
Whole-Body Vibration Prevents Neuronal, Neurochemical, and Behavioral Effects of Morphine Withdrawal in a Rat Model
 , , , , and
, , , , and 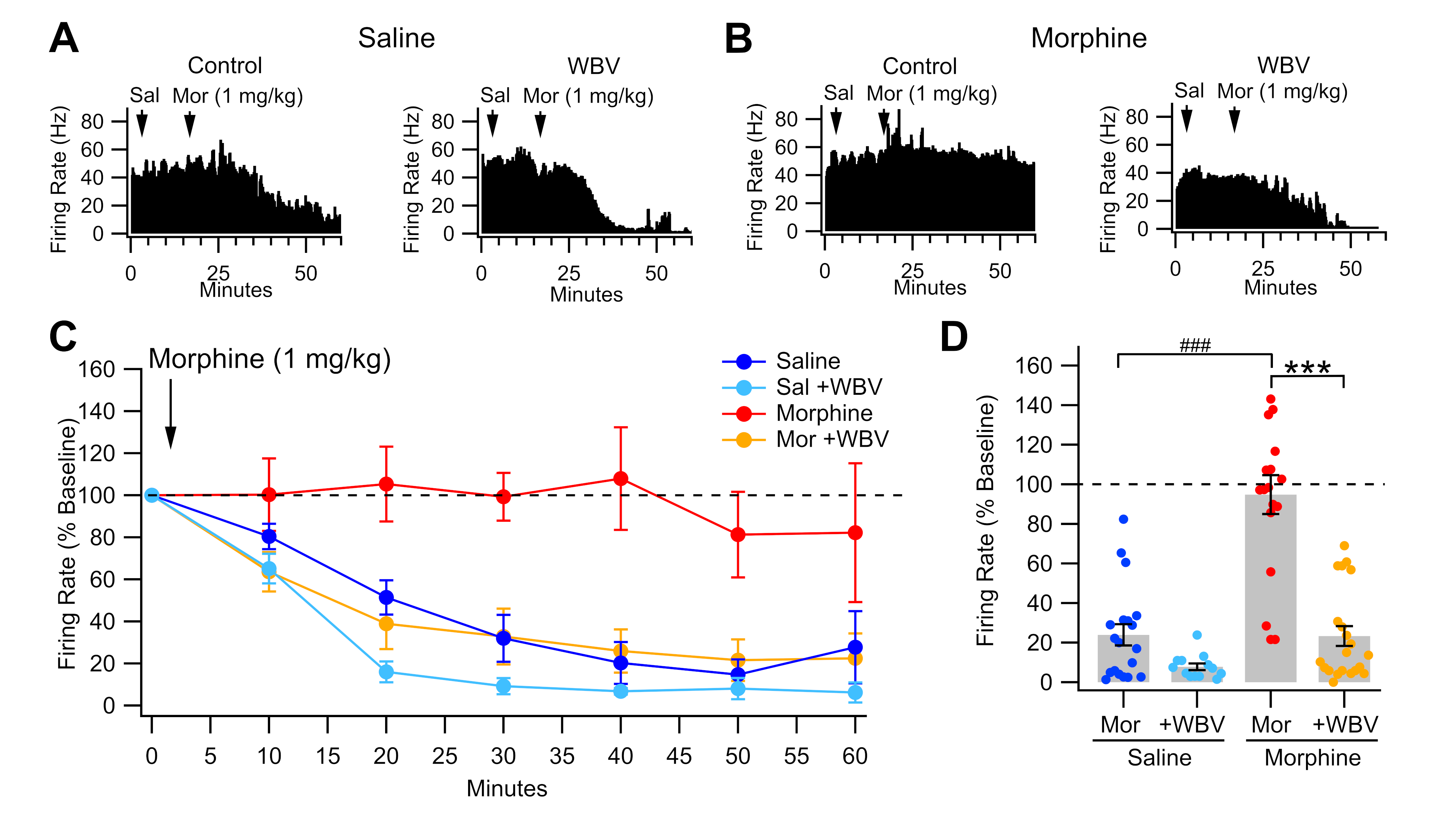{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of WBV on Morphine-Induced Changes to VTA GABA Neurons
2.2. Effect of WBV on Morphine-Induced Changes to NAc DA Release
2.3. Effect of WBV on Morphine-Induced Changes in NAc CIN DOR Expression
2.4. Effect of WBV on Anxiety Induced by Morphine Withdrawal
3. Discussion
4. Materials and Methods
4.1. Animal Subjects
4.2. Whole-Body Vibration Apparatus
4.3. Morphine Administration and Whole-Body Vibration
4.4. In Vivo Single Cell Recordings and Characterization of VTA GABA Neurons
4.5. In Vivo Microdialysis and High-Performance Liquid Chromatography of NAc DA
4.6. Immunohistochemistry and Confocal Microscopy
4.7. Elevated-Plus Maze
4.8. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Passik, S.D. Issues in long-term opioid therapy: Unmet needs, risks, and solutions. Mayo Clin. Proc. 2009, 84, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Christie, C.; Baker, C.; Cooper, R.; Kennedy, P.J.; Madras, B.; Bondi, P. The President’s Commission on Combating Drug Addiction and the Opioid Crisis; US Government Printing Office: Washington, DC, USA, 2017. [Google Scholar]
- Substance Abuse and Mental Health Services Administration. Results from the 2008 National Survey on Drug Use and Health: National Findings; Substance Abuse and Mental Health Services Administration: Rockville, MD, USA, 2009. [Google Scholar]
- Bell, J.; Strang, J. Medication Treatment of Opioid Use Disorder. Biol. Psychiatry 2020, 87, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Sofuoglu, M.; DeVito, E.E.; Carroll, K.M. Pharmacological and Behavioral Treatment of Opioid Use Disorder. Psychiatr. Res. Clin. Pract. 2019, 1, 4–15. [Google Scholar] [CrossRef]
- Koob, G.F. A role for brain stress systems in addiction. Neuron 2008, 59, 11–34. [Google Scholar] [CrossRef]
- Sinha, R. Chronic stress, drug use, and vulnerability to addiction. Ann. N. Y. Acad. Sci. 2008, 1141, 105–130. [Google Scholar] [CrossRef]
- Varghese, S.P.; Montalvo-Ortiz, J.L.; Csernansky, J.G.; Eiger, R.I.; Herrold, A.A.; Koola, M.M.; Dong, H. Early Life Stress as a Risk Factor for Substance use Disorders: Clinical and Neurobiological Substrates. Indian J. Psychol. Med. 2015, 37, 36–41. [Google Scholar] [CrossRef]
- McKendrick, G.; Garrett, H.; Jones, H.E.; McDevitt, D.S.; Sharma, S.; Silberman, Y.; Graziane, N.M. Ketamine Blocks Morphine-Induced Conditioned Place Preference and Anxiety-Like Behaviors in Mice. Front. Behav. Neurosci. 2020, 14, 75. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine and reward: The anhedonia hypothesis 30 years on. Neurotox. Res. 2008, 14, 169–183. [Google Scholar] [CrossRef]
- Ahn, S.; Phillips, A.G. Modulation by central and basolateral amygdalar nuclei of dopaminergic correlates of feeding to satiety in the rat nucleus accumbens and medial prefrontal cortex. J. Neurosci. 2002, 22, 10958–10965. [Google Scholar] [CrossRef]
- Ahn, S.; Phillips, A.G. Independent modulation of basal and feeding-evoked dopamine efflux in the nucleus accumbens and medial prefrontal cortex by the central and basolateral amygdalar nuclei in the rat. Neuroscience 2003, 116, 295–305. [Google Scholar] [CrossRef]
- Phillips, A.G.; Ahn, S.; Howland, J.G. Amygdalar control of the mesocorticolimbic dopamine system: Parallel pathways to motivated behavior. Neurosci. Biobehav. Rev. 2003, 27, 543–554. [Google Scholar] [CrossRef] [PubMed]
- Agmo, A.; Federman, I.; Navarro, V.; Padua, M.; Velazquez, G. Reward and reinforcement produced by drinking water: Role of opioids and dopamine receptor subtypes. Pharmacol. Biochem. Behav. 1993, 46, 183–194. [Google Scholar] [CrossRef]
- Agmo, A.; Galvan, A.; Talamantes, B. Reward and reinforcement produced by drinking sucrose: Two processes that may depend on different neurotransmitters. Pharmacol. Biochem. Behav. 1995, 52, 403–414. [Google Scholar] [CrossRef]
- Tzschentke, T.M. The medial prefrontal cortex as a part of the brain reward system. Amino Acids 2000, 19, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Brain reward circuitry: Insights from unsensed incentives. Neuron 2002, 36, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Gratton, A.; Wise, R.A. Brain stimulation reward in the lateral hypothalamic medial forebrain bundle: Mapping of boundaries and homogeneity. Brain Res. 1983, 274, 25–30. [Google Scholar] [CrossRef]
- Wise, R.A. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Nestler, E.J. Psychogenomics: Opportunities for understanding addiction. J. Neurosci. 2001, 21, 8324–8327. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Churchill, L.; Klitenick, M.A. The circuitry mediating the translation of motivational stimuli into adaptive motor responses. In Limbic Motor Circuits and Neuropsychiatry; Kalivas, P.W., Barnes, C.D., Eds.; CRC Press, Inc.: Boca Raton, FL, USA, 1993; pp. 237–287. [Google Scholar]
- Kalivas, P.W.; Volkow, N.D. The neural basis of addiction: A pathology of motivation and choice. Am. J. Psychiatry 2005, 162, 1403–1413. [Google Scholar] [CrossRef]
- Gallegos, R.A.; Criado, J.R.; Lee, R.S.; Henriksen, S.J.; Steffensen, S.C. Adaptive responses of GABAergic neurons in the ventral tegmental area to chronic ethanol. J. Pharmacol. Exp. Ther. 1999, 291, 1045–1053. [Google Scholar]
- Ludlow, K.H.; Bradley, K.D.; Allison, D.W.; Taylor, S.R.; Yorgason, J.T.; Hansen, D.M.; Walton, C.H.; Sudweeks, S.N.; Steffensen, S.C. Acute and chronic ethanol modulate dopamine D2-subtype receptor responses in ventral tegmental area GABA neurons. Alcohol. Clin. Exp. Res. 2009, 33, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Stobbs, S.H.; Ohran, A.J.; Lassen, M.B.; Allison, D.W.; Brown, J.E.; Steffensen, S.C. Ethanol suppression of ventral tegmental area GABA neuron electrical transmission involves NMDA receptors. J. Pharmacol. Exp. Ther. 2004, 311, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, S.C.; Taylor, S.R.; Horton, M.L.; Barber, E.N.; Lyle, L.T.; Stobbs, S.H.; Allison, D.W. Cocaine disinhibits dopamine neurons in the ventral tegmental area via use-dependent blockade of GABA neuron voltage-sensitive sodium channels. Eur. J. Neurosci. 2008, 28, 2028–2040. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.T.; Tan, K.R.; O’Connor, E.C.; Nikonenko, I.; Muller, D.; Luscher, C. Ventral tegmental area GABA projections pause accumbal cholinergic interneurons to enhance associative learning. Nature 2012, 492, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.R.; Yvon, C.; Turiault, M.; Mirzabekov, J.J.; Doehner, J.; Labouebe, G.; Deisseroth, K.; Tye, K.M.; Luscher, C. GABA neurons of the VTA drive conditioned place aversion. Neuron 2012, 73, 1173–1183. [Google Scholar] [CrossRef]
- Ohta, Y.; Murakami, T.E.; Kawahara, M.; Haruta, M.; Takehara, H.; Tashiro, H.; Sasagawa, K.; Ohta, J.; Akay, M.; Akay, Y.M. Investigating the Influence of GABA Neurons on Dopamine Neurons in the Ventral Tegmental Area Using Optogenetic Techniques. Int. J. Mol. Sci. 2022, 23, 1114. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Stobbs, S.H.; Colago, E.E.; Lee, R.S.; Koob, G.F.; Gallegos, R.A.; Henriksen, S.J. Contingent and non-contingent effects of heroin on mu-opioid receptor-containing ventral tegmental area GABA neurons. Exp. Neurol. 2006, 202, 139–151. [Google Scholar] [CrossRef]
- Carboni, E.; Imperato, A.; Perezzani, L.; Di Chiara, G. Amphetamine, cocaine, phencyclidine and nomifensine increase extracellular dopamine concentrations preferentially in the nucleus accumbens of freely moving rats. Neuroscience 1989, 28, 653–661. [Google Scholar] [CrossRef]
- Yoshimoto, K.; McBride, W.J.; Lumeng, L.; Li, T.K. Alcohol stimulates the release of dopamine and serotonin in the nucleus accumbens. Alcohol 1992, 9, 17–22. [Google Scholar] [CrossRef]
- Bocklisch, C.; Pascoli, V.; Wong, J.C.; House, D.R.; Yvon, C.; de Roo, M.; Tan, K.R.; Luscher, C. Cocaine disinhibits dopamine neurons by potentiation of GABA transmission in the ventral tegmental area. Science 2013, 341, 1521–1525. [Google Scholar] [CrossRef]
- Bonci, A.; Williams, J.T. Increased probability of GABA release during withdrawal from morphine. J. Neurosci. 1997, 17, 796–803. [Google Scholar] [CrossRef] [PubMed]
- Koeltzow, T.E.; White, F.J. Behavioral depression during cocaine withdrawal is associated with decreased spontaneous activity of ventral tegmental area dopamine neurons. Behav. Neurosci. 2003, 117, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Karkhanis, A.N.; Huggins, K.N.; Rose, J.H.; Jones, S.R. Switch from excitatory to inhibitory actions of ethanol on dopamine levels after chronic exposure: Role of kappa opioid receptors. Neuropharmacology 2016, 110, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Maisonneuve, I.M.; Ho, A.; Kreek, M.J. Chronic administration of a cocaine “binge” alters basal extracellular levels in male rats: An in vivo microdialysis study. J. Pharmacol. Exp. Ther. 1995, 272, 652–657. [Google Scholar] [PubMed]
- Rose, J.H.; Karkhanis, A.N.; Chen, R.; Gioia, D.; Lopez, M.F.; Becker, H.C.; McCool, B.A.; Jones, S.R. Supersensitive kappa opioid receptors promotes ethanol withdrawal-related behaviors and reduce dopamine signaling in the nucleus accumbens. Int. J. Neuropsychopharmacol. 2016, 19, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lyness, W.H.; Smith, F.L. Influence of dopaminergic and serotonergic neurons on intravenous ethanol self-administration in the rat. Pharmacol. Biochem. Behav. 1992, 42, 187–192. [Google Scholar] [CrossRef]
- Nelson, A.C.; Williams, S.B.; Pistorius, S.S.; Park, H.J.; Woodward, T.J.; Payne, A.J.; Obray, J.D.; Shin, S.I.; Mabey, J.K.; Steffensen, S.C. Ventral tegmental area GABA neurons are resistant to GABA(A) receptor-mediated inhibition during ethanol withdrawal. Front. Neurosci. 2018, 12, 131. [Google Scholar] [CrossRef]
- Ting, A.K.R.; van der Kooy, D. The neurobiology of opiate motivation. Cold Spring Harb. Perspect. Med. 2012, 2, 1–15. [Google Scholar] [CrossRef]
- Ting, A.K.R.; Vargas-Perez, H.; Mabey, J.K.; Shin, S.I.; Steffensen, S.C.; van der Kooy, D. Ventral tegmental area GABA neurons and opiate motivation. Psychopharmacology 2013, 227, 697–709. [Google Scholar] [CrossRef]
- Vargas-Perez, H.; Bahi, A.; Bufalino, M.R.; Ting, A.K.R.; Maal-Bared, G.; Lam, J.; Fahmy, A.; Clarke, L.; Blanchard, J.K.; Larsen, B.R.; et al. BDNF signaling in the VTA links the drug-dependent state to drug withdrawal aversions. J. Neurosci. 2014, 34, 7899–7909. [Google Scholar] [CrossRef]
- Vargas-Perez, H.; Ting, A.K.R.; Walton, C.H.; Hansen, D.M.; Razavi, R.; Clarke, L.; Bufalino, M.R.; Allison, D.W.; Steffensen, S.C.; van der Kooy, D. Ventral tegmental area BDNF induces an opiate-dependent-like reward state in naive rats. Science 2009, 324, 1732–1734. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.; Ryu, Y.; Gwak, Y.S.; Kim, N.J.; Kim, J.M.; Lee, J.Y.; Kim, S.A.; Lee, B.H.; Steffensen, S.C.; Jang, E.Y.; et al. Spinal pathways involved in somatosensory inhibition of the psychomotor actions of cocaine. Sci. Rep. 2017, 7, 5359. [Google Scholar] [CrossRef] [PubMed]
- Jin, W.; Kim, M.S.; Jang, E.Y.; Lee, J.Y.; Lee, J.G.; Kim, H.Y.; Yoon, S.S.; Lee, B.H.; Chang, S.; Kim, J.H.; et al. Acupuncture reduces relapse to cocaine-seeking behavior via activation of GABA neurons in the ventral tegmental area. Addict. Biol. 2018, 23, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.A.; Lee, B.H.; Bae, J.H.; Kim, K.J.; Steffensen, S.C.; Ryu, Y.H.; Leem, J.W.; Yang, C.H.; Kim, H.Y. Peripheral afferent mechanisms underlying acupuncture inhibition of cocaine behavioral effects in rats. PLoS ONE 2013, 8, e81018. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.S.; Yang, E.J.; Lee, B.H.; Jang, E.Y.; Kim, H.Y.; Choi, S.M.; Steffensen, S.C.; Yang, C.H. Effects of acupuncture on stress-induced relapse to cocaine-seeking in rats. Psychopharmacology 2012, 222, 303–311. [Google Scholar] [CrossRef]
- Yang, C.H.; Yoon, S.S.; Hansen, D.M.; Wilcox, J.D.; Blumell, B.R.; Park, J.J.; Steffensen, S.C. Acupuncture inhibits GABA neuron activity in the ventral tegmental area and reduces ethanol self-administration. Alcohol. Clin. Exp. Res. 2010, 34, 2137–2146. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.J.; Ryu, Y.; Lee, B.H.; Chang, S.; Fan, Y.; Gwak, Y.S.; Yang, C.H.; Bills, K.B.; Steffensen, S.C.; Koo, J.S.; et al. Acupuncture inhibition of methamphetamine-induced behaviors, dopamine release and hyperthermia in the nucleus accumbens: Mediation of group II mGluR. Addict. Biol. 2019, 24, 206–217. [Google Scholar] [CrossRef]
- Chang, S.; Fan, Y.; Lee, S.M.; Ryu, Y.; Lee, B.H.; Kim, S.C.; Bills, K.B.; Steffensen, S.C.; Yang, C.H.; Kim, H.Y. Acupuncture reduces cocaine psychomotor responses by activating the rostromedial tegmental nucleus. Addict. Biol. 2021, 26, e13052. [Google Scholar] [CrossRef]
- Yang, C.H.; Lee, B.H.; Sohn, S.H. A possible mechanism underlying the effectiveness of acupuncture in the treatment of drug addiction. Evid.-Based Complement. Altern. Med. 2008, 5, 463284. [Google Scholar] [CrossRef]
- Hipolito, L.; Sanchez-Catalan, M.J.; Zanolini, I.; Polache, A.; Granero, L. Shell/core differences in mu- and delta-opioid receptor modulation of dopamine efflux in nucleus accumbens. Neuropharmacology 2008, 55, 183–189. [Google Scholar] [CrossRef]
- Oroszi, T.; Geerts, E.; de Boer, S.F.; Schoemaker, R.G.; van der Zee, E.A.; Nyakas, C. Whole body vibration improves spatial memory, anxiety-like behavior, and motor performance in aged male and female rats. Front. Aging Neurosci. 2022, 13, 801828. [Google Scholar] [CrossRef] [PubMed]
- Peng, G.; Yang, L.; Wu, C.Y.; Zhang, L.L.; Wu, C.Y.; Li, F.; Shi, H.W.; Hou, J.; Zhang, L.M.; Ma, X.; et al. Whole body vibration training improves depression-like behaviors in a rat chronic restraint stress model. Neurochem. Int. 2021, 142, 104926. [Google Scholar] [CrossRef] [PubMed]
- Wunram, H.L.; Hamacher, S.; Hellmich, M.; Volk, M.; Jänicke, F.; Reinhard, F.; Bloch, W.; Zimmer, P.; Graf, C.; Schönau, E.; et al. Whole body vibration added to treatment as usual is effective in adolescents with depression: A partly randomized, three-armed clinical trial in inpatients. Eur. Child Adolesc. Psychiatry 2018, 27, 645–662. [Google Scholar] [CrossRef] [PubMed]
- McLain, R.F.; Raiszadeh, K. Mechanoreceptor endings of the cervical, thoracic, and lumbar spine. Iowa Orthop. J. 1995, 15, 147–155. [Google Scholar] [PubMed]
- Bills, K.B.; Clarke, T.; Major, G.H.; Jacobson, C.B.; Blotter, J.D.; Feland, J.B.; Steffensen, S.C. Targeted subcutaneous vibration with single-neuron electrophysiology as a novel method for understanding the central effects of peripheral vibrational therapy in a rodent model. Dose-Response 2019, 17, 1559325818825172. [Google Scholar] [CrossRef] [PubMed]
- Bills, K.B.; Obray, J.D.; Clarke, T.; Parsons, M.; Brundage, J.; Yang, C.H.; Kim, H.Y.; Yorgason, J.T.; Blotter, J.D.; Steffensen, S.C. Mechanical stimulation of cervical vertebrae modulates the discharge activity of ventral tegmental area neurons and dopamine release in the nucleus accumbens. Brain Stimul. 2020, 13, 403–411. [Google Scholar] [CrossRef]
- Bills, K.B.; Otteson, D.Z.; Jones, G.C.; Brundage, J.N.; Baldwin, E.K.; Small, C.A.; Kim, H.Y.; Yorgason, J.T.; Blotter, J.D.; Steffensen, S.C. Mechanical stimulation alters chronic ethanol-induced changes to VTA GABA neurons, NAc DA release and measures of withdrawal. Int. J. Mol. Sci. 2022, 23, 12630. [Google Scholar] [CrossRef]
- Simola, N.; Granon, S. Ultrasonic vocalizations as a tool in studying emotional states in rodent models of social behavior and brain disease. Neuropharmacology 2019, 159, 107420. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef]
- Xia, Y.; Driscoll, J.R.; Wilbrecht, L.; Margolis, E.B.; Fields, H.L.; Hjelmstad, G.O. Nucleus accumbens medium spiny neurons target non-dopaminergic neurons in the ventral tegmental area. J. Neurosci. 2011, 31, 7811–7816. [Google Scholar] [CrossRef]
- Jalabert, M.; Bourdy, R.; Courtin, J.; Veinante, P.; Manzoni, O.J.; Barrot, M.; Georges, F. Neuronal circuits underlying acute morphine action on dopamine neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 16446–16450. [Google Scholar] [CrossRef]
- Georges, F.; Moine, C.L.; Aston-Jones, G. No effect of morphine on ventral tegmental dopamine neurons during withdrawal. J. Neurosci. 2006, 26, 5720–5726. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, M.A.; Welch, S.P. μ-Opioid receptor down-regulation and cAMP-dependent protein kinase phosphorylation in a mouse model of chronic morphine tolerance. Mol. Brain Res. 1998, 55, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhang, Y.; Kalyuzhny, A.E.; Pan, Z.Z. Emergence of functional δ-opioid receptors induced by long-term treatment with morphine. Mol. Pharmacol. 2006, 69, 1137–1145. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhang, L.; Law, P.-Y.; Wei, L.-N.; Loh, H.H. Long-term morphine treatment decreases the association of μ-opioid receptor (MOR1) mRNA with polysomes through miRNA23b. Mol. Pharmacol. 2009, 75, 744–750. [Google Scholar] [CrossRef]
- Burgdorf, J.S.; Brudzynski, S.M.; Moskal, J.R. Using rat ultrasonic vocalization to study the neurobiology of emotion: From basic science to the development of novel therapeutics for affective disorders. Curr. Opin. Neurobiol. 2020, 60, 192–200. [Google Scholar] [CrossRef]
- Brudzynski, S.M. The ascending mesolimbic cholinergic system—A specific division of the reticular activating system involved in the initiation of negative emotional states. J. Mol. Neurosci. 2014, 53, 436–445. [Google Scholar] [CrossRef]
- Maldonado, R.; Koob, G.F. Destruction of the locus coeruleus decreases physical signs of opiate withdrawal. Brain Res. 1993, 605, 128–138. [Google Scholar] [CrossRef]
- Steffensen, S.C.; Svingos, A.L.; Pickel, V.M.; Henriksen, S.J. Electrophysiological characterization of GABAergic neurons in the ventral tegmental area. J. Neurosci. 1998, 18, 8003–8015. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, G.C.; Small, C.A.; Otteson, D.Z.; Hafen, C.W.; Breinholt, J.T.; Flora, P.D.; Burris, M.D.; Sant, D.W.; Ruchti, T.R.; Yorgason, J.T.; et al. Whole-Body Vibration Prevents Neuronal, Neurochemical, and Behavioral Effects of Morphine Withdrawal in a Rat Model. Int. J. Mol. Sci. 2023, 24, 14147. https://doi.org/10.3390/ijms241814147
Jones GC, Small CA, Otteson DZ, Hafen CW, Breinholt JT, Flora PD, Burris MD, Sant DW, Ruchti TR, Yorgason JT, et al. Whole-Body Vibration Prevents Neuronal, Neurochemical, and Behavioral Effects of Morphine Withdrawal in a Rat Model. International Journal of Molecular Sciences. 2023; 24(18):14147. https://doi.org/10.3390/ijms241814147
Chicago/Turabian StyleJones, Gavin C., Christina A. Small, Dallin Z. Otteson, Caylor W. Hafen, Jacob T. Breinholt, Paul D. Flora, Matthew D. Burris, David W. Sant, Tysum R. Ruchti, Jordan T. Yorgason, and et al. 2023. "Whole-Body Vibration Prevents Neuronal, Neurochemical, and Behavioral Effects of Morphine Withdrawal in a Rat Model" International Journal of Molecular Sciences 24, no. 18: 14147. https://doi.org/10.3390/ijms241814147