
NLRC5 Deficiency Reduces LPS-Induced Microglial Activation via Inhibition of NF-κB Signaling and Ameliorates Mice’s Depressive-like Behavior
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
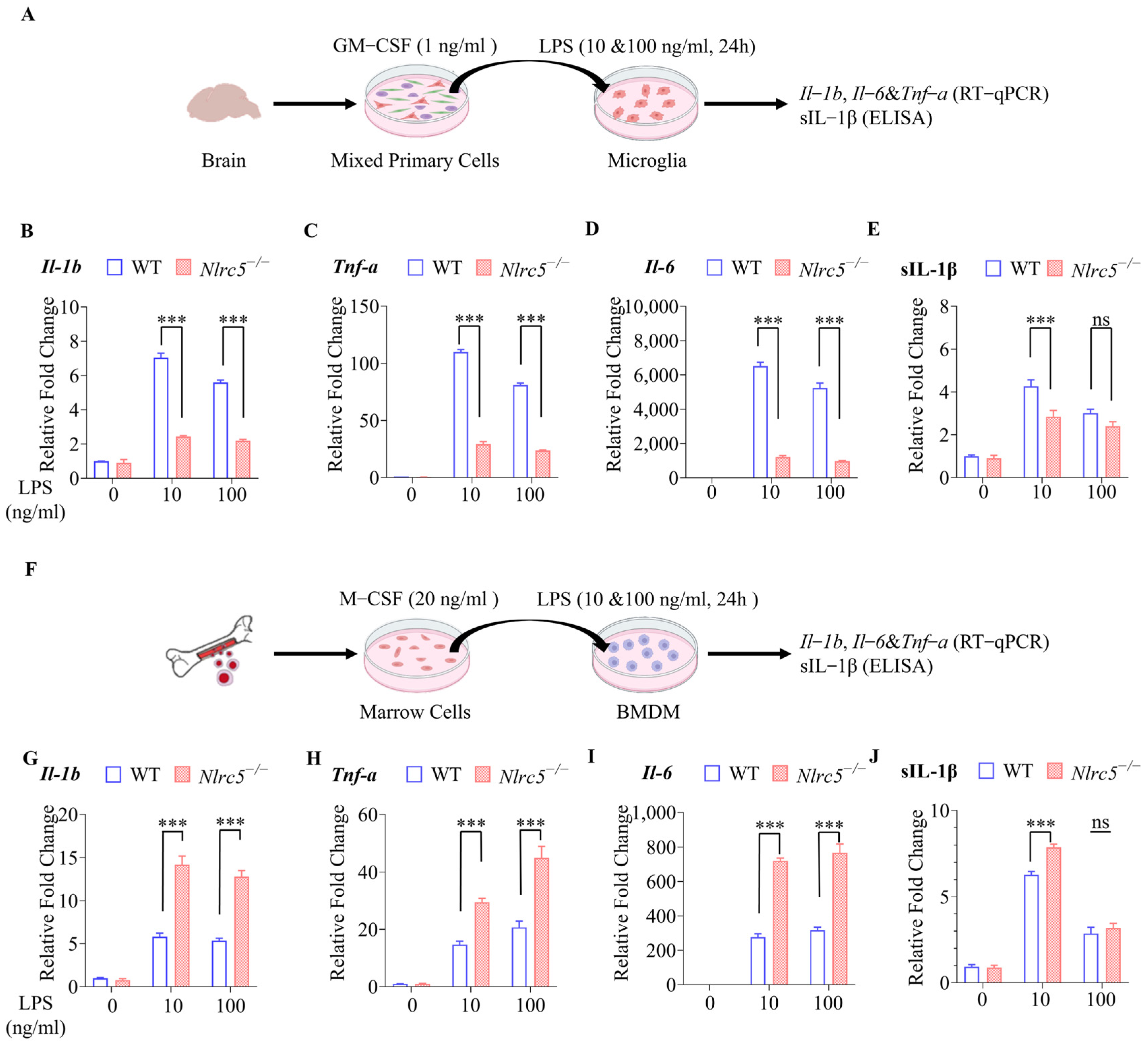
2.1. Nlrc5 Deficiency Inhibited the Expression of Pro-Inflammatory Cytokines under LPS Stimulation in Primary Cultured Microglia but Not in BMDMs
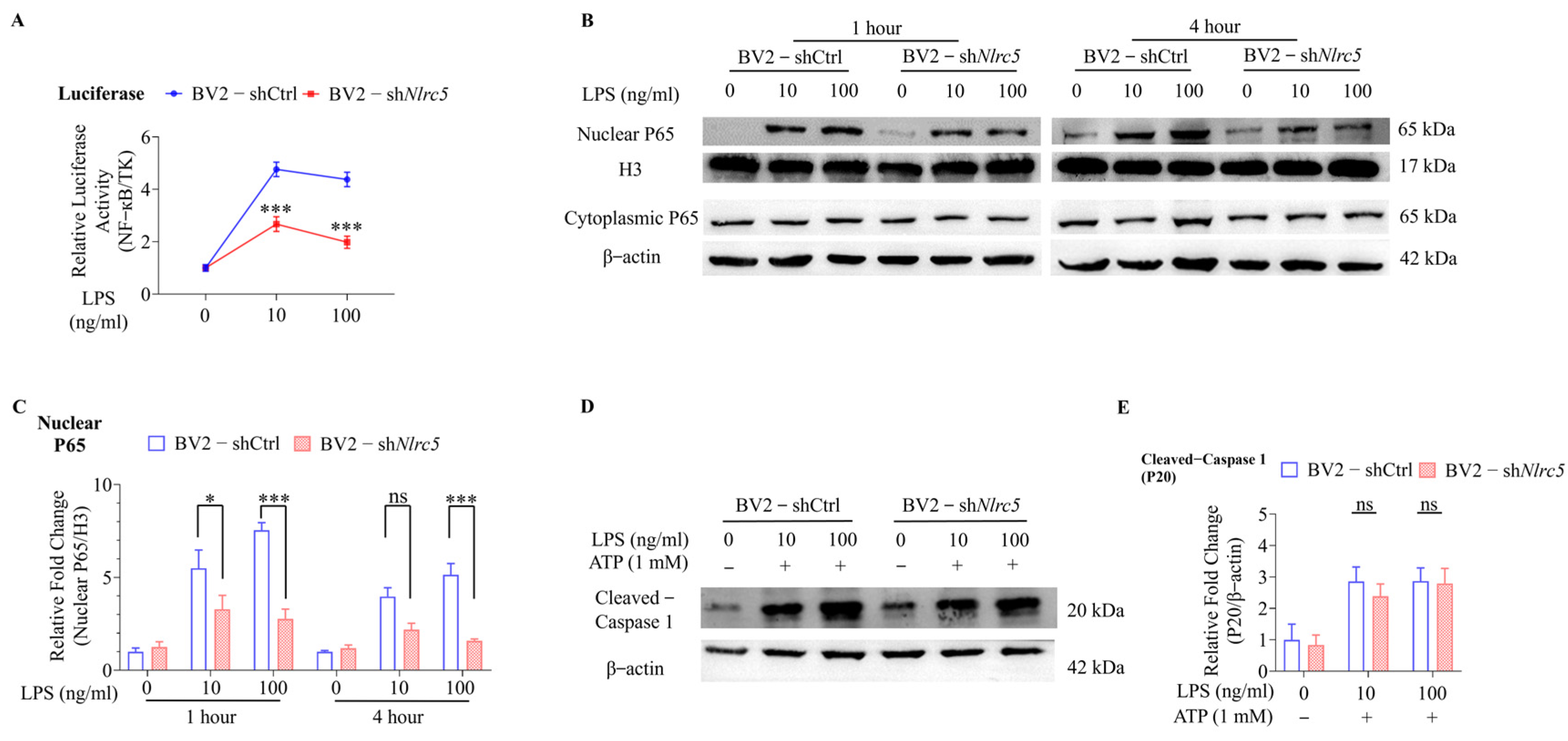
2.2. NLRC5 Regulates Microglial Activation via the NF-κB Signaling Pathway
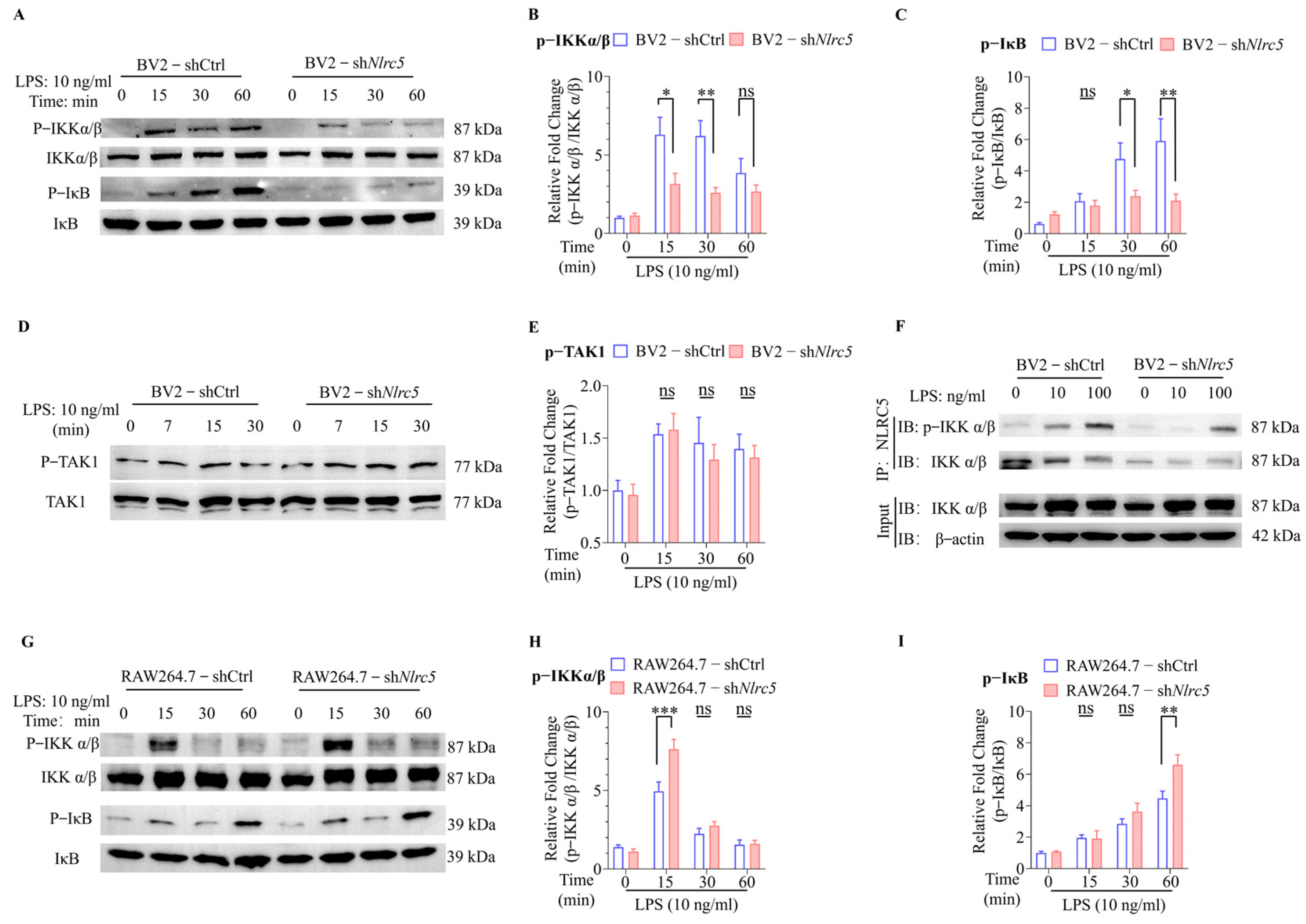
2.3. NLRC5 Regulates Microglial Activation by Promoting the Phosphorylation of IKKα/β
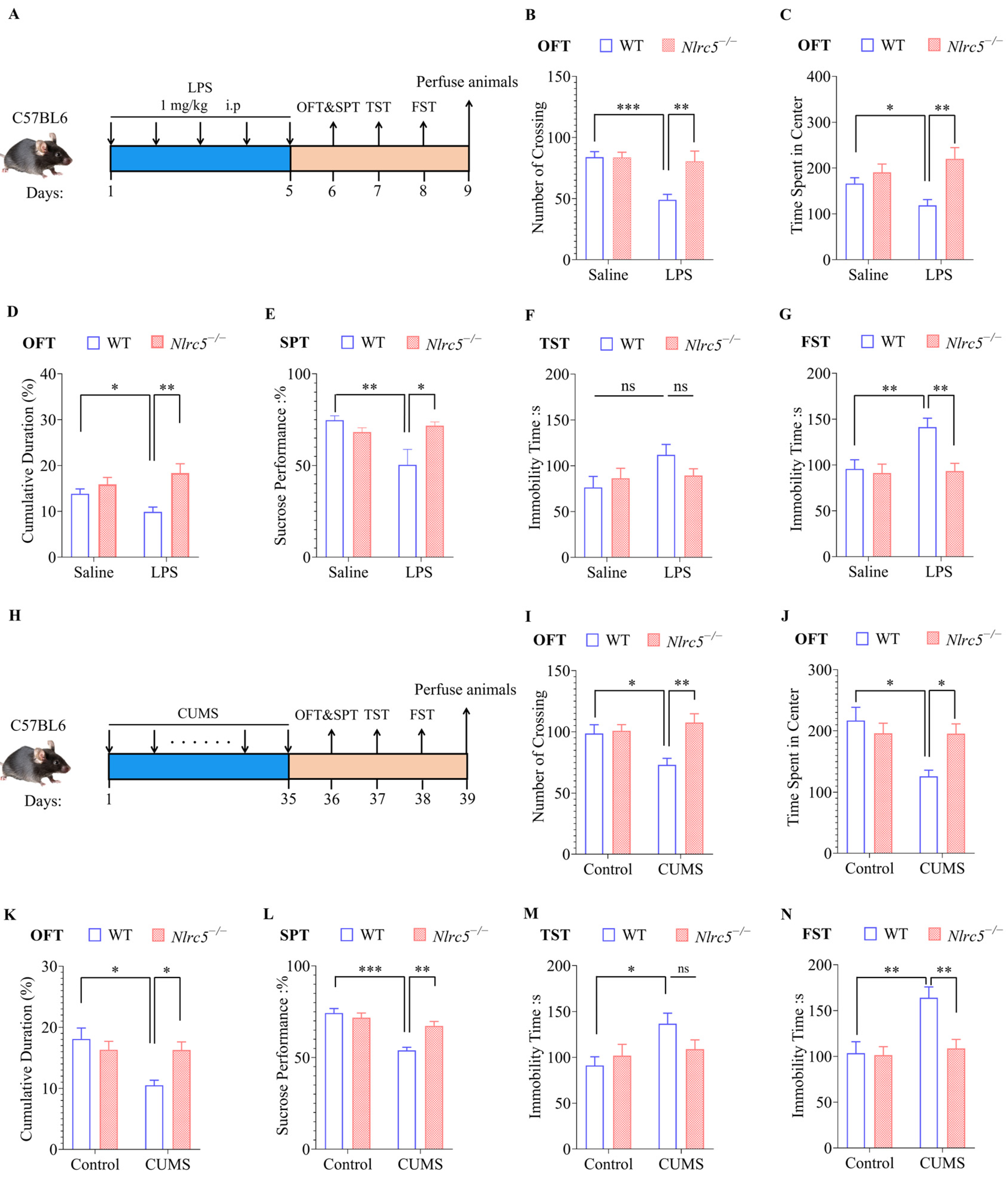
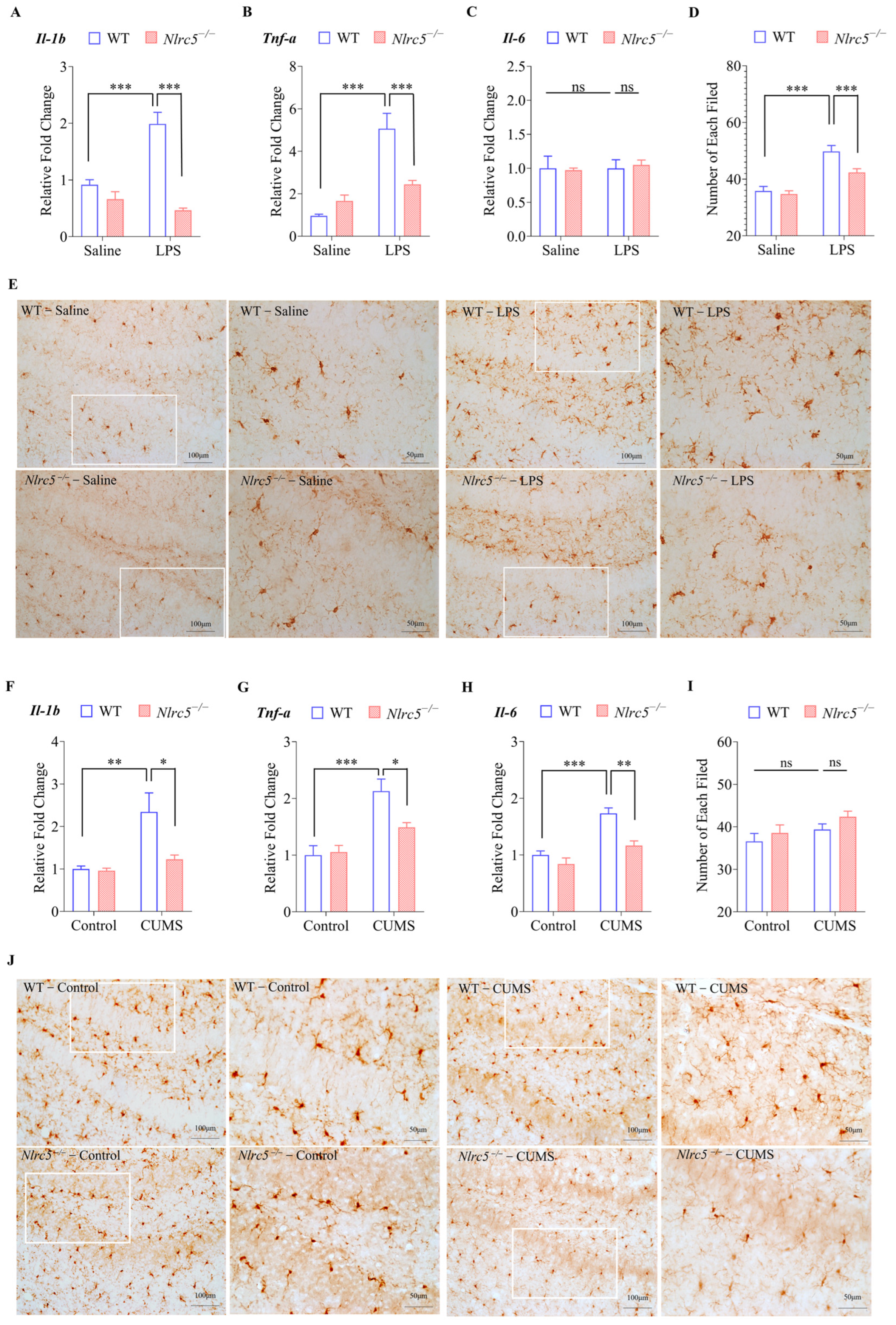
2.4. Nlrc5 Deficiency Ameliorated LPS- or CUMS-Induced Depressive and Anxiety-like Behaviors in Mice
2.5. Nlrc5 Deficiency Inhibited Microglial Activation Induced by LPS or CUMS
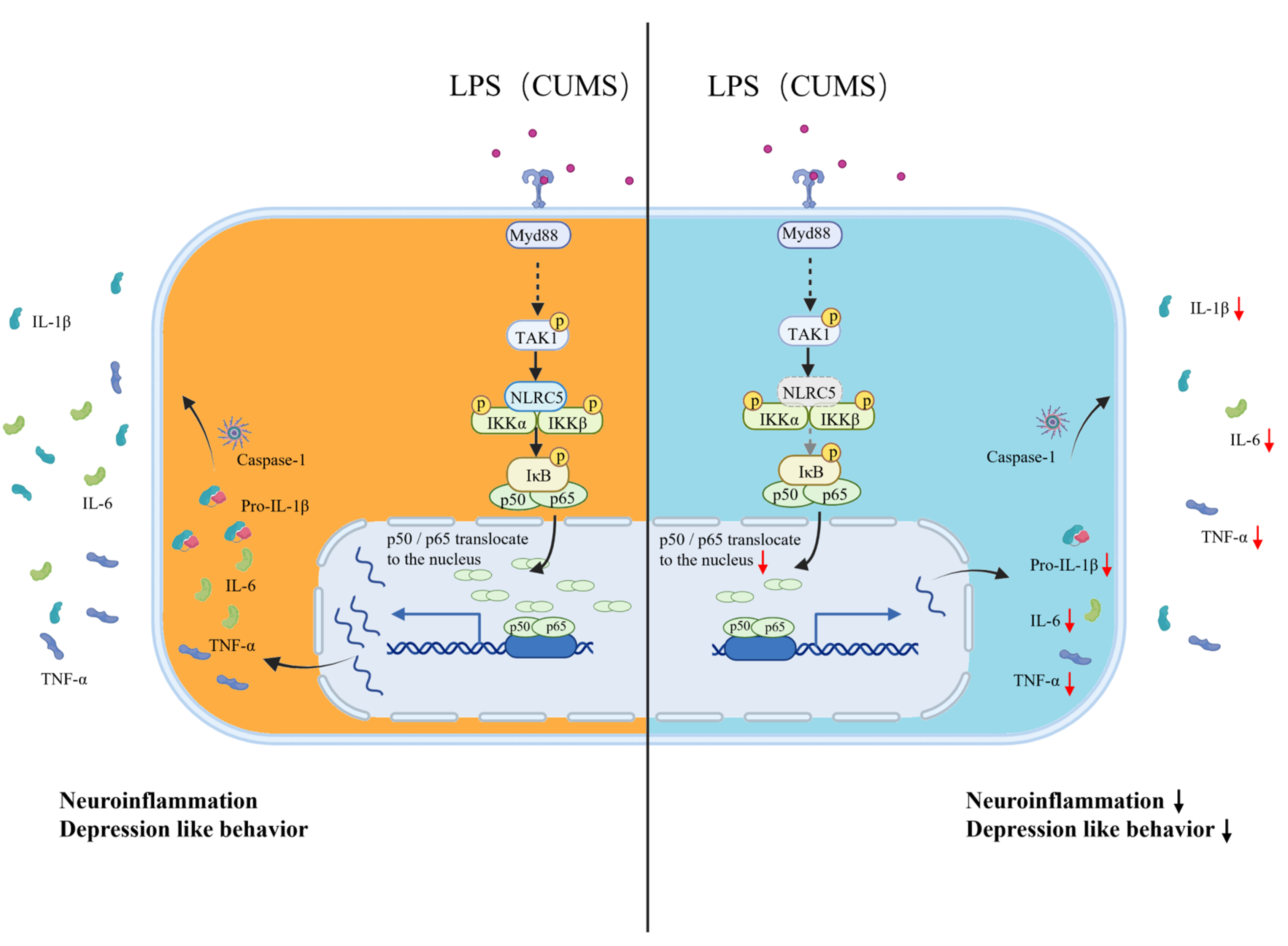
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cell Culture
4.3. Western Blotting
4.4. Real-Time Quantitative PCR (RT-qPCR)
4.5. Enzyme-Linked Immunosorbent Assay (ELISA)
4.6. Plasmid Transfection and Luciferase Reporter Assay
4.7. Immunoprecipitation
4.8. Mouse Model
4.9. Behavior Test
4.10. Immunostaining and Image Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borst, K.; Dumas, A.A.; Prinz, M. Microglia: Immune and non-immune functions. Immunity 2021, 54, 2194–2208. [Google Scholar]
- Wake, H.; Moorhouse, A.J.; Miyamoto, A.; Nabekura, J. Microglia: Actively surveying and shaping neuronal circuit structure and function. Trends Neurosci. 2013, 36, 209–217. [Google Scholar]
- Sharon, G.; Sampson, T.R.; Geschwind, D.H.; Mazmanian, S.K. The Central Nervous System and the Gut Microbiome. Cell 2016, 167, 915–932. [Google Scholar]
- Zrzavy, T.; Hametner, S.; Wimmer, I.; Butovsky, O.; Weiner, H.L.; Lassmann, H. Loss of ‘homeostatic’ microglia and patterns of their activation in active multiple sclerosis. Brain 2017, 140, 1900–1913. [Google Scholar]
- Ménard, C.; Hodes, G.E.; Russo, S.J. Pathogenesis of depression: Insights from human and rodent studies. Neuroscience 2016, 321, 138–162. [Google Scholar] [PubMed]
- Rajendran, L.; Paolicelli, R.C. Microglia-Mediated Synapse Loss in Alzheimer’ Disease. J. Neurosci. 2018, 38, 2911–2919. [Google Scholar] [PubMed]
- Wang, J.Q.; Liu, Y.R.; Xia, Q.; Chen, R.N.; Liang, J.; Xia, Q.R.; Li, J. Emerging Roles for NLRC5 in Immune Diseases. Front. Pharmacol. 2019, 10, 1352. [Google Scholar] [PubMed]
- Wu, Y.; Shi, T.; Li, J. NLRC5: A paradigm for NLRs in immunological and inflammatory reaction. Cancer Lett. 2019, 451, 92–99. [Google Scholar] [PubMed]
- Wang, C.; Fan, L.; Khawaja, R.R.; Liu, B.; Zhan, L.; Kodama, L.; Chin, M.; Li, Y.; Le, D.; Zhou, Y.; et al. Microglial NF-κB drives tau spreading and toxicity in a mouse model of tauopathy. Nat. Commun. 2022, 13, 1969. [Google Scholar] [PubMed]
- Cui, J.; Zhu, L.; Xia, X.; Wang, H.Y.; Legras, X.; Hong, J.; Ji, J.; Shen, P.; Zheng, S.; Chen, Z.J.; et al. NLRC5 negatively regulates the NF-kappaB and type I interferon signaling pathways. Cell 2010, 141, 483–496. [Google Scholar]
- Wang, Y.; Huang, C.; Bian, E.; Lei, T.; Lv, X.; Li, J. NLRC5 negatively regulates inflammatory responses in LPS-induced acute lung injury through NF-κB and p38 MAPK signal pathways. Toxicol. Appl. Pharmacol. 2020, 403, 115150. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Yao, J.N.; Zhang, L.F.; Wang, C.F.; Zhang, X.X.; Gao, B. Effect of NLRC5 on activation and reversion of hepatic stellate cells by regulating the nuclear factor-κB signaling pathway. World J. Gastroenterol. 2019, 25, 3044–3055. [Google Scholar] [CrossRef] [PubMed]
- Luan, P.; Zhuang, J.; Zou, J.; Li, H.; Shuai, P.; Xu, X.; Zhao, Y.; Kou, W.; Ji, S.; Peng, A.; et al. NLRC5 deficiency ameliorates diabetic nephropathy through alleviating inflammation. Faseb J. 2018, 32, 1070–1084. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Fu, Y.; Sheng, L.; Hu, Y.; Su, L.; Luo, J.; Yan, C.; Chi, W. The Regulatory NOD-Like Receptor NLRC5 Promotes Ganglion Cell Death in Ischemic Retinopathy by Inducing Microglial Pyroptosis. Front. Cell Dev. Biol. 2021, 9, 669696. [Google Scholar] [CrossRef]
- Hu, Y.; Shen, Y.; Wu, X.; Ba, R.; Xu, H.; Lu, K.; Shao, Y.; Sun, C.; Zhang, Y.; Miao, F.; et al. Expression pattern of NLRC5 in the postnatal mouse brain. Acta Histochem. 2022, 124, 151939. [Google Scholar] [CrossRef]
- Liu, Z.; Shen, C.; Li, H.; Tong, J.; Wu, Y.; Ma, Y.; Wang, J.; Wang, Z.; Li, Q.; Zhang, X.; et al. NOD-like receptor NLRC5 promotes neuroinflammation and inhibits neuronal survival in Parkinson’s disease models. J. Neuroinflammation 2023, 20, 96. [Google Scholar]
- Hoogland, I.C.; Houbolt, C.; van Westerloo, D.J.; van Gool, W.A.; van de Beek, D. Systemic inflammation and microglial activation: Systematic review of animal experiments. J. Neuroinflamm. 2015, 12, 114. [Google Scholar] [CrossRef]
- Agostinho, P.; Madeira, D.; Dias, L.; Simões, A.P.; Cunha, R.A.; Canas, P.M. Purinergic signaling orchestrating neuron-glia communication. Pharmacol. Res. 2020, 162, 105253. [Google Scholar] [PubMed]
- Wang, H.; He, Y.; Sun, Z.; Ren, S.; Liu, M.; Wang, G.; Yang, J. Microglia in depression: An overview of microglia in the pathogenesis and treatment of depression. J. Neuroinflamm. 2022, 19, 132. [Google Scholar]
- Iwata, M.; Ota, K.T.; Li, X.Y.; Sakaue, F.; Li, N.; Dutheil, S.; Banasr, M.; Duric, V.; Yamanashi, T.; Kaneko, K.; et al. Psychological Stress Activates the Inflammasome via Release of Adenosine Triphosphate and Stimulation of the Purinergic Type 2X7 Receptor. Biol. Psychiatry 2016, 80, 12–22. [Google Scholar]
- Borbély, É.; Simon, M.; Fuchs, E.; Wiborg, O.; Czéh, B.; Helyes, Z. Novel drug developmental strategies for treatment-resistant depression. Br. J. Pharmacol. 2022, 179, 1146–1186. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, R.; Davis, R.L. Transcriptome sequencing implicates dorsal striatum-specific gene network, immune response and energy metabolism pathways in bipolar disorder. Mol. Psychiatry 2017, 22, 441–449. [Google Scholar] [CrossRef]
- Davis, B.K.; Roberts, R.A.; Huang, M.T.; Willingham, S.B.; Conti, B.J.; Brickey, W.J.; Barker, B.R.; Kwan, M.; Taxman, D.J.; Accavitti-Loper, M.A.; et al. Cutting edge: NLRC5-dependent activation of the inflammasome. J. Immunol. 2011, 186, 1333–1337. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.M.; Ma, X.; Zeng, Z.; Luo, Z.; Li, Y.; Nian, X.; Ma, Y.; Shi, Z.; Song, R.; Zhu, Z.; et al. Senecavirus a 3D Interacts with NLRP3 to Induce IL-1β Production by Activating NF-κB and Ion Channel Signals. Microbiol. Spectr. 2022, 10, e0209721. [Google Scholar] [CrossRef]
- Cserép, C.; Pósfai, B.; Dénes, Á. Shaping Neuronal Fate: Functional Heterogeneity of Direct Microglia-Neuron Interactions. Neuron 2021, 109, 222–240. [Google Scholar] [CrossRef]
- Kuenzel, S.; Till, A.; Winkler, M.; Häsler, R.; Lipinski, S.; Jung, S.; Grötzinger, J.; Fickenscher, H.; Schreiber, S.; Rosenstiel, P. The nucleotide-binding oligomerization domain-like receptor NLRC5 is involved in IFN-dependent antiviral immune responses. J. Immunol. 2010, 184, 1990–2000. [Google Scholar] [CrossRef] [PubMed]
- Benkő, S.; Kovács, E.G.; Hezel, F.; Kufer, T.A. NLRC5 Functions beyond MHC I Regulation—What Do We Know So Far? Front. Immunol. 2017, 8, 150. [Google Scholar] [CrossRef] [PubMed]
- Benko, S.; Magalhaes, J.G.; Philpott, D.J.; Girardin, S.E. NLRC5 limits the activation of inflammatory pathways. J. Immunol. 2010, 185, 1681–1691. [Google Scholar] [CrossRef]
- Periyasamy, P.; Thangaraj, A.; Bendi, V.S.; Buch, S. HIV-1 Tat-mediated microglial inflammation involves a novel miRNA-34a-NLRC5-NFκB signaling axis. Brain Behav. Immun. 2019, 80, 227–237. [Google Scholar] [CrossRef] [PubMed]
- Epelman, S.; Lavine, K.J.; Randolph, G.J. Origin and functions of tissue macrophages. Immunity 2014, 41, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.; Cai, C.; Sun, T.; Wang, Q.; Xie, W.; Wang, R.; Cui, J. Reversible ubiquitination shapes NLRC5 function and modulates NF-κB activation switch. J. Cell Biol. 2015, 211, 1025–1040. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.K.; Miller, S.D. D. Microglia initiate central nervous system innate and adaptive immune responses through multiple TLRs. J. Immunol. 2004, 173, 3916–3924. [Google Scholar] [CrossRef] [PubMed]
- Zusso, M.; Lunardi, V.; Franceschini, D.; Pagetta, A.; Lo, R.; Stifani, S.; Frigo, A.C.; Giusti, P.; Moro, S. Ciprofloxacin and levofloxacin attenuate microglia inflammatory response via TLR4/NF-kB pathway. J. Neuroinflamm. 2019, 16, 148. [Google Scholar] [CrossRef]
- Kreisel, T.; Frank, M.G.; Licht, T.; Reshef, R.; Ben-Menachem-Zidon, O.; Baratta, M.V.; Maier, S.F.; Yirmiya, R. Dynamic microglial alterations underlie stress-induced depressive-like behavior and suppressed neurogenesis. Mol. Psychiatry 2014, 19, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Gong, Y.; Tong, L.; Yang, R.; Hu, W.; Xu, X.; Wang, W.; Wang, P.; Lu, X.; Gao, M.; Wu, Y.; et al. Dynamic changes in hippocampal microglia contribute to depressive-like behavior induced by early social isolation. Neuropharmacology 2018, 135, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Troubat, R.; Leman, S.; Pinchaud, K.; Surget, A.; Barone, P.; Roger, S.; Le Guisquet, A.M.; Brizard, B.; Belzung, C.; Camus, V. Brain immune cells characterization in UCMS exposed P2X7 knock-out mouse. Brain Behav. Immun. 2021, 94, 159–174. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, L.; Bai, Y.; Han, B.; He, C.; Gong, L.; Huang, R.; Shen, L.; Chao, J.; Liu, P.; et al. CircDYM ameliorates depressive-like behavior by targeting miR-9 to regulate microglial activation via HSP90 ubiquitination. Mol. Psychiatry 2020, 25, 1175–1190. [Google Scholar] [CrossRef]
- Paolicelli, R.C.; Sierra, A.; Stevens, B.; Tremblay, M.E.; Aguzzi, A.; Ajami, B.; Amit, I.; Audinat, E.; Bechmann, I.; Bennett, M.; et al. Microglia states and nomenclature: A field at its crossroads. Neuron 2022, 110, 3458–3483. [Google Scholar] [CrossRef]
- Li, L.; Yu, M.; Pang, H.; Chen, L.; Liu, J.; Hou, S. NLRC5 protects neurons from oxygen-glucose deprivation-induced injury through activating the Nrf2/HO-1 pathway. J. Recept. Signal Transduct. Res. 2021, 41, 53–58. [Google Scholar] [CrossRef]
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef]
- Durkee, C.; Kofuji, P.; Navarrete, M.; Araque, A. Astrocyte and neuron cooperation in long-term depression. Trends Neurosci. 2021, 44, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Shen, Y.; Cui, P.; Hu, Y.; Zhang, Y.; Miao, F.; Zhang, A.; Zhang, J. Neuronal NLRC5 regulates MHC class I expression in Neuro-2a cells and also during hippocampal development. J. Neurochem. 2020, 152, 182–194. [Google Scholar] [CrossRef]
- Wen, Z.F.; Liu, H.; Gao, R.; Zhou, M.; Ma, J.; Zhang, Y.; Zhao, J.; Chen, Y.; Zhang, T.; Huang, F.; et al. Tumor cell-released autophagosomes (TRAPs) promote immunosuppression through induction of M2-like macrophages with increased expression of PD-L1. J. Immunother. Cancer 2018, 6, 151. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, C.; Shen, Y.; Liu, P.; Shen, Y.; Hu, Y.; Li, P.; Zhang, Y.; Miao, F.; Zhang, J. NLRC5 Deficiency Reduces LPS-Induced Microglial Activation via Inhibition of NF-κB Signaling and Ameliorates Mice’s Depressive-like Behavior. Int. J. Mol. Sci. 2023, 24, 13265. https://doi.org/10.3390/ijms241713265
Sun C, Shen Y, Liu P, Shen Y, Hu Y, Li P, Zhang Y, Miao F, Zhang J. NLRC5 Deficiency Reduces LPS-Induced Microglial Activation via Inhibition of NF-κB Signaling and Ameliorates Mice’s Depressive-like Behavior. International Journal of Molecular Sciences. 2023; 24(17):13265. https://doi.org/10.3390/ijms241713265
Chicago/Turabian StyleSun, Chen, Yuqing Shen, Piaopiao Liu, Yi Shen, Yue Hu, Ping Li, Ying Zhang, Fengqin Miao, and Jianqiong Zhang. 2023. "NLRC5 Deficiency Reduces LPS-Induced Microglial Activation via Inhibition of NF-κB Signaling and Ameliorates Mice’s Depressive-like Behavior" International Journal of Molecular Sciences 24, no. 17: 13265. https://doi.org/10.3390/ijms241713265