantitative Proteomics Analysis of Synovial Fluid-Derived Exosomes in Inflammatory Arthritis. Front. Immunol. 2022, 13, 800902. [Google Scholar] [CrossRef] [PubMed]
- Ripmeester, E.G.J.; Caron, M.M.J.; van den Akker, G.G.H.; Surtel, D.A.M.; Cremers, A.; Balaskas, P.; Dyer, P.; Housmans, B.A.C.; Chabronova, A.; Smagul, A.; et al. Impaired chondrocyte U3 snoRNA expression in osteoarthritis impacts the chondrocyte protein translation apparatus. Sci. Rep. 2020, 10, 13426. [Google Scholar] [CrossRef]
- Peffers, M.J.; Chabronova, A.; Balaskas, P.; Fang, Y.; Dyer, P.; Cremers, A.; Emans, P.J.; Feczko, P.Z.; Caron, M.M.; Welting, T.J.M. SnoRNA signatures in cartilage ageing and osteoarthritis. Sci. Rep. 2020, 10, 10641. [Google Scholar] [CrossRef]
- Chabronova, A.; van den Akker, G.G.H.; Housmans, B.A.C.; Caron, M.M.J.; Cremers, A.; Surtel, D.A.M.; Wichapong, K.; Peffers, M.M.J.; van Rhijn, L.W.; Marchand, V.; et al. Ribosomal RNA-based epitranscriptomic regulation of chondrocyte translation and proteome in osteoarthritis. Osteoarthr. Cartil. 2023, 31, 374–385. [Google Scholar] [CrossRef]
- Baker, M.E.; Lee, S.; Clinton, M.; Hackl, M.; Castanheira, C.; Peffers, M.J.; Taylor, S.E. Investigation of MicroRNA Biomarkers in Equine Distal Interphalangeal Joint Osteoarthritis. Int. J. Mol. Sci. 2022, 23, 15526. [Google Scholar] [CrossRef]
- Castanheira, C.; Anderson, J.R.; Fang, Y.; Milner, P.I.; Goljanek-Whysall, K.; House, L.; Clegg, P.D.; Peffers, M.J. Mouse microRNA signatures in joint ageing and post-traumatic osteoarthritis. Osteoarthr. Cart. Cartil. Open 2021, 3, 100186. [Google Scholar] [CrossRef]
- Castanheira, C.; Balaskas, P.; Falls, C.; Ashraf-Kharaz, Y.; Clegg, P.; Burke, K.; Fang, Y.; Dyer, P.; Welting, T.J.M.; Peffers, M.J. Equine synovial fluid small non-coding RNA signatures in early osteoarthritis. BMC Vet. Res. 2021, 17, 26. [Google Scholar] [CrossRef]
- Hulme, C.H.; Peffers, M.J.; Harrington, G.M.B.; Wilson, E.; Perry, J.; Roberts, S.; Gallacher, P.; Jermin, P.; Wright, K.T. Identification of Candidate Synovial Fluid Biomarkers for the Prediction of Patient Outcome After Microfracture or Osteotomy. Am. J. Sports Med. 2021, 49, 1512–1523. [Google Scholar] [CrossRef]
- Hulme, C.H.; Wilson, E.L.; Peffers, M.J.; Roberts, S.; Simpson, D.M.; Richardson, J.B.; Gallacher, P.; Wright, K.T. Autologous chondrocyte implantation-derived synovial fluids display distinct responder and non-responder proteomic profiles. Arthritis Res. Ther. 2017, 19, 150. [Google Scholar] [CrossRef]
- Peffers, M.J.; Cillero-Pastor, B.; Eijkel, G.B.; Clegg, P.D.; Heeren, R.M. Matrix assisted laser desorption ionization mass spectrometry imaging identifies markers of ageing and osteoarthritic cartilage. Arthritis Res. Ther. 2014, 16, R110. [Google Scholar] [CrossRef]
- Peffers, M.J.; McDermott, B.; Clegg, P.D.; Riggs, C.M. Comprehensive protein profiling of synovial fluid in osteoarthritis following protein equalization. Osteoarthr. Cartil. 2015, 23, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Timur, U.T.; Jahr, H.; Anderson, J.; Green, D.C.; Emans, P.J.; Smagul, A.; van Rhijn, L.W.; Peffers, M.J.; Welting, T.J.M. Identification of tissue-dependent proteins in knee OA synovial fluid. Osteoarthr. Cartil. 2021, 29, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Anderson, A.R.; Phelan, M.M.; Caamaño-Gutiérrez, E.; Clegg, P.D.; Rubio-Martinez, L.K.; Peffers, M.J. Metabolomic and Proteomic Stratification of Equine Osteoarthritis. BioRxiv 2020. [Google Scholar] [CrossRef]
- Walters, M.; Skovgaard, K.; Heegaard, P.; Peffers, M.; Fang, Y.; Bundgaard, L.; Skovgaard, L.; Jacobsen, S. Changes in small non-coding RNA expression in synovial fluid during disease progression in an equine model of experimental osteoarthritis. Osteoarthr. Cartil. 2021, 29, S155–S156. [Google Scholar] [CrossRef]
- Jin, L.; Bi, Y.; Hu, C.; Qu, J.; Shen, S.; Wang, X.; Tian, Y. A comparative study of evaluating missing value imputation methods in label-free proteomics. Sci. Rep. 2021, 11, 1760. [Google Scholar] [CrossRef]
- Gardner, M.L.; Freitas, M.A. Multiple Imputation Approaches Applied to the Missing Value Problem in Bottom-Up Proteomics. Int. J. Mol. Sci. 2021, 22, 9650. [Google Scholar] [CrossRef]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef]
- Burgelman, M.; Dujardin, P.; Vandendriessche, C.; Vandenbroucke, R.E. Free complement and complement containing extracellular vesicles as potential biomarkers for neuroinflammatory and neurodegenerative disorders. Front. Immunol. 2022, 13, 1055050. [Google Scholar] [CrossRef]
- Park, D.J.; Duggan, E.; Ho, K.; Dorschner, R.A.; Dobke, M.; Nolan, J.P.; Eliceiri, B.P. Serpin-loaded extracellular vesicles promote tissue repair in a mouse model of impaired wound healing. J. Nanobiotechnology 2022, 20, 474. [Google Scholar] [CrossRef]
- Karasu, E.; Eisenhardt, S.U.; Harant, J.; Huber-Lang, M. Extracellular Vesicles: Packages Sent With Complement. Front. Immunol. 2018, 9, 721. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Chen, A.; Su, Y.; You, M.; Guo, H.; Tan, S.; He, Q.; Hu, B. Extracellular vesicles: A new communication paradigm of complement in neurological diseases. Brain Res. Bull. 2023, 199, 110667. [Google Scholar] [CrossRef] [PubMed]
- Ahlmann-Eltze, C.; Anders, S. ProDA: Probabilistic Dropout Analysis for Identifying Differentially Abundant Proteins in Label-Free Mass Spectrometry. BioRxiv 2019, 180, 968–983.e24. [Google Scholar] [CrossRef]
- Argelaguet, R.; Arnol, D.; Bredikhin, D.; Deloro, Y.; Velten, B.; Marioni, J.C.; Stegle, O. MOFA+: A statistical framework for comprehensive integration of multi-modal single-cell data. Genome Biol. 2020, 21, 111. [Google Scholar] [CrossRef] [PubMed]
- Housmans, B.A.C.; van den Akker, G.G.H.; Neefjes, M.; Timur, U.T.; Cremers, A.; Peffers, M.J.; Caron, M.M.J.; van Rhijn, L.W.; Emans, P.J.; Boymans, T.; et al. Direct comparison of non-osteoarthritic and osteoarthritic synovial fluid-induced intracellular chondrocyte signaling and phenotype changes. Osteoarthr. Cartil. 2023, 31, 60–71. [Google Scholar] [CrossRef]
- Raggi, F.; Bartolucci, M.; Cangelosi, D.; Rossi, C.; Pelassa, S.; Trincianti, C.; Petretto, A.; Filocamo, G.; Civino, A.; Eva, A.; et al. Proteomic profiling of extracellular vesicles in synovial fluid and plasma from Oligoarticular Juvenile Idiopathic Arthritis patients reveals novel immunopathogenic biomarkers. Front. Immunol. 2023, 14, 1134747. [Google Scholar] [CrossRef]
- Reymond, S.; Gruaz, L.; Sanchez, J.C. Depletion of abundant plasma proteins for extracellular vesicle proteome characterization: Benefits and pitfalls. Anal. Bioanal. Chem. 2023, 415, 3177–3187. [Google Scholar] [CrossRef]
- Cao, X.; Sandberg, A.; Araujo, J.E.; Cvetkovski, F.; Berglund, E.; Eriksson, L.E.; Pernemalm, M. Evaluation of Spin Columns for Human Plasma Depletion to Facilitate MS-Based Proteomics Analysis of Plasma. J. Proteome Res. 2021, 20, 4610–4620. [Google Scholar] [CrossRef]
- Zhang, X.; Huebner, J.L.; Kraus, V.B. Extracellular Vesicles as Biological Indicators and Potential Sources of Autologous Therapeutics in Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 8351. [Google Scholar] [CrossRef]
- Mustonen, A.M.; Lehmonen, N.; Paakkonen, T.; Raekallio, M.; Kakela, R.; Niemela, T.; Mykkanen, A.; Sihvo, S.P.; Nieminen, P. Equine osteoarthritis modifies fatty acid signatures in synovial fluid and its extracellular vesicles. Arthritis Res. Ther. 2023, 25, 39. [Google Scholar] [CrossRef] [PubMed]
- Mustonen, A.M.; Lehmonen, N.; Oikari, S.; Capra, J.; Raekallio, M.; Mykkanen, A.; Paakkonen, T.; Rilla, K.; Niemela, T.; Nieminen, P. Counts of hyaluronic acid-containing extracellular vesicles decrease in naturally occurring equine osteoarthritis. Sci. Rep. 2022, 12, 17550. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Mokuda, S.; Tokunaga, T.; Kohno, H.; Ishitoku, M.; Araki, K.; Sugimoto, T.; Yoshida, Y.; Yamamoto, T.; Matsumoto, M.; et al. Expression of factor XIII originating from synovial fibroblasts and macrophages induced by interleukin-6 signaling. Inflamm. Regen. 2023, 43, 2. [Google Scholar] [CrossRef] [PubMed]
- Raghu, H.; Cruz, C.; Rewerts, C.L.; Frederick, M.D.; Thornton, S.; Mullins, E.S.; Schoenecker, J.G.; Degen, J.L.; Flick, M.J. Transglutaminase factor XIII promotes arthritis through mechanisms linked to inflammation and bone erosion. Blood 2015, 125, 427–437. [Google Scholar] [CrossRef]
- Kolhe, R.; Owens, V.; Sharma, A.; Lee, T.J.; Zhi, W.; Ghilzai, U.; Mondal, A.K.; Liu, Y.; Isales, C.M.; Hamrick, M.W.; et al. Sex-Specific Differences in Extracellular Vesicle Protein Cargo in Synovial Fluid of Patients with Osteoarthritis. Life 2020, 10, 337. [Google Scholar] [CrossRef]
- Gkretsi, V.; Simopoulou, T.; Tsezou, A. Lipid metabolism and osteoarthritis: Lessons from atherosclerosis. Prog. Lipid. Res. 2011, 50, 133–140. [Google Scholar] [CrossRef]
- Gentle, I.E. Supramolecular Complexes in Cell Death and Inflammation and Their Regulation by Autophagy. Front. Cell. Dev. Biol. 2019, 7, 73. [Google Scholar] [CrossRef]
- Neefjes, M.; Housmans, B.A.C.; van den Akker, G.G.H.; van Rhijn, L.W.; Welting, T.J.M.; van der Kraan, P.M. Reporter gene comparison demonstrates interference of complex body fluids with secreted luciferase activity. Sci. Rep. 2021, 11, 1359. [Google Scholar] [CrossRef]
- Wen, A.Y.; Sakamoto, K.M.; Miller, L.S. The role of the transcription factor CREB in immune function. J. Immunol. 2010, 185, 6413–6419. [Google Scholar] [CrossRef]
- Sotiropoulos, A.; Gineitis, D.; Copeland, J.; Treisman, R. Signal-regulated activation of serum response factor is mediated by changes in actin dynamics. Cell 1999, 98, 159–169. [Google Scholar] [CrossRef]
- Frisbie, D.D.; Kawcak, C.E.; McIlwraith, C.W. Evaluation of the effect of extracorporeal shock wave treatment on experimentally induced osteoarthritis in middle carpal joints of horses. Am. J. Vet. Res. 2009, 70, 449–454. [Google Scholar] [CrossRef]
- Demichev, V.; Messner, C.B.; Vernardis, S.I.; Lilley, K.S.; Ralser, M. DIA-NN: Neural networks and interference correction enable deep proteome coverage in high throughput. Nat. Methods 2020, 17, 41–44. [Google Scholar] [CrossRef]
- Housmans, B.A.C.; Neefjes, M.; Surtel, D.A.M.; Vitik, M.; Cremers, A.; van Rhijn, L.W.; van der Kraan, P.M.; van den Akker, G.G.H.; Welting, T.J.M. Synovial fluid from end-stage osteoarthritis induces proliferation and fibrosis of articular chondrocytes via MAPK and RhoGTPase signaling. Osteoarthr. Cartil. 2022, 30, 862–874. [Google Scholar] [CrossRef]
- Willforss, J.; Chawade, A.; Levander, F. NormalyzerDE: Online Tool for Improved Normalization of Omics Expression Data and High-Sensitivity Differential Expression Analysis. J. Proteome Res. 2019, 18, 732–740. [Google Scholar] [CrossRef]
- Zhang, X.; Smits, A.H.; van Tilburg, G.B.; Ovaa, H.; Huber, W.; Vermeulen, M. Proteome-wide identification of ubiquitin interactions using UbIA-MS. Nat. Protoc. 2018, 13, 530–550. [Google Scholar] [CrossRef]
- Law, C.W.; Alhamdoosh, M.; Su, S.; Dong, X.; Tian, L.; Smyth, G.K.; Ritchie, M.E. RNA-seq analysis is easy as 1-2-3 with limma, Glimma and edgeR. F1000Res 2016, 5, 1408. [Google Scholar] [CrossRef]
- Rappoport, N.; Shamir, R. Multi-omic and multi-view clustering algorithms: Review and cancer benchmark. Nucleic Acids Res. 2018, 46, 10546–10562. [Google Scholar] [CrossRef]
- Kumar, L.; Futschik, M.E. Mfuzz: A software package for soft clustering of microarray data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef]
- Velten, B.; Braunger, J.M.; Argelaguet, R.; Arnol, D.; Wirbel, J.; Bredikhin, D.; Zeller, G.; Stegle, O. Identifying temporal and spatial patterns of variation from multimodal data using MEFISTO. Nat. Methods 2022, 19, 179–186. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]
- Huang, C.-Y.; Vesvoranan, O.; Yin, X.; Montoya, A.; Londono, V.; Sawatari, Y.; Garcia-Godoy, F. Anti-Inflammatory Effects of Conditioned Medium of Periodontal Ligament-Derived Stem Cells on Chondrocytes, Synoviocytes, and Meniscus Cells. Stem Cells Dev. 2021, 30, 537–547. [Google Scholar] [CrossRef]
- Cosenza, S.; Ruiz, M.; Toupet, K.; Jorgensen, C.; Noël, D. Mesenchymal stem cells derived exosomes and microparticles protect cartilage and bone from degradation in osteoarthritis. Sci. Rep. 2021, 7, 16214. [Google Scholar] [CrossRef]
- Hotham, W.E.; Thompson, C.; Szu-Ting, L.; Henson, F.M.D. The anti-inflammatory effects of equine bone marrow stem cell‐derived extracellular vesicles on autologous chondrocytes. Veter. Rec. Open 2021, 8, e22. [Google Scholar] [CrossRef]
- Ni, Z.; Kuang, L.; Chen, H.; Xie, Y.; Zhang, B.; Ouyang, J.; Wu, J.; Zhou, S.; Chen, L.; Su, N.; et al. The exosome-like vesicles from osteoarthritic chondrocyte enhanced mature IL-1beta production of macrophages and aggra-vated synovitis in osteoarthritis. Cell Death Dis. 2019, 10, 522. [Google Scholar]
- Liu, X.; Shortt, C.; Zhang, F.; Bater, M.Q.; Cowman, M.K.; Kirsch, T. Extracellular Vesicles Released from Articular Chondrocytes Play a Major Role in Cell–Cell Communication. J. Orthop. Res. 2019, 38, 731–739. [Google Scholar] [CrossRef]
- Zhang, S.; Chuah, S.J.; Lai, R.C.; Hui, J.H.P.; Lim, S.K.; Toh, W.S. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials 2018, 156, 16–27. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biofluid | Day (Versus Day 0) | p-Value < 0.05 |
|---|---|---|
| Plasma | 10 | 4 |
| Plasma | 35 | 8 |
| Plasma | 42 | 5 |
| Plasma | 49 | 12 |
| Plasma | 56 | 8 |
| Plasma | 63 | 3 |
| Plasma | continuous | 36 |
| Synovial fluid | 10 | 34 |
| Synovial fluid | 35 | 6 |
| Synovial fluid | 42 | 12 |
| Synovial fluid | 49 | 3 |
| Synovial fluid | 56 | 6 |
| Synovial fluid | 63 | 6 |
| Synovial fluid | continuous | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anderson, J.R.; Johnson, E.; Jenkins, R.; Jacobsen, S.; Green, D.; Walters, M.; Bundgaard, L.; Hausmans, B.A.C.; van den Akker, G.; Welting, T.J.M.; et al. Multi-Omic Temporal Landscape of Plasma and Synovial Fluid-Derived Extracellular Vesicles Using an Experimental Model of Equine Osteoarthritis. Int. J. Mol. Sci. 2023, 24, 14888. https://doi.org/10.3390/ijms241914888
Anderson JR, Johnson E, Jenkins R, Jacobsen S, Green D, Walters M, Bundgaard L, Hausmans BAC, van den Akker G, Welting TJM, et al. Multi-Omic Temporal Landscape of Plasma and Synovial Fluid-Derived Extracellular Vesicles Using an Experimental Model of Equine Osteoarthritis. International Journal of Molecular Sciences. 2023; 24(19):14888. https://doi.org/10.3390/ijms241914888
Chicago/Turabian StyleAnderson, James R., Emily Johnson, Rosalind Jenkins, Stine Jacobsen, Daniel Green, Marie Walters, Louise Bundgaard, Bas A. C. Hausmans, Guus van den Akker, Tim J. M. Welting, and et al. 2023. "Multi-Omic Temporal Landscape of Plasma and Synovial Fluid-Derived Extracellular Vesicles Using an Experimental Model of Equine Osteoarthritis" International Journal of Molecular Sciences 24, no. 19: 14888. https://doi.org/10.3390/ijms241914888