Canagliflozin Inhibited the Activity of Hemolysin and Reduced the Inflammatory Response Caused by Streptococcus suis

Abstract
:1. Introduction
2. Results
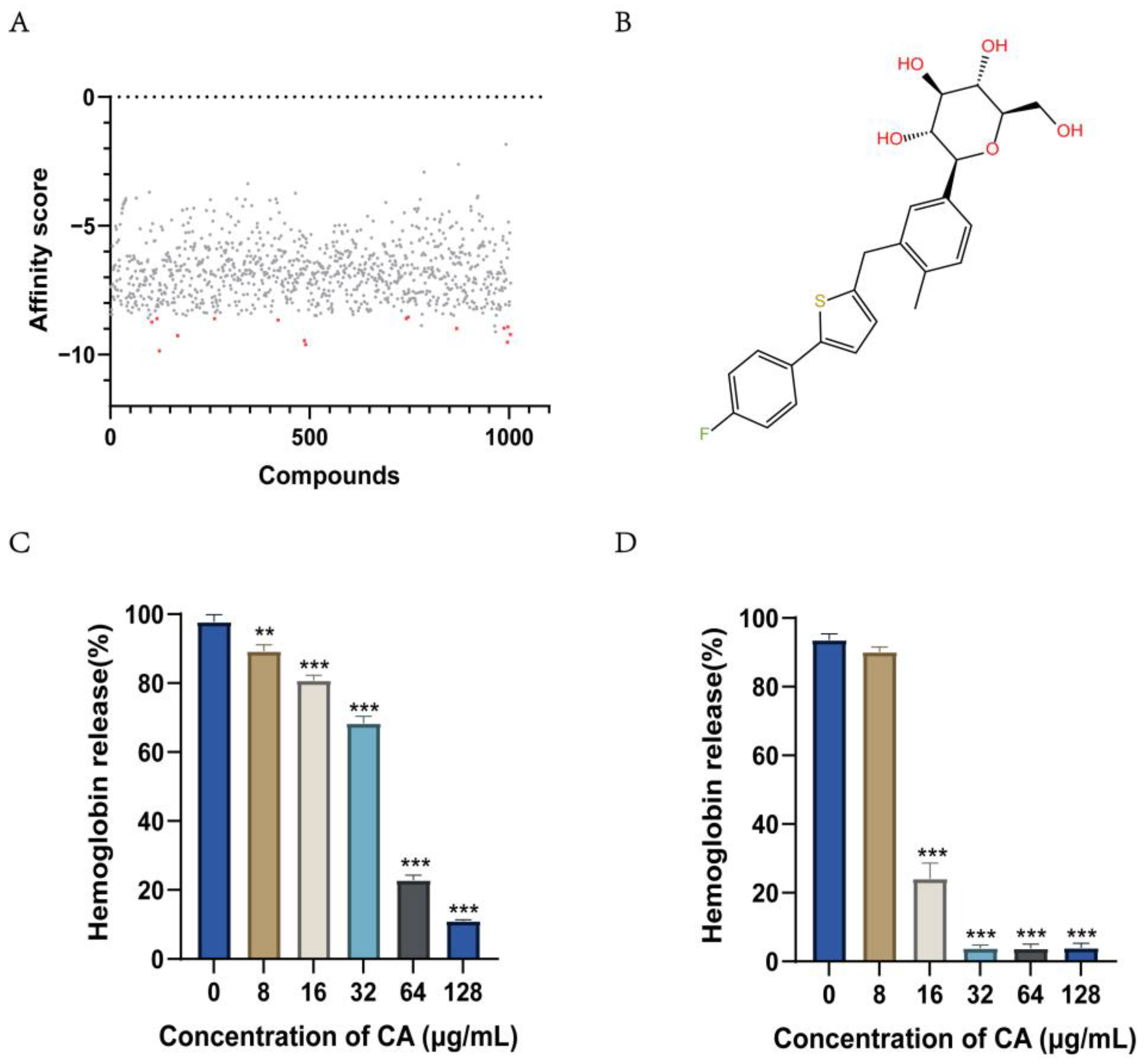
2.1. Screening for Compounds Targeting Hemolysin
2.2. Canagliflozin Inhibits the Activity of the S. suis Hemolysin Protein SLY
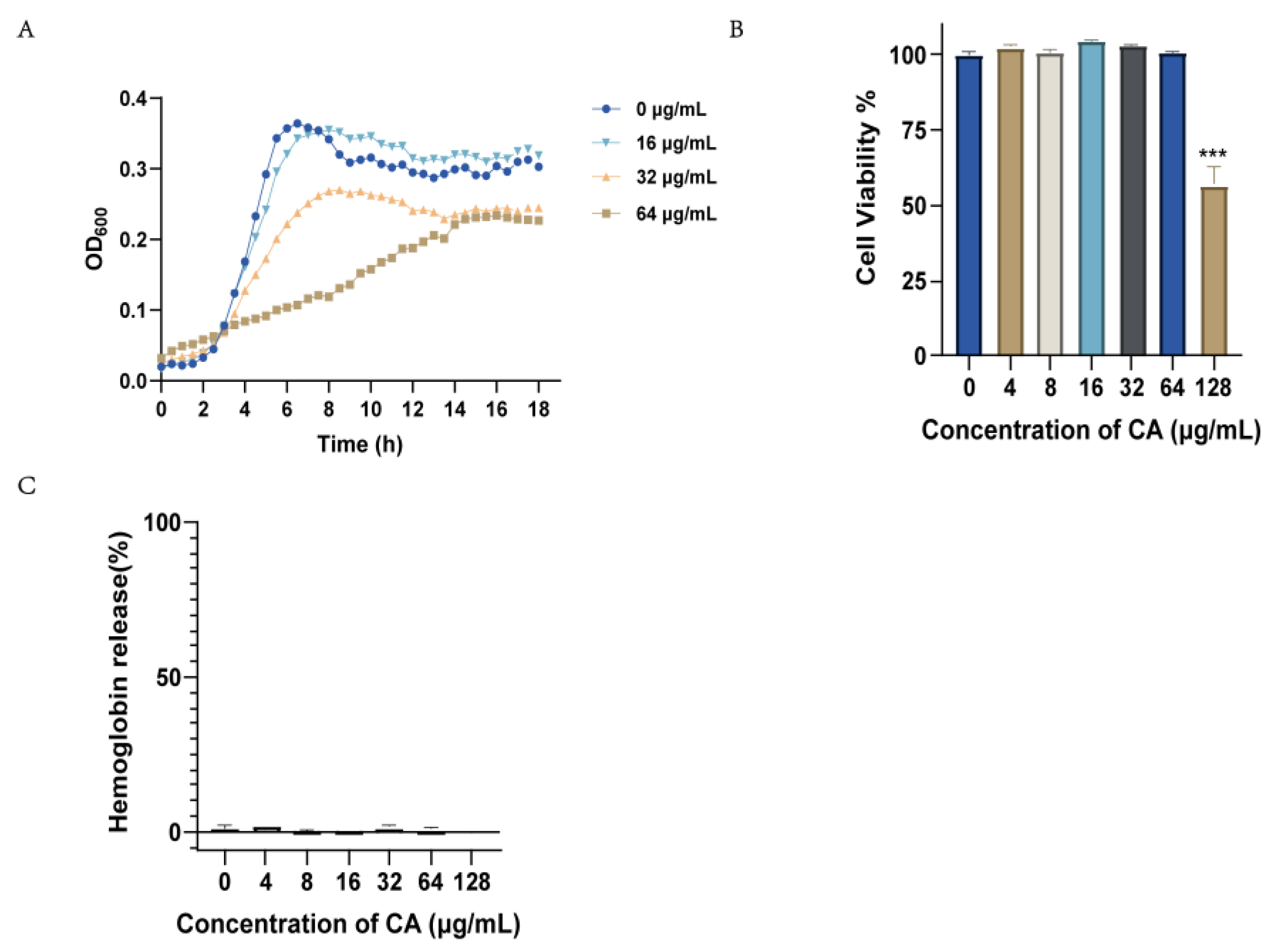
2.3. Canagliflozin Activity against S. suis In Vitro
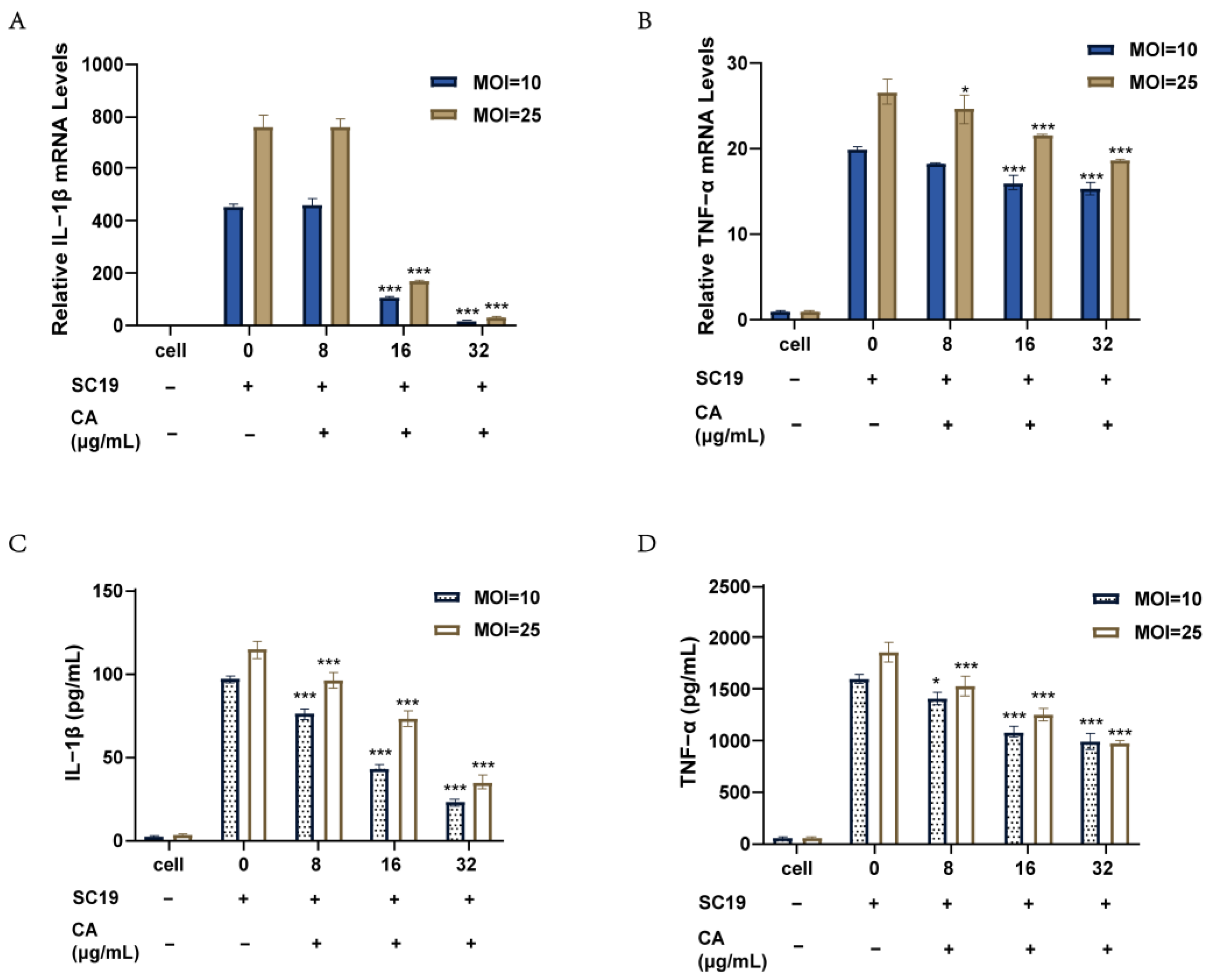
2.4. Canagliflozin Reduced the Inflammatory Response of RAW264.7 Cells Induced by SC19
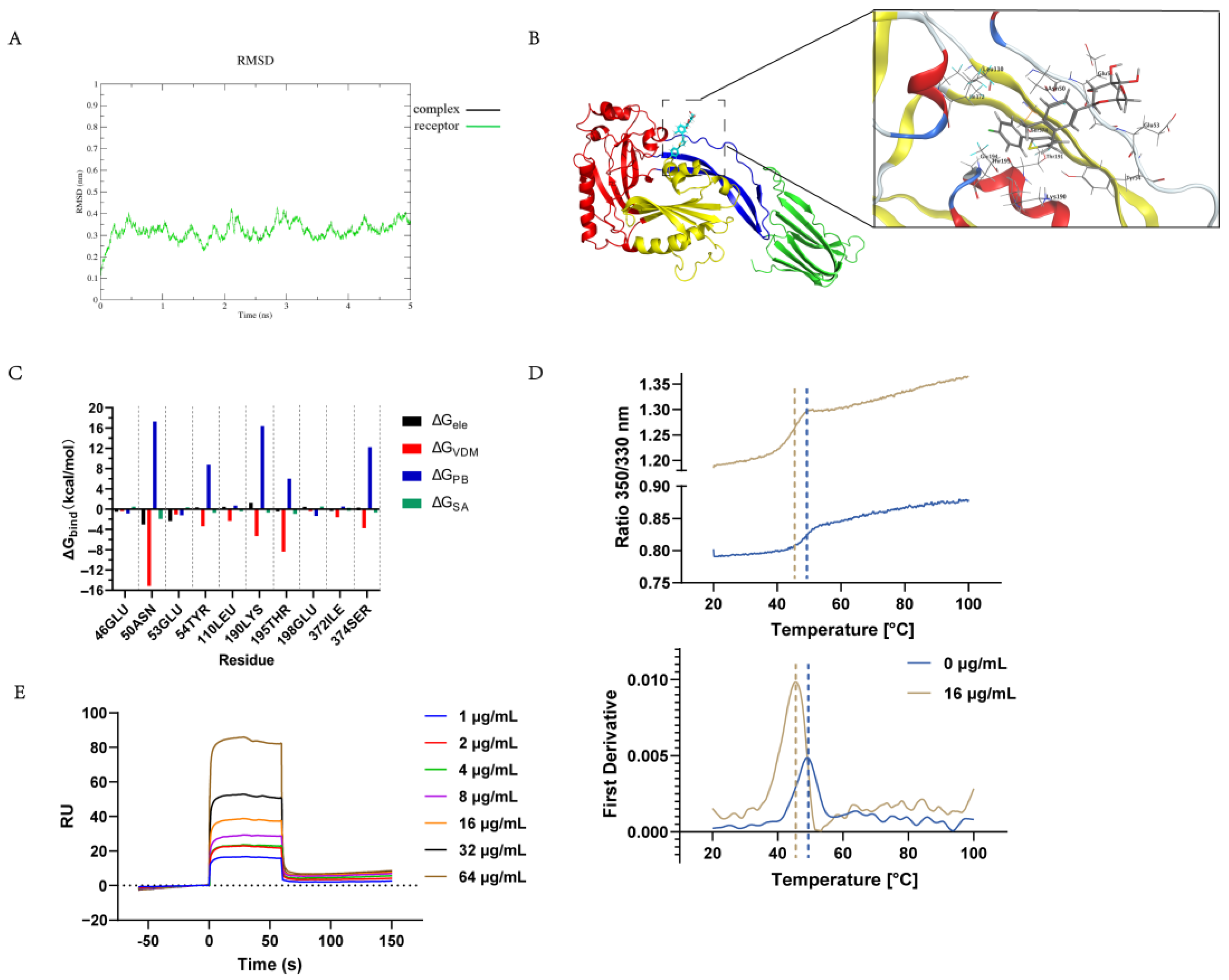
2.5. Determination of the Canagliflozin Targeting of the S. suis Hemolysin Protein In Vitro
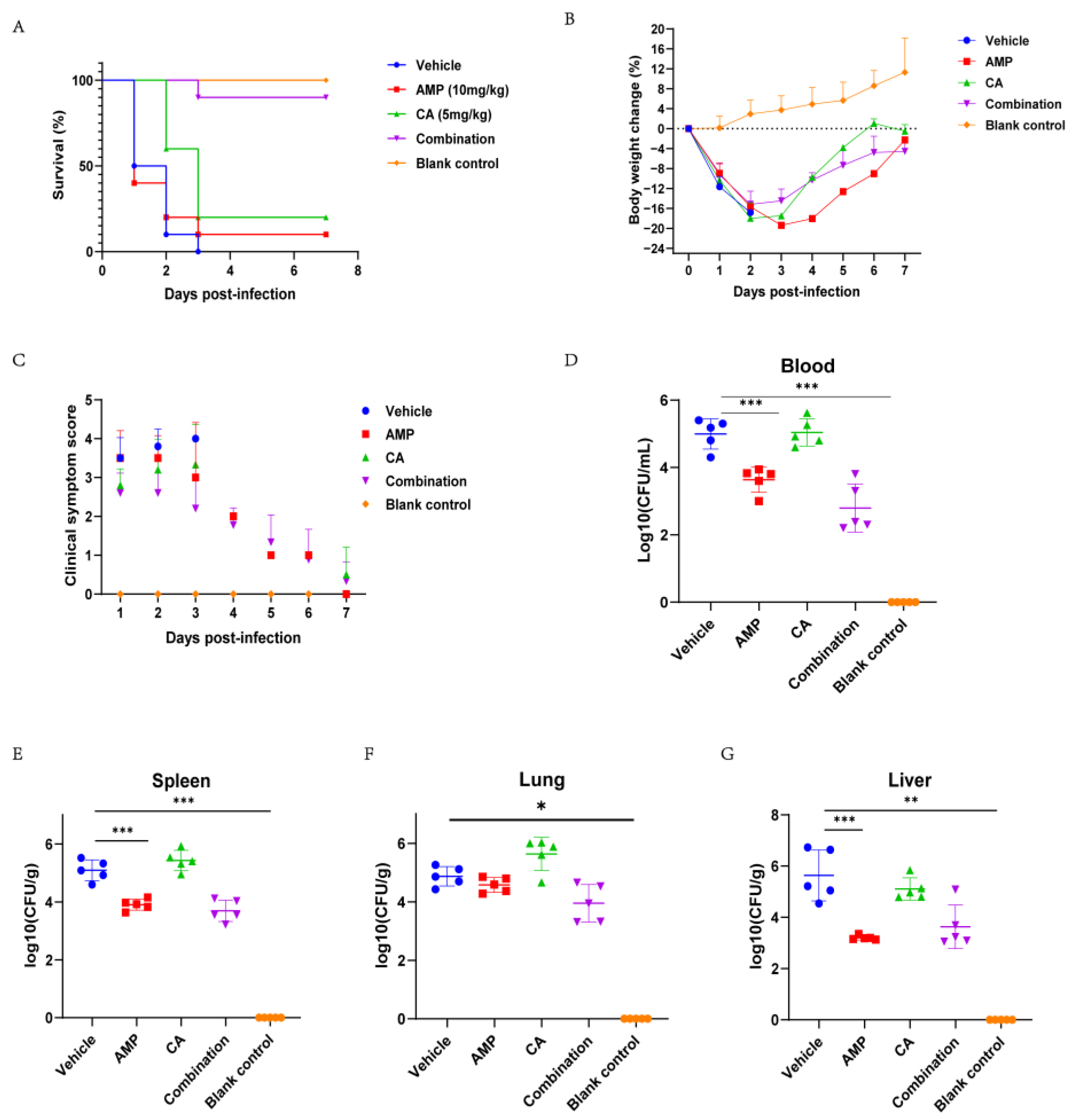
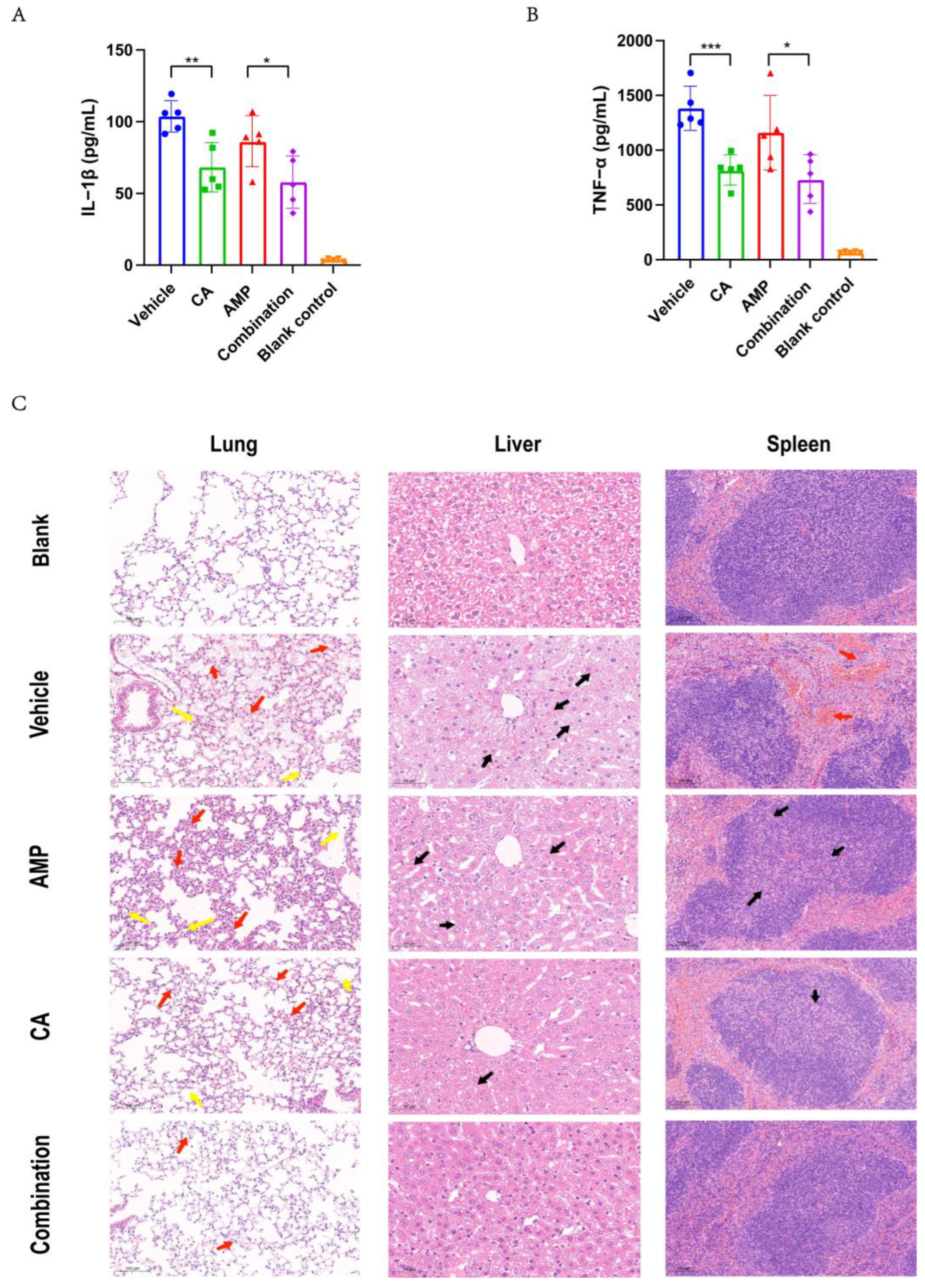
2.6. Canagliflozin Improved the Therapeutic Effect of Ampicillin on SC19 Infection In Vivo
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Growth Conditions, and Preparation of Chemicals
4.2. Screening Method
4.3. Bacterial Culture
4.4. Minimum Inhibitory Concentration Test
4.5. Measurement of Growth Curves
4.6. Recombinant SLY Expression and Purification
4.7. Determination of Antihemolytic Activity
4.8. Cell Infection Study
4.9. Toxicity Test
4.10. Nano Differential Scanning Fluorimetry
4.11. Molecular Dynamics Simulation
4.12. Surface Plasmon Resonance (SPR)
4.13. Animal Studies
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Li, J.; Yang, L.; Zhang, S.; Li, Y.; Jia, X.; Sun, L.; Yin, Y.; Qin, C.; Wang, B.; et al. Assessment of the pathogenesis of Streptococcus suis type 2 infection in piglets for understanding streptococcal toxic shock-like syndrome, meningitis, and sequelae. Vet. Microbiol. 2014, 173, 299–309. [Google Scholar] [CrossRef]
- Werinder, A.; Aspán, A.; Jacobson, M.; Backhans, A.; Sjölund, M.; Guss, B.; Söderlund, R. Genome characteristics related to the virulence of Streptococcus suis in Swedish pigs. Vet. Microbiol. 2023, 284, 109839. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, L.; Lv, W.; Han, L.; Xiang, Y.; Fu, L.; Jin, M.; Zhou, R.; Chen, H.; Zhang, A. An NLRP3 inflammasome-triggered cytokine storm contributes to Streptococcal toxic shock-like syndrome (STSLS). PLoS Pathog. 2019, 15, e1007795. [Google Scholar] [CrossRef] [PubMed]
- Lachance, C.; Gottschalk, M.; Gerber, P.P.; Lemire, P.; Xu, J.; Segura, M. Exacerbated type II interferon response drives hypervirulence and toxic shock by an emergent epidemic strain of Streptococcus suis. Infect. Immun. 2013, 81, 1928–1939. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Jing, H.; Chen, Z.; Zheng, H.; Zhu, X.; Wang, H.; Wang, S.; Liu, L.; Zu, R.; Luo, L.; et al. Human Streptococcus suis outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 914–920. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, D.; Liu, M.; Yang, X.; Zong, B.; Wang, X.; Chen, H.; Bei, W.; Tan, C. Effect of the glycosyltransferases on the capsular polysaccharide synthesis of Streptococcus suis serotype 2. Microbiol. Res. 2016, 185, 45–54. [Google Scholar] [CrossRef]
- Yang, X.P.; Fu, J.Y.; Yang, R.C.; Liu, W.T.; Zhang, T.; Yang, B.; Miao, L.; Dou, B.B.; Tan, C.; Chen, H.C.; et al. EGFR transactivation contributes to neuroinflammation in Streptococcus suis meningitis. J. Neuroinflamm. 2016, 13, 274. [Google Scholar] [CrossRef]
- Lu, H.; Lu, W.; Zhu, Y.; Wang, C.; Shi, L.; Li, X.; Wu, Z.; Wang, G.; Dong, W.; Tan, C.; et al. Auranofin Has Advantages over First-Line Drugs in the Treatment of Severe Streptococcus suis Infections. Antibiotics 2020, 10, 26. [Google Scholar] [CrossRef]
- Jacobs, A.A.; Loeffen, P.L.; van den Berg, A.J.; Storm, P.K. Identification, purification, and characterization of a thiol-activated hemolysin (suilysin) of Streptococcus suis. Infect. Immun. 1994, 62, 1742–1748. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Segura, M. The pathogenesis of the meningitis caused by Streptococcus suis: The unresolved questions. Vet. Microbiol. 2000, 76, 259–272. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Huang, B.; Du, H.; Zhang, X.C.; Xu, J.; Li, X.; Rao, Z. Crystal structure of cytotoxin protein suilysin from Streptococcus suis. Protein Cell 2010, 1, 96–105. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Niu, X.; Li, G.; Deng, X.; Wang, J. Amentoflavone Ameliorates Streptococcus suis-Induced Infection In Vitro and In Vivo. Appl. Environ. Microbiol. 2018, 84, e01804-18. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zong, B.; Wang, X.; Zhu, Y.; Hu, L.; Li, P.; Zhang, A.; Chen, H.; Liu, M.; Tan, C. Fisetin Lowers Streptococcus suis serotype 2 Pathogenicity in Mice by Inhibiting the Hemolytic Activity of Suilysin. Front. Microbiol. 2018, 9, 1723. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Shen, X.; Wei, Y.; Si, X.; Deng, X.; Wang, J. Quercetin reduces Streptococcus suis virulence by inhibiting suilysin activity and inflammation. Int. Immunopharmacol. 2019, 69, 71–78. [Google Scholar] [CrossRef]
- Lu, H.; Wang, C.; Lu, W.; Li, X.; Wu, Z.; Wang, G.; Dong, W.; Tan, C.; Liu, M. Apigenin and Ampicillin as Combined Strategy to Treat Severe Streptococcus suis Infection. Molecules 2021, 26, 1980. [Google Scholar] [CrossRef]
- Escaich, S. Antivirulence as a new antibacterial approach for chemotherapy. Curr. Opin. Chem. Biol. 2008, 12, 400–408. [Google Scholar] [CrossRef]
- Li, Q.; Wang, L.; Xu, J.; Liu, S.; Song, Z.; Chen, T.; Deng, X.; Wang, J.; Lv, Q. Quercitrin Is a Novel Inhibitor of Salmonella enterica Serovar Typhimurium Type III Secretion System. Molecules 2023, 28, 5455. [Google Scholar] [CrossRef]
- Kim, Y.J.; Cubitt, B.; Chen, E.; Hull, M.V.; Chatterjee, A.K.; Cai, Y.; Kuhn, J.H.; de la Torre, J.C. The ReFRAME library as a comprehensive drug repurposing library to identify mammarenavirus inhibitors. Antiviral. Res. 2019, 169, 104558. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, Y.; Xu, Z.; Ma, X.; Chen, X.; Liu, W. Repurposing of the gold drug auranofin and a review of its derivatives as antibacterial therapeutics. Drug. Discov. Today 2022, 27, 1961–1973. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Q.; Wang, R.; Wang, H.; Wong, Y.T.; Wang, M.; Hao, Q.; Yan, A.; Kao, R.Y.; Ho, P.L.; et al. Resensitizing carbapenem- and colistin-resistant bacteria to antibiotics using auranofin. Nat. Commun. 2020, 11, 5263. [Google Scholar] [CrossRef]
- Wang, C.; Lu, H.; Li, X.; Zhu, Y.; Ji, Y.; Lu, W.; Wang, G.; Dong, W.; Liu, M.; Wang, X.; et al. Identification of an anti-virulence drug that reverses antibiotic resistance in multidrug resistant bacteria. Biomed. Pharmacother. 2022, 153, 113334. [Google Scholar] [CrossRef]
- Zuo, J.; Shen, Y.; Wang, H.; Gao, S.; Yuan, S.; Song, D.; Wang, Y.; Wang, Y. Effects of metformin on Streptococcus suis LuxS/AI-2 quorum sensing system and biofilm formation. Microb. Pathog. 2023, 181, 106183. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, Y.; Yang, K.; Li, R.; Xiao, X.; Zhu, K.; Wang, Z. Metformin Restores Tetracyclines Susceptibility against Multidrug Resistant Bacteria. Adv. Sci. 2020, 7, 1902227. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef]
- Heidt, M.C.; Mohamed, W.; Hain, T.; Vogt, P.R.; Chakraborty, T.; Domann, E. Human infective endocarditis caused by Streptococcus suis serotype 2. J. Clin. Microbiol. 2005, 43, 4898–4901. [Google Scholar] [CrossRef] [PubMed]
- Dobrindt, U.; Hacker, J. Targeting virulence traits: Potential strategies to combat extraintestinal pathogenic E. coli infections. Curr. Opin. Microbiol. 2008, 11, 409–413. [Google Scholar] [CrossRef] [PubMed]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug. Discov 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Takeuchi, D.; Akeda, Y.; Nakayama, T.; Kerdsin, A.; Sano, Y.; Kanda, T.; Hamada, S.; Dejsirilert, S.; Oishi, K. The contribution of suilysin to the pathogenesis of Streptococcus suis meningitis. J. Infect. Dis. 2014, 209, 1509–1519. [Google Scholar] [CrossRef]
- Johnston, R.; Uthman, O.; Cummins, E.; Clar, C.; Royle, P.; Colquitt, J.; Tan, B.K.; Clegg, A.; Shantikumar, S.; Court, R.; et al. Canagliflozin, dapagliflozin and empagliflozin monotherapy for treating type 2 diabetes: Systematic review and economic evaluation. Health Technol. Assess. 2017, 21, 1–218. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Arakawa, K.; Ueta, K.; Matsushita, Y.; Kuriyama, C.; Martin, T.; Du, F.; Liu, Y.; Xu, J.; Conway, B.; et al. Effect of canagliflozin on renal threshold for glucose, glycemia, and body weight in normal and diabetic animal models. PLoS ONE 2012, 7, e30555. [Google Scholar] [CrossRef] [PubMed]
- Martinon, F.; Tschopp, J. NLRs join TLRs as innate sensors of pathogens. Trends Immunol. 2005, 26, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Sochocka, M.; Zwolinska, K.; Leszek, J. The Infectious Etiology of Alzheimer’s Disease. Curr. Neuropharmacol. 2017, 15, 996–1009. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zheng, H.; Zhang, J.; Jing, H.; Wang, L.; Xiong, Y.; Wang, W.; Zhou, Z.; Sun, Q.; Luo, X.; et al. Clinical, experimental, and genomic differences between intermediately pathogenic, highly pathogenic, and epidemic Streptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef]
- Segura, M.; Vanier, G.; Al-Numani, D.; Lacouture, S.; Olivier, M.; Gottschalk, M. Proinflammatory cytokine and chemokine modulation by Streptococcus suis in a whole-blood culture system. FEMS Immunol. Med. Microbiol. 2006, 47, 92–106. [Google Scholar] [CrossRef]
- Dominguez-Punaro Mde, L.; Segura, M.; Radzioch, D.; Rivest, S.; Gottschalk, M. Comparison of the susceptibilities of C57BL/6 and A/J mouse strains to Streptococcus suis serotype 2 infection. Infect. Immun. 2008, 76, 3901–3910. [Google Scholar] [CrossRef]
- Sarraju, A.; Li, J.; Cannon, C.P.; Chang, T.I.; Agarwal, R.; Bakris, G.; Charytan, D.M.; de Zeeuw, D.; Greene, T.; Heerspink, H.J.L.; et al. Effects of canagliflozin on cardiovascular, renal, and safety outcomes in participants with type 2 diabetes and chronic kidney disease according to history of heart failure: Results from the CREDENCE trial. Am. Heart J. 2021, 233, 141–148. [Google Scholar] [CrossRef]
- Peters, K.E.; Bringans, S.D.; O’Neill, R.S.; Lumbantobing, T.S.C.; Lui, J.K.C.; Davis, T.M.E.; Hansen, M.K.; Lipscombe, R.J. Canagliflozin Attenuates PromarkerD Diabetic Kidney Disease Risk Prediction Scores. J. Clin. Med. 2023, 12, 3247. [Google Scholar] [CrossRef]
- Jardine, M.J.; Zhou, Z.; Mahaffey, K.W.; Oshima, M.; Agarwal, R.; Bakris, G.; Bajaj, H.S.; Bull, S.; Cannon, C.P.; Charytan, D.M.; et al. Renal, Cardiovascular, and Safety Outcomes of Canagliflozin by Baseline Kidney Function: A Secondary Analysis of the CREDENCE Randomized Trial. J. Am. Soc. Nephrol. JASN 2020, 31, 1128–1139. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoo, H.J.; Park, E.J.; Na, D.H. Nano Differential Scanning Fluorimetry-Based Thermal Stability Screening and Optimal Buffer Selection for Immunoglobulin G. Pharmaceuticals 2021, 15, 29. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Lee, J.M.; Jung, Y.; Habtemariam, T.; Salah, A.W.; Fermin, C.D.; Kim, M. Combination of cysteine- and oligomerization domain-mediated protein immobilization on a surface plasmon resonance (SPR) gold chip surface. Analyst 2011, 136, 2506–2511. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC (μg/mL) | |
|---|---|---|
| Canagliflozin | Ampicillin | |
| S. suis SC19 2 | 128 | <1 |
| S. pneumoniae ATCC 49619 1 | 64 | <1 |
| S. aureus ATCC 25923 1 | 64 | <1 |
| S. aureus USA300 1 | >128 | 64 |
| S. aureus ATCC 43300 1 | 64 | 8 |
| L. monocytogenes ATCC19115 1 | 64 | 2 |
| B. subtilis NCD-2 1 | 64 | 2 |
| S. suis K6 2 | 128 | <1 |
| S. suis 1422 2 | 128 | <1 |
| S. suis 1519 2 | 128 | <1 |
| S. suis 1808028 2 | 128 | <1 |
| S. suis 1765 2 | 128 | 64 |
| S. suis 1801101 2 | >128 | <1 |
| S. suis 1804005 2 | >128 | <1 |
| S. aureus 1605020 2 | 128 | <1 |
| S. aureus 2117 2 | 128 | 64 |
| S. aureus 1802097 2 | >128 | <1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Li, Q.; Zhang, Z.; Wang, C.; Huo, X.; Lai, H.; Lu, H.; Lu, W.; Qian, Y.; Dong, W.; et al. Canagliflozin Inhibited the Activity of Hemolysin and Reduced the Inflammatory Response Caused by Streptococcus suis. Int. J. Mol. Sci. 2023, 24, 13074. https://doi.org/10.3390/ijms241713074
Li X, Li Q, Zhang Z, Wang C, Huo X, Lai H, Lu H, Lu W, Qian Y, Dong W, et al. Canagliflozin Inhibited the Activity of Hemolysin and Reduced the Inflammatory Response Caused by Streptococcus suis. International Journal of Molecular Sciences. 2023; 24(17):13074. https://doi.org/10.3390/ijms241713074
Chicago/Turabian StyleLi, Xiaodan, Qingyuan Li, Zhaoran Zhang, Chenchen Wang, Xinyu Huo, Hongjiang Lai, Hao Lu, Wenjia Lu, Yulin Qian, Wenqi Dong, and et al. 2023. "Canagliflozin Inhibited the Activity of Hemolysin and Reduced the Inflammatory Response Caused by Streptococcus suis" International Journal of Molecular Sciences 24, no. 17: 13074. https://doi.org/10.3390/ijms241713074