
Long-Term Effects of ART on the Health of the Offspring
 , and
, and
Abstract
:1. Introduction
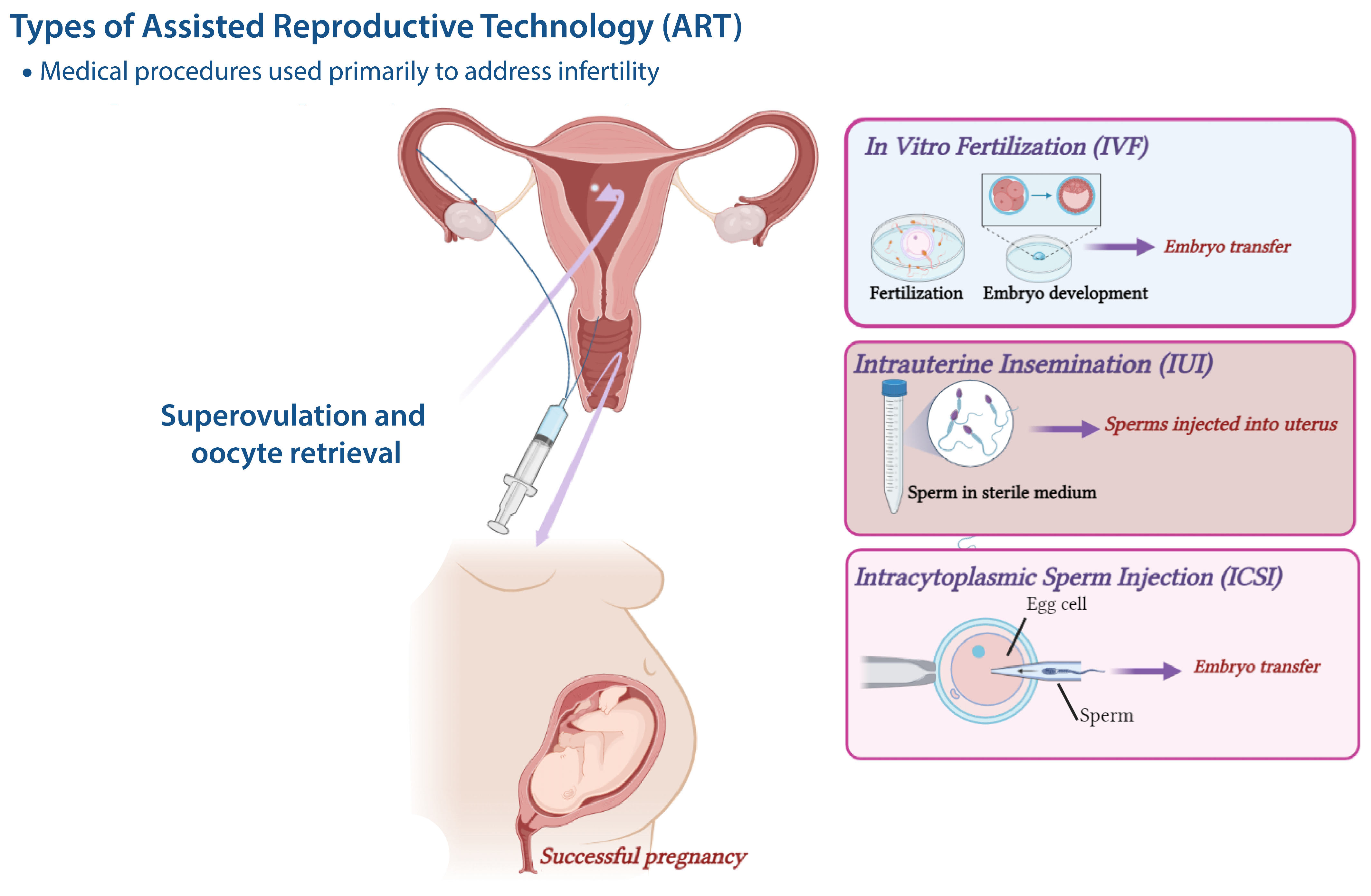
2. Assisted Reproductive Technologies (ART)
3. Preimplantation Stress and Development
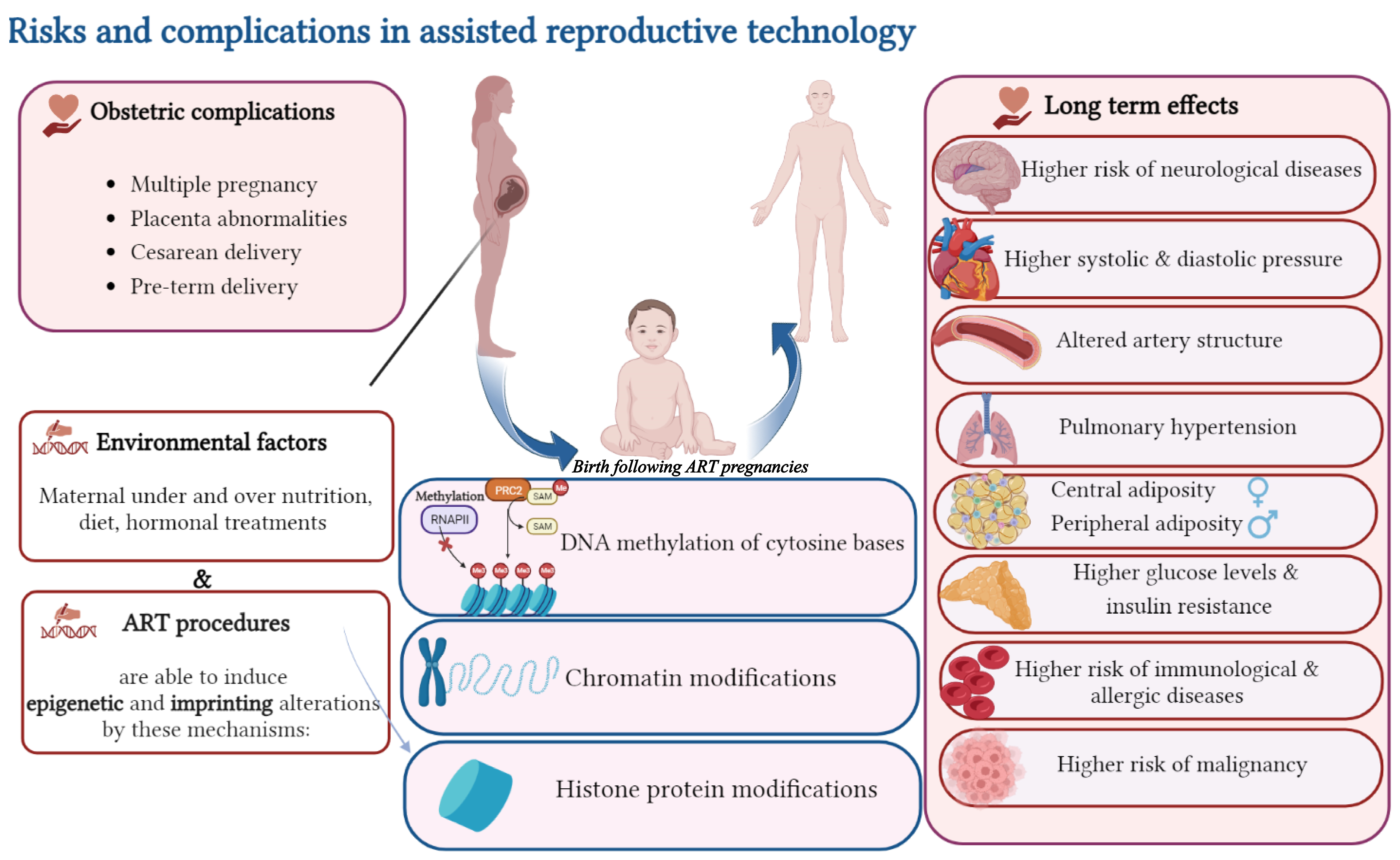
4. ART and Imprinting Disorders (IDs)
4.1. Effects of Cryopreservation and Vitrification on Epigenetic Alteration
4.2. Effects of Culture Media on Epigenetic Alteration
4.3. Effects of In Vitro Maturation (IVM) on Epigenetic Alteration
4.4. Effects of ICSI on Epigenetic Alteration
5. ART and Genomic Imprinting in the Placenta
6. ART and Underlying Parental Characteristics
7. ART and Congenital Malformations
8. ART and Neurological Disorders
9. ART and Cancer Risk
10. ART and Cardio Metabolic Disorders
11. ART and Altered Immune Functions
12. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fauser, B.C.; Devroey, P.; Diedrich, K.; Balaban, B.; Bonduelle, M.; Delemarre-van de Waal, H.A.; Estella, C.; Ezcurra, D.; Geraedts, J.P.; Howles, C.M.; et al. Health outcomes of children born after IVF/ICSI: A review of current expert opinion and literature. Reprod. Biomed. Online 2014, 28, 162–182. [Google Scholar] [CrossRef]
- Ahmadi, H.; Fathi, F.; Karimi, H.; Amidi, F.; Mehdinejadiani, S.; Moeini, A.; Bahram Rezai, M.; Hoseini, S.; Sobhani, A. Altered TH1, TH2, TH17 balance in assisted reproductive technology conceived mice. J. Reprod. Immunol. 2020, 139, 103117. [Google Scholar] [CrossRef]
- Hoorsan, H.; Mirmiran, P.; Chaichian, S.; Moradi, Y.; Hoorsan, R.; Jesmi, F. Congenital Malformations in Infants of Mothers Undergoing Assisted Reproductive Technologies: A Systematic Review and Meta-analysis Study. J. Prev. Med. Public Health 2017, 50, 347–360. [Google Scholar] [CrossRef]
- Simpson, J.L. Birth defects and assisted reproductive technologies. Semin. Fetal Neonatal Med. 2014, 19, 177–182. [Google Scholar] [CrossRef]
- Moll, A.C.; Imhof, S.M.; Cruysberg, J.R.; Schouten-van Meeteren, A.Y.; Boers, M.; van Leeuwen, F.E. Incidence of retinoblastoma in children born after in-vitro fertilisation. Lancet 2003, 361, 309–310. [Google Scholar] [CrossRef]
- Petridou, E.T.; Sergentanis, T.N.; Panagopoulou, P.; Moschovi, M.; Polychronopoulou, S.; Baka, M.; Pourtsidis, A.; Athanassiadou, F.; Kalmanti, M.; Sidi, V.; et al. In vitro fertilization and risk of childhood leukemia in Greece and Sweden. Pediatr. Blood Cancer 2012, 58, 930–936. [Google Scholar] [CrossRef]
- Massaro, P.A.; MacLellan, D.L.; Anderson, P.A.; Romao, R.L. Does intracytoplasmic sperm injection pose an increased risk of genitourinary congenital malformations in offspring compared to in vitro fertilization? A systematic review and meta-analysis. J. Urol. 2015, 193, 1837–1842. [Google Scholar] [CrossRef]
- Kamel, R.M. Assisted reproductive technology after the birth of louise brown. J. Reprod. Infertil. 2013, 14, 96–109. [Google Scholar] [CrossRef]
- Wang, L.Y.; Wang, N.; Le, F.; Li, L.; Lou, H.Y.; Liu, X.Z.; Zheng, Y.M.; Qian, Y.Q.; Chen, Y.L.; Jiang, X.H.; et al. Superovulation Induced Changes of Lipid Metabolism in Ovaries and Embryos and Its Probable Mechanism. PLoS ONE 2015, 10, e0132638. [Google Scholar] [CrossRef]
- Carpinello, O.J.; DeCherney, A.H.; Hill, M.J. Developmental Origins of Health and Disease: The History of the Barker Hypothesis and Assisted Reproductive Technology. Semin. Reprod. Med. 2018, 36, 177–182. [Google Scholar] [CrossRef]
- Rinaudo, P.; Schultz, R.M. Effects of embryo culture on global pattern of gene expression in preimplantation mouse embryos. Reproduction 2004, 128, 301–311. [Google Scholar] [CrossRef]
- Fernández-Gonzalez, R.; Moreira, P.; Bilbao, A.; Jiménez, A.; Pérez-Crespo, M.; Ramírez, M.A.; Rodríguez De Fonseca, F.; Pintado, B.; Gutiérrez-Adán, A. Long-term effect of in vitro culture of mouse embryos with serum on mRNA expression of imprinting genes, development, and behavior. Proc. Natl. Acad. Sci. USA 2004, 101, 5880–5885. [Google Scholar] [CrossRef] [PubMed]
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care, 2017. Hum. Reprod. 2017, 32, 1786–1801. [Google Scholar] [CrossRef]
- Cohen, J.; Trounson, A.; Dawson, K.; Jones, H.; Hazekamp, J.; Nygren, K.G.; Hamberger, L. The early days of IVF outside the UK. Hum. Reprod. Update 2005, 11, 439–459. [Google Scholar] [CrossRef]
- Lenz, S.; Lauritsen, J.G. Ultrasonically guided percutaneous aspiration of human follicles under local anesthesia: A new method of collecting oocytes for in vitro fertilization. Fertil. Steril. 1982, 38, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Trounson, A.; Mohr, L. Human pregnancy following cryopreservation, thawing and transfer of an eight-cell embryo. Nature 1983, 305, 707–709. [Google Scholar] [CrossRef]
- Trounson, A.; Peura, A.; Kirby, C. Ultrarapid freezing: A new low-cost and effective method of embryo cryopreservation. Fertil. Steril. 1987, 48, 843–850. [Google Scholar] [CrossRef]
- Patrizio, P.; Silber, S.; Ord, T.; Balmaceda, J.P.; Asch, R.H. Two births after microsurgical sperm aspiration in congenital absence of vas deferens. Lancet 1988, 2, 1364. [Google Scholar] [CrossRef]
- Palermo, G.; Joris, H.; Devroey, P.; Van Steirteghem, A.C. Pregnancies after intracytoplasmic injection of single spermatozoon into an oocyte. Lancet 1992, 340, 17–18. [Google Scholar] [CrossRef] [PubMed]
- Cha, K.Y.; Koo, J.J.; Ko, J.J.; Choi, D.H.; Han, S.Y.; Yoon, T.K. Pregnancy after in vitro fertilization of human follicular oocytes collected from nonstimulated cycles, their culture in vitro and their transfer in a donor oocyte program. Fertil. Steril. 1991, 55, 109–113. [Google Scholar] [CrossRef]
- Germond, M.; Dessole, S.; Senn, A.; Loumaye, E.; Howles, C.; Beltrami, V. Successful in-vitro fertilisation and embryo transfer after treatment with recombinant human FSH. Lancet 1992, 339, 1170. [Google Scholar]
- Xu, K.; Shi, Z.M.; Veeck, L.L.; Hughes, M.R.; Rosenwaks, Z. First unaffected pregnancy using preimplantation genetic diagnosis for sickle cell anemia. JAMA 1999, 281, 1701–1706. [Google Scholar] [CrossRef]
- Boer, K.A.; McArthur, S.; Murray, C.; Jansen, R.P.S. O-54. First live birth following blastocyst biopsy and PGD analysis. Reprod. BioMed. Online 2002, 4, 35. [Google Scholar] [CrossRef]
- Barker, D.J. The developmental origins of adult disease. J. Am. Coll. Nutr. 2004, 23, 588s–595s. [Google Scholar] [CrossRef]
- Stanner, S.A.; Bulmer, K.; Andrès, C.; Lantseva, O.E.; Borodina, V.; Poteen, V.V.; Yudkin, J.S. Does malnutrition in utero determine diabetes and coronary heart disease in adulthood? Results from the Leningrad siege study, a cross sectional study. BMJ 1997, 315, 1342–1348. [Google Scholar] [CrossRef] [PubMed]
- Fleming, T.P.; Lucas, E.S.; Watkins, A.J.; Eckert, J.J. Adaptive responses of the embryo to maternal diet and consequences for post-implantation development. Reprod. Fertil. Dev. 2011, 24, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Watkins, A.J.; Ursell, E.; Panton, R.; Papenbrock, T.; Hollis, L.; Cunningham, C.; Wilkins, A.; Perry, V.H.; Sheth, B.; Kwong, W.Y.; et al. Adaptive responses by mouse early embryos to maternal diet protect fetal growth but predispose to adult onset disease. Biol. Reprod. 2008, 78, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Ecker, D.J.; Stein, P.; Xu, Z.; Williams, C.J.; Kopf, G.S.; Bilker, W.B.; Abel, T.; Schultz, R.M. Long-term effects of culture of preimplantation mouse embryos on behavior. Proc. Natl. Acad. Sci. USA 2004, 101, 1595–1600. [Google Scholar] [CrossRef]
- Kwong, W.Y.; Wild, A.E.; Roberts, P.; Willis, A.C.; Fleming, T.P. Maternal undernutrition during the preimplantation period of rat development causes blastocyst abnormalities and programming of postnatal hypertension. Development 2000, 127, 4195–4202. [Google Scholar] [CrossRef]
- Adamiak, S.J.; Mackie, K.; Watt, R.G.; Webb, R.; Sinclair, K.D. Impact of nutrition on oocyte quality: Cumulative effects of body composition and diet leading to hyperinsulinemia in cattle. Biol. Reprod. 2005, 73, 918–926. [Google Scholar] [CrossRef]
- Cockburn, K.; Rossant, J. Making the blastocyst: Lessons from the mouse. J. Clin. Investig. 2010, 120, 995–1003. [Google Scholar] [CrossRef]
- Dey, S.K.; Lim, H.; Das, S.K.; Reese, J.; Paria, B.C.; Daikoku, T.; Wang, H. Molecular cues to implantation. Endocr. Rev. 2004, 25, 341–373. [Google Scholar] [CrossRef]
- Biggers, J.D.; Whittingham, D.G.; Donahue, R.P. The pattern of energy metabolism in the mouse oöcyte and zygote. Proc. Natl. Acad. Sci. USA 1967, 58, 560–567. [Google Scholar] [CrossRef]
- Brinster, R.L.; Thomson, J.L. Development of eight-cell mouse embryos in vitro. Exp. Cell Res. 1966, 42, 308–315. [Google Scholar] [CrossRef]
- Lane, M.; Gardner, D.K. Lactate regulates pyruvate uptake and metabolism in the preimplantation mouse embryo. Biol. Reprod. 2000, 62, 16–22. [Google Scholar] [CrossRef]
- Lane, M.; Gardner, D.K. Ammonium induces aberrant blastocyst differentiation, metabolism, pH regulation, gene expression and subsequently alters fetal development in the mouse. Biol. Reprod. 2003, 69, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Edwards, L.J.; Williams, D.A.; Gardner, D.K. Intracellular pH of the mouse preimplantation embryo: Amino acids act as buffers of intracellular pH. Hum. Reprod. 1998, 13, 3441–3448. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.K.; Leese, H.J. The role of glucose and pyruvate transport in regulating nutrient utilization by preimplantation mouse embryos. Development 1988, 104, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Bermejo-Alvarez, P.; Rizos, D.; Lonergan, P.; Gutierrez-Adan, A. Transcriptional sexual dimorphism during preimplantation embryo development and its consequences for developmental competence and adult health and disease. Reproduction 2011, 141, 563–570. [Google Scholar] [CrossRef]
- Feuer, S.; Rinaudo, P. Preimplantation stress and development. Birth Defects Res. C Embryo Today 2012, 96, 299–314. [Google Scholar] [CrossRef]
- Nadal, A.; Ropero, A.B.; Laribi, O.; Maillet, M.; Fuentes, E.; Soria, B. Nongenomic actions of estrogens and xenoestrogens by binding at a plasma membrane receptor unrelated to estrogen receptor alpha and estrogen receptor beta. Proc. Natl. Acad. Sci. USA 2000, 97, 11603–11608. [Google Scholar] [CrossRef]
- Takai, Y.; Tsutsumi, O.; Ikezuki, Y.; Kamei, Y.; Osuga, Y.; Yano, T.; Taketan, Y. Preimplantation exposure to bisphenol A advances postnatal development. Reprod. Toxicol. 2001, 15, 71–74. [Google Scholar] [CrossRef]
- Hassa, H.; Gurer, F.; Tanir, H.M.; Kaya, M.; Gunduz, N.B.; Sariboyaci, A.E.; Bal, C. Effect of cigarette smoke and alpha-tocopherol (vitamin E) on fertilization, cleavage, and embryo development rates in mice: An experimental in vitro fertilization mice model study. Eur. J. Obstet. Gynecol. Reprod. Biol. 2007, 135, 177–182. [Google Scholar] [CrossRef]
- Polyzos, A.; Schmid, T.E.; Piña-Guzmán, B.; Quintanilla-Vega, B.; Marchetti, F. Differential sensitivity of male germ cells to mainstream and sidestream tobacco smoke in the mouse. Toxicol. Appl. Pharmacol. 2009, 237, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, M.; Horiuchi, T.; Yanagimachi, R. Effects of light on development of mammalian zygotes. Proc. Natl. Acad. Sci. USA 2007, 104, 14289–14293. [Google Scholar] [CrossRef]
- Gil, M.A.; Maside, C.; Cuello, C.; Parrilla, I.; Vazquez, J.M.; Roca, J.; Martinez, E.A. Effects of Hoechst 33,342 staining and ultraviolet irradiation on mitochondrial distribution and DNA copy number in porcine oocytes and preimplantation embryos. Mol. Reprod. Dev. 2012, 79, 651–663. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Saka, N.; Takahashi, H.; Kanai, Y.; Schultz, R.M.; Okano, A. Assessment of DNA damage in individual hamster embryos by comet assay. Mol. Reprod. Dev. 1999, 54, 1–7. [Google Scholar] [CrossRef]
- Oh, S.J.; Gong, S.P.; Lee, S.T.; Lee, E.J.; Lim, J.M. Light intensity and wavelength during embryo manipulation are important factors for maintaining viability of preimplantation embryos in vitro. Fertil. Steril. 2007, 88, 1150–1157. [Google Scholar] [CrossRef]
- Bognar, Z.; Csabai, T.J.; Pallinger, E.; Balassa, T.; Farkas, N.; Schmidt, J.; Görgey, E.; Berta, G.; Szekeres-Bartho, J.; Bodis, J. The effect of light exposure on the cleavage rate and implantation capacity of preimplantation murine embryos. J. Reprod. Immunol. 2019, 132, 21–28. [Google Scholar] [CrossRef]
- Bódis, J.; Gödöny, K.; Várnagy, Á.; Kovács, K.; Koppán, M.; Nagy, B.; Erostyák, J.; Herczeg, R.; Szekeres-Barthó, J.; Gyenesei, A.; et al. How to Reduce the Potential Harmful Effects of Light on Blastocyst Development during IVF. Med. Princ. Pract. 2020, 29, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Delle Piane, L.; Lin, W.; Liu, X.; Donjacour, A.; Minasi, P.; Revelli, A.; Maltepe, E.; Rinaudo, P.F. Effect of the method of conception and embryo transfer procedure on mid-gestation placenta and fetal development in an IVF mouse model. Hum. Reprod. 2010, 25, 2039–2046. [Google Scholar] [CrossRef]
- Giritharan, G.; Li, M.W.; Di Sebastiano, F.; Esteban, F.J.; Horcajadas, J.A.; Lloyd, K.C.; Donjacour, A.; Maltepe, E.; Rinaudo, P.F. Effect of ICSI on gene expression and development of mouse preimplantation embryos. Hum. Reprod. 2010, 25, 3012–3024. [Google Scholar] [CrossRef]
- Valenzuela, O.A.; Couturier-Tarrade, A.; Choi, Y.H.; Aubrière, M.C.; Ritthaler, J.; Chavatte-Palmer, P.; Hinrichs, K. Impact of equine assisted reproductive technologies (standard embryo transfer or intracytoplasmic sperm injection (ICSI) with in vitro culture and embryo transfer) on placenta and foal morphometry and placental gene expression. Reprod. Fertil. Dev. 2018, 30, 371–379. [Google Scholar] [CrossRef]
- Giritharan, G.; Delle Piane, L.; Donjacour, A.; Esteban, F.J.; Horcajadas, J.A.; Maltepe, E.; Rinaudo, P. In vitro culture of mouse embryos reduces differential gene expression between inner cell mass and trophectoderm. Reprod. Sci. 2012, 19, 243–252. [Google Scholar] [CrossRef]
- Kleijkers, S.H.; Eijssen, L.M.; Coonen, E.; Derhaag, J.G.; Mantikou, E.; Jonker, M.J.; Mastenbroek, S.; Repping, S.; Evers, J.L.; Dumoulin, J.C.; et al. Differences in gene expression profiles between human preimplantation embryos cultured in two different IVF culture media. Hum. Reprod. 2015, 30, 2303–2311. [Google Scholar] [CrossRef]
- Rinaudo, P.F.; Giritharan, G.; Talbi, S.; Dobson, A.T.; Schultz, R.M. Effects of oxygen tension on gene expression in preimplantation mouse embryos. Fertil. Steril. 2006, 86, 1265.e1–1265.e36. [Google Scholar] [CrossRef]
- Leese, H.J.; Baumann, C.G.; Brison, D.R.; McEvoy, T.G.; Sturmey, R.G. Metabolism of the viable mammalian embryo: Quietness revisited. Mol. Hum. Reprod. 2008, 14, 667–672. [Google Scholar] [CrossRef]
- Consensus Group, C. ‘There is only one thing that is truly important in an IVF laboratory: Everything’ Cairo Consensus Guidelines on IVF Culture Conditions. Reprod. Biomed. Online 2020, 40, 33–60. [Google Scholar] [CrossRef]
- Khodavirdilou, R.; Pournaghi, M.; Oghbaei, H.; Rastgar Rezaei, Y.; Javid, F.; Khodavirdilou, L.; Shakibfar, F.; Latifi, Z.; Hakimi, P.; Nouri, M.; et al. Toxic effect of light on oocyte and pre-implantation embryo: A systematic review. Arch. Toxicol. 2021, 95, 3161–3169. [Google Scholar] [CrossRef]
- Kolahi, K.S.; Donjacour, A.; Liu, X.; Lin, W.; Simbulan, R.K.; Bloise, E.; Maltepe, E.; Rinaudo, P. Effect of substrate stiffness on early mouse embryo development. PLoS ONE 2012, 7, e41717. [Google Scholar] [CrossRef]
- Argyle, C.E.; Harper, J.C.; Davies, M.C. Oocyte cryopreservation: Where are we now? Hum. Reprod. Update 2016, 22, 440–449. [Google Scholar] [CrossRef] [PubMed]
- Bielanski, A.; Vajta, G. Risk of contamination of germplasm during cryopreservation and cryobanking in IVF units. Hum. Reprod. 2009, 24, 2457–2467. [Google Scholar] [CrossRef] [PubMed]
- Lavara, R.; Baselga, M.; Marco-Jiménez, F.; Vicente, J.S. Long-term and transgenerational effects of cryopreservation on rabbit embryos. Theriogenology 2014, 81, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Grissom, N.M.; George, R.; Reyes, T.M. Suboptimal nutrition in early life affects the inflammatory gene expression profile and behavioral responses to stressors. Brain Behav. Immun. 2017, 63, 115–126. [Google Scholar] [CrossRef]
- Kawwass, J.F.; Badell, M.L. Maternal and Fetal Risk Associated with Assisted Reproductive Technology. Obstet. Gynecol. 2018, 132, 763–772. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J.; Rumbold, A.R.; Marino, J.L.; Willson, K.; Giles, L.C.; Whitrow, M.J.; Scheil, W.; Moran, L.J.; Thompson, J.G.; Lane, M.; et al. Maternal factors and the risk of birth defects after IVF and ICSI: A whole of population cohort study. BJOG 2017, 124, 1537–1544. [Google Scholar] [CrossRef]
- Cabry, R.; Merviel, P.; Madkour, A.; Lefranc, E.; Scheffler, F.; Desailloud, R.; Bach, V.; Benkhalifa, M. The impact of endocrine disruptor chemicals on oocyte/embryo and clinical outcomes in IVF. Endocr. Connect. 2020, 9, R134–R142. [Google Scholar] [CrossRef]
- Krisher, R.L. In vivo and in vitro environmental effects on mammalian oocyte quality. Annu. Rev. Anim. Biosci. 2013, 1, 393–417. [Google Scholar] [CrossRef]
- Walters, E.; Edwards, R.G. On a fallacious invocation of the Barker hypothesis of anomalies in newborn rats due to mothers’ food restriction in preimplantation phases. Reprod. Biomed. Online 2003, 7, 580–582. [Google Scholar] [CrossRef]
- Neitzke, U.; Harder, T.; Schellong, K.; Melchior, K.; Ziska, T.; Rodekamp, E.; Dudenhausen, J.W.; Plagemann, A. Intrauterine growth restriction in a rodent model and developmental programming of the metabolic syndrome: A critical appraisal of the experimental evidence. Placenta 2008, 29, 246–254. [Google Scholar] [CrossRef]
- Jansson, T.; Lambert, G.W. Effect of intrauterine growth restriction on blood pressure, glucose tolerance and sympathetic nervous system activity in the rat at 3-4 months of age. J. Hypertens. 1999, 17, 1239–1248. [Google Scholar] [CrossRef]
- Harlow, G.M.; Quinn, P. Development of preimplantation mouse embryos in vivo and in vitro. Aust. J. Biol. Sci. 1982, 35, 187–193. [Google Scholar] [CrossRef]
- Schwarzer, C.; Esteves, T.C.; Araúzo-Bravo, M.J.; Le Gac, S.; Nordhoff, V.; Schlatt, S.; Boiani, M. ART culture conditions change the probability of mouse embryo gestation through defined cellular and molecular responses. Hum. Reprod. 2012, 27, 2627–2640. [Google Scholar] [CrossRef]
- Angiolini, E.; Fowden, A.; Coan, P.; Sandovici, I.; Smith, P.; Dean, W.; Burton, G.; Tycko, B.; Reik, W.; Sibley, C.; et al. Regulation of placental efficiency for nutrient transport by imprinted genes. Placenta 2006, 27 (Suppl. A), S98–S102. [Google Scholar] [CrossRef]
- Bloise, E.; Lin, W.; Liu, X.; Simbulan, R.; Kolahi, K.S.; Petraglia, F.; Maltepe, E.; Donjacour, A.; Rinaudo, P. Impaired placental nutrient transport in mice generated by in vitro fertilization. Endocrinology 2012, 153, 3457–3467. [Google Scholar] [CrossRef]
- Fernández-Gonzalez, R.; Moreira, P.N.; Pérez-Crespo, M.; Sánchez-Martín, M.; Ramirez, M.A.; Pericuesta, E.; Bilbao, A.; Bermejo-Alvarez, P.; de Dios Hourcade, J.; de Fonseca, F.R.; et al. Long-term effects of mouse intracytoplasmic sperm injection with DNA-fragmented sperm on health and behavior of adult offspring. Biol. Reprod. 2008, 78, 761–772. [Google Scholar] [CrossRef]
- Scott, K.A.; Yamazaki, Y.; Yamamoto, M.; Lin, Y.; Melhorn, S.J.; Krause, E.G.; Woods, S.C.; Yanagimachi, R.; Sakai, R.R.; Tamashiro, K.L. Glucose parameters are altered in mouse offspring produced by assisted reproductive technologies and somatic cell nuclear transfer. Biol. Reprod. 2010, 83, 220–227. [Google Scholar] [CrossRef]
- Watkins, A.J.; Platt, D.; Papenbrock, T.; Wilkins, A.; Eckert, J.J.; Kwong, W.Y.; Osmond, C.; Hanson, M.; Fleming, T.P. Mouse embryo culture induces changes in postnatal phenotype including raised systolic blood pressure. Proc. Natl. Acad. Sci. USA 2007, 104, 5449–5454. [Google Scholar] [CrossRef]
- Wu, C.; Morris, J.R. Genes, genetics, and epigenetics: A correspondence. Science 2001, 293, 1103–1105. [Google Scholar] [CrossRef]
- Santos, F.; Dean, W. Epigenetic reprogramming during early development in mammals. Reproduction 2004, 127, 643–651. [Google Scholar] [CrossRef]
- Kiefer, J.C. Epigenetics in development. Dev. Dyn. 2007, 236, 1144–1156. [Google Scholar] [CrossRef] [PubMed]
- Henningsen, A.A.; Gissler, M.; Rasmussen, S.; Opdahl, S.; Wennerholm, U.B.; Spangsmose, A.L.; Tiitinen, A.; Bergh, C.; Romundstad, L.B.; Laivuori, H.; et al. Imprinting disorders in children born after ART: A Nordic study from the CoNARTaS group. Hum. Reprod. 2020, 35, 1178–1184. [Google Scholar] [CrossRef]
- Eroglu, A.; Layman, L.C. Role of ART in imprinting disorders. Semin. Reprod. Med. 2012, 30, 92–104. [Google Scholar] [CrossRef] [PubMed]
- Owen, C.M.; Segars, J.H., Jr. Imprinting disorders and assisted reproductive technology. Semin. Reprod. Med. 2009, 27, 417–428. [Google Scholar] [CrossRef]
- Cortessis, V.K.; Azadian, M.; Buxbaum, J.; Sanogo, F.; Song, A.Y.; Sriprasert, I.; Wei, P.C.; Yu, J.; Chung, K.; Siegmund, K.D. Comprehensive meta-analysis reveals association between multiple imprinting disorders and conception by assisted reproductive technology. J. Assist. Reprod. Genet. 2018, 35, 943–952. [Google Scholar] [CrossRef]
- Amor, D.J.; Halliday, J. A review of known imprinting syndromes and their association with assisted reproduction technologies. Hum. Reprod. 2008, 23, 2826–2834. [Google Scholar] [CrossRef] [PubMed]
- Horánszky, A.; Becker, J.L.; Zana, M.; Ferguson-Smith, A.C.; Dinnyés, A. Epigenetic Mechanisms of ART-Related Imprinting Disorders: Lessons From iPSC and Mouse Models. Genes 2021, 12, 1704. [Google Scholar] [CrossRef] [PubMed]
- Chi, F.; Zhao, M.; Li, K.; Lin, A.Q.; Li, Y.; Teng, X. DNA methylation status of imprinted H19 and KvDMR1 genes in human placentas after conception using assisted reproductive technology. Ann. Transl. Med. 2020, 8, 854. [Google Scholar] [CrossRef]
- Lazaraviciute, G.; Kauser, M.; Bhattacharya, S.; Haggarty, P.; Bhattacharya, S. A systematic review and meta-analysis of DNA methylation levels and imprinting disorders in children conceived by IVF/ICSI compared with children conceived spontaneously. Hum. Reprod. Update 2014, 20, 840–852. [Google Scholar] [CrossRef]
- Yu, B.; Smith, T.H.; Battle, S.L.; Ferrell, S.; Hawkins, R.D. Superovulation alters global DNA methylation in early mouse embryo development. Epigenetics 2019, 14, 780–790. [Google Scholar] [CrossRef]
- Chen, X.; Huang, Y.; Huang, H.; Guan, Y.; Li, M.; Jiang, X.; Yu, M.; Yang, X. Effects of superovulation, in vitro fertilization, and oocyte in vitro maturation on imprinted gene Grb10 in mouse blastocysts. Arch. Gynecol. Obstet. 2018, 298, 1219–1227. [Google Scholar] [CrossRef]
- Sato, A.; Otsu, E.; Negishi, H.; Utsunomiya, T.; Arima, T. Aberrant DNA methylation of imprinted loci in superovulated oocytes. Hum. Reprod. 2007, 22, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Fortier, A.L.; Lopes, F.L.; Darricarrère, N.; Martel, J.; Trasler, J.M. Superovulation alters the expression of imprinted genes in the midgestation mouse placenta. Hum. Mol. Genet. 2008, 17, 1653–1665. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wei, Q.; Deng, H.; Li, G.; Ma, L.; Zeng, H. Negative regulation of Grb10 Interacting GYF Protein 2 on insulin-like growth factor-1 receptor signaling pathway caused diabetic mice cognitive impairment. PLoS ONE 2014, 9, e108559. [Google Scholar] [CrossRef]
- Chaudhry, M.; Wang, X.; Bamne, M.N.; Hasnain, S.; Demirci, F.Y.; Lopez, O.L.; Kamboh, M.I. Genetic variation in imprinted genes is associated with risk of late-onset Alzheimer’s disease. J. Alzheimers Dis. 2015, 44, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Bonakdar, E.; Edriss, M.A.; Bakhtari, A.; Jafarpour, F.; Asgari, V.; Hosseini, S.M.; Boroujeni, N.S.; Hajian, M.; Rahmani, H.R.; Nasr-Esfahani, M.H. A physiological, rather than a superovulated, post-implantation environment can attenuate the compromising effect of assisted reproductive techniques on gene expression in developing mice embryos. Mol. Reprod. Dev. 2015, 82, 191–206. [Google Scholar] [CrossRef]
- Saenz-de-Juano, M.D.; Billooye, K.; Smitz, J.; Anckaert, E. The loss of imprinted DNA methylation in mouse blastocysts is inflicted to a similar extent by in vitro follicle culture and ovulation induction. Mol. Hum. Reprod. 2016, 22, 427–441. [Google Scholar] [CrossRef]
- Ratajczak, M.Z. Igf2-H19, an imprinted tandem gene, is an important regulator of embryonic development, a guardian of proliferation of adult pluripotent stem cells, a regulator of longevity, and a ‘passkey’ to cancerogenesis. Folia Histochem. Cytobiol. 2012, 50, 171–179. [Google Scholar] [CrossRef]
- Ma, Y.; Ma, Y.; Wen, L.; Lei, H.; Chen, S.; Wang, X. Changes in DNA methylation and imprinting disorders in E9.5 mouse fetuses and placentas derived from vitrified eight-cell embryos. Mol. Reprod. Dev. 2019, 86, 404–415. [Google Scholar] [CrossRef]
- Bhattacharya, S. Maternal and perinatal outcomes after fresh versus frozen embryo transfer-what is the risk-benefit ratio? Fertil. Steril. 2016, 106, 241–243. [Google Scholar] [CrossRef]
- Toraño, E.G.; García, M.G.; Fernández-Morera, J.L.; Niño-García, P.; Fernández, A.F. The Impact of External Factors on the Epigenome: In Utero and over Lifetime. Biomed. Res. Int. 2016, 2016, 2568635. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, E.Y.; Cui, X.S.; Tae, J.C.; Lee, W.D.; Kim, N.H.; Park, S.P.; Lim, J.H. Increase in DNA fragmentation and apoptosis-related gene expression in frozen-thawed bovine blastocysts. Zygote 2006, 14, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.; Geng, L.; Huang, R.; Peng, W.; Chen, X.; Jiang, X.; Yu, M.; Li, M.; Huang, Y.; Yang, X. Effect of vitrification on in vitro development and imprinted gene Grb10 in mouse embryos. Reproduction 2017, 154, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, L.; Deng, T.; Zou, P.; Wang, Y.; Quan, F.; Zhang, Y. Effects of oocyte vitrification on epigenetic status in early bovine embryos. Theriogenology 2016, 86, 868–878. [Google Scholar] [CrossRef]
- Derakhshan-Horeh, M.; Abolhassani, F.; Jafarpour, F.; Moini, A.; Karbalaie, K.; Hosseini, S.M.; Nasr-Esfahani, M.H. Vitrification at Day3 stage appears not to affect the methylation status of H19/IGF2 differentially methylated region of in vitro produced human blastocysts. Cryobiology 2016, 73, 168–174. [Google Scholar] [CrossRef]
- Hiura, H.; Hattori, H.; Kobayashi, N.; Okae, H.; Chiba, H.; Miyauchi, N.; Kitamura, A.; Kikuchi, H.; Yoshida, H.; Arima, T. Genome-wide microRNA expression profiling in placentae from frozen-thawed blastocyst transfer. Clin. Epigenetics 2017, 9, 79. [Google Scholar] [CrossRef]
- Velker, B.A.M.; Denomme, M.M.; Krafty, R.T.; Mann, M.R.W. Maintenance of Mest imprinted methylation in blastocyst-stage mouse embryos is less stable than other imprinted loci following superovulation or embryo culture. Environ. Epigenet 2017, 3, dvx015. [Google Scholar] [CrossRef] [PubMed]
- Market-Velker, B.A.; Fernandes, A.D.; Mann, M.R. Side-by-side comparison of five commercial media systems in a mouse model: Suboptimal in vitro culture interferes with imprint maintenance. Biol. Reprod. 2010, 83, 938–950. [Google Scholar] [CrossRef]
- Fleming, T.P.; Kwong, W.Y.; Porter, R.; Ursell, E.; Fesenko, I.; Wilkins, A.; Miller, D.J.; Watkins, A.J.; Eckert, J.J. The embryo and its future. Biol. Reprod. 2004, 71, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Fair, T. The ART of studying early embryo development: Progress and challenges in ruminant embryo culture. Theriogenology 2014, 81, 49–55. [Google Scholar] [CrossRef]
- Lonergan, P.; Fair, T.; Corcoran, D.; Evans, A.C. Effect of culture environment on gene expression and developmental characteristics in IVF-derived embryos. Theriogenology 2006, 65, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Lonergan, P.; Rizos, D.; Kanka, J.; Nemcova, L.; Mbaye, A.M.; Kingston, M.; Wade, M.; Duffy, P.; Boland, M.P. Temporal sensitivity of bovine embryos to culture environment after fertilization and the implications for blastocyst quality. Reproduction 2003, 126, 337–346. [Google Scholar] [CrossRef]
- Fauque, P.; Mondon, F.; Letourneur, F.; Ripoche, M.A.; Journot, L.; Barbaux, S.; Dandolo, L.; Patrat, C.; Wolf, J.P.; Jouannet, P.; et al. In vitro fertilization and embryo culture strongly impact the placental transcriptome in the mouse model. PLoS ONE 2010, 5, e9218. [Google Scholar] [CrossRef] [PubMed]
- Donjacour, A.; Liu, X.; Lin, W.; Simbulan, R.; Rinaudo, P.F. In Vitro Fertilization Affects Growth and Glucose Metabolism in a Sex-Specific Manner in an Outbred Mouse Model. Biol. Reprod. 2014, 90, 80. [Google Scholar] [CrossRef] [PubMed]
- Sui, L.; An, L.; Tan, K.; Wang, Z.; Wang, S.; Miao, K.; Ren, L.; Tao, L.; He, S.; Yu, Y.; et al. Dynamic proteomic profiles of in vivo- and in vitro-produced mouse postimplantation extraembryonic tissues and placentas. Biol. Reprod. 2014, 91, 155. [Google Scholar] [CrossRef]
- Crosier, A.E.; Farin, C.E.; Rodriguez, K.F.; Blondin, P.; Alexander, J.E.; Farin, P.W. Development of skeletal muscle and expression of candidate genes in bovine fetuses from embryos produced in vivo or in vitro. Biol. Reprod. 2002, 67, 401–408. [Google Scholar] [CrossRef]
- Barros, C.M.; Satrapa, R.A.; Castilho, A.C.; Fontes, P.K.; Razza, E.M.; Ereno, R.L.; Nogueira, M.F. Effect of superstimulatory treatments on the expression of genes related to ovulatory capacity, oocyte competence and embryo development in cattle. Reprod. Fertil. Dev. 2012, 25, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Kerjean, A.; Couvert, P.; Heams, T.; Chalas, C.; Poirier, K.; Chelly, J.; Jouannet, P.; Paldi, A.; Poirot, C. In vitro follicular growth affects oocyte imprinting establishment in mice. Eur. J. Hum. Genet. 2003, 11, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Gioia, L.; Barboni, B.; Turriani, M.; Capacchietti, G.; Pistilli, M.G.; Berardinelli, P.; Mattioli, M. The capability of reprogramming the male chromatin after fertilization is dependent on the quality of oocyte maturation. Reproduction 2005, 130, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Borghol, N.; Lornage, J.; Blachère, T.; Sophie Garret, A.; Lefèvre, A. Epigenetic status of the H19 locus in human oocytes following in vitro maturation. Genomics 2006, 87, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Kuhtz, J.; Romero, S.; De Vos, M.; Smitz, J.; Haaf, T.; Anckaert, E. Human in vitro oocyte maturation is not associated with increased imprinting error rates at LIT1, SNRPN, PEG3 and GTL2. Hum. Reprod. 2014, 29, 1995–2005. [Google Scholar] [CrossRef] [PubMed]
- Pliushch, G.; Schneider, E.; Schneider, T.; El Hajj, N.; Rösner, S.; Strowitzki, T.; Haaf, T. In vitro maturation of oocytes is not associated with altered deoxyribonucleic acid methylation patterns in children from in vitro fertilization or intracytoplasmic sperm injection. Fertil. Steril. 2015, 103, 720–727. [Google Scholar] [CrossRef]
- Hewitson, L.; Takahashi, D.; Dominko, T.; Simerly, C.; Schatten, G. Fertilization and embryo development to blastocysts after intracytoplasmic sperm injection in the rhesus monkey. Hum. Reprod. 1998, 13, 3449–3455. [Google Scholar] [CrossRef]
- Dozortsev, D.; Wakaiama, T.; Ermilov, A.; Yanagimachi, R. Intracytoplasmic sperm injection in the rat. Zygote 1998, 6, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Ferré, L.B.; Alvarez-Gallardo, H.; Romo, S.; Fresno, C.; Stroud, T.; Stroud, B.; Lindsey, B.; Kjelland, M.E. Transvaginal ultrasound-guided oocyte retrieval in cattle: State-of-the-art and its impact on the in vitro fertilization embryo production outcome. Reprod. Domest. Anim. 2023, 58, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Kohda, T.; Ogonuki, N.; Inoue, K.; Furuse, T.; Kaneda, H.; Suzuki, T.; Kaneko-Ishino, T.; Wakayama, T.; Wakana, S.; Ogura, A.; et al. Intracytoplasmic sperm injection induces transcriptome perturbation without any transgenerational effect. Biochem. Biophys. Res. Commun. 2011, 410, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Bridges, P.J.; Jeoung, M.; Kim, H.; Kim, J.H.; Lee, D.R.; Ko, C.; Baker, D.J. Methodology matters: IVF versus ICSI and embryonic gene expression. Reprod. Biomed. Online 2011, 23, 234–244. [Google Scholar] [CrossRef]
- Wildman, D.E. IFPA award in placentology lecture: Phylogenomic origins and evolution of the mammalian placenta. Placenta 2016, 48 (Suppl. 1), S31–S39. [Google Scholar] [CrossRef]
- Okae, H.; Hiura, H.; Nishida, Y.; Funayama, R.; Tanaka, S.; Chiba, H.; Yaegashi, N.; Nakayama, K.; Sasaki, H.; Arima, T. Re-investigation and RNA sequencing-based identification of genes with placenta-specific imprinted expression. Hum. Mol. Genet. 2012, 21, 548–558. [Google Scholar] [CrossRef]
- Nelissen, E.C.; van Montfoort, A.P.; Dumoulin, J.C.; Evers, J.L. Epigenetics and the placenta. Hum. Reprod. Update 2011, 17, 397–417. [Google Scholar] [CrossRef] [PubMed]
- Sakian, S.; Louie, K.; Wong, E.C.; Havelock, J.; Kashyap, S.; Rowe, T.; Taylor, B.; Ma, S. Altered gene expression of H19 and IGF2 in placentas from ART pregnancies. Placenta 2015, 36, 1100–1105. [Google Scholar] [CrossRef]
- Katari, S.; Turan, N.; Bibikova, M.; Erinle, O.; Chalian, R.; Foster, M.; Gaughan, J.P.; Coutifaris, C.; Sapienza, C. DNA methylation and gene expression differences in children conceived in vitro or in vivo. Hum. Mol. Genet. 2009, 18, 3769–3778. [Google Scholar] [CrossRef]
- Shi, X.; Ni, Y.; Zheng, H.; Chen, S.; Zhong, M.; Wu, F.; Xia, R.; Luo, Y. Abnormal methylation patterns at the IGF2/H19 imprinting control region in phenotypically normal babies conceived by assisted reproductive technologies. Eur. J. Obstet. Gynecol. Reprod. Biol. 2011, 158, 52–55. [Google Scholar] [CrossRef] [PubMed]
- Nelissen, E.C.; Dumoulin, J.C.; Daunay, A.; Evers, J.L.; Tost, J.; van Montfoort, A.P. Placentas from pregnancies conceived by IVF/ICSI have a reduced DNA methylation level at the H19 and MEST differentially methylated regions. Hum. Reprod. 2013, 28, 1117–1126. [Google Scholar] [CrossRef]
- Hanna, C.W.; Demond, H.; Kelsey, G. Epigenetic regulation in development: Is the mouse a good model for the human? Hum. Reprod. Update 2018, 24, 556–576. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, H.; Sato, A.; Otsu, E.; Hiura, H.; Tomatsu, C.; Utsunomiya, T.; Sasaki, H.; Yaegashi, N.; Arima, T. Aberrant DNA methylation of imprinted loci in sperm from oligospermic patients. Hum. Mol. Genet. 2007, 16, 2542–2551. [Google Scholar] [CrossRef]
- Young, L.E.; Fernandes, K.; McEvoy, T.G.; Butterwith, S.C.; Gutierrez, C.G.; Carolan, C.; Broadbent, P.J.; Robinson, J.J.; Wilmut, I.; Sinclair, K.D. Epigenetic change in IGF2R is associated with fetal overgrowth after sheep embryo culture. Nat. Genet. 2001, 27, 153–154. [Google Scholar] [CrossRef] [PubMed]
- de Waal, E.; Vrooman, L.A.; Fischer, E.; Ord, T.; Mainigi, M.A.; Coutifaris, C.; Schultz, R.M.; Bartolomei, M.S. The cumulative effect of assisted reproduction procedures on placental development and epigenetic perturbations in a mouse model. Hum. Mol. Genet. 2015, 24, 6975–6985. [Google Scholar] [CrossRef] [PubMed]
- de Waal, E.; Mak, W.; Calhoun, S.; Stein, P.; Ord, T.; Krapp, C.; Coutifaris, C.; Schultz, R.M.; Bartolomei, M.S. In vitro culture increases the frequency of stochastic epigenetic errors at imprinted genes in placental tissues from mouse concepti produced through assisted reproductive technologies. Biol. Reprod. 2014, 90, 22. [Google Scholar] [CrossRef]
- Wisborg, K.; Ingerslev, H.J.; Henriksen, T.B. IVF and stillbirth: A prospective follow-up study. Hum. Reprod. 2010, 25, 1312–1316. [Google Scholar] [CrossRef]
- Hart, R.; Norman, R.J. The longer-term health outcomes for children born as a result of IVF treatment: Part I--General health outcomes. Hum. Reprod. Update 2013, 19, 232–243. [Google Scholar] [CrossRef]
- Pinborg, A.; Wennerholm, U.B.; Romundstad, L.B.; Loft, A.; Aittomaki, K.; Söderström-Anttila, V.; Nygren, K.G.; Hazekamp, J.; Bergh, C. Why do singletons conceived after assisted reproduction technology have adverse perinatal outcome? Systematic review and meta-analysis. Hum. Reprod. Update 2013, 19, 87–104. [Google Scholar] [CrossRef]
- Henningsen, A.K.; Pinborg, A.; Lidegaard, Ø.; Vestergaard, C.; Forman, J.L.; Andersen, A.N. Perinatal outcome of singleton siblings born after assisted reproductive technology and spontaneous conception: Danish national sibling-cohort study. Fertil. Steril. 2011, 95, 959–963. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Shetty, A.; Hamilton, M.; Bhattacharya, S.; Maheshwari, A. Obstetric and perinatal outcomes in singleton pregnancies resulting from IVF/ICSI: A systematic review and meta-analysis. Hum. Reprod. Update 2012, 18, 485–503. [Google Scholar] [CrossRef]
- Helmerhorst, F.M.; Perquin, D.A.; Donker, D.; Keirse, M.J. Perinatal outcome of singletons and twins after assisted conception: A systematic review of controlled studies. BMJ 2004, 328, 261. [Google Scholar] [CrossRef]
- Davies, M.J.; Moore, V.M.; Willson, K.J.; Van Essen, P.; Priest, K.; Scott, H.; Haan, E.A.; Chan, A. Reproductive technologies and the risk of birth defects. N. Engl. J. Med. 2012, 366, 1803–1813. [Google Scholar] [CrossRef]
- Buckett, W.M.; Chian, R.C.; Holzer, H.; Dean, N.; Usher, R.; Tan, S.L. Obstetric outcomes and congenital abnormalities after in vitro maturation, in vitro fertilization, and intracytoplasmic sperm injection. Obstet. Gynecol. 2007, 110, 885–891. [Google Scholar] [CrossRef]
- Seggers, J.; Pontesilli, M.; Ravelli, A.C.J.; Painter, R.C.; Hadders-Algra, M.; Heineman, M.J.; Repping, S.; Mol, B.W.J.; Roseboom, T.J.; Ensing, S. Effects of in vitro fertilization and maternal characteristics on perinatal outcomes: A population-based study using siblings. Fertil. Steril. 2016, 105, 590–598.e2. [Google Scholar] [CrossRef]
- Horton, J.; Sterrenburg, M.; Lane, S.; Maheshwari, A.; Li, T.C.; Cheong, Y. Reproductive, obstetric, and perinatal outcomes of women with adenomyosis and endometriosis: A systematic review and meta-analysis. Hum. Reprod. Update 2019, 25, 592–632. [Google Scholar] [CrossRef] [PubMed]
- Sterling, L.; Liu, J.; Okun, N.; Sakhuja, A.; Sierra, S.; Greenblatt, E. Pregnancy outcomes in women with polycystic ovary syndrome undergoing in vitro fertilization. Fertil. Steril. 2016, 105, 791–797.e2. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, R.; Gissler, M.; Tiitinen, A. Obstetric outcome among women with unexplained infertility after IVF: A matched case-control study. Hum. Reprod. 2002, 17, 1755–1761. [Google Scholar] [CrossRef] [PubMed]
- Luke, B.; Gopal, D.; Cabral, H.; Stern, J.E.; Diop, H. Pregnancy, birth, and infant outcomes by maternal fertility status: The Massachusetts Outcomes Study of Assisted Reproductive Technology. Am. J. Obstet. Gynecol. 2017, 217, 327.e1–327.e14. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Liang, Z.; Cai, R.; Jin, S.; Xia, T.; Wang, C.; Kuang, Y. Association of adverse birth outcomes with in vitro fertilization after controlling infertility factors based on a singleton live birth cohort. Sci. Rep. 2022, 12, 4528. [Google Scholar] [CrossRef] [PubMed]
- Schieve, L.A.; Meikle, S.F.; Ferre, C.; Peterson, H.B.; Jeng, G.; Wilcox, L.S. Low and very low birth weight in infants conceived with use of assisted reproductive technology. N. Engl. J. Med. 2002, 346, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Żyła, M.M.; Wilczyński, J.; Nowakowska-Głąb, A.; Maniecka-Bryła, I.; Nowakowska, D. Pregnancy and Delivery in Women with Uterine Malformations. Adv. Clin. Exp. Med. 2015, 24, 873–879. [Google Scholar] [CrossRef]
- Boyle, A.K.; Rinaldi, S.F.; Norman, J.E.; Stock, S.J. Preterm birth: Inflammation, fetal injury and treatment strategies. J. Reprod. Immunol. 2017, 119, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Sunkara, S.K.; Antonisamy, B.; Redla, A.C.; Kamath, M.S. Female causes of infertility are associated with higher risk of preterm birth and low birth weight: Analysis of 117 401 singleton live births following IVF. Hum. Reprod. 2021, 36, 676–682. [Google Scholar] [CrossRef]
- Dunietz, G.L.; Holzman, C.; Zhang, Y.; Li, C.; Todem, D.; Boulet, S.L.; McKane, P.; Kissin, D.M.; Copeland, G.; Bernson, D.; et al. Assisted reproduction and risk of preterm birth in singletons by infertility diagnoses and treatment modalities: A population-based study. J. Assist. Reprod. Genet. 2017, 34, 1529–1535. [Google Scholar] [CrossRef]
- Kawwass, J.F.; Crawford, S.; Kissin, D.M.; Session, D.R.; Boulet, S.; Jamieson, D.J. Tubal factor infertility and perinatal risk after assisted reproductive technology. Obstet. Gynecol. 2013, 121, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Mariappen, U.; Keane, K.N.; Hinchliffe, P.M.; Dhaliwal, S.S.; Yovich, J.L. Neither male age nor semen parameters influence clinical pregnancy or live birth outcomes from IVF. Reprod. Biol. 2018, 18, 324–329. [Google Scholar] [CrossRef]
- Pinborg, A.; Loft, A.; Aaris Henningsen, A.K.; Rasmussen, S.; Andersen, A.N. Infant outcome of 957 singletons born after frozen embryo replacement: The Danish National Cohort Study 1995–2006. Fertil Steril 2010, 94, 1320–1327. [Google Scholar] [CrossRef]
- Mohseni, R.; Mohammed, S.H.; Safabakhsh, M.; Mohseni, F.; Monfared, Z.S.; Seyyedi, J.; Mejareh, Z.N.; Alizadeh, S. Birth Weight and Risk of Cardiovascular Disease Incidence in Adulthood: A Dose-Response Meta-analysis. Curr. Atheroscler. Rep. 2020, 22, 12. [Google Scholar] [CrossRef]
- Skilton, M.R.; Siitonen, N.; Würtz, P.; Viikari, J.S.; Juonala, M.; Seppälä, I.; Laitinen, T.; Lehtimäki, T.; Taittonen, L.; Kähönen, M.; et al. High birth weight is associated with obesity and increased carotid wall thickness in young adults: The cardiovascular risk in young Finns study. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1064–1068. [Google Scholar] [CrossRef] [PubMed]
- Hansen, M.; Kurinczuk, J.J.; Bower, C.; Webb, S. The risk of major birth defects after intracytoplasmic sperm injection and in vitro fertilization. N. Engl. J. Med. 2002, 346, 725–730. [Google Scholar] [CrossRef]
- Klemetti, R.; Gissler, M.; Sevón, T.; Koivurova, S.; Ritvanen, A.; Hemminki, E. Children born after assisted fertilization have an increased rate of major congenital anomalies. Fertil. Steril. 2005, 84, 1300–1307. [Google Scholar] [CrossRef] [PubMed]
- Smithers, P.R.; Halliday, J.; Hale, L.; Talbot, J.M.; Breheny, S.; Healy, D. High frequency of cesarean section, antepartum hemorrhage, placenta previa, and preterm delivery in in-vitro fertilization twin pregnancies. Fertil. Steril. 2003, 80, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Manoura, A.; Korakaki, E.; Hatzidaki, E.; Bikouvarakis, S.; Papageorgiou, M.; Giannakopoulou, C. Perinatal outcome of twin pregnancies after in vitro fertilization. Acta Obstet. Gynecol. Scand. 2004, 83, 1079–1084. [Google Scholar] [CrossRef]
- Bosch, B.; Sutcliffe, A. Congenital Anomalies Following Assisted Reproductive Technology. In Complications and Outcomes of Assisted Reproduction; Rizk, B., Gerris, J., Eds.; Cambridge University Press: Cambridge, UK, 2017; pp. 15–23. [Google Scholar] [CrossRef]
- Katalinic, A.; Rösch, C.; Ludwig, M. Pregnancy course and outcome after intracytoplasmic sperm injection: A controlled, prospective cohort study. Fertil. Steril. 2004, 81, 1604–1616. [Google Scholar] [CrossRef]
- El Hajj, N.; Haaf, T. Epigenetic disturbances in in vitro cultured gametes and embryos: Implications for human assisted reproduction. Fertil. Steril. 2013, 99, 632–641. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Norman, R.J.; Kristiansson, P. The effect of various infertility treatments on the risk of preterm birth. Hum. Reprod. 2002, 17, 945–949. [Google Scholar] [CrossRef]
- Luke, B.; Brown, M.B.; Wantman, E.; Forestieri, N.E.; Browne, M.L.; Fisher, S.C.; Yazdy, M.M.; Ethen, M.K.; Canfield, M.A.; Watkins, S.; et al. The risk of birth defects with conception by ART. Hum. Reprod. 2021, 36, 116–129. [Google Scholar] [CrossRef]
- Hansen, M.; Kurinczuk, J.J.; Milne, E.; de Klerk, N.; Bower, C. Assisted reproductive technology and birth defects: A systematic review and meta-analysis. Hum. Reprod. Update 2013, 19, 330–353. [Google Scholar] [CrossRef] [PubMed]
- Olson, C.K.; Keppler-Noreuil, K.M.; Romitti, P.A.; Budelier, W.T.; Ryan, G.; Sparks, A.E.; Van Voorhis, B.J. In vitro fertilization is associated with an increase in major birth defects. Fertil. Steril. 2005, 84, 1308–1315. [Google Scholar] [CrossRef]
- Kanyó, K.; Konc, J. A follow-up study of children born after diode laser assisted hatching. Eur. J. Obstet. Gynecol. Reprod. Biol. 2003, 110, 176–180. [Google Scholar] [CrossRef]
- Nassar, A.H.; Usta, I.M.; Rechdan, J.B.; Harb, T.S.; Adra, A.M.; Abu-Musa, A.A. Pregnancy outcome in spontaneous twins versus twins who were conceived through in vitro fertilization. Am. J. Obstet. Gynecol. 2003, 189, 513–518. [Google Scholar] [CrossRef]
- Hansen, M.; Kurinczuk, J.J.; de Klerk, N.; Burton, P.; Bower, C. Assisted reproductive technology and major birth defects in Western Australia. Obstet. Gynecol. 2012, 120, 852–863. [Google Scholar] [CrossRef]
- Zhu, J.L.; Basso, O.; Obel, C.; Bille, C.; Olsen, J. Infertility, infertility treatment, and congenital malformations: Danish national birth cohort. BMJ 2006, 333, 679. [Google Scholar] [CrossRef]
- La Rovere, M.; Franzago, M.; Stuppia, L. Epigenetics and Neurological Disorders in ART. Int. J. Mol. Sci. 2019, 20, 4169. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lv, Z.; Yang, Y.; Dong, G.; Yu, Y.; Cui, Y.; Tong, M.; Wang, L.; Zhou, Z.; Zhu, H.; et al. Blastomere biopsy influences epigenetic reprogramming during early embryo development, which impacts neural development and function in resulting mice. Cell Mol. Life Sci. 2014, 71, 1761–1774. [Google Scholar] [CrossRef]
- Strömberg, B.; Dahlquist, G.; Ericson, A.; Finnström, O.; Köster, M.; Stjernqvist, K. Neurological sequelae in children born after in-vitro fertilisation: A population-based study. Lancet 2002, 359, 461–465. [Google Scholar] [CrossRef] [PubMed]
- Lidegaard, O.; Pinborg, A.; Andersen, A.N. Imprinting diseases and IVF: Danish National IVF cohort study. Hum. Reprod. 2005, 20, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Källén, A.J.; Finnström, O.O.; Lindam, A.P.; Nilsson, E.M.; Nygren, K.G.; Olausson, P.M. Cerebral palsy in children born after in vitro fertilization. Is the risk decreasing? Eur. J. Paediatr. Neurol. 2010, 14, 526–530. [Google Scholar] [CrossRef]
- Hvidtjørn, D.; Grove, J.; Schendel, D.; Svaerke, C.; Schieve, L.A.; Uldall, P.; Ernst, E.; Jacobsson, B.; Thorsen, P. Multiplicity and early gestational age contribute to an increased risk of cerebral palsy from assisted conception: A population-based cohort study. Hum. Reprod. 2010, 25, 2115–2123. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.L.; Hvidtjørn, D.; Basso, O.; Obel, C.; Thorsen, P.; Uldall, P.; Olsen, J. Parental infertility and cerebral palsy in children. Hum. Reprod. 2010, 25, 3142–3145. [Google Scholar] [CrossRef]
- Bay, B.; Mortensen, E.L.; Kesmodel, U.S. Assisted reproduction and child neurodevelopmental outcomes: A systematic review. Fertil. Steril. 2013, 100, 844–853. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Mannan, O.; Sutcliffe, A. I was born following ART: How will I get on at school? Semin. Fetal Neonatal Med. 2014, 19, 245–249. [Google Scholar] [CrossRef]
- Punamäki, R.L.; Tiitinen, A.; Lindblom, J.; Unkila-Kallio, L.; Flykt, M.; Vänskä, M.; Poikkeus, P.; Tulppala, M. Mental health and developmental outcomes for children born after ART: A comparative prospective study on child gender and treatment type. Hum. Reprod. 2016, 31, 100–107. [Google Scholar] [CrossRef]
- Rumbold, A.R.; Moore, V.M.; Whitrow, M.J.; Oswald, T.K.; Moran, L.J.; Fernandez, R.C.; Barnhart, K.T.; Davies, M.J. The impact of specific fertility treatments on cognitive development in childhood and adolescence: A systematic review. Hum. Reprod. 2017, 32, 1489–1507. [Google Scholar] [CrossRef]
- Maimburg, R.D.; Vaeth, M. Do children born after assisted conception have less risk of developing infantile autism? Hum. Reprod. 2007, 22, 1841–1843. [Google Scholar] [CrossRef]
- Hvidtjørn, D.; Grove, J.; Schendel, D.; Schieve, L.A.; Sværke, C.; Ernst, E.; Thorsen, P. Risk of autism spectrum disorders in children born after assisted conception: A population-based follow-up study. J. Epidemiol. Community Health 2011, 65, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Kissin, D.M.; Zhang, Y.; Boulet, S.L.; Fountain, C.; Bearman, P.; Schieve, L.; Yeargin-Allsopp, M.; Jamieson, D.J. Association of assisted reproductive technology (ART) treatment and parental infertility diagnosis with autism in ART-conceived children. Hum. Reprod. 2015, 30, 454–465. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Gao, J.; He, X.; Cai, Y.; Wang, L.; Fan, X. Association between assisted reproductive technology and the risk of autism spectrum disorders in the offspring: A meta-analysis. Sci. Rep. 2017, 7, 46207. [Google Scholar] [CrossRef]
- Hargreave, M.; Jensen, A.; Toender, A.; Andersen, K.K.; Kjaer, S.K. Fertility treatment and childhood cancer risk: A systematic meta-analysis. Fertil. Steril. 2013, 100, 150–161. [Google Scholar] [CrossRef] [PubMed]
- Chiavarini, M.; Ostorero, A.; Naldini, G.; Fabiani, R. Cancer Risk in Children and Young Adults (Offspring) Born after Medically Assisted Reproduction: A Systematic Review and Meta-Analysis. J 2019, 2, 430–448. [Google Scholar] [CrossRef]
- Wang, T.; Chen, L.; Yang, T.; Wang, L.; Zhao, L.; Zhang, S.; Ye, Z.; Chen, L.; Zheng, Z.; Qin, J. Cancer risk among children conceived by fertility treatment. Int. J. Cancer 2019, 144, 3001–3013. [Google Scholar] [CrossRef]
- Williams, C.L.; Bunch, K.J.; Murphy, M.F.G.; Stiller, C.A.; Botting, B.J.; Wallace, W.H.; Davies, M.C.; Sutcliffe, A.G. Cancer risk in children born after donor ART. Hum. Reprod. 2018, 33, 140–146. [Google Scholar] [CrossRef]
- Sundh, K.J.; Henningsen, A.K.; Källen, K.; Bergh, C.; Romundstad, L.B.; Gissler, M.; Pinborg, A.; Skjaerven, R.; Tiitinen, A.; Vassard, D.; et al. Cancer in children and young adults born after assisted reproductive technology: A Nordic cohort study from the Committee of Nordic ART and Safety (CoNARTaS). Hum. Reprod. 2014, 29, 2050–2057. [Google Scholar] [CrossRef]
- Lerner-Geva, L.; Boyko, V.; Ehrlich, S.; Mashiach, S.; Hourvitz, A.; Haas, J.; Margalioth, E.; Levran, D.; Calderon, I.; Orvieto, R.; et al. Possible risk for cancer among children born following assisted reproductive technology in Israel. Pediatr. Blood Cancer 2017, 64, e26292. [Google Scholar] [CrossRef] [PubMed]
- Spaan, M.; van den Belt-Dusebout, A.W.; van den Heuvel-Eibrink, M.M.; Hauptmann, M.; Lambalk, C.B.; Burger, C.W.; van Leeuwen, F.E. Risk of cancer in children and young adults conceived by assisted reproductive technology. Hum. Reprod. 2019, 34, 740–750. [Google Scholar] [CrossRef]
- Källén, B.; Finnström, O.; Lindam, A.; Nilsson, E.; Nygren, K.G.; Olausson, P.O. Cancer risk in children and young adults conceived by in vitro fertilization. Pediatrics 2010, 126, 270–276. [Google Scholar] [CrossRef]
- Barker, D.J.; Osmond, C.; Golding, J.; Kuh, D.; Wadsworth, M.E. Growth in utero, blood pressure in childhood and adult life, and mortality from cardiovascular disease. BMJ 1989, 298, 564–567. [Google Scholar] [CrossRef]
- Barker, D.J. The origins of the developmental origins theory. J. Intern. Med. 2007, 261, 412–417. [Google Scholar] [CrossRef]
- Morton, J.S.; Cooke, C.L.; Davidge, S.T. In Utero Origins of Hypertension: Mechanisms and Targets for Therapy. Physiol. Rev. 2016, 96, 549–603. [Google Scholar] [CrossRef] [PubMed]
- Schenewerk, A.L.; Ramírez, F.; Foote, C.; Ji, T.; Martínez-Lemus, L.A.; Rivera, R.M. Effects of the use of assisted reproduction and high-caloric diet consumption on body weight and cardiovascular health of juvenile mouse offspring. Reproduction 2014, 147, 111–123. [Google Scholar] [CrossRef]
- Rexhaj, E.; Pireva, A.; Paoloni-Giacobino, A.; Allemann, Y.; Cerny, D.; Dessen, P.; Sartori, C.; Scherrer, U.; Rimoldi, S.F. Prevention of vascular dysfunction and arterial hypertension in mice generated by assisted reproductive technologies by addition of melatonin to culture media. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1151–H1156. [Google Scholar] [CrossRef]
- Wang, L.Y.; Le, F.; Wang, N.; Li, L.; Liu, X.Z.; Zheng, Y.M.; Lou, H.Y.; Xu, X.R.; Chen, Y.L.; Zhu, X.M.; et al. Alteration of fatty acid metabolism in the liver, adipose tissue, and testis of male mice conceived through assisted reproductive technologies: Fatty acid metabolism in ART mice. Lipids Health Dis. 2013, 12, 5. [Google Scholar] [CrossRef]
- Yeung, E.H.; Druschel, C. Cardiometabolic health of children conceived by assisted reproductive technologies. Fertil. Steril. 2013, 99, 318–326. [Google Scholar] [CrossRef]
- Scherrer, U.; Rexhaj, E.; Allemann, Y.; Sartori, C.; Rimoldi, S.F. Cardiovascular dysfunction in children conceived by assisted reproductive technologies. Eur. Heart J. 2015, 36, 1583–1589. [Google Scholar] [CrossRef]
- Wikstrand, M.H.; Niklasson, A.; Strömland, K.; Hellström, A. Abnormal vessel morphology in boys born after intracytoplasmic sperm injection. Acta Paediatr. 2008, 97, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Pan, H.T.; Lin, X.H.; Zhang, J.Y.; Jiang, Y.; Tian, S.; Chen, L.T.; Liu, M.E.; Xiong, Y.M.; Huang, H.F.; et al. Altered protein expression profiles in umbilical veins: Insights into vascular dysfunctions of the children born after in vitro fertilization. Biol. Reprod. 2014, 91, 71. [Google Scholar] [CrossRef] [PubMed]
- de Jong, F.; Monuteaux, M.C.; van Elburg, R.M.; Gillman, M.W.; Belfort, M.B. Systematic review and meta-analysis of preterm birth and later systolic blood pressure. Hypertension 2012, 59, 226–234. [Google Scholar] [CrossRef]
- Kaijser, M.; Bonamy, A.K.; Akre, O.; Cnattingius, S.; Granath, F.; Norman, M.; Ekbom, A. Perinatal risk factors for ischemic heart disease: Disentangling the roles of birth weight and preterm birth. Circulation 2008, 117, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.Y.; Liu, X.M.; Jin, L.; Wang, T.T.; Ullah, K.; Sheng, J.Z.; Huang, H.F. Cardiovascular and metabolic profiles of offspring conceived by assisted reproductive technologies: A systematic review and meta-analysis. Fertil. Steril. 2017, 107, 622–631.e5. [Google Scholar] [CrossRef] [PubMed]
- Scherrer, U.; Rimoldi, S.F.; Rexhaj, E.; Stuber, T.; Duplain, H.; Garcin, S.; de Marchi, S.F.; Nicod, P.; Germond, M.; Allemann, Y.; et al. Systemic and pulmonary vascular dysfunction in children conceived by assisted reproductive technologies. Circulation 2012, 125, 1890–1896. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Wu, L.; Zhao, J.; Wu, F.; Davies, M.J.; Wittert, G.A.; Norman, R.J.; Robker, R.L.; Heilbronn, L.K. Altered glucose metabolism in mouse and humans conceived by IVF. Diabetes 2014, 63, 3189–3198. [Google Scholar] [CrossRef]
- Ceelen, M.; van Weissenbruch, M.M.; Vermeiden, J.P.; van Leeuwen, F.E.; Delemarre-van de Waal, H.A. Cardiometabolic differences in children born after in vitro fertilization: Follow-up study. J. Clin. Endocrinol. Metab. 2008, 93, 1682–1688. [Google Scholar] [CrossRef]
- Belva, F.; Henriet, S.; Liebaers, I.; Van Steirteghem, A.; Celestin-Westreich, S.; Bonduelle, M. Medical outcome of 8-year-old singleton ICSI children (born >or=32 weeks’ gestation) and a spontaneously conceived comparison group. Hum. Reprod. 2007, 22, 506–515. [Google Scholar] [CrossRef]
- Belva, F.; Painter, R.; Bonduelle, M.; Roelants, M.; Devroey, P.; De Schepper, J. Are ICSI adolescents at risk for increased adiposity? Hum. Reprod. 2012, 27, 257–264. [Google Scholar] [CrossRef]
- Sakka, S.D.; Loutradis, D.; Kanaka-Gantenbein, C.; Margeli, A.; Papastamataki, M.; Papassotiriou, I.; Chrousos, G.P. Absence of insulin resistance and low-grade inflammation despite early metabolic syndrome manifestations in children born after in vitro fertilization. Fertil. Steril. 2010, 94, 1693–1699. [Google Scholar] [CrossRef]
- Heber, M.F.; Ptak, G.E. The effects of assisted reproduction technologies on metabolic health and disease†. Biol. Reprod. 2021, 104, 734–744. [Google Scholar] [CrossRef]
- Carson, C.; Sacker, A.; Kelly, Y.; Redshaw, M.; Kurinczuk, J.J.; Quigley, M.A. Asthma in children born after infertility treatment: Findings from the UK Millennium Cohort Study. Hum. Reprod. 2013, 28, 471–479. [Google Scholar] [CrossRef]
- Cui, L.; Zhou, W.; Xi, B.; Ma, J.; Hu, J.; Fang, M.; Hu, K.; Qin, Y.; You, L.; Cao, Y.; et al. Increased risk of metabolic dysfunction in children conceived by assisted reproductive technology. Diabetologia 2020, 63, 2150–2157. [Google Scholar] [CrossRef]
- Karimi, H.; Mahdavi, P.; Fakhari, S.; Faryabi, M.R.; Esmaeili, P.; Banafshi, O.; Mohammadi, E.; Fathi, F.; Mokarizadeh, A. Altered helper T cell-mediated immune responses in male mice conceived through in vitro fertilization. Reprod. Toxicol. 2017, 69, 196–203. [Google Scholar] [CrossRef]
- Xu, X.; Wu, H.; Bian, Y.; Cui, L.; Man, Y.; Wang, Z.; Zhang, X.; Zhang, C.; Geng, L. The altered immunological status of children conceived by assisted reproductive technology. Reprod. Biol. Endocrinol. 2021, 19, 171. [Google Scholar] [CrossRef]
- Zhang, Y.; Cui, Y.; Zhou, Z.; Sha, J.; Li, Y.; Liu, J. Altered global gene expressions of human placentae subjected to assisted reproductive technology treatments. Placenta 2010, 31, 251–258. [Google Scholar] [CrossRef]
- Paolino, M.; Koglgruber, R.; Cronin, S.J.F.; Uribesalgo, I.; Rauscher, E.; Harreiter, J.; Schuster, M.; Bancher-Todesca, D.; Pranjic, B.; Novatchkova, M.; et al. RANK links thymic regulatory T cells to fetal loss and gestational diabetes in pregnancy. Nature 2021, 589, 442–447. [Google Scholar] [CrossRef]
- Hargreave, M.; Jensen, A.; Hansen, M.K.; Dehlendorff, C.; Winther, J.F.; Schmiegelow, K.; Kjær, S.K. Association Between Fertility Treatment and Cancer Risk in Children. JAMA 2019, 322, 2203–2210. [Google Scholar] [CrossRef]
- Papi, A.; Blasi, F.; Canonica, G.W.; Morandi, L.; Richeldi, L.; Rossi, A. Treatment strategies for asthma: Reshaping the concept of asthma management. Allergy Asthma Clin. Immunol. 2020, 16, 75. [Google Scholar] [CrossRef]
- Pawankar, R. Allergic diseases and asthma: A global public health concern and a call to action. World Allergy Organ. J. 2014, 7, 12. [Google Scholar] [CrossRef]
- Harris, R.A.; Nagy-Szakal, D.; Kellermayer, R. Human metastable epiallele candidates link to common disorders. Epigenetics 2013, 8, 157–163. [Google Scholar] [CrossRef]
- Reese, S.E.; Xu, C.J.; den Dekker, H.T.; Lee, M.K.; Sikdar, S.; Ruiz-Arenas, C.; Merid, S.K.; Rezwan, F.I.; Page, C.M.; Ullemar, V.; et al. Epigenome-wide meta-analysis of DNA methylation and childhood asthma. J. Allergy Clin. Immunol. 2019, 143, 2062–2074. [Google Scholar] [CrossRef]
- Declercq, E.; Luke, B.; Belanoff, C.; Cabral, H.; Diop, H.; Gopal, D.; Hoang, L.; Kotelchuck, M.; Stern, J.E.; Hornstein, M.D. Perinatal outcomes associated with assisted reproductive technology: The Massachusetts Outcomes Study of Assisted Reproductive Technologies (MOSART). Fertil. Steril. 2015, 103, 888–895. [Google Scholar] [CrossRef]
- Huang, L.; Chen, Q.; Zhao, Y.; Wang, W.; Fang, F.; Bao, Y. Is elective cesarean section associated with a higher risk of asthma? A meta-analysis. J. Asthma 2015, 52, 16–25. [Google Scholar] [CrossRef]
- Hart, R.; Norman, R.J. The longer-term health outcomes for children born as a result of IVF treatment. Part II--Mental health and development outcomes. Hum. Reprod. Update 2013, 19, 244–250. [Google Scholar] [CrossRef]
- Kettner, L.O.; Henriksen, T.B.; Bay, B.; Ramlau-Hansen, C.H.; Kesmodel, U.S. Assisted reproductive technology and somatic morbidity in childhood: A systematic review. Fertil. Steril. 2015, 103, 707–719. [Google Scholar] [CrossRef]
- Ericson, A.; Nygren, K.G.; Olausson, P.O.; Källén, B. Hospital care utilization of infants born after IVF. Hum. Reprod. 2002, 17, 929–932. [Google Scholar] [CrossRef]
- Tsabouri, S.; Lavasidis, G.; Efstathiadou, A.; Papasavva, M.; Bellou, V.; Bergantini, H.; Priftis, K.; Ntzani, E.E. Association between childhood asthma and history of assisted reproduction techniques: A systematic review and meta-analysis. Eur. J. Pediatr. 2021, 180, 2007–2017. [Google Scholar] [CrossRef]
- Källén, B.; Finnström, O.; Nygren, K.G.; Otterblad Olausson, P. Asthma in Swedish children conceived by in vitro fertilisation. Arch. Dis. Child. 2013, 98, 92–96. [Google Scholar] [CrossRef]
- García-Blanco, A.; Diago, V.; Hervás, D.; Ghosn, F.; Vento, M.; Cháfer-Pericás, C. Anxiety and depressive symptoms, and stress biomarkers in pregnant women after in vitro fertilization: A prospective cohort study. Hum. Reprod. 2018, 33, 1237–1246. [Google Scholar] [CrossRef]
- Wijs, L.A.; Doherty, D.A.; Keelan, J.A.; Penova-Veselinovic, B.; Burton, P.; Yovich, J.L.; Hall, G.L.; Sly, P.D.; Holt, P.G.; Hart, R.J. Asthma and allergies in a cohort of adolescents conceived with ART. Reprod. Biomed. Online 2022, 45, 1255–1265. [Google Scholar] [CrossRef]
- Glover, V. Prenatal stress and its effects on the fetus and the child: Possible underlying biological mechanisms. Adv. Neurobiol. 2015, 10, 269–283. [Google Scholar] [CrossRef]
- Ahmadi, H.; Fathi, F.; Moeini, A.; Amidi, F.; Sobhani, A. Evaluation of prooxidant-antioxidant balance in in vitro fertilization-conceived mice. Clin. Exp. Reprod. Med. 2018, 45, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Zijlmans, M.A.; Korpela, K.; Riksen-Walraven, J.M.; de Vos, W.M.; de Weerth, C. Maternal prenatal stress is associated with the infant intestinal microbiota. Psychoneuroendocrinology 2015, 53, 233–245. [Google Scholar] [CrossRef]
- Moustaki, M.; Tsabouri, S.; Priftis, K.N.; Douros, K. Prenatal Stress Enhances Susceptibility to Allergic Diseases of Offspring. Endocr. Metab. Immune Disord. Drug Targets 2017, 17, 255–263. [Google Scholar] [CrossRef]
- Price, T.M.; Murphy, S.K.; Younglai, E.V. Perspectives: The possible influence of assisted reproductive technologies on transgenerational reproductive effects of environmental endocrine disruptors. Toxicol. Sci. 2007, 96, 218–226. [Google Scholar] [CrossRef]
- Baart, E.B.; Martini, E.; Eijkemans, M.J.; Van Opstal, D.; Beckers, N.G.; Verhoeff, A.; Macklon, N.S.; Fauser, B.C. Milder ovarian stimulation for in-vitro fertilization reduces aneuploidy in the human preimplantation embryo: A randomized controlled trial. Hum. Reprod. 2007, 22, 980–988. [Google Scholar] [CrossRef]
- Lainas, T.G.; Sfontouris, I.A.; Zorzovilis, I.Z.; Petsas, G.K.; Lainas, G.T.; Alexopoulou, E.; Kolibianakis, E.M. Flexible GnRH antagonist protocol versus GnRH agonist long protocol in patients with polycystic ovary syndrome treated for IVF: A prospective randomised controlled trial (RCT). Hum. Reprod. 2010, 25, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Schover, L.R.; Partridge, A.H.; Patrizio, P.; Wallace, W.H.; Hagerty, K.; Beck, L.N.; Brennan, L.V.; Oktay, K. American Society of Clinical Oncology recommendations on fertility preservation in cancer patients. J. Clin. Oncol. 2006, 24, 2917–2931. [Google Scholar] [CrossRef]
- Henningsen, A.A.; Gissler, M.; Skjaerven, R.; Bergh, C.; Tiitinen, A.; Romundstad, L.B.; Wennerholm, U.B.; Lidegaard, O.; Nyboe Andersen, A.; Forman, J.L.; et al. Trends in perinatal health after assisted reproduction: A Nordic study from the CoNARTaS group. Hum. Reprod. 2015, 30, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.M.; Hodgson, R.; Wong, T.Y.; Merrilees, M.; Norman, R.J.; Johnson, N.P. Single embryo transfer for all? Aust. N. Z. J. Obstet. Gynaecol. 2016, 56, 514–517. [Google Scholar] [CrossRef]
- Pandian, Z.; Marjoribanks, J.; Ozturk, O.; Serour, G.; Bhattacharya, S. Number of embryos for transfer following in vitro fertilisation or intra-cytoplasmic sperm injection. Cochrane Database Syst. Rev. 2013, 2013, Cd003416. [Google Scholar] [CrossRef]
- Gardner, D.K.; Lane, M.; Stevens, J.; Schoolcraft, W.B. Noninvasive assessment of human embryo nutrient consumption as a measure of developmental potential. Fertil. Steril. 2001, 76, 1175–1180. [Google Scholar] [CrossRef]
- Lane, M.; Gardner, D.K. Selection of viable mouse blastocysts prior to transfer using a metabolic criterion. Hum. Reprod. 1996, 11, 1975–1978. [Google Scholar] [CrossRef]
- Montskó, G.; Zrínyi, Z.; Janáky, T.; Szabó, Z.; Várnagy, Á.; Kovács, G.L.; Bódis, J. Noninvasive embryo viability assessment by quantitation of human haptoglobin alpha-1 fragment in the in vitro fertilization culture medium: An additional tool to increase success rate. Fertil. Steril. 2015, 103, 687–693. [Google Scholar] [CrossRef]
- Pallinger, E.; Bognar, Z.; Bodis, J.; Csabai, T.; Farkas, N.; Godony, K.; Varnagy, A.; Buzas, E.; Szekeres-Bartho, J. A simple and rapid flow cytometry-based assay to identify a competent embryo prior to embryo transfer. Sci. Rep. 2017, 7, 39927. [Google Scholar] [CrossRef]
- Feuerstein, P.; Cadoret, V.; Dalbies-Tran, R.; Guerif, F.; Bidault, R.; Royere, D. Gene expression in human cumulus cells: One approach to oocyte competence. Hum. Reprod. 2007, 22, 3069–3077. [Google Scholar] [CrossRef]
- Assou, S.; Haouzi, D.; Mahmoud, K.; Aouacheria, A.; Guillemin, Y.; Pantesco, V.; Rème, T.; Dechaud, H.; De Vos, J.; Hamamah, S. A non-invasive test for assessing embryo potential by gene expression profiles of human cumulus cells: A proof of concept study. Mol. Hum. Reprod. 2008, 14, 711–719. [Google Scholar] [CrossRef]
- Le Gac, S.; Nordhoff, V. Microfluidics for mammalian embryo culture and selection: Where do we stand now? Mol. Hum. Reprod. 2017, 23, 213–226. [Google Scholar] [CrossRef]
- Kushnir, V.A.; Smith, G.D.; Adashi, E.Y. The Future of IVF: The New Normal in Human Reproduction. Reprod. Sci. 2022, 29, 849–856. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Types of Stress | Effects on Embryo Development and Long-Term Effects | Definitions | References | |
|---|---|---|---|---|
| In vitro | IVF |
| Adding a defined amount of sperm to the oocyte in a culture medium | [40,51] |
| ICSI |
| A single spermatozoon is injected into the cytoplasm of a mature oocyte. | [52,53] | |
| Culture media composition |
| Variations in nutrient availability are sources of stresses. | [54,55] | |
| Oxygen tension |
| Oxygen concentration during culture and oxidative stress affect gene expression and intracellular signaling. | [56] | |
| Temperature |
| Temperature fluctuations affect gamete and embryo viability. | [57] | |
| PH |
| PH modulates metabolic activity, cellular proliferation, transcriptional activity, protein localization and synthesis. | [36,58] | |
| Light |
| White light exposure of the embryo results in impaired implantation capacity. | [45,49,59] | |
| Substrate stiffness |
| fertilization and embryo development are more successful on a collagen matrix than on a standard polystyrene petri dish. | [60] | |
| Cryopreservation |
| Cryoinjuries including ice crystal formation, structural damage to water bound enzymes, separation of membrane proteins from lipids, altered membrane permeability and osmotic stress due to changes in cell volume should be considered. | [61,62,63] | |
| In vivo | Suboptimal diet |
| Nutritional conditions in utero are associated with glucose intolerance, obesity, and cardiac dysfunction in adulthood. | [64] |
| Maternal diseases |
| Diabetes, hypertension, epilepsy, obesity, and cardiopulmonary disorders known as in vivo stress factors. | [65,66] | |
| Endocrine disruptors |
| Poor oocyte maturation and competency, embryonic defects and poor IVF outcomes are possible complications of endocrine disorders. | [67] | |
| Toxins |
| Tobacco, nicotine can cause delayed migration of embryos from the fallopian tubes into the uterus, growth retardation, pregnancy loss. | [44,68] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmadi, H.; Aghebati-Maleki, L.; Rashidiani, S.; Csabai, T.; Nnaemeka, O.B.; Szekeres-Bartho, J. Long-Term Effects of ART on the Health of the Offspring. Int. J. Mol. Sci. 2023, 24, 13564. https://doi.org/10.3390/ijms241713564
Ahmadi H, Aghebati-Maleki L, Rashidiani S, Csabai T, Nnaemeka OB, Szekeres-Bartho J. Long-Term Effects of ART on the Health of the Offspring. International Journal of Molecular Sciences. 2023; 24(17):13564. https://doi.org/10.3390/ijms241713564
Chicago/Turabian StyleAhmadi, Hamid, Leili Aghebati-Maleki, Shima Rashidiani, Timea Csabai, Obodo Basil Nnaemeka, and Julia Szekeres-Bartho. 2023. "Long-Term Effects of ART on the Health of the Offspring" International Journal of Molecular Sciences 24, no. 17: 13564. https://doi.org/10.3390/ijms241713564