
Effect of Fetal Bovine Serum or Basic Fibroblast Growth Factor on Cell Survival and the Proliferation of Neural Stem Cells: The Influence of Homocysteine Treatment
 , , and
, , and
Abstract
:1. Introduction
2. Results
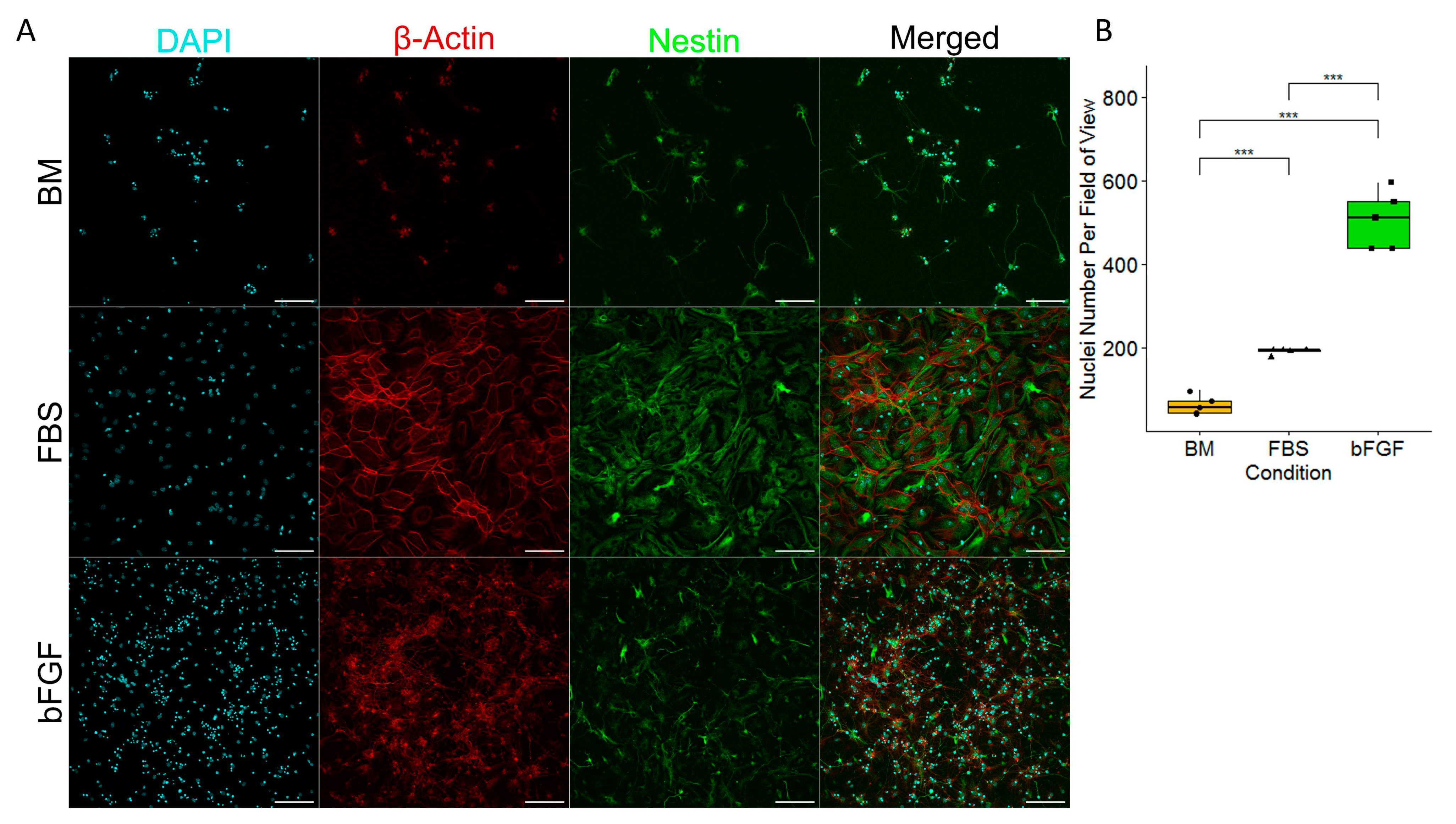
2.1. Cell Cultures of Precursors of the Nervous System Require Either FBS or bFGF to Prevent Cell Death and Achieve a Satisfactory Level of the Presence of Actin and Nestin Filaments
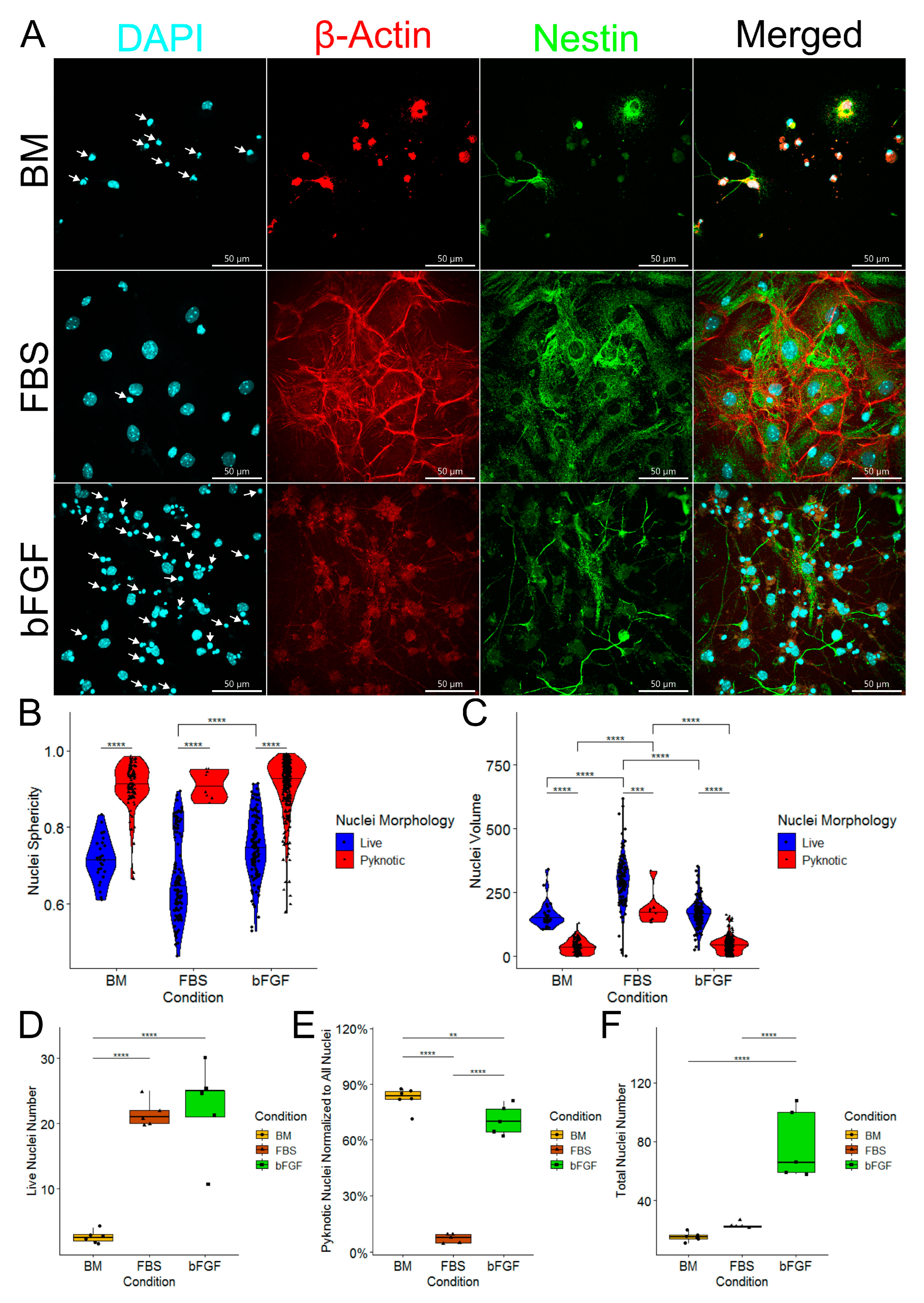
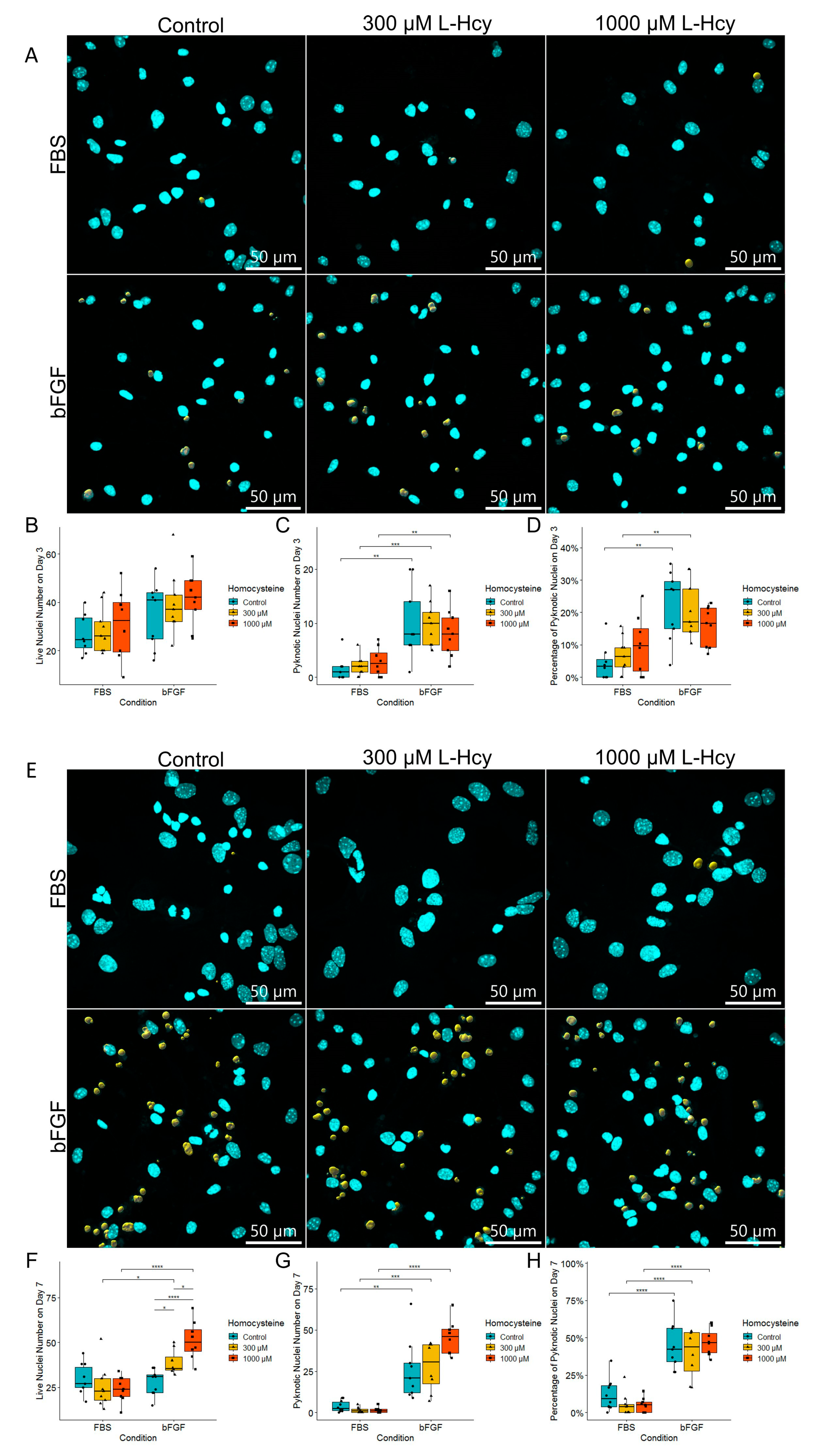
2.2. Medium Enriched by FBS Yields a Significantly Lower Number of Pyknotic Nuclei Compared to bFGF-Enriched Medium and Basal Medium
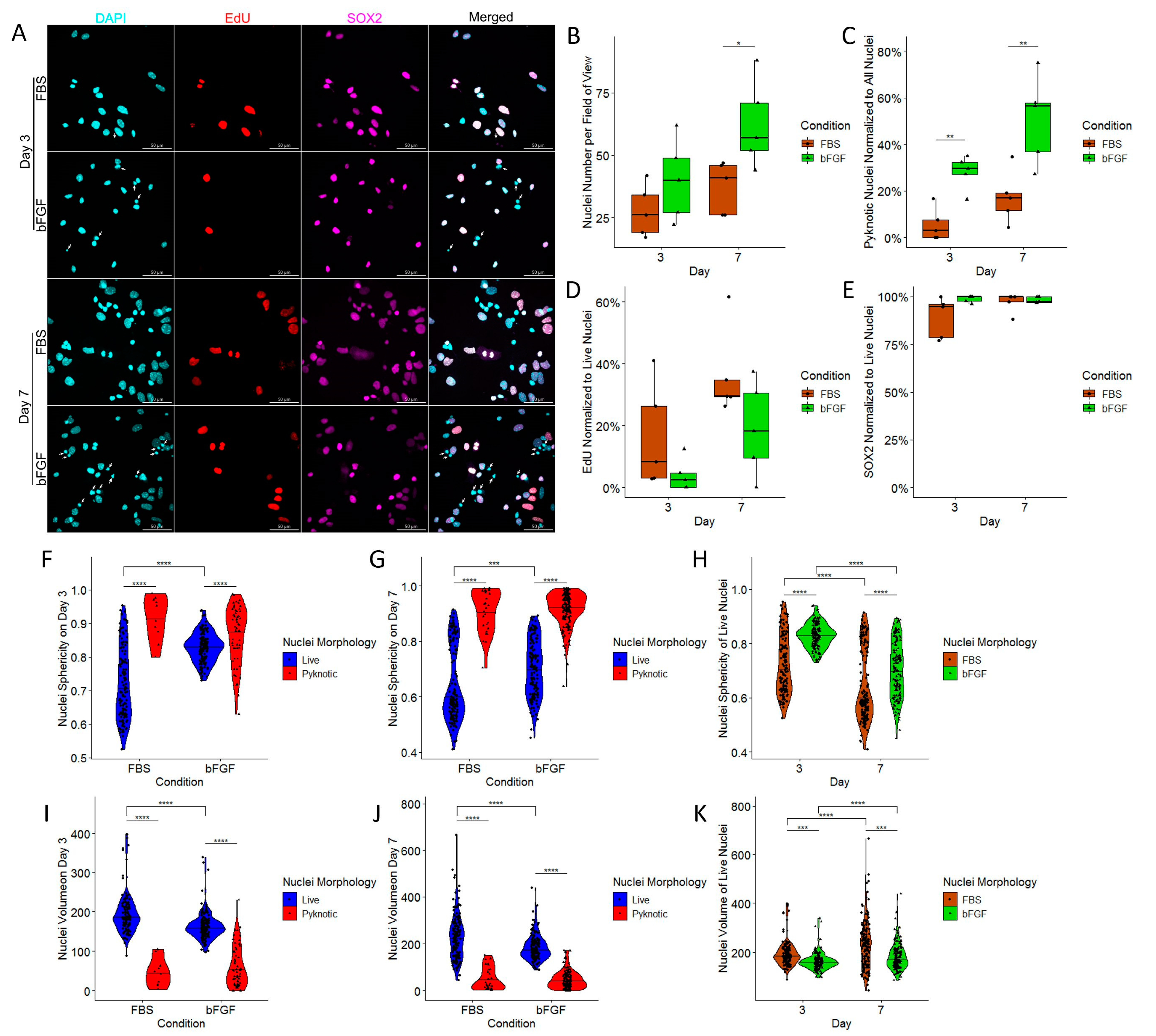
2.3. Treatment with bFGF Increases the Number of Neuronal Precursors While Marginally Decreasing Cell Proliferation
2.4. FBS-Treated NSCs Develop into Reactive Astrocytes and Neurons, Whereas bFGF-Treated NSCs Develop into Astrocytes with Quiescent Morphology and Neurons that Increase in Volume over Time
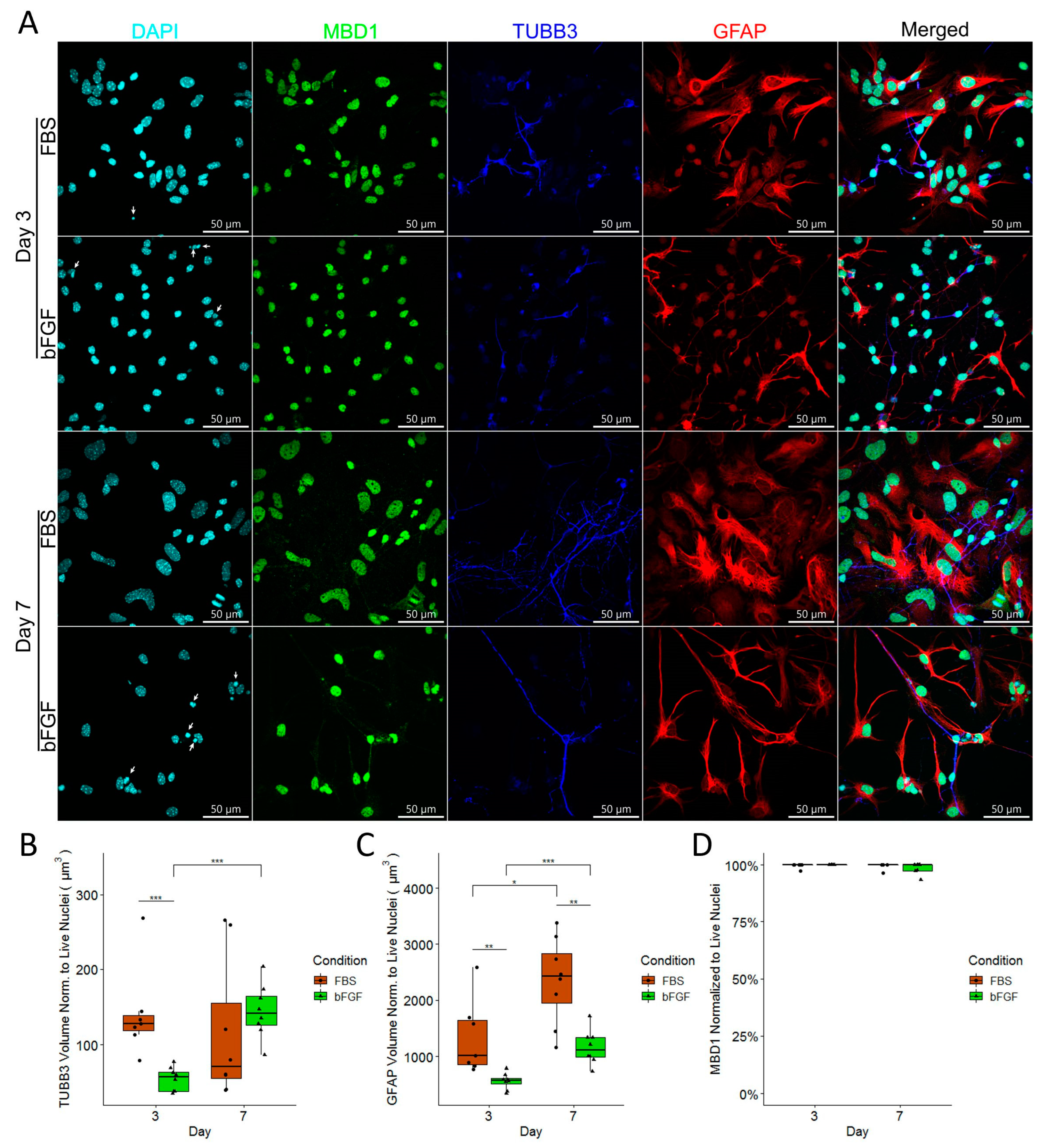
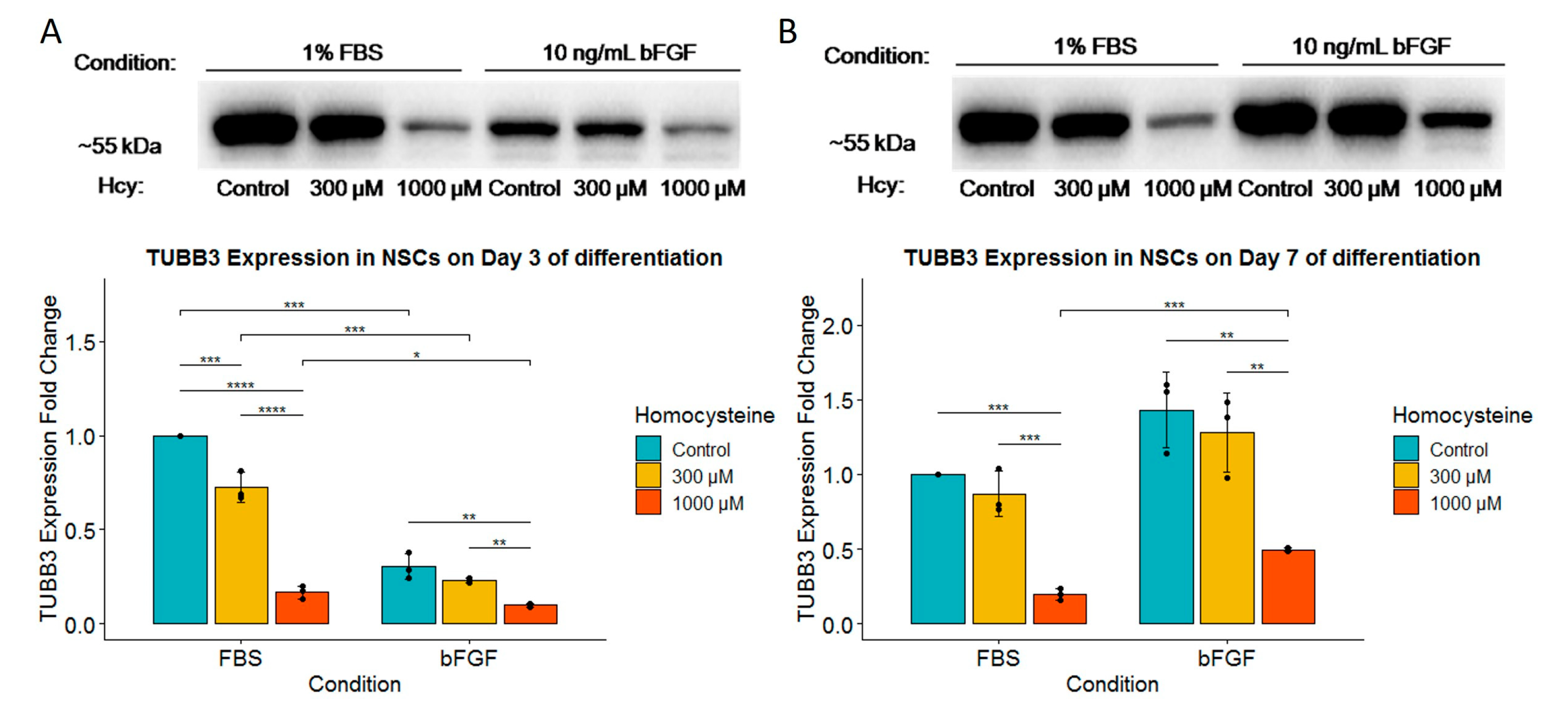
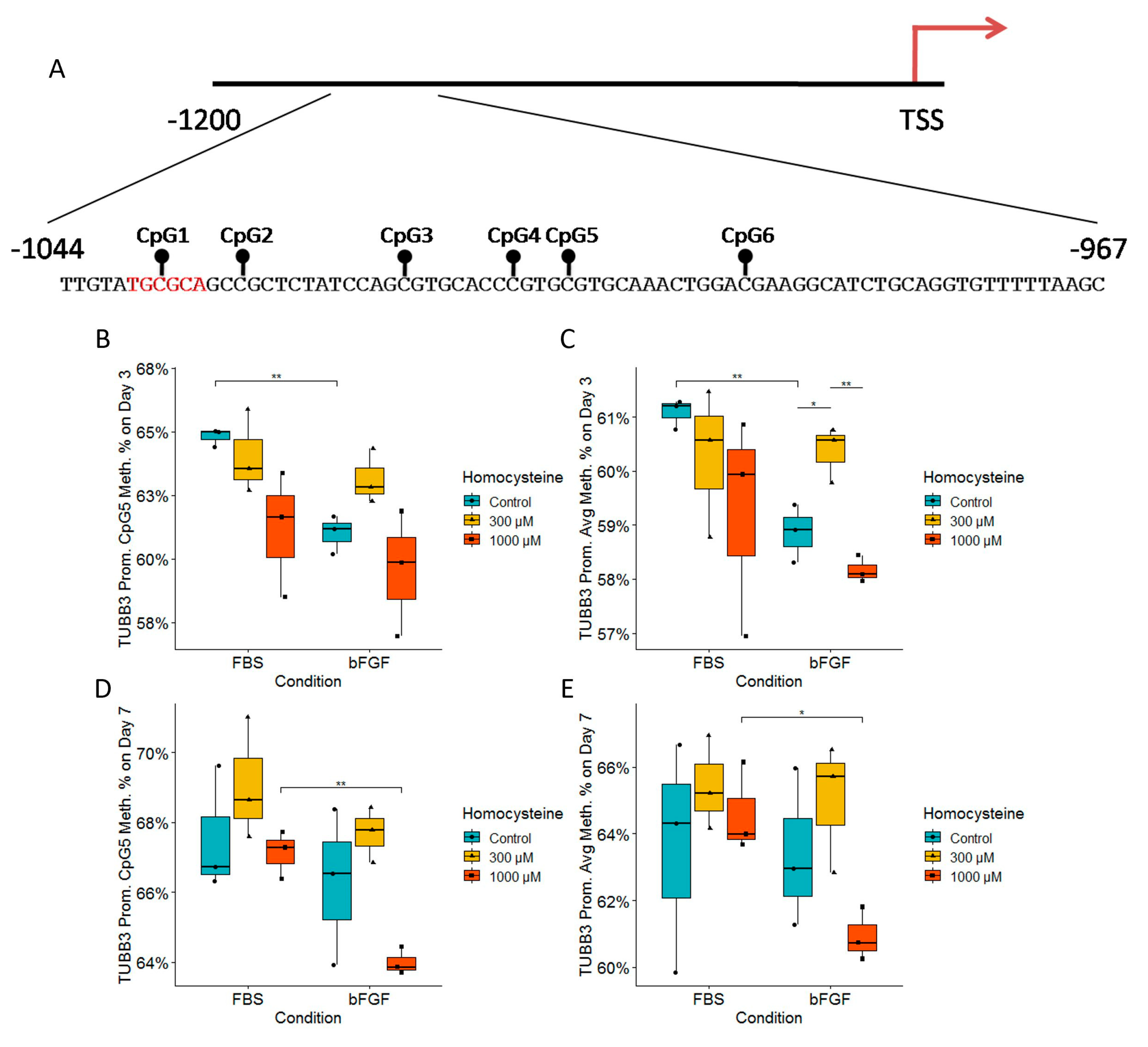
2.5. Homocysteine Decreases Expression of TUBB3 in Both FBS- and bFGF-Treated Immature Neural Cells in a Dose-Dependent Manner without Influence on the Total Number of Cells
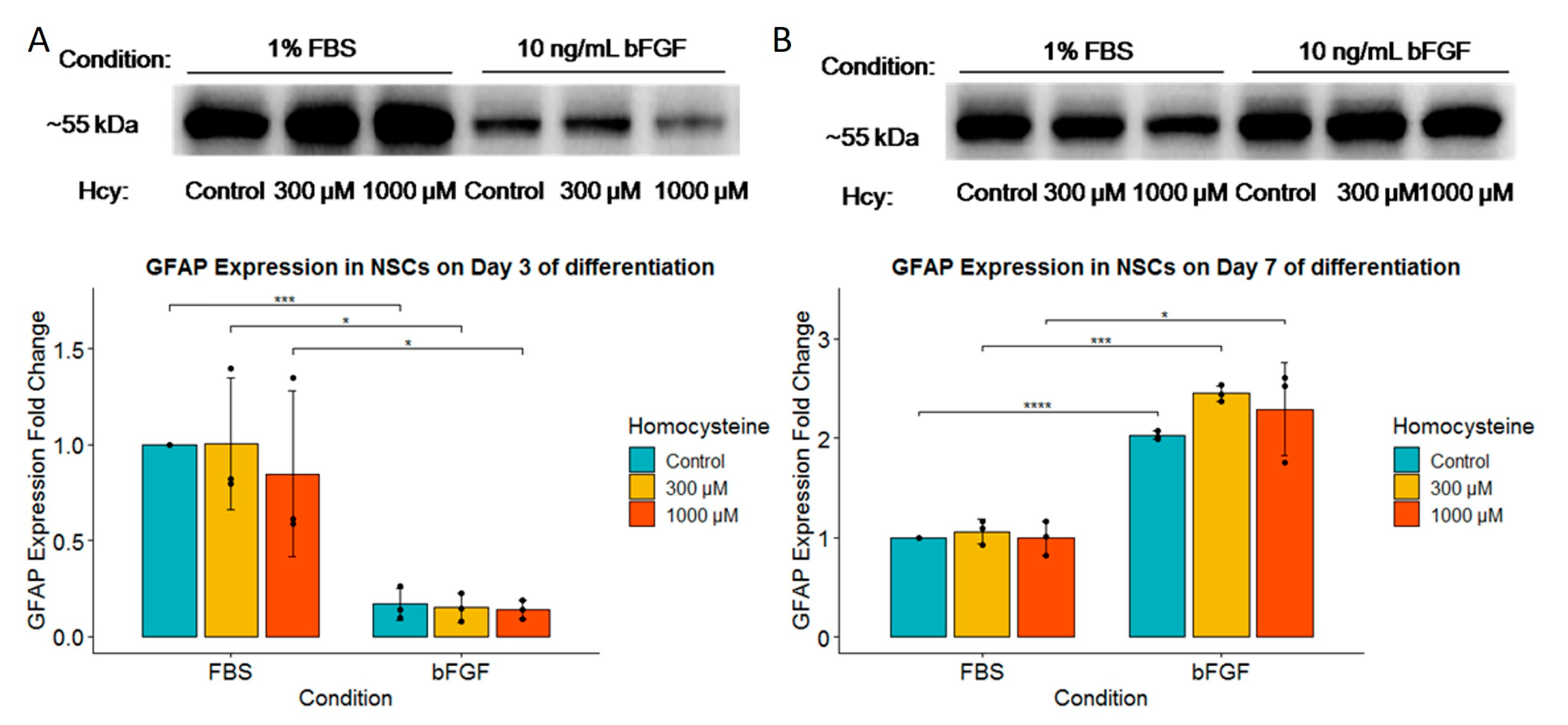
2.6. Expression of GFAP Was Much More Strongly Induced by FBS than by bFGF, and It Was Not Influenced by Homocysteine
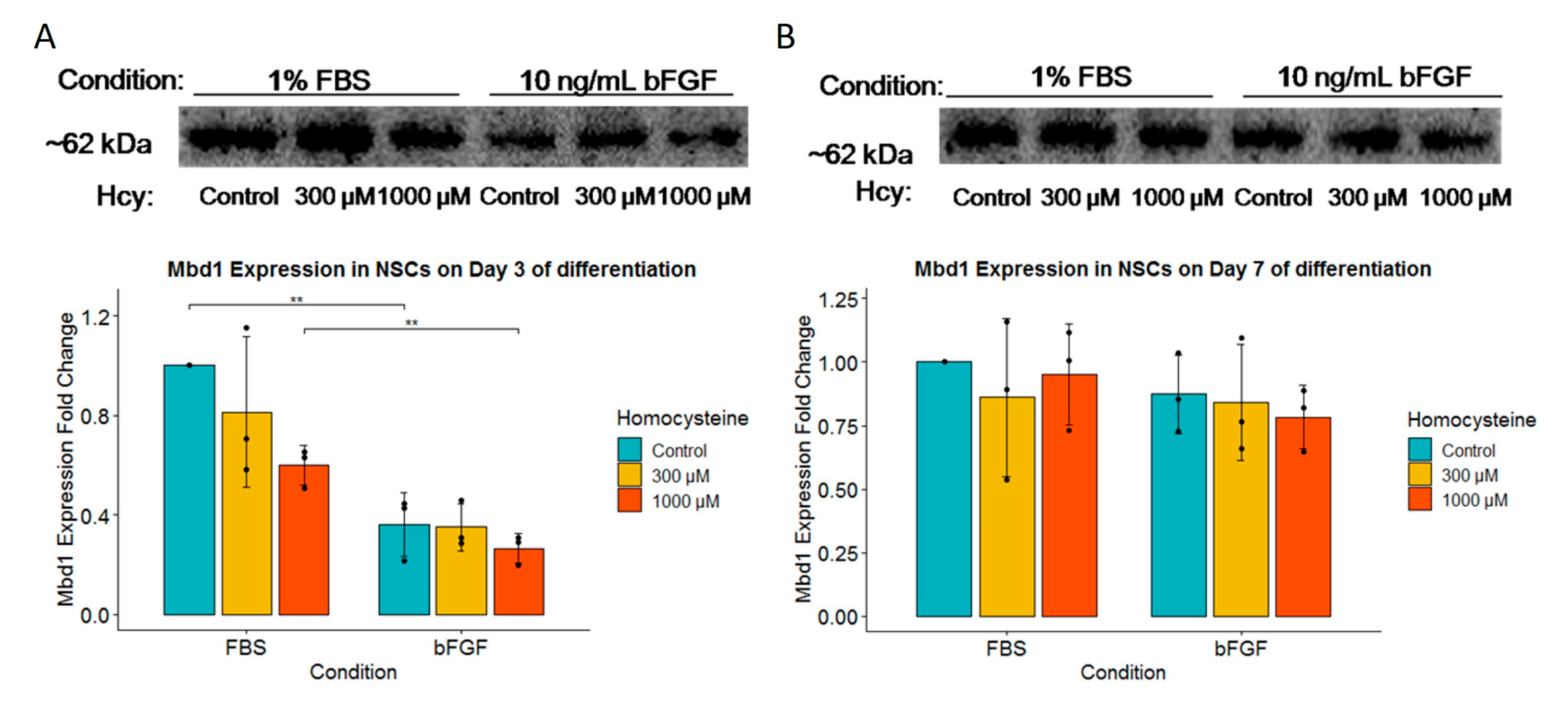
2.7. Although MBD1 Expression Is Unaffected by Hcy Treatment, Early Neuronal Precursors Treated with FBS Have Higher MBD1 Expression
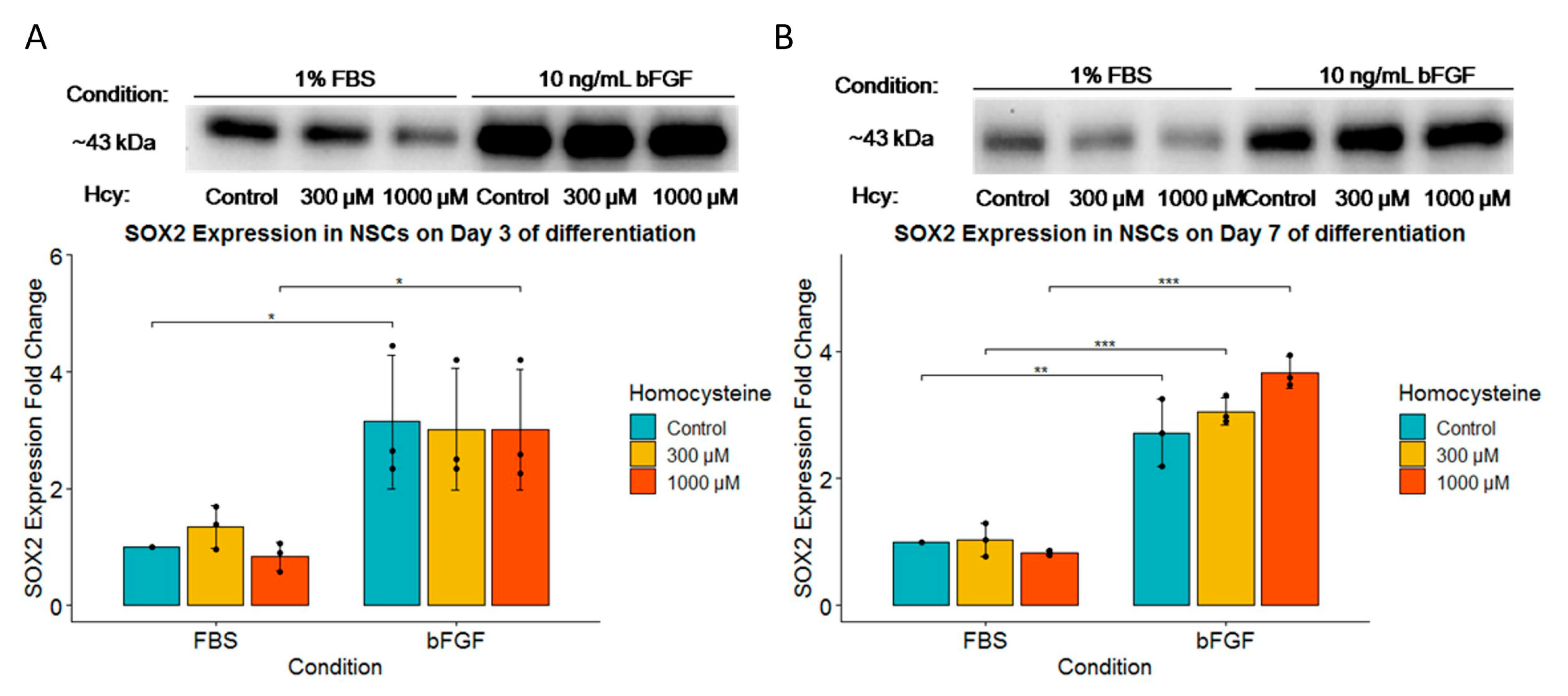
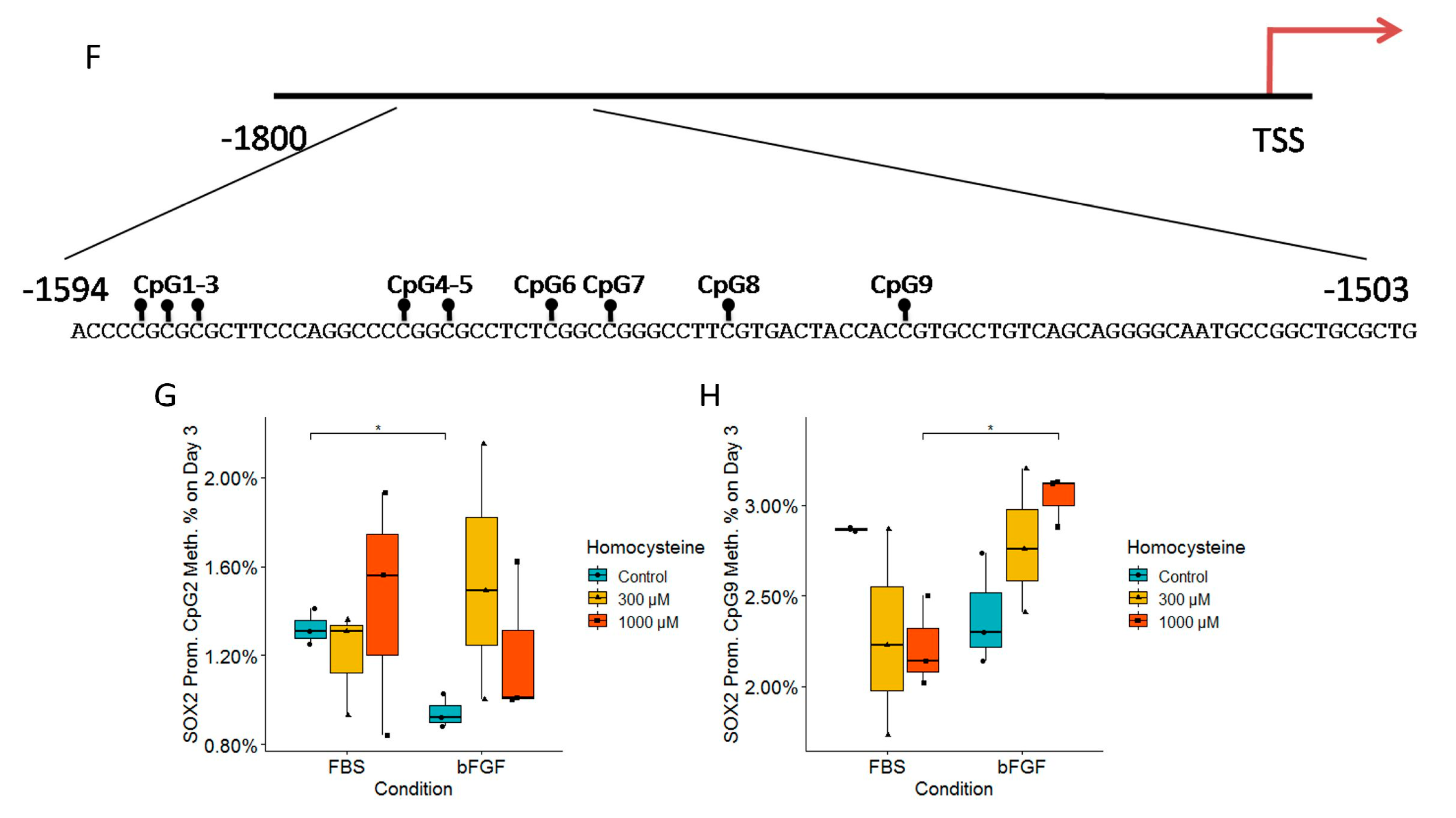
2.8. bFGF Significantly Increases SOX2 Expression on Both Day 3 and Day 7, While SOX2 Expression Is Unaffected by Hcy Treatment
2.9. Analysis of the DNA Methylation Status of Tubb3 and Sox2 Promoters Revealed the Impact of Treated Media on Epigenetics
3. Discussion
4. Materials and Methods
4.1. Isolation and Passaging of Neural Stem Cells
4.2. Differentiation and Treatment of Neural Stem Cells
4.3. Immunocytochemistry
4.4. Pyknotic and Live Nuclei Demarcation in Imaris Software Version 9.9.1
4.5. Western Blot
4.6. DNA Methylation Analysis
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Radoszkiewicz, K.; Hribljan, V.; Isakovic, J.; Mitrecic, D.; Sarnowska, A. Critical Points for Optimizing Long-Term Culture and Neural Differentiation Capacity of Rodent and Human Neural Stem Cells to Facilitate Translation into Clinical Settings. Exp. Neurol. 2023, 363, 114353. [Google Scholar] [CrossRef]
- Chen, X.; Li, Z.; Cheng, Y.; Kardami, E.; Loh, Y.P. Low and High Molecular Weight FGF-2 Have Differential Effects on Astrocyte Proliferation, but Are Both Protective Against Aβ-Induced Cytotoxicity. Front. Mol. Neurosci. 2020, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Xie, D.; Wu, T.; Xu, W.; Meng, Q.; Cao, K.; Hu, J. Evaluation of the Protective Roles of Alpha-Lipoic Acid Supplementation on Nanomaterial-Induced Toxicity: A Meta-Analysis of in Vitro and in Vivo Studies. Front. Nutr. 2022, 9, 991524. [Google Scholar] [CrossRef] [PubMed]
- Olfson, M.; Stroup, T.S.; Huang, C.; Wall, M.M.; Gerhard, T. Age and Incidence of Dementia Diagnosis. J. Gen. Intern. Med. 2021, 36, 2167–2169. [Google Scholar] [CrossRef]
- Boots, A.; Wiegersma, A.M.; Vali, Y.; van den Hof, M.; Langendam, M.W.; Limpens, J.; Backhouse, E.V.; Shenkin, S.D.; Wardlaw, J.M.; Roseboom, T.J.; et al. Shaping the Risk for Late-Life Neurodegenerative Disease: A Systematic Review on Prenatal Risk Factors for Alzheimer’s Disease-Related Volumetric Brain Biomarkers. Neurosci. Biobehav. Rev. 2023, 146, 105019. [Google Scholar] [CrossRef]
- Walter, K.R.; Ricketts, D.K.; Presswood, B.H.; Smith, S.M.; Mooney, S.M. Prenatal Alcohol Exposure Causes Persistent Microglial Activation and Age- and Sex-Specific Effects on Cognition and Metabolic Outcomes in an Alzheimer’s Disease Mouse Model. Am. J. Drug Alcohol Abuse 2023, 49, 302–320. [Google Scholar] [CrossRef]
- Koklesova, L.; Mazurakova, A.; Samec, M.; Biringer, K.; Samuel, S.M.; Büsselberg, D.; Kubatka, P.; Golubnitschaja, O. Homocysteine Metabolism as the Target for Predictive Medical Approach, Disease Prevention, Prognosis, and Treatments Tailored to the Person. EPMA J. 2021, 12, 477–505. [Google Scholar] [CrossRef]
- Huang, T.; Yuan, G.; Zhang, Z.; Zou, Z.; Li, D. Cardiovascular Pathogenesis in Hyperhomocysteinemia. Asia Pac. J. Clin. Nutr. 2008, 17, 8–16. [Google Scholar]
- Zhuo, J.-M.; Wang, H.; Praticò, D. Is Hyperhomocysteinemia an Alzheimer’s Disease (AD) Risk Factor, an AD Marker, or Neither? Trends Pharmacol. Sci. 2011, 32, 562–571. [Google Scholar] [CrossRef]
- Zhang, Q.; Bai, B.; Mei, X.; Wan, C.; Cao, H.; Li, D.; Wang, S.; Zhang, M.; Wang, Z.; Wu, J.; et al. Elevated H3K79 Homocysteinylation Causes Abnormal Gene Expression during Neural Development and Subsequent Neural Tube Defects. Nat. Commun. 2018, 9, 3436. [Google Scholar] [CrossRef]
- Tang, K.-F.; Li, Y.-L.; Wang, H.-Y. Quantitative Assessment of Maternal Biomarkers Related to One-Carbon Metabolism and Neural Tube Defects. Sci. Rep. 2015, 5, 8510. [Google Scholar] [CrossRef] [PubMed]
- Cecchini, M.S.; Bourckhardt, G.F.; Jaramillo, M.L.; Ammar, D.; Müller, Y.M.R.; Nazari, E.M. Exposure to Homocysteine Leads to Cell Cycle Damage and Reactive Gliosis in the Developing Brain. Reprod. Toxicol. 2019, 87, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Shiue, K.; Schomberg, D.; Zhou, F.C. Cellular Epigenetic Modifications of Neural Stem Cell Differentiation. Cell Transplant. 2009, 18, 1197–1211. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Qin, S.; Luo, S.; Cui, S.; Huang, G.; Zhang, X. Homocysteine Induces Cytotoxicity and Proliferation Inhibition in Neural Stem Cells via DNA Methylation in Vitro. FEBS J. 2014, 281, 2088–2096. [Google Scholar] [CrossRef]
- Mansukhani, A.; Ambrosetti, D.; Holmes, G.; Cornivelli, L.; Basilico, C. Sox2 Induction by FGF and FGFR2 Activating Mutations Inhibits Wnt Signaling and Osteoblast Differentiation. J. Cell Biol. 2005, 168, 1065–1076. [Google Scholar] [CrossRef]
- Louis, S.A.; Mak, C.K.H.; Reynolds, B.A. Methods to Culture, Differentiate, and Characterize Neural Stem Cells from the Adult and Embryonic Mouse Central Nervous System. In Basic Cell Culture Protocols; Humana: Totowa, NJ, USA, 2013; pp. 479–506. [Google Scholar]
- Eidet, J.R.; Pasovic, L.; Maria, R.; Jackson, C.J.; Utheim, T.P. Objective Assessment of Changes in Nuclear Morphology and Cell Distribution Following Induction of Apoptosis. Diagn. Pathol. 2014, 9, 92. [Google Scholar] [CrossRef]
- Hou, L.; Liu, K.; Li, Y.; Ma, S.; Ji, X.; Liu, L. Necrotic Pyknosis Is a Morphologically and Biochemically Distinct Event from Apoptotic Pyknosis. J. Cell Sci. 2016, 129, 3084–3090. [Google Scholar] [CrossRef]
- Wohlan, K.; Goy, S.; Olling, A.; Srivaratharajan, S.; Tatge, H.; Genth, H.; Gerhard, R. Pyknotic Cell Death Induced by C Lostridium Difficile TcdB: Chromatin Condensation and Nuclear Blister Are Induced Independently of the Glucosyltransferase Activity. Cell. Microbiol. 2014, 16, 1678–1692. [Google Scholar] [CrossRef]
- Chen, B.; Co, C.; Ho, C.-C. Cell Shape Dependent Regulation of Nuclear Morphology. Biomaterials 2015, 67, 129–136. [Google Scholar] [CrossRef]
- Israsena, N.; Hu, M.; Fu, W.; Kan, L.; Kessler, J.A. The Presence of FGF2 Signaling Determines Whether β-Catenin Exerts Effects on Proliferation or Neuronal Differentiation of Neural Stem Cells. Dev. Biol. 2004, 268, 220–231. [Google Scholar] [CrossRef]
- Luo, S.; Zhang, X.; Yu, M.; Yan, H.; Liu, H.; Wilson, J.X.; Huang, G. Folic Acid Acts Through DNA Methyltransferases to Induce the Differentiation of Neural Stem Cells into Neurons. Cell Biochem. Biophys. 2013, 66, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Aswad, H.; Jalabert, A.; Rome, S. Depleting Extracellular Vesicles from Fetal Bovine Serum Alters Proliferation and Differentiation of Skeletal Muscle Cells in Vitro. BMC Biotechnol. 2016, 16, 32. [Google Scholar] [CrossRef] [PubMed]
- Jagečić, D.; Petrović, D.J.; Šimunić, I.; Isaković, J.; Mitrečić, D. The Oxygen and Glucose Deprivation of Immature Cells of the Nervous System Exerts Distinct Effects on Mitochondria, Mitophagy, and Autophagy, Depending on the Cells’ Differentiation Stage. Brain Sci. 2023, 13, 910. [Google Scholar] [CrossRef] [PubMed]
- Pilgrim, C.R.; McCahill, K.A.; Rops, J.G.; Dufour, J.M.; Russell, K.A.; Koch, T.G. A Review of Fetal Bovine Serum in the Culture of Mesenchymal Stromal Cells and Potential Alternatives for Veterinary Medicine. Front. Vet. Sci. 2022, 9, 859025. [Google Scholar] [CrossRef] [PubMed]
- van der Valk, J.; Gstraunthaler, G. Fetal Bovine Serum (FBS)—A Pain in the Dish? Altern. Lab. Anim. 2017, 45, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.G.; Winkler, J.; Kempermann, G.; Thal, L.J.; Gage, F.H. Epidermal Growth Factor and Fibroblast Growth Factor-2 Have Different Effects on Neural Progenitors in the Adult Rat Brain. J. Neurosci. 1997, 17, 5820–5829. [Google Scholar] [CrossRef] [PubMed]
- Mudò, G.; Bonomo, A.; Di Liberto, V.; Frinchi, M.; Fuxe, K.; Belluardo, N. The FGF-2/FGFRs Neurotrophic System Promotes Neurogenesis in the Adult Brain. J. Neural Transm. 2009, 116, 995–1005. [Google Scholar] [CrossRef]
- Jin, K.; Sun, Y.; Xie, L.; Batteur, S.; Mao, X.O.; Smelick, C.; Logvinova, A.; Greenberg, D.A. Neurogenesis and Aging: FGF-2 and HB-EGF Restore Neurogenesis in Hippocampus and Subventricular Zone of Aged Mice. Aging Cell 2003, 2, 175–183. [Google Scholar] [CrossRef]
- Kiyota, T.; Ingraham, K.L.; Jacobsen, M.T.; Xiong, H.; Ikezu, T. FGF2 Gene Transfer Restores Hippocampal Functions in Mouse Models of Alzheimer’s Disease and Has Therapeutic Implications for Neurocognitive Disorders. Proc. Natl. Acad. Sci. USA 2011, 108, E1339–E1348. [Google Scholar] [CrossRef]
- Rabaneda, L.G.; Carrasco, M.; López-Toledano, M.A.; Murillo-Carretero, M.; Ruiz, F.A.; Estrada, C.; Castro, C. Homocysteine Inhibits Proliferation of Neuronal Precursors in the Mouse Adult Brain by Impairing the Basic Fibroblast Growth Factor Signaling Cascade and Reducing Extracellular Regulated Kinase 1/2-dependent Cyclin E Expression. FASEB J. 2008, 22, 3823–3835. [Google Scholar] [CrossRef]
- Wang, D.; Chen, Y.; Ruan, M.; Zhou, A.; Qian, Y.; Chen, C. Homocysteine Inhibits Neural Stem Cells Survival by Inducing DNA Interstrand Cross-Links via Oxidative Stress. Neurosci. Lett. 2016, 635, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Maler, J.; Seifert, W.; Hüther, G.; Wiltfang, J.; Rüther, E.; Kornhuber, J.; Bleich, S. Homocysteine Induces Cell Death of Rat Astrocytes in Vitro. Neurosci. Lett. 2003, 347, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Zhang, X.; Luo, S.; Liu, H.; Wang, X.; Gao, Y.; Wilson, J.X.; Huang, G. Effects of Homocysteine on ERK Signaling and Cell Proliferation in Fetal Neural Stem Cells In Vitro. Cell Biochem. Biophys. 2013, 66, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Yin, Y.; Feng, X.; Chen, Y.; Shi, J.; Weng, H.; Wang, D. Homocysteine Aggravates DNA Damage by Impairing the FA/Brca1 Pathway in NE4C Murine Neural Stem Cells. Int. J. Med. Sci. 2020, 17, 2477–2486. [Google Scholar] [CrossRef]
- Prah, J.; Winters, A.; Chaudhari, K.; Hersh, J.; Liu, R.; Yang, S.-H. A Novel Serum Free Primary Astrocyte Culture Method That Mimic Quiescent Astrocyte Phenotype. J. Neurosci. Methods 2019, 320, 50–63. [Google Scholar] [CrossRef]
- Stern, F.; Berner, Y.N.; Polyak, Z.; Komarnitsky, M.; Sela, B.-A.; Hopp, M.; Dror, Y. Homocysteine Effect on Protein Degradation Rates. Clin. Biochem. 2004, 37, 1002–1009. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, G.; Cowan, N.J.; Cabral, F. Mutations Affecting β-Tubulin Folding and Degradation. J. Biol. Chem. 2006, 281, 13628–13635. [Google Scholar] [CrossRef]
- Sharma, G.S.; Bhattacharya, R.; Singh, L.R. Functional Inhibition of Redox Regulated Heme Proteins: A Novel Mechanism towards Oxidative Stress Induced by Homocysteine. Redox Biol. 2021, 46, 102080. [Google Scholar] [CrossRef]
- Ye, M.; Li, H.; Luo, H.; Zhou, Y.; Luo, W.; Lin, Z. Potential Antioxidative Activity of Homocysteine in Erythrocytes under Oxidative Stress. Antioxidants 2023, 12, 202. [Google Scholar] [CrossRef]
- Guo, J.; Kim, H.S.; Asmis, R.; Ludueña, R.F. Interactions of β Tubulin Isotypes with Glutathione in Differentiated Neuroblastoma Cells Subject to Oxidative Stress. Cytoskeleton 2018, 75, 283–289. [Google Scholar] [CrossRef]
- Chen, N.; Liu, J.; Qiao, Z.; Liu, Y.; Yang, Y.; Jiang, C.; Wang, X.; Wang, C. Chemical Proteomic Profiling of Protein: N -Homocysteinylation with a Thioester Probe. Chem. Sci. 2018, 9, 2826–2850. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H.; Zhang, L.; Bardeguez, A.; Aviv, A. Homocysteine Thiolactone and Protein Homocysteinylation in Human Endothelial Cells Implications for Atherosclerosis. Circ. Res. 2000, 87, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Paoli, P.; Sbrana, F.; Tiribilli, B.; Caselli, A.; Pantera, B.; Cirri, P.; De Donatis, A.; Formigli, L.; Nosi, D.; Manao, G.; et al. Protein N-Homocysteinylation Induces the Formation of Toxic Amyloid-like Protofibrils. J. Mol. Biol. 2010, 400, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Leeman, D.S.; Hebestreit, K.; Ruetz, T.; Webb, A.E.; McKay, A.; Pollina, E.A.; Dulken, B.W.; Zhao, X.; Yeo, R.W.; Ho, T.T.; et al. Lysosome Activation Clears Aggregates and Enhances Quiescent Neural Stem Cell Activation during Aging. Science 2018, 359, 1277–1283. [Google Scholar] [CrossRef]
- Bossenmeyer-Pourié, C.; Smith, A.D.; Lehmann, S.; Deramecourt, V.; Sablonnière, B.; Camadro, J.-M.; Pourié, G.; Kerek, R.; Helle, D.; Umoret, R.; et al. N-Homocysteinylation of Tau and MAP1 Is Increased in Autopsy Specimens of Alzheimer’s Disease and Vascular Dementia. J. Pathol. 2019, 248, 291–303. [Google Scholar] [CrossRef]
- Karima, O.; Riazi, G.; Khodadadi, S.; Aryapour, H.; Nasiri Khalili, M.A.; Yousefi, L.; Moosavi-Movahedi, A.A. Altered Tubulin Assembly Dynamics with N-Homocysteinylated Human 4R/1N Tau in Vitro. FEBS Lett. 2012, 586, 3914–3919. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, D.-X.; Wang, F.-W.; Zhang, Q.; Du, Z.-X.; Zhan, J.-M.; Yuan, Q.-H.; Ling, E.-A.; Hao, A.-J. L-Cysteine Promotes the Proliferation and Differentiation of Neural Stem Cells via the CBS/H2S Pathway. Neuroscience 2013, 237, 106–117. [Google Scholar] [CrossRef]
- Singh, S.; Padovani, D.; Leslie, R.A.; Chiku, T.; Banerjee, R. Relative Contributions of Cystathionine β-Synthase and γ-Cystathionase to H2S Biogenesis via Alternative Trans-Sulfuration Reactions. J. Biol. Chem. 2009, 284, 22457–22466. [Google Scholar] [CrossRef]
- Kruger, W.D.; Wang, L.; Jhee, K.H.; Singh, R.H.; Elsas, L.J. Cystathionine?-Synthase Deficiency in Georgia (USA): Correlation of Clinical and Biochemical Phenotype with Genotype. Hum. Mutat. 2003, 22, 434–441. [Google Scholar] [CrossRef]
- Liu, D.; Wang, Z.; Zhan, J.; Zhang, Q.; Wang, J.; Zhang, Q.; Xian, X.; Luan, Q.; Hao, A. Hydrogen Sulfide Promotes Proliferation and Neuronal Differentiation of Neural Stem Cells and Protects Hypoxia-Induced Decrease in Hippocampal Neurogenesis. Pharmacol. Biochem. Behav. 2014, 116, 55–63. [Google Scholar] [CrossRef]
- Sharma, G.S.; Kumar, T.; Dar, T.A.; Singh, L.R. Protein N-Homocysteinylation: From Cellular Toxicity to Neurodegeneration. Biochim. Biophys. Acta 2015, 1850, 2239–2245. [Google Scholar] [CrossRef] [PubMed]
- Jakubowski, H. Protein N-Homocysteinylation: Implications for Atherosclerosis. Biomed. Pharmacother. 2001, 55, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Sibrian-Vazquez, M.; Escobedo, J.O.; Lim, S.; Samoei, G.K.; Strongin, R.M. Homocystamides Promote Free-Radical and Oxidative Damage to Proteins. Proc. Natl. Acad. Sci. USA 2010, 107, 551–554. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Amaral, A.; McCann, A.; Brennan, L. Homocysteine Levels Impact Directly on Epigenetic Reprogramming in Astrocytes. Neurochem. Int. 2011, 58, 833–838. [Google Scholar] [CrossRef]
- Cao, S.; Du, J.; Lv, Y.; Lin, H.; Mao, Z.; Xu, M.; Liu, M.; Liu, Y. PAX3 Inhibits β-Tubulin-III Expression and Neuronal Differentiation of Neural Stem Cell. Biochem. Biophys. Res. Commun. 2017, 485, 307–311. [Google Scholar] [CrossRef]
- Clouaire, T.; de las Heras, J.I.; Merusi, C.; Stancheva, I. Recruitment of MBD1 to Target Genes Requires Sequence-Specific Interaction of the MBD Domain with Methylated DNA. Nucleic Acids Res. 2010, 38, 4620–4634. [Google Scholar] [CrossRef]
- Improda, T.; Morgera, V.; Vitale, M.; Chiariotti, L.; Passaro, F.; Feola, A.; Porcellini, A.; Cuomo, M.; Pezone, A. Specific Methyl-CpG Configurations Define Cell Identity through Gene Expression Regulation. Int. J. Mol. Sci. 2023, 24, 9951. [Google Scholar] [CrossRef]
- Bunina, D.; Abazova, N.; Diaz, N.; Noh, K.M.; Krijgsveld, J.; Zaugg, J.B. Genomic Rewiring of SOX2 Chromatin Interaction Network during Differentiation of ESCs to Postmitotic Neurons. Cell Syst. 2020, 10, 480–494. [Google Scholar] [CrossRef]
- Miyagi, S.; Saito, T.; Mizutani, K.; Masuyama, N.; Gotoh, Y.; Iwama, A.; Nakauchi, H.; Masui, S.; Niwa, H.; Nishimoto, M.; et al. The Sox-2 Regulatory Regions Display Their Activities in Two Distinct Types of Multipotent Stem Cells. Mol. Cell. Biol. 2004, 24, 4207–4220. [Google Scholar] [CrossRef]
- Peng, H.; Chen, Q.; Zheng, J. Isolation and Culture of Neural Stem/Progenitor Cells. In Current Laboratory Methods in Neuroscience Research; Springer: New York, NY, USA, 2014; pp. 129–136. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gen | Primer | Sequence | bp | Annealing Temp. | Number of Cycles |
|---|---|---|---|---|---|
| TUBB3 | F | 5′-GGGGAGGGGTATTTTTTGAGAATAA-3′ | 199 | 56 °C | 50 |
| R | 5′-BIOTIN-ACACTTCAATAATCCACAAACATT-3′ | ||||
| Seq | 5′-GGATGTTTATTTTTAGAGAAAG-3′ | ||||
| Sequence to analyze | 5′-TTGTATGYGTAGTYGTTTTATTTAGYGTGTATTYGTGYGTGTAAATTGGAYGAAGGTATTTGTAGGTGTTTTTAAGT-3′ | ||||
| SOX2 | F | 5′-GTTTGGGTTTGTTTGGTG-3′ | 249 | 56 °C | 50 |
| R | 5′-BIOTIN-AACTTCCTAACATCCCAC-3′ | ||||
| Seq | 5′-TGTATTTGTATTTTTGG-3′ | ||||
| Sequence to analyze | 5′-ATTTYGYGYGTTTTTTAGGTTTYGGYGTTTTTYGGTYGGGTTTTYGTGATTATTATYGTGTTTGTTAGTAGGG-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrović, D.J.; Jagečić, D.; Krasić, J.; Sinčić, N.; Mitrečić, D. Effect of Fetal Bovine Serum or Basic Fibroblast Growth Factor on Cell Survival and the Proliferation of Neural Stem Cells: The Influence of Homocysteine Treatment. Int. J. Mol. Sci. 2023, 24, 14161. https://doi.org/10.3390/ijms241814161
Petrović DJ, Jagečić D, Krasić J, Sinčić N, Mitrečić D. Effect of Fetal Bovine Serum or Basic Fibroblast Growth Factor on Cell Survival and the Proliferation of Neural Stem Cells: The Influence of Homocysteine Treatment. International Journal of Molecular Sciences. 2023; 24(18):14161. https://doi.org/10.3390/ijms241814161
Chicago/Turabian StylePetrović, Dražen Juraj, Denis Jagečić, Jure Krasić, Nino Sinčić, and Dinko Mitrečić. 2023. "Effect of Fetal Bovine Serum or Basic Fibroblast Growth Factor on Cell Survival and the Proliferation of Neural Stem Cells: The Influence of Homocysteine Treatment" International Journal of Molecular Sciences 24, no. 18: 14161. https://doi.org/10.3390/ijms241814161