
Matrine Targets Intestinal Lactobacillus acidophilus to Inhibit Porcine Circovirus Type 2 Infection in Mice

 ,
,
Abstract
:1. Introduction
2. Results
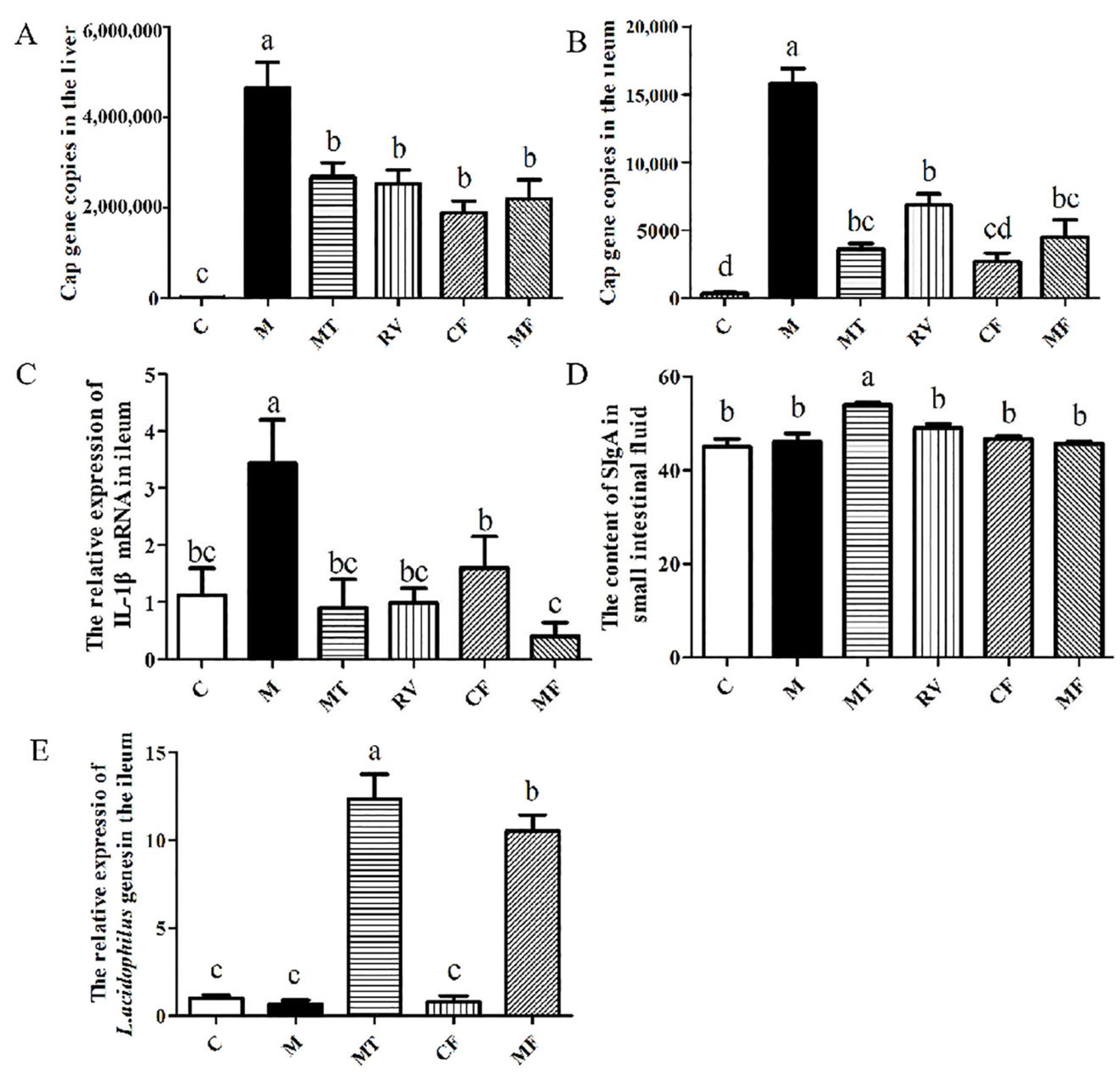
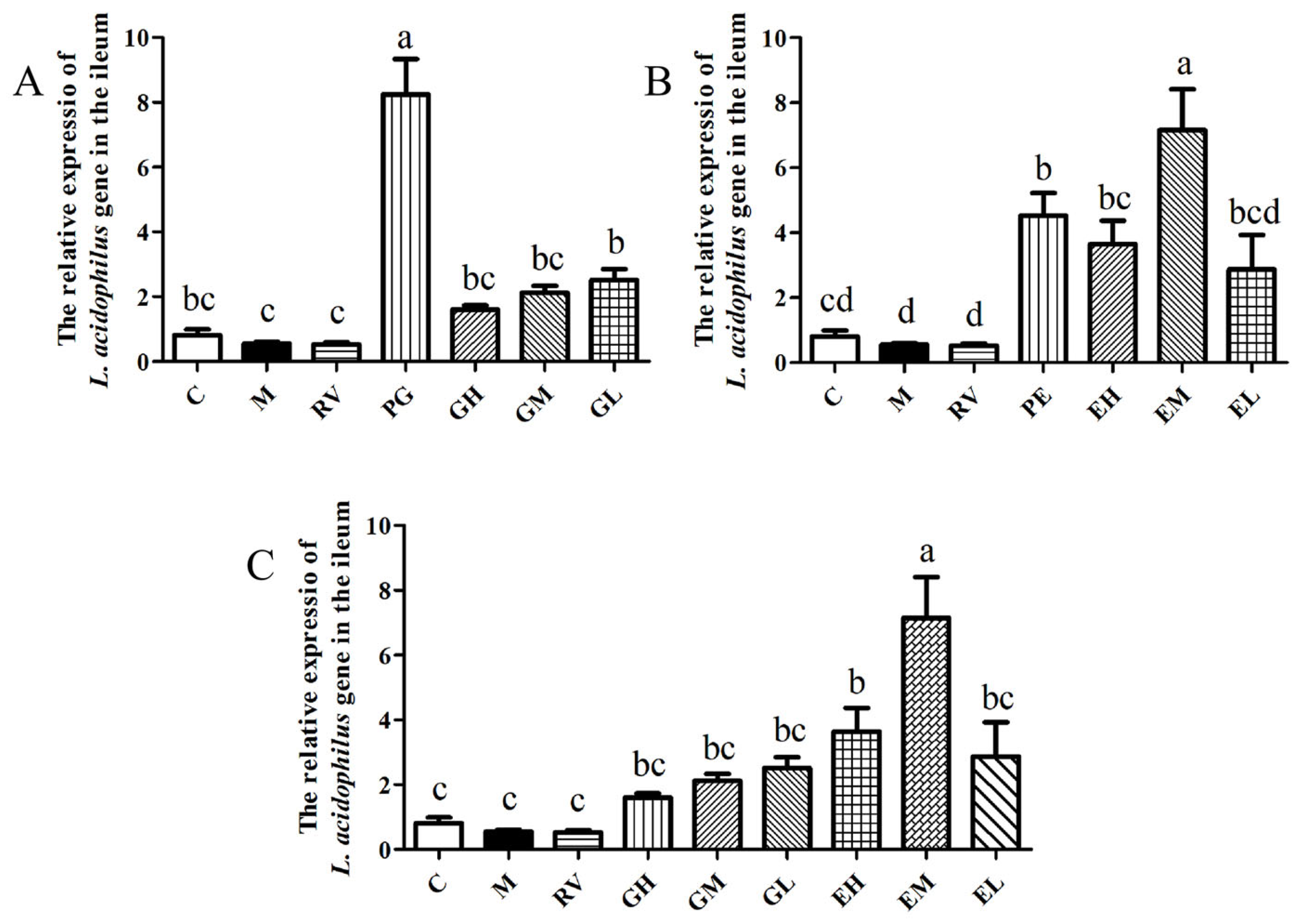
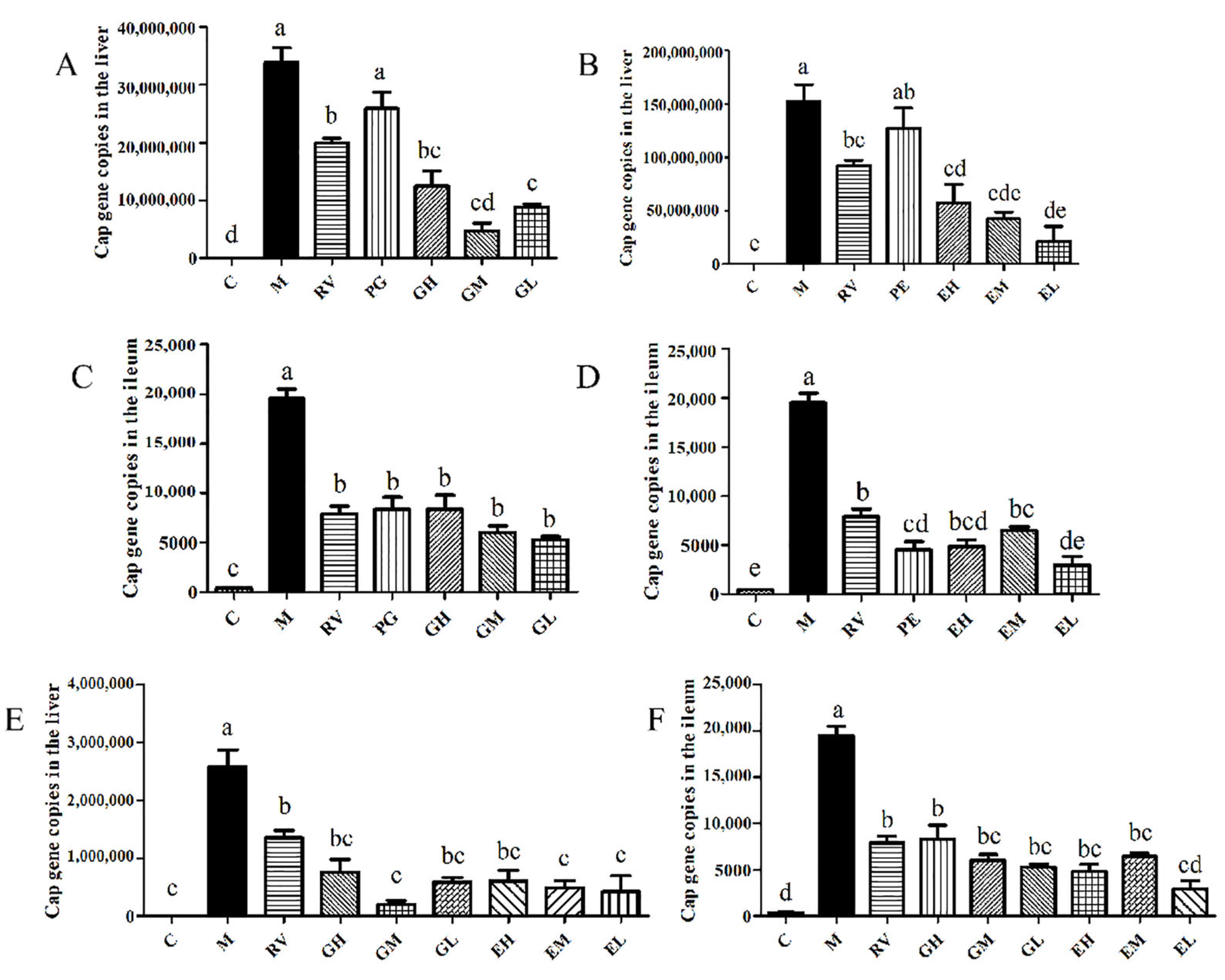
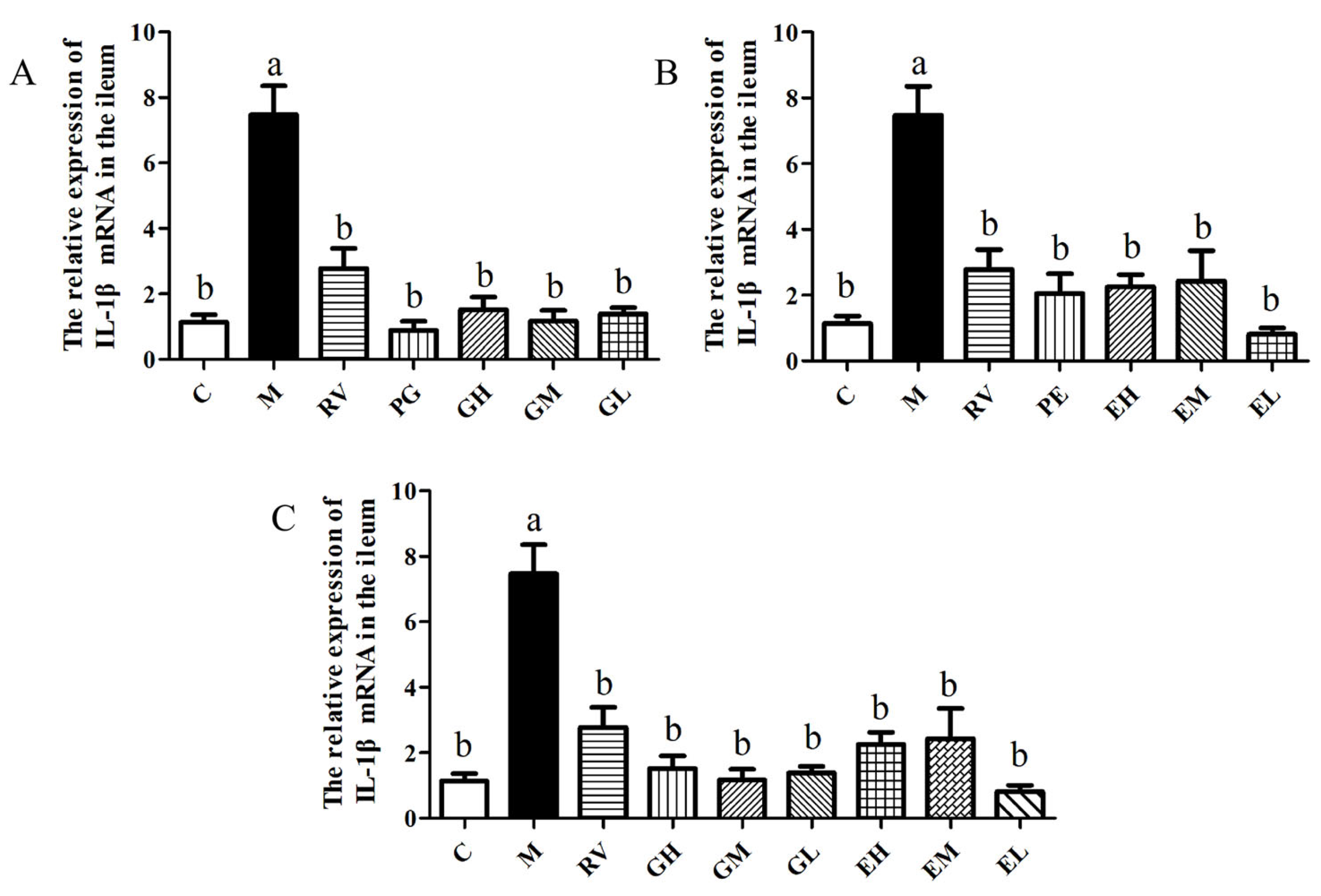
2.1. Matrine-Treated Fecal Bacterial Solution Reduced the Relative Expression of IL-1β mRNA and Promoted the Colonization of L. acidophilus, Indicating that It Has Anti-PCV2 Effects
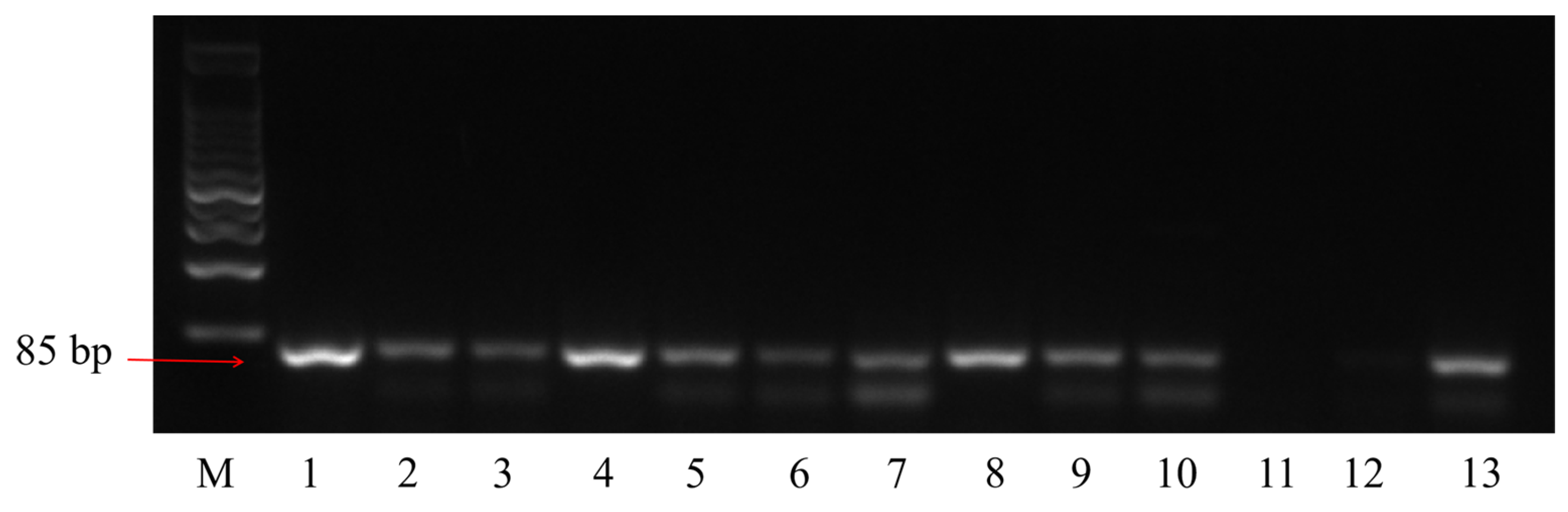
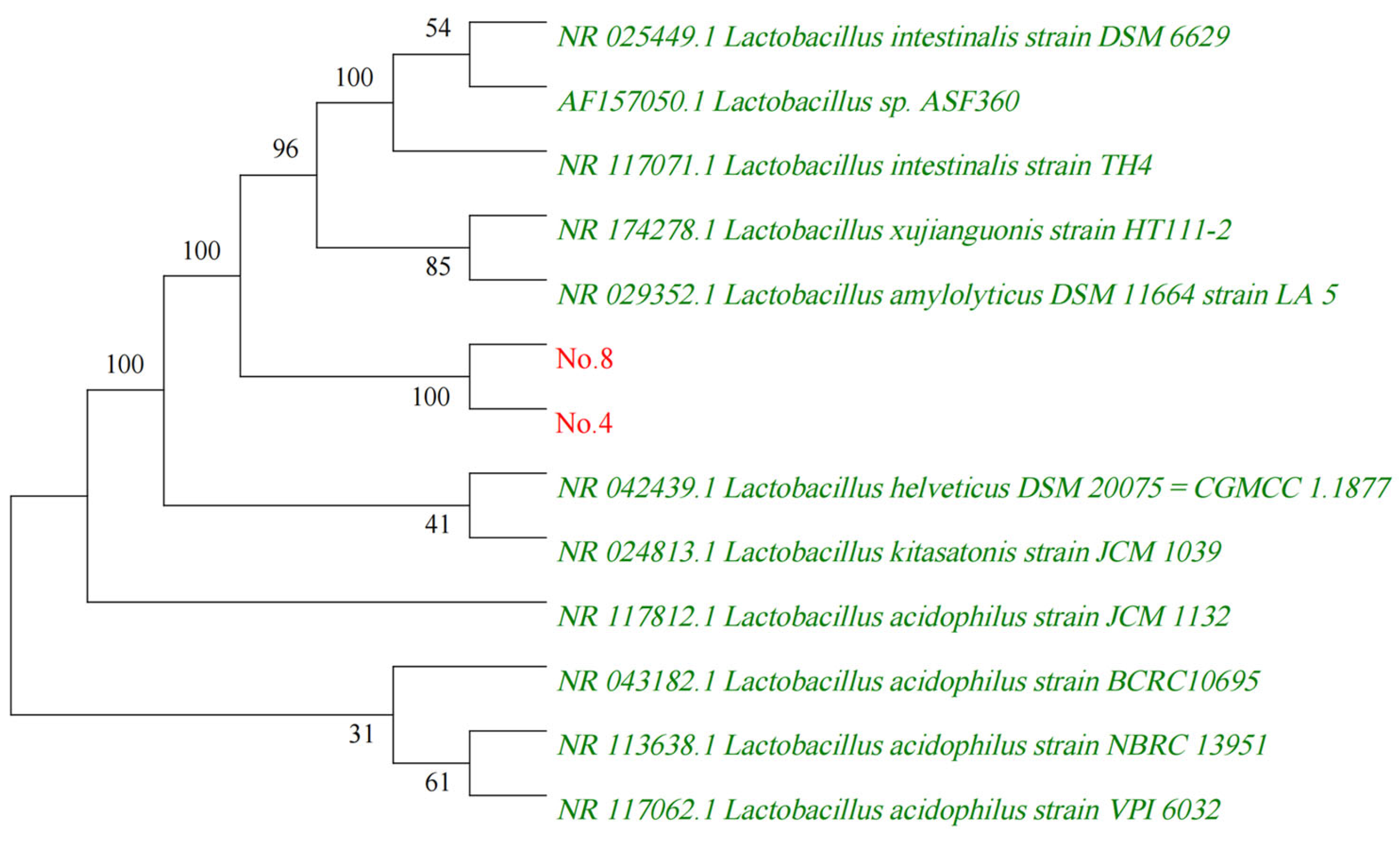
2.2. Identification of L. acidophilus
2.3. L. acidophilus Reduced the Relative Expression of IL-1β mRNA In Vivo and Had Anti-PCV2 Effects
3. Discussion
4. Materials and Methods
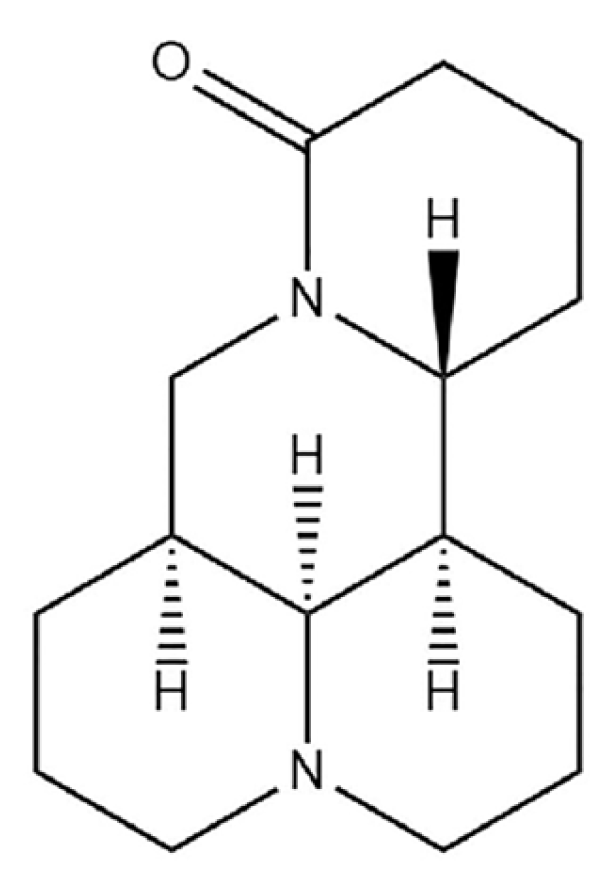
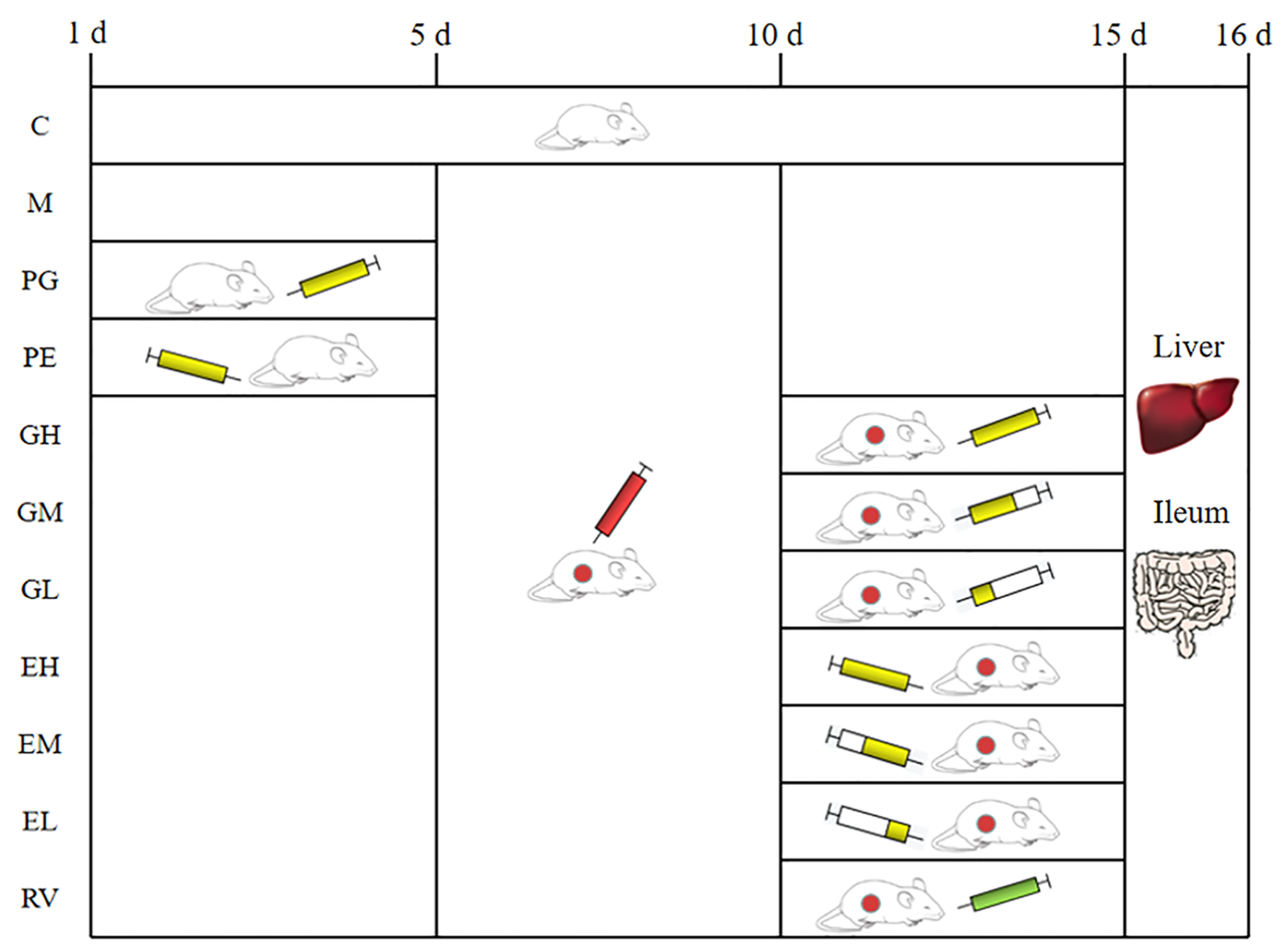
4.1. Compounds, Virus and KM Mice
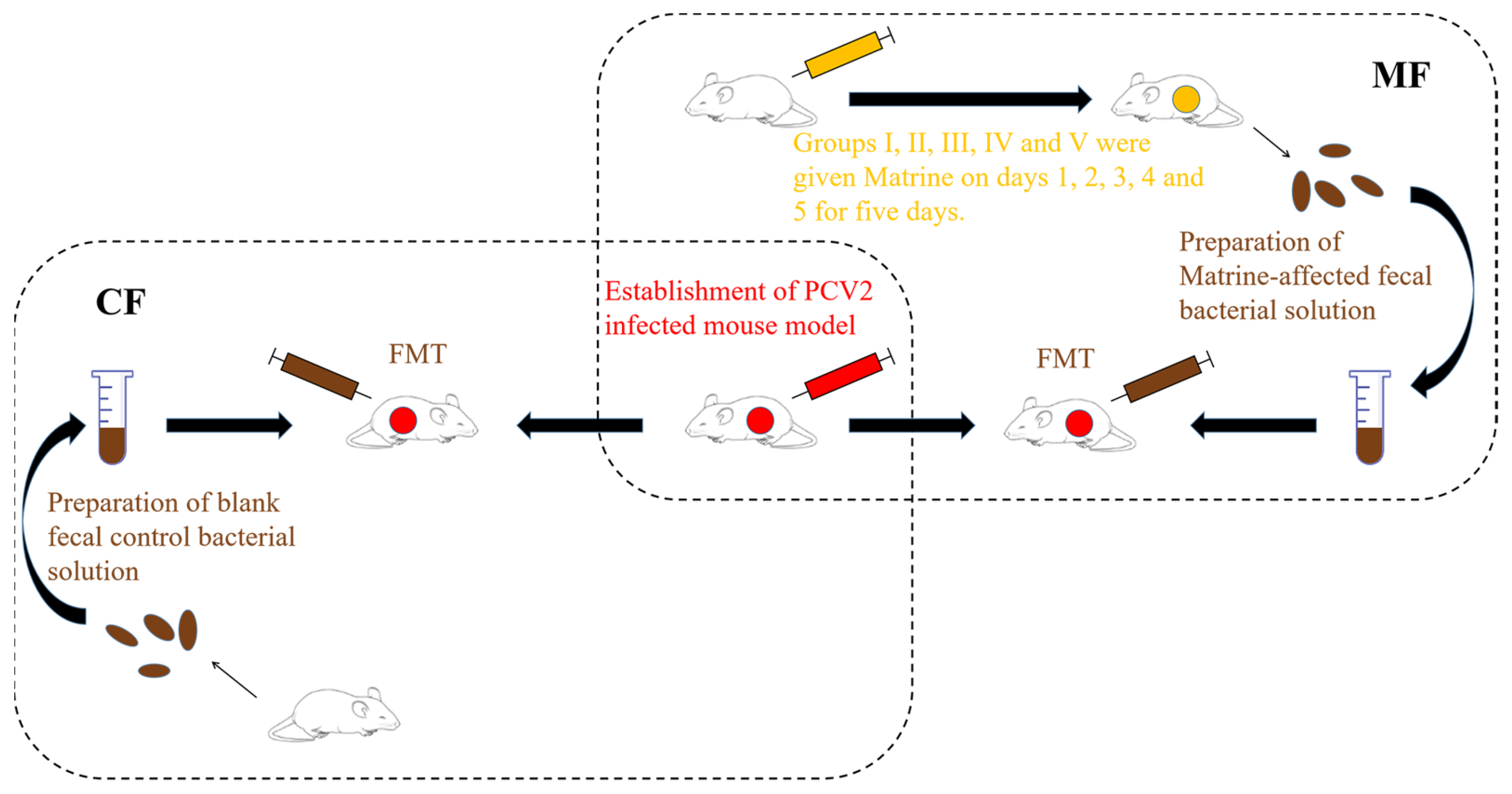
4.2. Preparation of Fecal bacteria Solution and FMT
4.3. Effect of Matrine-Treated Fecal Bacteria against PCV2
4.4. The Expressions of CaI, IL-1β and L. acidophilus Genes Were Determined by qPCR
4.5. The Content of SIgA in Small Intestinal Fluid Was Detected by ELISA
4.6. Isolation and Culture of L. acidophilus in Feces
4.7. Identification of L. acidophilus
4.8. Anti-PCV2 Effect of L. acidophilus In Vivo
4.9. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Cap | Capsid |
| FMT | Fecal microbiota transplantation |
| KM | Kunming |
| L. acidophilus | Lactobacillus acidophilus |
| ORFs | Open reading frame |
| PCVAD | Porcine circovirus associated disease |
| PCV2 | Porcine circovirus type 2 |
| qPCR | Real-time Quantitative polymerase chain reaction |
| SIgA | Secretory immunoglobulin A. |
References
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Investig. 2007, 19, 591–615. [Google Scholar] [CrossRef]
- Hamel, A.; Lin, L.; Nayar, G. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. J. Virol. 1998, 72, 5262–5267. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Li, G.; Zheng, W.; Shu, J.; Chen, J.; Yang, F.; Wu, Y.; He, Y. Development of a Combined Genetic Engineering Vaccine for Porcine Circovirus Type 2 and Mycoplasma Hyopneumoniae by a Baculovirus Expression System. Int. J. Mol. Sci. 2019, 20, 4425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Jiang, W.; Jiang, P.; Li, Y.; Feng, Z.; Xu, J. Construction and immunogenicity of recombinant adenovirus expressing the capsid protein of porcine circovirus 2 (PCV2) in mice. Vaccine 2006, 24, 3374–3380. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Zhang, W.; Ma, S.; Qiu, Z.; Li, B.; Xu, C.; He, H.; Fan, S.; Wu, K.; Chen, J.; et al. Fusion expression and immune effect of PCV2 cap protein tandem multiantigen epitopes with CD154/GM-CSF. Vet. Sci. 2021, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Segalés, J.; Allan, G.M.; Domingo, M. Porcine circovirus diseases. Anim. Health Res. Rev. 2005, 6, 119–142. [Google Scholar] [CrossRef]
- Sinha, A.; Schalk, S.; Lager, K.M.; Wang, C.; Opriessnig, T. Singular PCV2a or PCV2b infection results in apoptosis of hepatocytes in clinically affected gnotobiotic pigs. Res. Vet. Sci. 2012, 92, 151–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allan, G.M.; McNeilly, F.; Kennedy, S.; Daft, B.; Clarke, E.G.; Ellis, J.A.; Haines, D.M.; Meehan, B.M.; Adair, B.M. Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. J. Vet. Diagn. Investig. 1998, 10, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Niederwerder, M.C.; Jaing, C.J.; Thissen, J.B.; Cino-Ozuna, A.G.; McLoughlin, K.S.; Rowland, R.R. Microbiome associations in pigs with the best and worst clinical outcomes following co-infection with porcine reproductive and respiratory syndrome virus (PRRSV) and porcine circovirus type 2 (PCV2). Vet. Microbiol. 2016, 188, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Ellis, J.; Hassard, L.; Clark, E.; Harding, J.; Allan, G.; Willson, P.; Strokappe, J.; Martin, K.; McNeilly, F.; Meehan, B.; et al. Isolation of circovirus from lesions of pigs with postweaning multisystemic wasting syndrome. Can. Vet. J. 1998, 39, 44–51. [Google Scholar]
- Zhang, J.Q.; Li, Y.M.; Liu, T.; He, W.T.; Chen, Y.T.; Chen, X.H.; Li, X.; Zhou, W.C.; Yi, J.F.; Ren, Z.J. Antitumor effect of matrine in human hepatoma G2 cells by inducing apoptosis and autophagy. World J. Gastroenterol. 2010, 16, 4281–4290. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, Z.Y.; Li, Y.Y.; Luo, Y.; Liu, M.L.; Dong, H.Y.; Wang, Y.X.; Liu, Y.; Zhao, P.T.; Jin, F.G.; et al. Antiinflammatory effects of matrine in LPS-induced acute lung injury in mice. Eur. J. Pharm. Sci. 2011, 44, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Wang, J.; Ma, Z.; Ma, S. Anti-asthmatic effects of matrine in a mouse model of allergic asthma. Fitoterapia 2014, 94, 183–189. [Google Scholar] [CrossRef]
- Sun, N.; Sun, P.; Lv, H.; Sun, Y.; Guo, J.; Wang, Z.; Luo, T.; Wang, S.; Li, H. Matrine displayed antiviral activity in porcine alveolar macrophages co-infected by porcine reproductive and respiratory syndrome virus and porcine circovirus type 2. Sci. Rep. 2016, 6, 24401. [Google Scholar] [CrossRef] [Green Version]
- Barbara, A.M.; Karen, E.N.; Mihai, P. Human microbiome project consortium. A framework for human microbiome research. Nature 2012, 486, 215–221. [Google Scholar]
- Yang, Y.; Misra, B.B.; Liang, L.; Bi, D.; Weng, W.; Wu, W.; Cai, S.; Qin, H.; Goel, A.; Li, X.; et al. Integrated microbiome and metabolome analysis reveals a novel interplay between commensal bacteria and metabolites in colorectal cancer. Theranostics 2019, 9, 4101–4114. [Google Scholar] [CrossRef]
- Sun, B.M.; Meng, L.; Liu, H.; Bao, D. Changes in intestinal flora in preeclampsia rats and effects of probiotics on their inflammation and blood pressure. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10155–10161. [Google Scholar]
- Wang, Y.; Wang, Z.; Wang, Y.; Li, F.; Jia, J.; Song, X.; Qin, S.; Wang, R.; Jin, F.; Kitazato, K.; et al. The gut-microglia connection: Implications for central nervous system diseases. Front. Immunol. 2018, 9, 2325. [Google Scholar] [CrossRef] [Green Version]
- Groves, H.T.; Cuthbertson, L.; James, P.; Moffatt, M.F.; Cox, M.J.; Tregoning, J.S. Respiratory disease following viral lung infection alters the murine gut microbiota. Front. Immunol. 2018, 9, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado Acosta, M.; Geoghegan, E.M.; Lepenies, B.; Ruzal, S.; Kielian, M.; Martinez, M.G. Surface (S) layer proteins of Lactobacillus acidophilus block virus infection via DC-SIGN interaction. Front. Microbiol. 2019, 10, 810. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Kang, X.; Yang, H.; Liu, H.; Yang, X.; Liu, Q.; Tian, H.; Xue, Y.; Ren, P.; Kuang, X.; et al. Lactobacillus acidophilus ameliorates obesity in mice through modulation of gut microbiota dysbiosis and intestinal permeability. Pharmacol. Res. 2022, 175, 106020. [Google Scholar] [CrossRef] [PubMed]
- Juan, Z.; Chen, J.; Ding, B.; Yongping, L.; Liu, K.; Wang, L.; Le, Y.; Liao, Q.; Shi, J.; Huang, J.; et al. Probiotic supplement attenuates chemotherapy-related cognitive impairment in patients with breast cancer: A randomised, double-blind, and placebo-controlled trial. Eur. J. Cancer 2022, 161, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.J.; Chen, C.C.; Huang, Y.C.; Tseng, C.C.; Hsu, J.T.; Lin, Y.F.; Fang, Y.J.; Wu, M.S.; Liou, J.M. The efficacy of Lactobacillus acidophilus and rhamnosus in the reduction of bacterial load of Helicobacter pylori and modification of gut microbiota-a double-blind, placebo-controlled, randomized trial. Helicobacter 2021, 26, e12857. [Google Scholar] [CrossRef]
- Wang, Y.; Fei, Y.; Liu, L.; Xiao, Y.; Pang, Y.; Kang, J.; Wang, Z. Polygonatum odoratum polysaccharides modulate gut microbiota and mitigate experimentally induced obesity in rats. Int. J. Mol. Sci. 2018, 19, 3587. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.L.; Zhao, L.H.; Lian, F.M.; Zhou, Q.; Xia, L.; Zhang, J.C.; Chen, X.Y.; Ji, H.Y. Clinical observations on the dose-effect relationship of gegen qin lian decoction on 54 out-patients with type 2 diabetes. J. Tradit. Chin. Med. 2011, 31, 56–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Li, P.; Zheng, Q.; Hou, J. Lactobacillus acidophilus S-layer protein-mediated inhibition of PEDV-induced apoptosis of Vero cells. Vet. Microbiol. 2019, 229, 159–167. [Google Scholar] [CrossRef]
- Zhang, W.; Azevedo, M.S.; Wen, K.; Gonzalez, A.; Saif, L.J.; Li, G.; Yousef, A.E.; Yuan, L. Probiotic Lactobacillus acidophilus enhances the immunogenicity of an oral rotavirus vaccine in gnotobiotic pigs. Vaccine 2008, 26, 3655–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences |
|---|---|
| β-actin | F: AGGGAAATCGTGCGTGACAT R: GGAAAAGAGCCTCAGGGCAT |
| IL-1β | F: GCCACCTTTTGACAGTGATGAGA R: GACAGCCCAGGTCAAAGGTT |
| Cap | F: GTCTACATTTCCAGCAGTTTG R: CTCCCGCCATACCATAA |
| L acidophilus | F: GAAAGAGCCCAAACCAAGTGATT R: CTTCCCAGATAATTCAACTATCGCTTA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Z.; Ling, X.; Sun, P.; Zheng, X.; Zhang, H.; Zhong, J.; Yin, W.; Fan, K.; Sun, Y.; Li, H.; et al. Matrine Targets Intestinal Lactobacillus acidophilus to Inhibit Porcine Circovirus Type 2 Infection in Mice. Int. J. Mol. Sci. 2023, 24, 11878. https://doi.org/10.3390/ijms241511878
Cao Z, Ling X, Sun P, Zheng X, Zhang H, Zhong J, Yin W, Fan K, Sun Y, Li H, et al. Matrine Targets Intestinal Lactobacillus acidophilus to Inhibit Porcine Circovirus Type 2 Infection in Mice. International Journal of Molecular Sciences. 2023; 24(15):11878. https://doi.org/10.3390/ijms241511878
Chicago/Turabian StyleCao, Zhigang, Xiaoya Ling, Panpan Sun, Xiaozhong Zheng, Hua Zhang, Jia Zhong, Wei Yin, Kuohai Fan, Yaogui Sun, Hongquan Li, and et al. 2023. "Matrine Targets Intestinal Lactobacillus acidophilus to Inhibit Porcine Circovirus Type 2 Infection in Mice" International Journal of Molecular Sciences 24, no. 15: 11878. https://doi.org/10.3390/ijms241511878