
Crosstalk of Inflammation and Coagulation in Bothrops Snakebite Envenoming: Endogenous Signaling Pathways and Pathophysiology
 , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Overview of the Toxic Effects of Bothrops Venoms
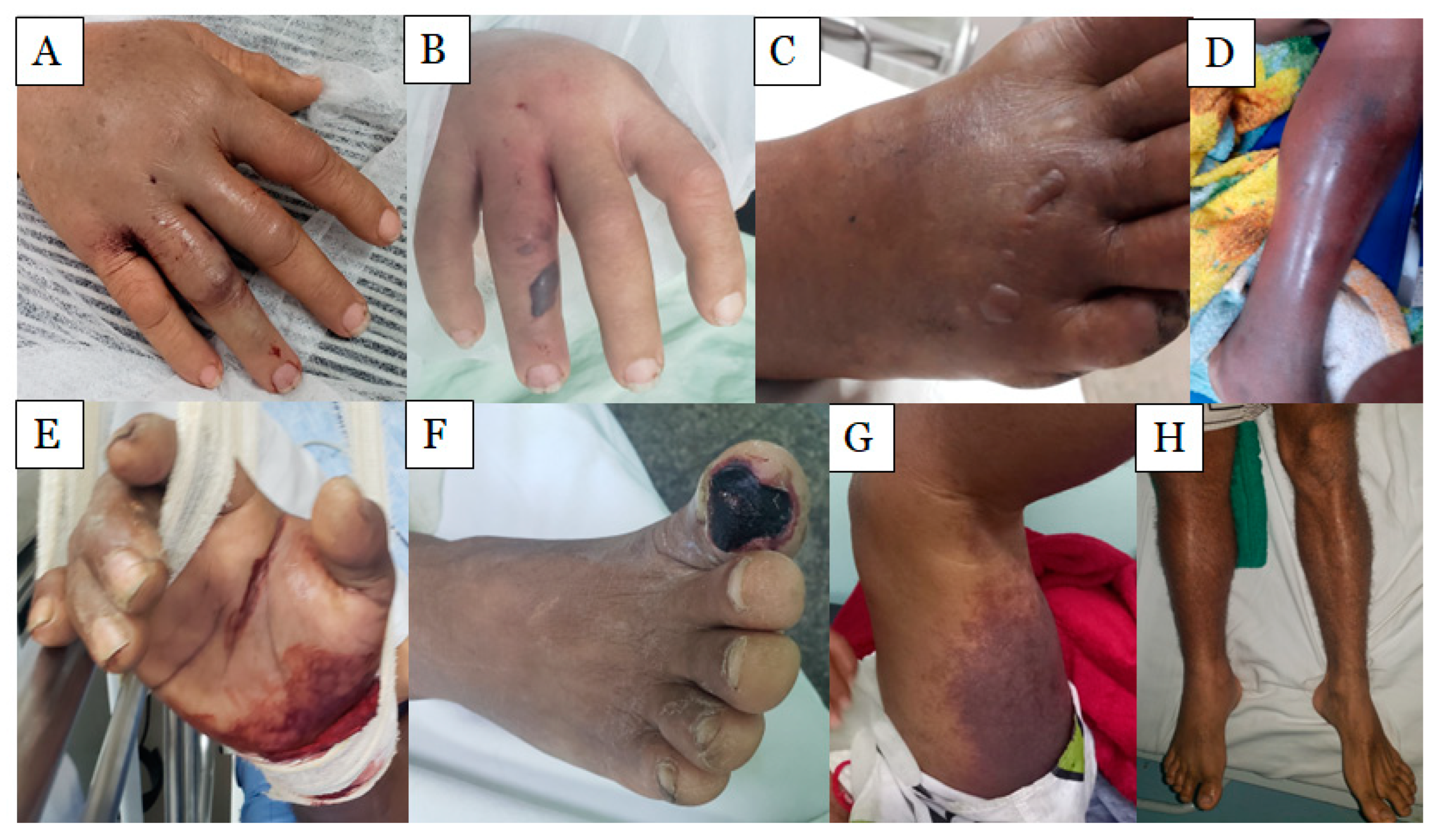
2.1. Local Effects
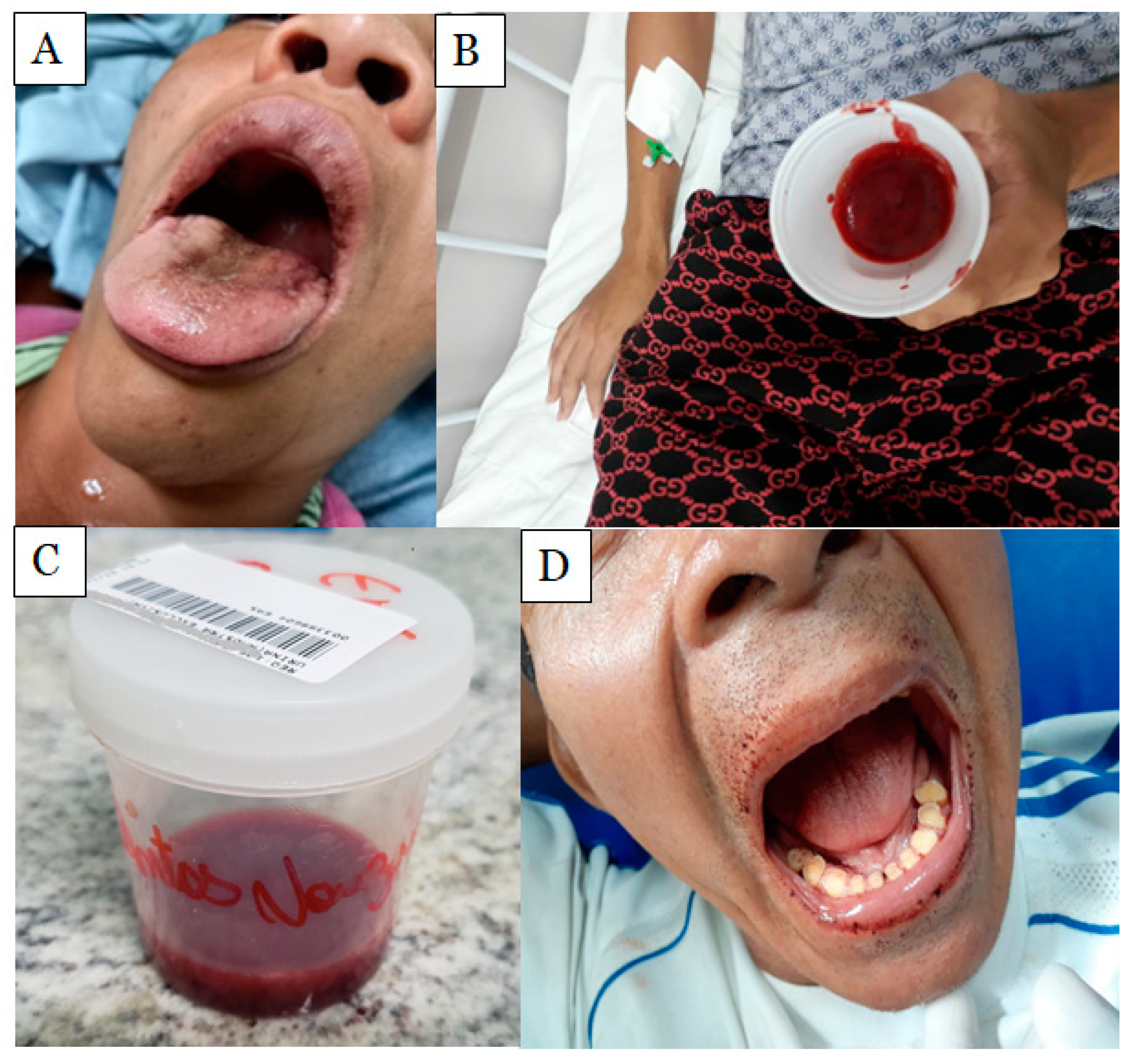
2.2. Systemic Effects
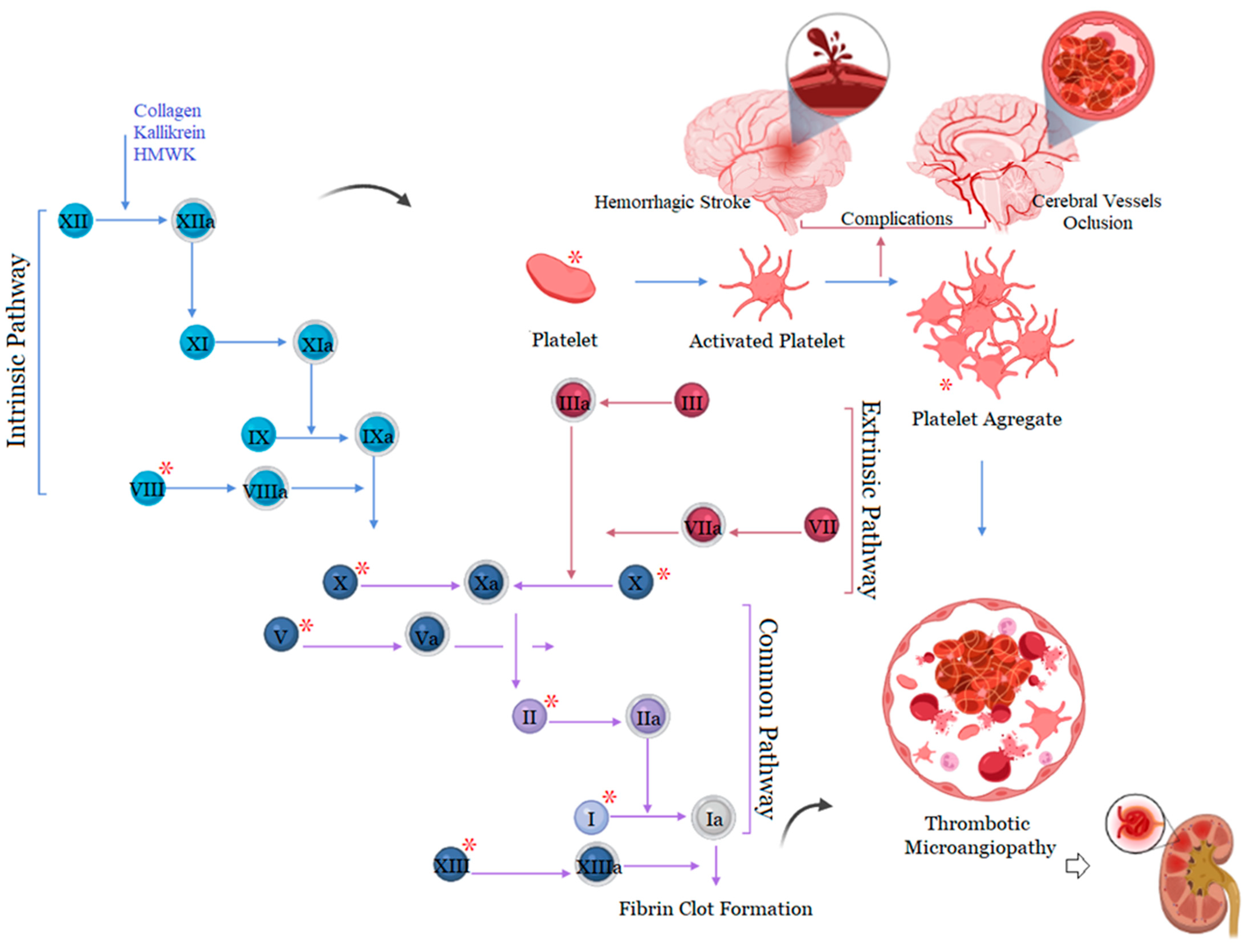
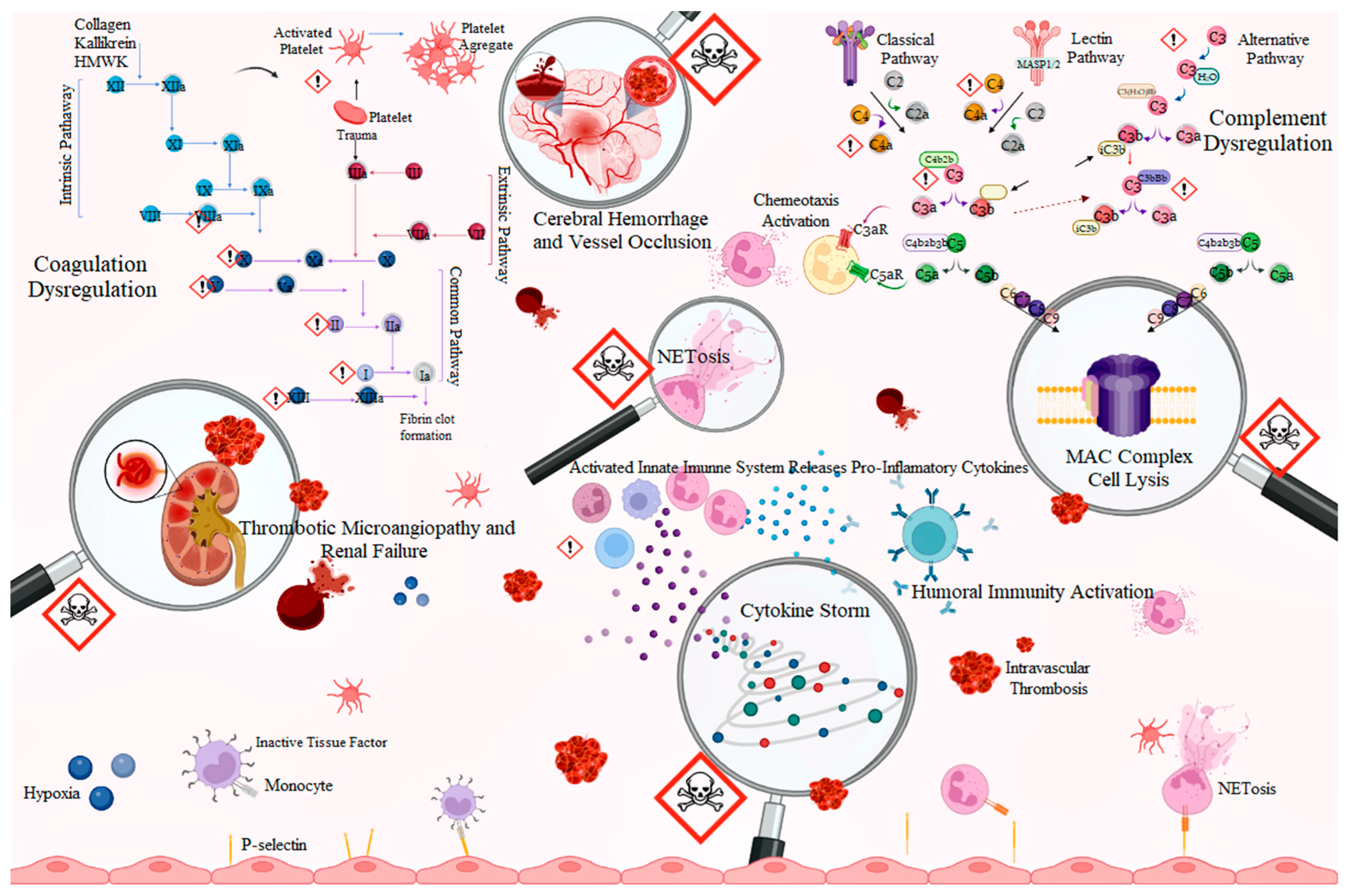
3. Bothrops Snakebite as a Prothrombotic State
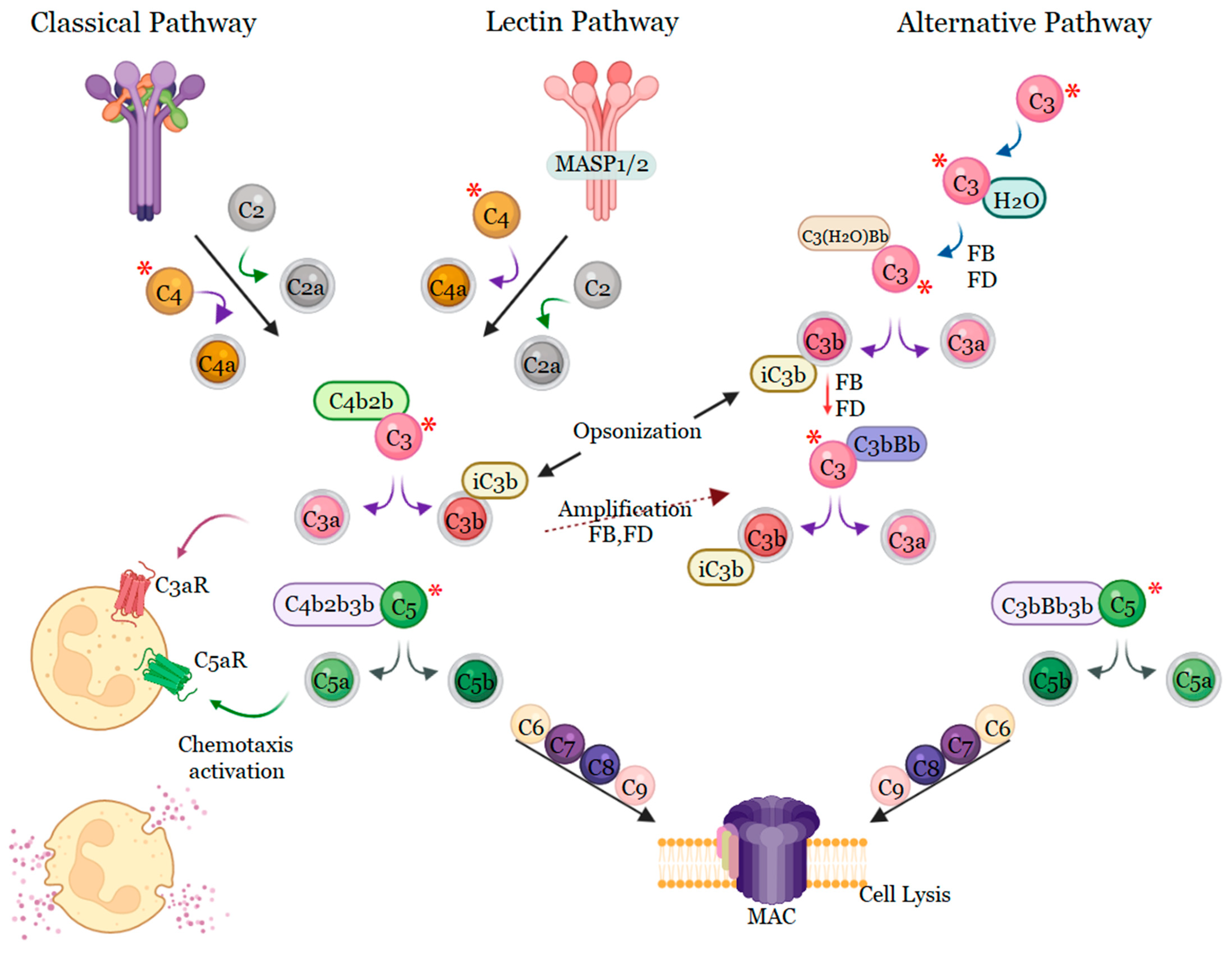
4. Complement System Mediators’ Activation by Bothrops Venoms
5. Role of Neutrophils
6. Cytokines and Their Role in Inflammation: An Overview
7. Platelets and Bothrops Venoms
8. Snake Venom-Induced Coagulation/Inflammation Cross-Talk and Participation of Tissue Factor
9. Thromboinflammation and Snakebite
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Chippaux, J.-P. Snakebite Envenomation Turns Again into a Neglected Tropical Disease! J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 38. [Google Scholar] [CrossRef] [PubMed]
- Minghui, R.; Malecela, M.N.; Cooke, E.; Abela-Ridder, B. WHO’s Snakebite Envenoming Strategy for Prevention and Control. Lancet Glob. Health 2019, 7, e837–e838. [Google Scholar] [CrossRef]
- Gutiérrez, J.M.; Calvete, J.J.; Habib, A.G.; Harrison, R.A.; Williams, D.J.; Warrell, D.A. Snakebite Envenoming. Nat. Rev. Dis. Prim. 2017, 3, 17063. [Google Scholar] [CrossRef] [PubMed]
- Chippaux, J.-P. Incidence and Mortality Due to Snakebite in the Americas. PLoS Negl. Trop. Dis. 2017, 11, e0005662. [Google Scholar] [CrossRef] [PubMed]
- Alencar, L.R.V.; Quental, T.B.; Grazziotin, F.G.; Alfaro, M.L.; Martins, M.; Venzon, M.; Zaher, H. Diversification in Vipers: Phylogenetic Relationships, Time of Divergence and Shifts in Speciation Rates. Mol. Phylogenet. Evol. 2016, 105, 50–62. [Google Scholar] [CrossRef]
- Carrasco, P.A.; Venegas, P.J.; Chaparro, J.C.; Scrocchi, G.J. Nomenclatural Instability in the Venomous Snakes of the Bothrops Complex: Implications in Toxinology and Public Health. Toxicon 2016, 119, 122–128. [Google Scholar] [CrossRef]
- Ferraz, C.R.; Arrahman, A.; Xie, C.; Casewell, N.R.; Lewis, R.J.; Kool, J.; Cardoso, F.C. Multifunctional Toxins in Snake Venoms and Therapeutic Implications: From Pain to Hemorrhage and Necrosis. Front. Ecol. Evol. 2019, 7, 218. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, P.L.M.M.; Paiva, J.H.H.G.L.; Martins, A.M.C.; Meneses, G.C.; Silva Júnior, G.B.; Buckley, N.; Daher, E.D.F. Clinical Assessment and Pathophysiology of Bothrops Venom-Related Acute Kidney Injury: A Scoping Review. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190076. [Google Scholar] [CrossRef]
- Núñez, V.; Cid, P.; Sanz, L.; de La Torre, P.; Angulo, Y.; Lomonte, B.; Gutiérrez, J.M.; Calvete, J.J. Snake Venomics and Antivenomics of Bothrops Atrox Venoms from Colombia and the Amazon Regions of Brazil, Perú and Ecuador Suggest the Occurrence of Geographic Variation of Venom Phenotype by a Trend towards Paedomorphism. J Proteom. 2009, 73, 57–78. [Google Scholar] [CrossRef]
- Sousa, L.F.; Nicolau, C.A.; Peixoto, P.S.; Bernardoni, J.L.; Oliveira, S.S.; Portes-Junior, J.A.; Mourão, R.H.V.; Lima-dos-Santos, I.; Sano-Martins, I.S.; Chalkidis, H.M.; et al. Comparison of Phylogeny, Venom Composition and Neutralization by Antivenom in Diverse Species of Bothrops Complex. PLoS Negl. Trop Dis. 2013, 7, e2442. [Google Scholar] [CrossRef] [Green Version]
- Simizo, A.; Kitano, E.S.; Sant’Anna, S.S.; Grego, K.F.; Tanaka-Azevedo, A.M.; Tashima, A.K. Comparative Gender Peptidomics of Bothrops Atrox Venoms: Are There Differences between Them? J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200055. [Google Scholar] [CrossRef] [PubMed]
- Hatakeyama, D.M.; Tasima, L.J.; Bravo-Tobar, C.A.; Serino-Silva, C.; Tashima, A.K.; Rodrigues, C.F.; Aguiar, W.D.; Galizio, N.D.; Lima, E.O.; Kavazoi, V.K.; et al. Venom Complexity of Bothrops Atrox (Common Lancehead) Siblings. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200018. [Google Scholar] [CrossRef] [PubMed]
- Sanz, L.; Pérez, A.; Quesada-Bernat, S.; Diniz-Sousa, R.; Calderón, L.A.; Soares, A.M.; Calvete, J.J.; Caldeira, C.A.S. Venomics and Antivenomics of the Poorly Studied Brazil’s Lancehead, Bothrops Brazili (Hoge, 1954), from the Brazilian State of Pará. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190103. [Google Scholar] [CrossRef]
- Pereira, L.D.; Messias, E.A.; Sorroche, B.P.; Oliveira, A.D.; Arantes, L.M.; Carvalho, A.C.; Tanaka-Azevedo, A.M.; Grego, K.F.; Carvalho, A.L.; Melendez, M.E. In-Depth Transcriptome Reveals the Potential Biotechnological Application of Bothrops Jararaca Venom Gland. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190058. [Google Scholar] [CrossRef]
- Tasoulis, T.; Isbister, G. A Review and Database of Snake Venom Proteomes. Toxins 2017, 9, 290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- dos Santos Cavalcante, J.; Júnior, F.A.; Jorge, R.J.; Almeida, C. Pain Modulated by Bothrops Snake Venoms: Mechanisms of Nociceptive Signaling and Therapeutic Perspectives. Toxicon 2021, 201, 105–114. [Google Scholar] [CrossRef]
- da Silva Souza, A.; Sachett, J.D.; Alcântara, J.A.; Freire, M.; Alecrim, M.D.; de Lima Ferreira, L.C.; Fan, H.W.; de Souza Sampaio, V.; Monteiro, W.M. Snakebites as Cause of Deaths in the Western Brazilian Amazon: Why and Who Dies? Deaths from Snakebites in the Amazon. Toxicon 2018, 145, 15–24. [Google Scholar] [CrossRef]
- Nascimento da Costa, T.; Mota-da-Silva, A.; Colombini, M.; Moura-da-Silva, A.M.; Medeiros de Souza, R.; Monteiro, W.M.; Bernarde, P.S. Relationship between Snake Size and Clinical, Epidemiological and Laboratory Aspects of Bothrops Atrox Snakebites in the Western Brazilian Amazon. Toxicon 2020, 186, 160–167. [Google Scholar] [CrossRef]
- Luciano, P.M.; Silva, G.E.B.; Azevedo-Marques, M.M. Acidente Botrópico Fatal. Med. (Ribeirão Preto) 2009, 42, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Schulz, R.S.; Sousa Queiroz, P.E.; Bastos, M.C.; Miranda, E.A.; Jesus, H.S.; Gatis, S.M.P. Treatment Of Wound Due To Ophidic Accident: Clinical Case. CuidArte Enferm. 2016, 10, 172–179. Available online: http://www.webfipa.net/facfipa/ner/sumarios/cuidarte/2016v2/172-179.pdf (accessed on 4 June 2023).
- Abero Valle, L.; da Fonseca Reis Silva, D.; Henrique Magalhães, P.; Aloísio Mattos, P.; Almeida Leal, J. Bilateral Amputation of Inferior Extremities Due to Serious Bothrops Accident: A Case Report. Arq. Med. Hosp. Fac. Cienc. Med. Santa Casa São Paulo. 2008, 53, 81–84. Available online: https://arquivosmedicos.fcmsantacasasp.edu.br/index.php/AMSCSP/article/view/404/456 (accessed on 4 June 2023).
- Graciano, A.R.; Naves De Carvalho, K.C. Compartimental Syndrome Associated To Snake Bite Of The Bothrops Gender: Case Report. Rev. Pesq. Saúde 2017, 18, 54–56. Available online: https://periodicoseletronicos.ufma.br/index.php/revistahuufma/article/view/7881/4876 (accessed on 4 June 2023).
- Cirille, S.; Chatot-Henry, D.; Didier, M.; Moravie, V.; Thomas, L.; Numeric, P.; Bucher, B. Multiple Cerebral Infarctions Following a Snakebite by Bothrops Caribbaeus. Am. J. Trop. Med. Hyg. 2002, 67, 287–288. [Google Scholar] [CrossRef] [Green Version]
- Malaque, C.M.S.; Duayer, I.F.; Santoro, M.L. Acute Kidney Injury Induced by Thrombotic Microangiopathy in Two Cases of Bothrops Envenomation. Clin. Toxicol. 2019, 57, 213–216. [Google Scholar] [CrossRef]
- Pucca, M.B.; Franco, M.V.S.; Medeiros, J.M.; Oliveira, I.S.; Ahmadi, S.; Cerni, F.A.; Zottich, U.; Bassoli, B.K.; Monteiro, W.M.; Laustsen, A.H. Chronic Kidney Failure Following Lancehead Bite Envenoming: A Clinical Report from the Amazon Region. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20200083. [Google Scholar] [CrossRef]
- Larréché, S.; Chippaux, J.-P.; Chevillard, L.; Mathé, S.; Résière, D.; Siguret, V.; Mégarbane, B. Bleeding and Thrombosis: Insights into Pathophysiology of Bothrops Venom-Related Hemostasis Disorders. Int. J. Mol. Sci. 2021, 22, 9643. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.L.; Sano-Martins, I.S.; Fan, H.W.; Cardoso, J.L.C.; Theakston, R.D.G.; Warrell, D.A. Haematological Evaluation of Patients Bitten by the Jararaca, Bothrops Jararaca, in Brazil. Toxicon 2008, 51, 1440–1448. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, S.S.; Alves, E.C.; Santos, A.S.; Pereira, J.P.T.; Sarraff, L.K.S.; Nascimento, E.F.; De-Brito-Sousa, J.D.; Sampaio, V.S.; Lacerda, M.V.G.; Sachett, J.A.G.; et al. Factors Associated with Systemic Bleeding in Bothrops Envenomation in a Tertiary Hospital in the Brazilian Amazon. Toxins 2019, 11, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, S.S.; Alves, E.C.; Santos, A.S.; Nascimento, E.F.; Pereira, J.P.T.; Silva, I.M.; Sachett, J.A.G.; Sarraff, L.K.S.; Freitas-de-Sousa, L.A.; Colombini, M.; et al. Bleeding Disorders in Bothrops Atrox Envenomations in the Brazilian Amazon: Participation of Hemostatic Factors and the Impact of Tissue Factor. Toxins 2020, 12, 554. [Google Scholar] [CrossRef]
- Teixeira, C.; Fernandes, C.M.; Leiguez, E.; Chudzinski-Tavassi, A.M. Inflammation Induced by Platelet-Activating Viperid Snake Venoms: Perspectives on Thromboinflammation. Front. Immunol. 2019, 10, 2082. [Google Scholar] [CrossRef] [Green Version]
- Zuliani, J.P.; Soares, A.M.; Gutiérrez, J.M. Polymorphonuclear Neutrophil Leukocytes in Snakebite Envenoming. Toxicon 2020, 187, 188–197. [Google Scholar] [CrossRef]
- Guo, L.; Rondina, M.T. The Era of Thromboinflammation: Platelets Are Dynamic Sensors and Effector Cells During Infectious Diseases. Front. Immunol. 2019, 10, 2204. [Google Scholar] [CrossRef] [Green Version]
- Cavalcante, J.S.; Brito, I.M.; De Oliveira, L.A.; De Barros, L.C.; Almeida, C.; Rossini, B.C.; Sousa, D.L.; Alves, R.S.; Jorge, R.J.B.; Santos, L.D. Experimental Bothrops Atrox Envenomation: Blood Plasma Proteome Effects after Local Tissue Damage and Perspectives on Thromboinflammation. Toxins 2022, 14, 613. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, J.S.; Borges da Silva, W.R.G.; de Oliveira, L.A.; Brito, I.M.C.; Muller, K.S.; Vidal, I.S.J.; dos Santos, L.D.; Jorge, R.J.B.; Almeida, C.; de Lima Bicho, C. Blood Plasma Proteome Alteration after Local Tissue Damage Induced by Bothrops Erythromelas Snake Venom in Mice. J. Proteom. 2022, 269, 104742. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Cavalcante, J.; de Almeida, C.A.; Clasen, M.A.; da Silva, E.L.; de Barros, L.C.; Marinho, A.D.; Rossini, B.C.; Marino, C.L.; Carvalho, P.C.; Jorge, R.J.; et al. A Fingerprint of Plasma Proteome Alteration after Local Tissue Damage Induced by Bothrops Leucurus Snake Venom in Mice. J. Proteom. 2022, 253, 104464. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Escalante, T.; Hernández, R.; Gastaldello, S.; Saravia-Otten, P.; Rucavado, A. Why Is Skeletal Muscle Regeneration Impaired after Myonecrosis Induced by Viperid Snake Venoms? Toxins 2018, 10, 182. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.M.; Rucavado, A.; Escalante, T.; Herrera, C.; Fernández, J.; Lomonte, B.; Fox, J.W. Unresolved Issues in the Understanding of the Pathogenesis of Local Tissue Damage Induced by Snake Venoms. Toxicon 2018, 148, 123–131. [Google Scholar] [CrossRef]
- Rucavado, A.; Nicolau, C.; Escalante, T.; Kim, J.; Herrera, C.; Gutiérrez, J.; Fox, J. Viperid Envenomation Wound Exudate Contributes to Increased Vascular Permeability via a DAMPs/TLR-4 Mediated Pathway. Toxins 2016, 8, 349. [Google Scholar] [CrossRef] [Green Version]
- Luchini, L.S.G.; Pidde, G.; Squaiella-Baptistão, C.C.; Tambourgi, D.V. Complement System Inhibition Modulates the Pro-Inflammatory Effects of a Snake Venom Metalloproteinase. Front. Immunol. 2019, 10, 1137. [Google Scholar] [CrossRef]
- Moreira, V.; Teixeira, C.; Borges da Silva, H.; D’Império Lima, M.R.; Dos-Santos, M.C. The Role of TLR2 in the Acute Inflammatory Response Induced by Bothrops Atrox Snake Venom. Toxicon 2016, 118, 121–128. [Google Scholar] [CrossRef]
- Bickler, P.E. Amplification of Snake Venom Toxicity by Endogenous Signaling Pathways. Toxins 2020, 12, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimenes, S.N.C.; Sachett, J.A.G.; Colombini, M.; Freitas-de-Sousa, L.A.; Ibiapina, H.N.S.; Costa, A.G.; Santana, M.F.; Park, J.-J.; Sherman, N.E.; Ferreira, L.C.L.; et al. Observation of Bothrops Atrox Snake Envenoming Blister Formation from Five Patients: Pathophysiological Insights. Toxins 2021, 13, 800. [Google Scholar] [CrossRef] [PubMed]
- Ibiapina, H.N.S.; Costa, A.G.; Sachett, J.A.G.; Silva, I.M.; Tarragô, A.M.; Neves, J.C.F.; Kerr, M.W.A.; Santana, M.F.; Martins-Filho, O.A.; Lacerda, M.V.G.; et al. An Immunological Stairway to Severe Tissue Complication Assembly in Bothrops Atrox Snakebites. Front. Immunol. 2019, 10, 1882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalante, T.; Rucavado, A.; Pinto, A.F.M.; Terra, R.M.S.; Gutiérrez, J.M.; Fox, J.W. Wound Exudate as a Proteomic Window to Reveal Different Mechanisms of Tissue Damage by Snake Venom Toxins. J. Proteome Res. 2009, 8, 5120–5131. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, J.; Escalante, T.; Rucavado, A.; Herrera, C. Hemorrhage Caused by Snake Venom Metalloproteinases: A Journey of Discovery and Understanding. Toxins 2016, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Gutiérrez, J.; Escalante, T.; Rucavado, A.; Herrera, C.; Fox, J. A Comprehensive View of the Structural and Functional Alterations of Extracellular Matrix by Snake Venom Metalloproteinases (SVMPs): Novel Perspectives on the Pathophysiology of Envenoming. Toxins 2016, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Macêdo, J.K.A.; Joseph, J.K.; Menon, J.; Escalante, T.; Rucavado, A.; Gutiérrez, J.M.; Fox, J.W. Proteomic Analysis of Human Blister Fluids Following Envenomation by Three Snake Species in India: Differential Markers for Venom Mechanisms of Action. Toxins 2019, 11, 246. [Google Scholar] [CrossRef] [Green Version]
- Zambelli, V.; Picolo, G.; Fernandes, C.; Fontes, M.; Cury, Y. Secreted Phospholipases A2 from Animal Venoms in Pain and Analgesia. Toxins 2017, 9, 406. [Google Scholar] [CrossRef] [Green Version]
- Baldo, C.; Jamora, C.; Yamanouye, N.; Zorn, T.M.; Moura-da-Silva, A.M. Mechanisms of Vascular Damage by Hemorrhagic Snake Venom Metalloproteinases: Tissue Distribution and In Situ Hydrolysis. PLoS Negl. Trop. Dis. 2010, 4, e727. [Google Scholar] [CrossRef] [Green Version]
- Herrera, C.; Escalante, T.; Voisin, M.-B.; Rucavado, A.; Morazán, D.; Macêdo, J.K.A.; Calvete, J.J.; Sanz, L.; Nourshargh, S.; Gutiérrez, J.M.; et al. Tissue Localization and Extracellular Matrix Degradation by PI, PII and PIII Snake Venom Metalloproteinases: Clues on the Mechanisms of Venom-Induced Hemorrhage. PLoS Negl. Trop. Dis. 2015, 9, e0003731. [Google Scholar] [CrossRef]
- Kini, R.; Koh, C. Metalloproteases Affecting Blood Coagulation, Fibrinolysis and Platelet Aggregation from Snake Venoms: Definition and Nomenclature of Interaction Sites. Toxins 2016, 8, 284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos Barreto, G.N.; de Oliveira, S.S.; Dos Anjos, I.V.; Chalkidis, H.D.; Mourão, R.H.; Moura-da-Silva, A.M.; Sano-Martins, I.S.; Gonçalves, L.R. Experimental Bothrops Atrox Envenomation: Efficacy of Antivenom Therapy and the Combination of Bothrops Antivenom with Dexamethasone. PLoS Negl. Trop. Dis. 2017, 11, e0005458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mora-Obando, D.; Fernández, J.; Montecucco, C.; Gutiérrez, J.M.; Lomonte, B. Synergism between Basic Asp49 and Lys49 Phospholipase A2 Myotoxins of Viperid Snake Venom In Vitro and In Vivo. PLoS ONE 2014, 9, e109846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunitha, K.; Hemshekhar, M.; Thushara, R.M.; Santhosh, M.S.; Sundaram, M.S.; Kemparaju, K.; Girish, K.S. Inflammation and Oxidative Stress in Viper Bite: An Insight within and Beyond. Toxicon 2015, 98, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, L.A.; França, F.O.S.; Barbaro, K.C.; Nunes, J.R.; Cardoso, J.L.C. Pulmonary Haemorrhage Causing Rapid Death after Bothrops Jararacussu Snakebite: A Case Report. Toxicon 2003, 42, 331–334. [Google Scholar] [CrossRef]
- Kamiguti, A.S.; Hay, C.R.M.; Theakston, R.D.G.; Zuzel, M. Insights into the Mechanism of Haemorrhage Caused by Snake Venom Metalloproteinases. Toxicon 1996, 34, 627–642. [Google Scholar] [CrossRef]
- Kamiguti, A.S.; Cardoso, J.L.C.; Theakston, R.D.G.; Sano-Martins, I.S.; Hutton, R.A.; Rugman, F.P.; Warrell, D.A.; Hay, C.R.M. Coagulopathy and Haemorrhage in Human Victims of Bothrops Jararaca Envenoming in Brazil. Toxicon 1991, 29, 961–972. [Google Scholar] [CrossRef]
- Silva de Oliveira, S.; Freitas-de-Sousa, L.A.; Alves, E.C.; de Lima Ferreira, L.C.; da Silva, I.M.; de Lacerda, M.V.G.; Fan, H.W.; Moura-da-Silva, A.M.; Monteiro, W.M. Fatal Stroke after Bothrops Snakebite in the Amazonas State, Brazil: A Case Report. Toxicon 2017, 138, 102–106. [Google Scholar] [CrossRef]
- de Queiroz, M.R.; de Sousa, B.B.; da Cunha Pereira, D.F.; Mamede, C.C.N.; Matias, M.S.; de Morais, N.C.G.; de Oliveira Costa, J.; de Oliveira, F. The Role of Platelets in Hemostasis and the Effects of Snake Venom Toxins on Platelet Function. Toxicon 2017, 133, 33–47. [Google Scholar] [CrossRef]
- Sitprija, V. Animal Toxins and the Kidney. Nat. Clin. Pr. Nephrol. 2008, 4, 616–627. [Google Scholar] [CrossRef]
- Havt, A.; Toyama, M.H.; do Nascimento, N.R.F.; Toyama, D.O.; Nobre, A.C.L.; Martins, A.M.C.; Barbosa, P.S.F.; Novello, J.C.; Boschero, A.C.; Carneiro, E.M.; et al. A New C-Type Animal Lectin Isolated from Bothrops Pirajai Is Responsible for the Snake Venom Major Effects in the Isolated Kidney. Int. J. Biochem. Cell Biol. 2005, 37, 130–141. [Google Scholar] [CrossRef]
- Marinho, A.D.; de Moraes Silveira, J.A.; Chaves Filho, A.J.; Jorge, A.R.; Júnior, F.A.; Pereira, V.B.; de Aquino, P.E.; Pereira, C.A.; Evangelista, J.S.; Macedo, D.S.; et al. Bothrops Pauloensis Snake Venom-Derived Asp-49 and Lys-49 Phospholipases A2 Mediates Acute Kidney Injury by Oxidative Stress and Release of Inflammatory Cytokines. Toxicon 2021, 190, 31–38. [Google Scholar] [CrossRef]
- Barbosa, P.S.F.; Havt, A.; Facó, P.E.G.; Sousa, T.M.; Bezerra, I.S.A.M.; Fonteles, M.C.; Toyama, M.H.; Marangoni, S.; Novello, J.C.; Monteiro, H.S.A. Renal Toxicity of Bothrops Moojeni Snake Venom and Its Main Myotoxins. Toxicon 2002, 40, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Slagboom, J.; Mladić, M.; Xie, C.; Kazandjian, T.D.; Vonk, F.; Somsen, G.W.; Casewell, N.R.; Kool, J. High Throughput Screening and Identification of Coagulopathic Snake Venom Proteins and Peptides Using Nanofractionation and Proteomics Approaches. PLoS Negl. Trop. Dis. 2020, 14, e0007802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stocker, K.; Barlow, G.H. The Coagulant Enzyme from Bothrops Atrox Venom (Batroxobin). In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1976; pp. 214–223. [Google Scholar]
- Antunes, T.C.; Yamashita, K.M.; Barbaro, K.C.; Saiki, M.; Santoro, M.L. Comparative Analysis of Newborn and Adult Bothrops Jararaca Snake Venoms. Toxicon 2010, 56, 1443–1458. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, W.M.; Contreras-Bernal, J.C.; Bisneto, P.F.; Sachett, J.; Mendonça da Silva, I.; Lacerda, M.; Guimarães da Costa, A.; Val, F.; Brasileiro, L.; Sartim, M.A.; et al. Bothrops Atrox, the Most Important Snake Involved in Human Envenomings in the Amazon: How Venomics Contributes to the Knowledge of Snake Biology and Clinical Toxinology. Toxicon X 2020, 6, 100037. [Google Scholar] [CrossRef]
- de Albuquerque Modesto, J.C.; Spencer, P.J.; Fritzen, M.; Valença, R.C.; Oliva, M.L.V.; da Silva, M.B.; Chudzinski-Tavassi, A.M.; Guarnieri, M.C. BE-I-PLA2, a Novel Acidic Phospholipase A2 from Bothrops Erythromelas Venom: Isolation, Cloning and Characterization as Potent Anti-Platelet and Inductor of Prostaglandin I2 Release by Endothelial Cells. Biochem. Pharm. 2006, 72, 377–384. [Google Scholar] [CrossRef]
- Hofmann, H.; Bon, C. Blood Coagulation Induced by the Venom of Bothrops Atrox. 2. Identification, Purification, and Properties of Two Factor X Activators. Biochemistry 1987, 26, 780–787. [Google Scholar] [CrossRef]
- Rosing, J.; Govers-Riemslag, J.W.P.; Yukelson, L.; Tans, G. Factor V Activation and Inactivation by Venom Proteases. Pathophysiol. Haemost. Thromb. 2001, 31, 241–246. [Google Scholar] [CrossRef]
- Kirby, E.P.; Niewiarowski, S.; Stocker, K.; Kettner, C.; Shaw, E.; Brudzynski, T.M. Thrombocytin, a Serine Protease from Bothrops Atrox Venom. 1. Purification and Characterization of the Enzyme. Biochemistry 1979, 18, 3564–3570. [Google Scholar] [CrossRef]
- Cintra, A.C.O.; de Toni, L.G.B.; Sartim, M.A.; Franco, J.J.; Caetano, R.C.; Murakami, M.T.; Sampaio, S.V. Batroxase, a New Metalloproteinase from B. Atrox Snake Venom with Strong Fibrinolytic Activity. Toxicon 2012, 60, 70–82. [Google Scholar] [CrossRef]
- Baldo, C.; Tanjoni, I.; León, I.R.; Batista, I.F.C.; Della-Casa, M.S.; Clissa, P.B.; Weinlich, R.; Lopes-Ferreira, M.; Lebrun, I.; Amarante-Mendes, G.P.; et al. BnP1, a Novel P-I Metalloproteinase from Bothrops Neuwiedi Venom: Biological Effects Benchmarking Relatively to Jararhagin, a P-III SVMP. Toxicon 2008, 51, 54–65. [Google Scholar] [CrossRef]
- Freitas-de-Sousa, L.A.; Amazonas, D.R.; Sousa, L.F.; Sant’Anna, S.S.; Nishiyama, M.Y.; Serrano, S.M.T.; Junqueira-de-Azevedo, I.L.M.; Chalkidis, H.M.; Moura-da-Silva, A.M.; Mourão, R.H.V. Comparison of Venoms from Wild and Long-Term Captive Bothrops Atrox Snakes and Characterization of Batroxrhagin, the Predominant Class PIII Metalloproteinase from the Venom of This Species. Biochimie 2015, 118, 60–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moretto Del-Rei, T.H.; Sousa, L.F.; Rocha, M.M.T.; Freitas-de-Sousa, L.A.; Travaglia-Cardoso, S.R.; Grego, K.; Sant’Anna, S.S.; Chalkidis, H.M.; Moura-da-Silva, A.M. Functional Variability of Bothrops Atrox Venoms from Three Distinct Areas across the Brazilian Amazon and Consequences for Human Envenomings. Toxicon 2019, 164, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, A.; Magalhães, H.P.B.; Richardson, M.; Gontijo, S.; Ferreira, R.N.; Almeida, A.P.; Sanchez, E.F. Purification and Properties of a Coagulant Thrombin-like Enzyme from the Venom of Bothrops Leucurus. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 565–575. [Google Scholar] [CrossRef]
- Fernandes de Oliveira, L.M.; Ullah, A.; Masood, R.; Zelanis, A.; Spencer, P.J.; Serrano, S.M.T.; Arni, R.K. Rapid Purification of Serine Proteinases from Bothrops Alternatus and Bothrops Moojeni Venoms. Toxicon 2013, 76, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Vivas-Ruiz, D.E.; Sandoval, G.A.; Mendoza, J.; Inga, R.R.; Gontijo, S.; Richardson, M.; Eble, J.A.; Yarleque, A.; Sanchez, E.F. Coagulant Thrombin-like Enzyme (Barnettobin) from Bothrops Barnetti Venom: Molecular Sequence Analysis of Its CDNA and Biochemical Properties. Biochimie 2013, 95, 1476–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vivas-Ruiz, D.E.; Sandoval, G.A.; Gonzalez-Kozlova, E.; Zarria-Romero, J.; Lazo, F.; Rodríguez, E.; Magalhães, H.P.B.; Chávez-Olortegui, C.; Oliveira, L.S.; Alvarenga, V.G.; et al. Fibrinogen-Clotting Enzyme, Pictobin, from Bothrops Pictus Snake Venom. Structural and Functional Characterization. Int. J. Biol. Macromol. 2020, 153, 779–795. [Google Scholar] [CrossRef]
- Amorim, F.; Menaldo, D.; Carone, S.; Silva, T.; Sartim, M.; De Pauw, E.; Quinton, L.; Sampaio, S. New Insights on Moojase, a Thrombin-Like Serine Protease from Bothrops Moojeni Snake Venom. Toxins 2018, 10, 500. [Google Scholar] [CrossRef] [Green Version]
- Cañas, C.A. Brainstem Ischemic Stroke after to Bothrops Atrox Snakebite. Toxicon 2016, 120, 124–127. [Google Scholar] [CrossRef]
- Ayeboua, L.; Plumelle, Y.; Smadja, D.; Merle, H.; Thomas, L.; Donnio, A. Occipital infarction revealed by quadranopsia following snakebite by bothrops lanceolatus. Am. J. Trop. Med. Hyg. 2005, 73, 583–585. [Google Scholar] [CrossRef]
- Thomas, L.; Chausson, N.; Uzan, J.; Kaidomar, S.; Vignes, R.; Plumelle, Y.; Bucher, B.; Smadja, D. Thrombotic Stroke Following Snake Bites by the “Fer-de-Lance” Bothrops Lanceolatus in Martinique despite Antivenom Treatment: A Report of Three Recent Cases. Toxicon 2006, 48, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Bucaretchi, F.; Pimenta, M.M.B.; Borrasca-Fernandes, C.F.; Prado, C.C.; Capitani, E.M.; Hyslop, S. Thrombotic Microangiopathy Following Bothrops Jararaca Snakebite: Case Report. Clin. Toxicol. 2019, 57, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Cañas, C.A.; Vecino, M.J.; Posso-Osorio, I. Atypical Hemolytic Uremic Syndrome in a Patient With Bothrops Asper Envenomation. Wilderness Env. Med. 2022, 33, 109–115. [Google Scholar] [CrossRef]
- Mota, S.M.B.; Albuquerque, P.L.M.M.; Silva Júnior, G.B.d.; Daher, E.D.F. Thrombotic Microangiopathy Due to Bothrops Erythromelas: A Case Report in Northeast Brazil. Rev. Inst. Med. Trop. Sao Paulo 2020, 62, e53. [Google Scholar] [CrossRef]
- Noutsos, T.; Currie, B.J.; Wijewickrama, E.S.; Isbister, G.K. Snakebite Associated Thrombotic Microangiopathy and Recommendations for Clinical Practice. Toxins 2022, 14, 57. [Google Scholar] [CrossRef] [PubMed]
- Senise, L.V.; Yamashita, K.M.; Santoro, M.L. Bothrops Jararaca Envenomation: Pathogenesis of Hemostatic Disturbances and Intravascular Hemolysis. Exp. Biol. Med. 2015, 240, 1528–1536. [Google Scholar] [CrossRef] [Green Version]
- Sartim, M.A.; Cezarette, G.N.; Jacob-Ferreira, A.L.; Frantz, F.G.; Faccioli, L.H.; Sampaio, S.V. Disseminated Intravascular Coagulation Caused by Moojenactivase, a Procoagulant Snake Venom Metalloprotease. Int. J. Biol. Macromol. 2017, 103, 1077–1086. [Google Scholar] [CrossRef]
- Thomazini, C.M.; Sachetto, A.T.; de Albuquerque, C.Z.; de Moura Mattaraia, V.G.; de Oliveira, A.K.; Serrano, S.M.; Lebrun, I.; Barbaro, K.C.; Santoro, M.L. Involvement of von Willebrand Factor and Botrocetin in the Thrombocytopenia Induced by Bothrops Jararaca Snake Venom. PLoS Negl. Trop. Dis. 2021, 15, e0009715. [Google Scholar] [CrossRef]
- Cedzyński, M.; Thielens, N.M.; Mollnes, T.E.; Vorup-Jensen, T. Editorial: The Role of Complement in Health and Disease. Front. Immunol. 2019, 10, 1869. [Google Scholar] [CrossRef]
- Pidde-Queiroz, G.; de Fátima Furtado, M.; Filgueiras, C.F.; Pessoa, L.A.; Spadafora-Ferreira, M.; Van den Berg, C.W.; Tambourgi, D.V. Human Complement Activation and Anaphylatoxins Generation Induced by Snake Venom Toxins from Bothrops Genus. Mol. Immunol. 2010, 47, 2537–2544. [Google Scholar] [CrossRef]
- Delafontaine, M.; Villas-Boas, I.M.; Pidde, G.; van den Berg, C.W.; Mathieu, L.; Blomet, J.; Tambourgi, D.V. Venom from Bothrops Lanceolatus, a Snake Species Native to Martinique, Potently Activates the Complement System. J. Immunol. Res. 2018, 2018, 3462136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berends, E.T.M.; Kuipers, A.; Ravesloot, M.M.; Urbanus, R.T.; Rooijakkers, S.H.M. Bacteria under Stress by Complement and Coagulation. FEMS Microbiol. Rev. 2014, 38, 1146–1171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, B.P. The Membrane Attack Complex as an Inflammatory Trigger. Immunobiology 2016, 221, 747–751. [Google Scholar] [CrossRef]
- Joiner, K.A.; Brown, E.J.; Frank, M.M. Complement and Bacteria: Chemistry and Biology in Host Defense. Annu. Rev. Immunol. 1984, 2, 461–492. [Google Scholar] [CrossRef] [PubMed]
- Palmer, L.J.; Damgaard, C.; Holmstrup, P.; Nielsen, C.H. Influence of Complement on Neutrophil Extracellular Trap Release Induced by Bacteria. J. Periodontal Res. 2016, 51, 70–76. [Google Scholar] [CrossRef]
- Lehman, H.K.; Segal, B.H. The Role of Neutrophils in Host Defense and Disease. J. Allergy Clin. Immunol. 2020, 145, 1535–1544. [Google Scholar] [CrossRef]
- Beavers, W.N.; Skaar, E.P. Neutrophil-Generated Oxidative Stress and Protein Damage in Staphylococcus Aureus. Pathog. Dis. 2016, 74, ftw060. [Google Scholar] [CrossRef] [Green Version]
- Manda-Handzlik, A.; Demkow, U. Neutrophils: The Role of Oxidative and Nitrosative Stress in Health and Disease. Pulm. Infect. 2015, 857, 51–60. [Google Scholar]
- Mortaz, E.; Alipoor, S.D.; Adcock, I.M.; Mumby, S.; Koenderman, L. Update on Neutrophil Function in Severe Inflammation. Front. Immunol. 2018, 9, 2171. [Google Scholar] [CrossRef] [Green Version]
- Ravindran, M.; Khan, M.A.; Palaniyar, N. Neutrophil Extracellular Trap Formation: Physiology, Pathology, and Pharmacology. Biomolecules 2019, 9, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosales, C. Neutrophil: A Cell with Many Roles in Inflammation or Several Cell Types? Front. Physiol. 2018, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Vorobjeva, N.V.; Chernyak, B.V. NETosis: Molecular Mechanisms, Role in Physiology and Pathology. Biochemistry 2020, 85, 1178–1190. [Google Scholar] [CrossRef]
- Wang, J. Neutrophils in Tissue Injury and Repair. Cell Tissue Res. 2018, 371, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Flores, C.A.; Zappellini, A.; Prado-Franceschi, J. Lipoxygenase-Derived Mediators May Be Involved in in Vivo Neutrophil Migration Induced by Bothrops Erythromelas and Bothrops Alternatus Venoms. Toxicon 1993, 31, 1551–1559. [Google Scholar] [CrossRef] [PubMed]
- Echeverría, S.; Leiguez, E.; Guijas, C.; do Nascimento, N.G.; Acosta, O.; Teixeira, C.; Leiva, L.C.; Rodríguez, J.P. Evaluation of Pro-Inflammatory Events Induced by Bothrops Alternatus Snake Venom. Chem. Biol. Interact. 2018, 281, 24–31. [Google Scholar] [CrossRef]
- Farsky, S.H.P.; Gonçalves, L.R.C.; Gutiérrez, J.M.; Correa, A.P.; Rucavado, A.; Gasque, P.; Tambourgi, D.V. Bothrops Asper Snake Venom and Its Metalloproteinase BaP–1 Activate the Complement System. Role in Leucocyte Recruitment. Mediat. Inflamm. 2000, 9, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Zamuner, S.R.; Teixeira, C.F.P. Cell Adhesion Molecules Involved in the Leukocyte Recruitment Induced by Venom of the Snake Bothrops Jararaca. Mediat. Inflamm. 2002, 11, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Escocard, R.D.; Kanashiro, M.M.; Petretski, J.H.; Azevedo-Silva, J.; de Carvalho, E.C.; da Silva, W.D.; Kipnis, T.L. Neutrophils Regulate the Expression of Cytokines, Chemokines and Nitric Oxide Synthase/Nitric Oxide in Mice Injected with Bothrops Atrox Venom. Immunobiology 2006, 211, 37–46. [Google Scholar] [CrossRef]
- Zamuner, S.R.; Gutiérrez, J.M.; Muscará, M.N.; Teixeira, S.A.; Teixeira, C.F.P. Bothrops Asper and Bothrops Jararaca Snake Venoms Trigger Microbicidal Functions of Peritoneal Leukocytes in Vivo. Toxicon 2001, 39, 1505–1513. [Google Scholar] [CrossRef]
- Wanderley, C.W.S.; Silva, C.M.S.; Wong, D.V.T.; Ximenes, R.M.; Morelo, D.F.C.; Cosker, F.; Aragão, K.S.; Fernandes, C.; Palheta-Júnior, R.C.; Havt, A.; et al. Bothrops Jararacussu Snake Venom-Induces a Local Inflammatory Response in a Prostanoid- and Neutrophil-Dependent Manner. Toxicon 2014, 90, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Arruda, V.A.; de Queiroz Guimarães, A.; Hyslop, S.; de Araújo, P.M.; Bon, C.; de Araújo, A.L. de Bothrops Lanceolatus (Fer de Lance) Venom Stimulates Leukocyte Migration into the Peritoneal Cavity of Mice. Toxicon 2003, 41, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Porto, B.N.; Telli, C.A.; Dutra, T.P.; Alves, L.S.; Bozza, M.T.; Fin, C.A.; Thiesen, F.V.; Renner, M.F. Biochemical and Biological Characterization of the Venoms of Bothriopsis Bilineata and Bothriopsis Taeniata (Serpentes: Viperidae). Toxicon 2007, 50, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Ni, Z.; Hu, Y.; Liang, W.; Ou, C.; He, J.; Liu, L.; Shan, H.; Lei, C.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef]
- Chaudhary, R.; Garg, J.; Houghton, D.E.; Murad, M.H.; Kondur, A.; Chaudhary, R.; Wysokinski, W.E.; McBane, R.D. Thromboinflammatory Biomarkers in COVID-19: Systematic Review and Meta-Analysis of 17,052 Patients. Mayo Clin Proc. Innov. Qual. Outcomes 2021, 5, 388–402. [Google Scholar] [CrossRef] [PubMed]
- Karki, R.; Kanneganti, T.-D. The ‘Cytokine Storm’: Molecular Mechanisms and Therapeutic Prospects. Trends Immunol. 2021, 42, 681–705. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine Storm and Leukocyte Changes in Mild versus Severe SARS-CoV-2 Infection: Review of 3939 COVID-19 Patients in China and Emerging Pathogenesis and Therapy Concepts. J. Leukoc. Biol. 2020, 108, 17–41. [Google Scholar] [CrossRef]
- Sun, X.; Wang, T.; Cai, D.; Hu, Z.; Chen, J.; Liao, H.; Zhi, L.; Wei, H.; Zhang, Z.; Qiu, Y.; et al. Cytokine Storm Intervention in the Early Stages of COVID-19 Pneumonia. Cytokine Growth Factor Rev. 2020, 53, 38–42. [Google Scholar] [CrossRef]
- Azkur, A.K.; Akdis, M.; Azkur, D.; Sokolowska, M.; Veen, W.; Brüggen, M.; O’Mahony, L.; Gao, Y.; Nadeau, K.; Akdis, C.A. Immune Response to SARS-CoV-2 and Mechanisms of Immunopathological Changes in COVID-19. Allergy 2020, 75, 1564–1581. [Google Scholar] [CrossRef]
- Gillot, C.; Favresse, J.; Mullier, F.; Lecompte, T.; Dogné, J.-M.; Douxfils, J. NETosis and the Immune System in COVID-19: Mechanisms and Potential Treatments. Front. Pharmacol. 2021, 12, 708302. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]
- Luna, K.; da Silva, M.; Pereira, V. Clinical and Immunological Aspects of Envenomations by Bothrops Snakes. J. Venom. Anim. Toxins Incl. Trop. Dis. 2011, 17, 130–141. [Google Scholar] [CrossRef] [Green Version]
- Júnior, F.N.; Jorge, A.R.; Marinho, A.D.; Silveira, J.D.; Alves, N.T.; Costa, P.H.; Chaves-Filho, A.J.; Lima, D.B.; Sampaio, T.L.; de Morais, G.B.; et al. Bothrops Alternatus Snake Venom Induces Cytokine Expression and Oxidative Stress on Renal Function. Curr. Top. Med. Chem. 2019, 19, 2058–2068. [Google Scholar] [CrossRef] [PubMed]
- Cedro, R.C.A.; Menaldo, D.L.; Costa, T.R.; Zoccal, K.F.; Sartim, M.A.; Santos-Filho, N.A.; Faccioli, L.H.; Sampaio, S.V. Cytotoxic and Inflammatory Potential of a Phospholipase A2 from Bothrops Jararaca Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 33. [Google Scholar] [CrossRef] [Green Version]
- Zuliani, J.P.; Fernandes, C.M.; Zamuner, S.R.; Gutiérrez, J.M.; Teixeira, C.F.P. Inflammatory Events Induced by Lys-49 and Asp-49 Phospholipases A2 Isolated from Bothrops Asper Snake Venom: Role of Catalytic Activity. Toxicon 2005, 45, 335–346. [Google Scholar] [CrossRef]
- Maria Fernandes, C.; Regina Zamuner, S.; Pavan Zuliani, J.; Rucavado, A.; Maria Gutiérrez, J.; de Fátima Pereira Teixeira, C. Inflammatory Effects of BaP1 a Metalloproteinase Isolated from Bothrops Asper Snake Venom: Leukocyte Recruitment and Release of Cytokines. Toxicon 2006, 47, 549–559. [Google Scholar] [CrossRef]
- Rucavado, A.; Escalante, T.; Teixeira, C.F.P.; Fernándes, C.M.; Díaz, C.; Gutiérrez, J.M. Increments in Cytokines and Matrix Metalloproteinases in Skeletal Muscle after Injection of Tissue-Damaging Toxins from the Venom of the Snake Bothrops Asper. Mediat. Inflamm 2002, 11, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Moreira, V.; Dos-Santos, M.C.; Nascimento, N.G.; da Silva, H.B.; Fernandes, C.M.; D’Império Lima, M.R.; Teixeira, C. Local Inflammatory Events Induced by Bothrops Atrox Snake Venom and the Release of Distinct Classes of Inflammatory Mediators. Toxicon 2012, 60, 12–20. [Google Scholar] [CrossRef]
- Luna, K.P.; Melo, C.M.; Pascoal, V.P.; Martins Filho, O.A.; Pereira, V.R. Bothrops Erythromelas Snake Venom Induces a Proinflammatory Response in Mice Splenocytes. Int. J. Interferon Cytokine Mediat. Res. 2011, 9, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Delafontaine, M.; Villas-Boas, I.; Mathieu, L.; Josset, P.; Blomet, J.; Tambourgi, D. Enzymatic and Pro-Inflammatory Activities of Bothrops Lanceolatus Venom: Relevance for Envenomation. Toxins 2017, 9, 244. [Google Scholar] [CrossRef] [Green Version]
- Nadur-Andrade, N.; Dale, C.S.; Oliveira, V.R.; Toniolo, E.F.; Feliciano, R.D.; da Silva, J.A., Jr.; Zamuner, S.R. Analgesic Effect of Photobiomodulation on Bothrops Moojeni Venom-Induced Hyperalgesia: A Mechanism Dependent on Neuronal Inhibition, Cytokines and Kinin Receptors Modulation. PLoS Negl. Trop. Dis. 2016, 10, e0004998. [Google Scholar] [CrossRef] [Green Version]
- Soares, F.G.S.; Ibiapina, H.N.; Sartim, M.A.; Mendonça-da-Silva, I.; Nascimento, E.F.; Ferreira, L.C.L.; Cerni, F.A.; Malheiro, A.; Pucca, M.B.; Wen, F.H.; et al. Lower Levels of CXCL-8 and IL-2 on Admission as Predictors of Early Adverse Reactions to Bothrops Antivenom in the Brazilian Amazon. Cytokine 2022, 152, 155825. [Google Scholar] [CrossRef]
- Lee, D.W.; Gardner, R.; Porter, D.L.; Louis, C.U.; Ahmed, N.; Jensen, M.; Grupp, S.A.; Mackall, C.L. Current Concepts in the Diagnosis and Management of Cytokine Release Syndrome. Blood 2014, 124, 188–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Templin, C.; Ghadri, J.R.; Diekmann, J.; Napp, L.C.; Bataiosu, D.R.; Jaguszewski, M.; Cammann, V.L.; Sarcon, A.; Geyer, V.; Neumann, C.A.; et al. Clinical Features and Outcomes of Takotsubo (Stress) Cardiomyopathy. N. Engl. J. Med. 2015, 373, 929–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burkard, P.; Vögtle, T.; Nieswandt, B. Platelets in Thrombo-Inflammation: Concepts, Mechanisms, and Therapeutic Strategies for Ischemic Stroke. Hamostaseologie 2020, 40, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Periayah, M.H.; Halim, A.S.; Zaharil, A.; Saad, M. Mechanism Action of Platelets and Crucial Blood Coagulation Pathways in Hemostasis. Int. J. Hematol.-Oncol. Stem Cell Res. 2017, 11, 319. [Google Scholar]
- Andersen, H.; Greenberg, D.L.; Fujikawa, K.; Xu, W.; Chung, D.W.; Davie, E.W. Protease-Activated Receptor 1 Is the Primary Mediator of Thrombin-Stimulated Platelet Procoagulant Activity. Proc. Natl. Acad. Sci. USA 1999, 96, 11189–11193. [Google Scholar] [CrossRef] [PubMed]
- Heuberger, D.M.; Schuepbach, R.A. Protease-Activated Receptors (PARs): Mechanisms of Action and Potential Therapeutic Modulators in PAR-Driven Inflammatory Diseases. Thromb. J. 2019, 17, 4. [Google Scholar] [CrossRef] [Green Version]
- Clemetson, K.J.; Clemetson, J.M. Platelet Receptors. In Platelets; Elsevier: Amsterdam, The Netherlands, 2019; pp. 169–192. [Google Scholar]
- Cameron-Vendrig, A.; Reheman, A.; Siraj, M.A.; Xu, X.R.; Wang, Y.; Lei, X.; Afroze, T.; Shikatani, E.; El-Mounayri, O.; Noyan, H.; et al. Glucagon-Like Peptide 1 Receptor Activation Attenuates Platelet Aggregation and Thrombosis. Diabetes 2016, 65, 1714–1723. [Google Scholar] [CrossRef] [Green Version]
- Margraf, A.; Zarbock, A. Platelets in Inflammation and Resolution. J. Immunol. 2019, 203, 2357–2367. [Google Scholar] [CrossRef]
- Mezger, M.; Nording, H.; Sauter, R.; Graf, T.; Heim, C.; von Bubnoff, N.; Ensminger, S.M.; Langer, H.F. Platelets and Immune Responses During Thromboinflammation. Front. Immunol. 2019, 10, 1731. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.R.; Zhang, D.; Oswald, B.E.; Carrim, N.; Wang, X.; Hou, Y.; Zhang, Q.; Lavalle, C.; McKeown, T.; Marshall, A.H.; et al. Platelets Are Versatile Cells: New Discoveries in Hemostasis, Thrombosis, Immune Responses, Tumor Metastasis and Beyond. Crit. Rev. Clin. Lab. Sci. 2016, 53, 409–430. [Google Scholar] [CrossRef] [PubMed]
- Arbesu, I.; Bucsaiova, M.; Fischer, M.B.; Mannhalter, C. Platelet-Borne Complement Proteins and Their Role in Platelet-Bacteria Interactions. J. Thromb. Haemost. 2016, 14, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Kozarcanin, H.; Lood, C.; Munthe-Fog, L.; Sandholm, K.; Hamad, O.A.; Bengtsson, A.A.; Skjoedt, M.-O.; Huber-Lang, M.; Garred, P.; Ekdahl, K.N.; et al. The Lectin Complement Pathway Serine Proteases (MASPs) Represent a Possible Crossroad between the Coagulation and Complement Systems in Thromboinflammation. J. Thromb. Haemost. 2016, 14, 531–545. [Google Scholar] [CrossRef] [Green Version]
- Hamad, O.A.; Ekdahl, K.N.; Nilsson, P.H.; Andersson, J.; Magotti, P.; Lambris, J.D.; Nilsson, B. Complement Activation Triggered by Chondroitin Sulfate Released by Thrombin Receptor-Activated Platelets. J. Thromb. Haemost. 2008, 6, 1413–1421. [Google Scholar] [CrossRef] [Green Version]
- del Conde, I.; Crúz, M.A.; Zhang, H.; López, J.A.; Afshar-Kharghan, V. Platelet Activation Leads to Activation and Propagation of the Complement System. J. Exp. Med. 2005, 201, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Peerschke, E.I.B.; Ghebrehiwet, B. Platelet Receptors for the Complement Component C1q: Implications for Hemostasis and Thrombosis. Immunobiology 1998, 199, 239–249. [Google Scholar] [CrossRef]
- Peerschke, E.I.B.; Yin, W.; Grigg, S.E.; Ghebrehiwet, B. Blood Platelets Activate the Classical Pathway of Human Complement. J. Thromb. Haemost. 2006, 4, 2035–2042. [Google Scholar] [CrossRef]
- Zuchtriegel, G.; Uhl, B.; Puhr-Westerheide, D.; Pörnbacher, M.; Lauber, K.; Krombach, F.; Reichel, C.A. Platelets Guide Leukocytes to Their Sites of Extravasation. PLoS Biol 2016, 14, e1002459. [Google Scholar] [CrossRef] [Green Version]
- Sreeramkumar, V.; Adrover, J.M.; Ballesteros, I.; Cuartero, M.I.; Rossaint, J.; Bilbao, I.; Nácher, M.; Pitaval, C.; Radovanovic, I.; Fukui, Y.; et al. Neutrophils Scan for Activated Platelets to Initiate Inflammation. Science (1979) 2014, 346, 1234–1238. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Gao, H.; Shi, C.; Erhardt, P.W.; Pavlovsky, A.; Soloviev, D.A.; Bledzka, K.; Ustinov, V.; Zhu, L.; Qin, J.; et al. Leukocyte Integrin Mac-1 Regulates Thrombosis via Interaction with Platelet GPIbα. Nat. Commun. 2017, 8, 15559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, K.M.; Nogueira, T.O.; Senise, L.V.; Cirillo, M.C.; Gonçalves, L.R.C.; Sano-Martins, I.S.; Giorgi, R.; Santoro, M.L. Involvement of Circulating Platelets on the Hyperalgesic Response Evoked by Carrageenan and Bothrops Jararaca Snake Venom. J. Thromb. Haemost. 2011, 9, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Sano-Martins, I. Platelet Dysfunction during Bothrops Jararaca Snake Envenomation in Rabbits. Thromb. Haemost. 2004, 92, 369–383. [Google Scholar] [CrossRef] [PubMed]
- Santos-Filho, N.A.; Silveira, L.B.; Oliveira, C.Z.; Bernardes, C.P.; Menaldo, D.L.; Fuly, A.L.; Arantes, E.C.; Sampaio, S.V.; Mamede, C.C.N.; Beletti, M.E.; et al. A New Acidic Myotoxic, Anti-Platelet and Prostaglandin I2 Inductor Phospholipase A2 Isolated from Bothrops Moojeni Snake Venom. Toxicon 2008, 52, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Silveira, L.B.; Marchi-Salvador, D.P.; Santos-Filho, N.A.; Silva, F.P.; Marcussi, S.; Fuly, A.L.; Nomizo, A.; da Silva, S.L.; Stábeli, R.G.; Arantes, E.C.; et al. Isolation and Expression of a Hypotensive and Anti-Platelet Acidic Phospholipase A2 from Bothrops Moojeni Snake Venom. J. Pharm. Biomed. Anal. 2013, 73, 35–43. [Google Scholar] [CrossRef]
- Andrião-Escarso, S.H.; Soares, A.M.; Fontes, M.R.M.; Fuly, A.L.; Corrêa, F.M.A.; Rosa, J.C.; Greene, L.J.; Giglio, J.R. Structural and Functional Characterization of an Acidic Platelet Aggregation Inhibitor and Hypotensive Phospholipase A2 from Bothrops jararacussu Snake Venom. Biochem.Pharm. 2002, 64, 723–732. [Google Scholar] [CrossRef]
- Serrano, S.M.T.; Reichl, A.P.; Mentele, R.; Auerswald, E.A.; Santoro, M.L.; Sampaio, C.A.M.; Camargo, A.C.M.; Assakura, M.T. A Novel Phospholipase A2, BJ-PLA2, from the Venom of the Snake Bothrops Jararaca: Purification, Primary Structure Analysis, and Its Characterization as a Platelet-Aggregation-Inhibiting Factor. Arch. Biochem. Biophys. 1999, 367, 26–32. [Google Scholar] [CrossRef]
- Ferreira, F.; Gomes, M.; de Souza, D.; Gimenes, S.; Castanheira, L.; Borges, M.; Rodrigues, R.; Yoneyama, K.; Brandeburgo, M.; Rodrigues, V. Molecular Cloning and Pharmacological Properties of an Acidic PLA2 from Bothrops pauloensis Snake Venom. Toxins 2013, 5, 2403–2419. [Google Scholar] [CrossRef] [Green Version]
- Fuly, A.L.; Soares, A.M.; Marcussi, S.; Giglio, J.R.; Guimarães, J.A. Signal Transduction Pathways Involved in the Platelet Aggregation Induced by a D-49 Phospholipase A2 Isolated from Bothrops jararacussu Snake Venom. Biochimie 2004, 86, 731–739. [Google Scholar] [CrossRef]
- Deluca, M.; Ward, C.M.; Ohmori, K.; Andrews, R.K.; Berndt, M.C. Jararhagin and Jaracetin: Novel Snake Venom Inhibitors of the Integrin Collagen Receptor, A2β1. Biochem. Biophys. Res. Commun. 1995, 206, 570–576. [Google Scholar] [CrossRef]
- Souza, D.H.F.; Iemma, M.R.C.; Ferreira, L.L.; Faria, J.P.; Oliva, M.L.V.; Zingali, R.B.; Niewiarowski, S.; Selistre-de-Araujo, H.S. The Disintegrin-like Domain of the Snake Venom Metalloprotease Alternagin Inhibits A2β1 Integrin-Mediated Cell Adhesion. Arch. Biochem. Biophys. 2000, 384, 341–350. [Google Scholar] [CrossRef]
- Sanchez, E.F.; Richardson, M.; Gremski, L.H.; Veiga, S.S.; Yarleque, A.; Niland, S.; Lima, A.M.; Estevao-Costa, M.I.; Eble, J.A. A Novel Fibrinolytic Metalloproteinase, Barnettlysin-I from Bothrops Barnetti (Barnett´s Pitviper) Snake Venom with Anti-Platelet Properties. Biochim. Et Biophys. Acta (BBA)–Gen. Subj. 2016, 1860, 542–556. [Google Scholar] [CrossRef] [PubMed]
- Sobrinho, J.C.; Kayano, A.M.; Simões-Silva, R.; Alfonso, J.J.; Gomez, A.F.; Gomez, M.C.V.; Zanchi, F.B.; Moura, L.A.; Souza, V.R.; Fuly, A.L.; et al. Anti-platelet aggregation activity of two novel acidic Asp49-phospholipases A2 from Bothrops brazili snake venom. Int. J. Biol. Macromol. 2018, 107 Pt A, 1014–1022. [Google Scholar] [CrossRef]
- Dias, E.H.V.; dos Santos Paschoal, T.; da Silva, A.P.; da Cunha Pereira, D.F.; de Sousa Simamoto, B.B.; Matias, M.S.; Santiago, F.M.; Rosa, J.C.; Soares, A.; Santos-Filho, N.A.; et al. BaltPLA2: A New Phospholipase A2 from Bothrops alternatus Snake Venom with Antiplatelet Aggregation Activity. Protein Pept. Lett. 2018, 25, 943–952. [Google Scholar] [CrossRef] [PubMed]
- Menaldo, D.L.; Bernardes, C.P.; Santos-Filho, N.A.; de Andrade Moura, L.; Fuly, A.L.; Arantes, E.C.; Sampaio, S.V. Biochemical Characterization and Comparative Analysis of Two Distinct Serine Proteases from Bothrops pirajai Snake Venom. Biochimie 2012, 94, 2545–2558. [Google Scholar] [CrossRef] [PubMed]
- Vilca-Quispe, A.; Ponce-Soto, L.A.; Winck, F.V.; Marangoni, S. Isolation and Characterization of a New Serine Protease with Thrombin-like Activity (TLBm) from the Venom of the Snake Bothrops marajoensis. Toxicon 2010, 55, 745–753. [Google Scholar] [CrossRef] [PubMed]
- Santos, B.F.; Serrano, S.M.T.; Kuliopulos, A.; Niewiarowski, S. Interaction of Viper Venom Serine Peptidases with Thrombin Receptors on Human Platelets. FEBS Lett. 2000, 477, 199–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marrakchi, N.; Barbouche, R.; Guermazi, S.; Karoui, H.; Bon, C.; el Ayeb, M. Cerastotin, a Serine Protease from Cerastes Cerastes Venom, with Platelet-Aggregating and Agglutinating Properties. Eur. J. Biochem. 1997, 247, 121–128. [Google Scholar] [CrossRef]
- Nishida, S.; Fujimura, Y.; Miura, S.; Yoshida, E.; Sugimoto, M.; Yoshioka, A.; Fukui, H.; Ozaki, Y.; Usami, Y. Purification and Characterization of Bothrombin, a Fibrinogen-Clotting Serine Protease from the Venom of Bothrops Jararaca. Biochemistry 1994, 33, 1843–1849. [Google Scholar] [CrossRef]
- de Queiroz, M.R.; Mamede, C.C.N.; Fonseca, K.C.; de Morais, N.C.G.; de Sousa, B.B.; Santos-Filho, N.A.; Beletti, M.E.; Arantes, E.C.; Stanziola, L.; de Oliveira, F. Rapid Purification of a New P-I Class Metalloproteinase from Bothrops Moojeni Venom with Antiplatelet Activity. Biomed.Res. Int. 2014, 2014, 352420. [Google Scholar] [CrossRef] [Green Version]
- Tanjoni, I.; Butera, D.; Bento, L.; Della-Casa, M.S.; Marques-Porto, R.; Takehara, H.A.; Gutiérrez, J.M.; Fernandes, I.; Moura-da-Silva, A.M. Snake Venom Metalloproteinases: Structure/Function Relationships Studies Using Monoclonal Antibodies. Toxicon 2003, 42, 801–808. [Google Scholar] [CrossRef]
- Sousa, B.B.; Mamede, C.C.; Matias, M.S.; Pereira, D.F.; Queiroz, M.R.; Dias, E.H.; Silva, A.C.; Dantas, N.O.; Costa, J.D.; Oliveira, F.D. A New Platelet-Aggregation-Inhibiting Factor Isolated from Bothrops moojeni Snake Venom. Biomed. Res. Int. 2017, 2017, 4315832. [Google Scholar] [CrossRef] [PubMed]
- Matias, M.S.; de Sousa, B.B.; da Cunha Pereira, D.F.; Dias, E.H.V.; Mamede, C.C.N.; de Queiroz, M.R.; Silva, A.C.A.; Dantas, N.O.; Soares, A.M.; de Oliveira Costa, J.; et al. BaltDC: Purification, Characterization and Infrared Spectroscopy of an Antiplatelet DC Protein Isolated from Bothrops Alternatus Snake Venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sartim, M.A.; Costa, T.R.; Laure, H.J.; Espíndola, M.S.; Frantz, F.G.; Sorgi, C.A.; Cintra, A.C.O.; Arantes, E.C.; Faccioli, L.H.; Rosa, J.C.; et al. Moojenactivase, a Novel pro-Coagulant PIIId Metalloprotease Isolated from Bothrops moojeni Snake Venom, Activates Coagulation Factors II and X and Induces Tissue Factor up-Regulation in Leukocytes. Arch. Toxicol. 2016, 90, 1261–1278. [Google Scholar] [CrossRef]
- Oliveira, L.S.; Estevão-Costa, M.I.; Alvarenga, V.G.; Vivas-Ruiz, D.E.; Yarleque, A.; Lima, A.M.; Cavaco, A.; Eble, J.A.; Sanchez, E.F. Atroxlysin-III, A Metalloproteinase from the Venom of the Peruvian Pit Viper Snake Bothrops atrox (Jergón) Induces Glycoprotein VI Shedding and Impairs Platelet Function. Molecule 2019, 26, 3489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suntravat, M.; Helmke, T.J.; Atphaisit, C.; Cuevas, E.; Lucena, S.E.; Uzcátegui, N.L.; Sánchez, E.E.; Rodriguez-Acosta, A. Expression, Purification, and Analysis of Three Recombinant ECD Disintegrins (r-Colombistatins) from P-III Class Snake Venom Metalloproteinases Affecting Platelet Aggregation and SK-MEL-28 Cell Adhesion. Toxicon 2016, 122, 43–49. [Google Scholar] [CrossRef] [Green Version]
- Izidoro, L.F.M.; Ribeiro, M.C.; Souza, G.R.L.; Sant’Ana, C.D.; Hamaguchi, A.; Homsi-Brandeburgo, M.I.; Goulart, L.R.; Beleboni, R.O.; Nomizo, A.; Sampaio, S.V.; et al. Biochemical and Functional Characterization of an L-Amino Acid Oxidase Isolated from Bothrops pirajai Snake Venom. Bioorg. Med. Chem. 2006, 14, 7034–7043. [Google Scholar] [CrossRef]
- Rodrigues, R.S.; da Silva, J.F.; Boldrini França, J.; Fonseca, F.P.P.; Otaviano, A.R.; Henrique Silva, F.; Hamaguchi, A.; Magro, A.J.; Braz, A.S.K.; dos Santos, J.I. Structural and Functional Properties of Bp-LAAO, a New l-Amino Acid Oxidase Isolated from Bothrops pauloensis Snake Venom. Biochimie 2009, 91, 490–501. [Google Scholar] [CrossRef]
- Stábeli, R.G.; Sant’Ana, C.D.; Ribeiro, P.H.; Costa, T.R.; Ticli, F.K.; Pires, M.G.; Nomizo, A.; Albuquerque, S.; Malta-Neto, N.R.; Marins, M.; et al. Cytotoxic L-Amino Acid Oxidase from Bothrops moojeni: Biochemical and Functional Characterization. Int. J. Biol.Macromol. 2007, 41, 132–140. [Google Scholar] [CrossRef]
- Naumann, G.B.; Silva, L.F.; Silva, L.; Faria, G.; Richardson, M.; Evangelista, K.; Kohlhoff, M.; Gontijo, C.M.F.; Navdaev, A.; de Rezende, F.F.; et al. Cytotoxicity and Inhibition of Platelet Aggregation Caused by an L-Amino Acid Oxidase from Bothrops leucurus Venom. Biochim. Et Biophys. Acta (BBA)–Gen. Subj. 2011, 1810, 683–694. [Google Scholar] [CrossRef] [Green Version]
- Read, M.; Smith, S.; Lamb, M.; Brinkhous, K. Role of Botrocetin in Platelet Agglutination: Formation of an Activated Complex of Botrocetin and von Willebrand Factor. Blood 1989, 74, 1031–1035. [Google Scholar] [CrossRef] [PubMed]
- da Cunha Pereira, D.F.; Ribeiro, M.S.; de Sousa Simamoto, B.B.; Dias, E.H.; de Oliveira Costa, J.; Santos-Filho, N.A.; Bordon, K.D.; Arantes, E.C.; Dantas, N.O.; Silva, A.C.; et al. Baltetin: A New C-Type Lectin-like Isolated from Bothrops alternatus Snake Venom Which Act as a Platelet Aggregation Inhibiting. J. Chromatogr. B 2021, 1173, 122695. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, T.; Kimura, H.; Shikama, Y.; Uchida, T.; Kariyone, S.; Hirano, T.; Kishimoto, T.; Takatsuki, F.; Akiyama, Y. Interleukin-6 Is a Potent Thrombopoietic Factor in Vivo in Mice. Blood 1989, 74, 1241–1244. [Google Scholar] [CrossRef] [Green Version]
- Asano, S.; Okano, A.; Ozawa, K.; Nakahata, T.; Ishibashi, T.; Koike, K.; Kimura, H.; Tanioka, Y.; Shibuya, A.; Hirano, T. In Vivo Effects of Recombinant Human Interleukin-6 in Primates: Stimulated Production of Platelets. Blood 1990, 75, 1602–1605. [Google Scholar] [CrossRef] [Green Version]
- Hill, R.J.; Warren, M.K.; Levin, J. Stimulation of Thrombopoiesis in Mice by Human Recombinant Interleukin 6. J. Clin. Investig. 1990, 85, 1242–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petricevich, V.L.; Teixeira, C.F.P.; Tambourgi, D.V.; Gutiérrez, J.M. Increments in Serum Cytokine and Nitric Oxide Levels in Mice Injected with Bothrops Asper and Bothrops Jararaca Snake Venoms. Toxicon 2000, 38, 1253–1266. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Agüero, M.L.; París, M.M.; Hu, S.; Peterson, P.K.; Gutiérrez, J.M.; Lomonte, B.; Faingezicht, I. Systemic Cytokine Response in Children Bitten by Snakes in Costa Rica. Pediatr. Emerg. Care 2001, 17, 425–429. [Google Scholar] [CrossRef]
- Barraviera, B.; Lomonte, B.; Tarkowski, A.; Hanson, L.Å.; Meira, D.A. acute-phase reactions, including cytokines, in patients bitten by Bothrops and Crotalus snakes in Brazil. J. Venom. Anim. Toxins 1995, 1, 11–22. [Google Scholar] [CrossRef]
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408. [Google Scholar] [CrossRef]
- Wellmann, I.A.M.; Ibiapina, H.N.S.; Sachett, J.A.G.; Sartim, M.A.; Silva, I.M.; Oliveira, S.S.; Tarragô, A.M.; Moura-da-Silva, A.M.; Lacerda, M.V.G.; Ferreira, L.C.; et al. Correlating Fibrinogen Consumption and Profiles of Inflammatory Molecules in Human Envenomation’s by Bothrops Atrox in the Brazilian Amazon. Front. Immunol 2020, 11, 1874. [Google Scholar] [CrossRef]
- Grover, S.P.; Mackman, N. Tissue Factor. Arter. Thromb. Vasc. Biol. 2018, 38, 709–725. [Google Scholar] [CrossRef] [Green Version]
- Zelaya, H.; Rothmeier, A.S.; Ruf, W. Tissue Factor at the Crossroad of Coagulation and Cell Signaling. J. Thromb. Haemost. 2018, 16, 1941–1952. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, K.M.; Alves, A.F.; Barbaro, K.C.; Santoro, M.L. Bothrops Jararaca Venom Metalloproteinases Are Essential for Coagulopathy and Increase Plasma Tissue Factor Levels during Envenomation. PLoS Negl. Trop. Dis. 2014, 8, e2814. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.L.M.; Fritzen, M.; Faria, F.; Da Motta, G.; Chudzinski-Tavassi, A.M. Releasing or Expression Modulating Mediator Involved in Hemostasis by Berythractivase and Jararhagin (SVMPs). Toxicon 2006, 47, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.B.; Schattner, M.; Ramos, C.R.R.; Junqueira-De-Azevedo, I.L.M.; Guarnieri, M.C.; Lazzari, M.A.; Sampaio, C.A.M.; Pozner, R.G.; Ventura, J.S.; Ho, P.L.; et al. A Prothrombin Activator from Bothrops erythromelas (Jararaca-Da-Seca) Snake Venom: Characterization and Molecular Cloning. Biochem. J. 2003, 369, 129–139. [Google Scholar] [CrossRef] [Green Version]
- Cezarette, G.N.; Sartim, M.A.; Sampaio, S.V. Inflammation and Coagulation Crosstalk Induced by BJcuL, a Galactose-Binding Lectin Isolated from Bothrops Jararacussu Snake Venom. Int. J. Biol. Macromol. 2020, 144, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Conran, N.; De Paula, E.V. Thromboinflammatory Mechanisms in Sickle Cell Disease–Challenging the Hemostatic Balance. Haematologica 2020, 105, 2380–2390. [Google Scholar] [CrossRef]
- Cavalcante, J.D.; de Almeida, D.E.; Moraes, M.S.; Santos, S.R.; Pincinato, P.M.; Riciopo, P.M.; de Oliveira, L.L.; Monteiro, W.M.; Ferreira-Junior, R.S. Challenges and Opportunities in Clinical Diagnostic Routine of Envenomation Using Blood Plasma Proteomics. Toxins 2023, 15, 180. [Google Scholar] [CrossRef]
- Kölm, R.; Schaller, M.; Roumenina, L.T.; Niemiec, I.; Kremer Hovinga, J.A.; Khanicheh, E.; Kaufmann, B.A.; Hopfer, H.; Trendelenburg, M. Von Willebrand Factor Interacts with Surface-Bound C1q and Induces Platelet Rolling. J. Immunol. 2016, 197, 3669–3679. [Google Scholar] [CrossRef] [Green Version]
- Rinder, C.S.; Smith, M.J.; Rinder, H.M.; Cortright, D.N.; Brodbeck, R.M.; Krause, J.E.; Smith, B.R. Leukocyte Effects of C5a-Receptor Blockade During Simulated Extracorporeal Circulation. Ann. Thorac. Surg. 2007, 83, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Lappegård, K.T.; Fung, M.; Bergseth, G.; Riesenfeld, J.; Mollnes, T.E. Artificial Surface-Induced Cytokine Synthesis: Effect of Heparin Coating and Complement Inhibition. Ann. Thorac. Surg. 2004, 78, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Riedl, M.; Noone, D.G.; Khan, M.A.; Pluthero, F.G.; Kahr, W.H.A.; Palaniyar, N.; Licht, C. Complement Activation Induces Neutrophil Adhesion and Neutrophil-Platelet Aggregate Formation on Vascular Endothelial Cells. Kidney Int. Rep. 2017, 2, 66–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blatt, A.Z.; Pathan, S.; Ferreira, V.P. Properdin: A Tightly Regulated Critical Inflammatory Modulator. Immunol. Rev. 2016, 274, 172–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Venom | Model | Cytokines | Reference |
|---|---|---|---|
| B. alternatus | mice peritoneal macrophages | TNFα, IL1, IL12 and IL6 | [107] |
| B. alternatus | Raw 264.7 cells | TNFα, IL1, IL12 and IL6 | [107] |
| B. asper | peritoneal cavity of mice | IL-6 and TNF-a | [126] |
| BaP1 from B. asper venom | macrophages in vitro | TNF-a | [128] |
| B. atrox | peritoneal cavity of mice | TNF-a, IL-6, IL-12p70, IL-10 and CCL-2 | [129] |
| B. erythromelas | mice splenocytes | IFN-γ, IL-6, IL-10 and NO | [130] |
| B. jararacussu | footpad samples | TNF-α and IL-1β | [112] |
| B. lanceolatus | human keratinocytes—HaCaT | IL-8, MCP-1, RANTES | [131] |
| B. lanceolatus | endothelial vascular cells—EAhy926 | IL-8, MCP-1, RANTES e IL-6 | [131] |
| B. moojeni | spinal cord | IL-10 | [132] |
| B. moojeni | footpad samples | IL-6, IL-10 and TNF-α | [132] |
| Protein Family | Protein Name | Species | Inhibition (−)/Activation (+) of Aggregation | References |
|---|---|---|---|---|
| PLA2 | Bothropstoxin-II | B. jararacussu | + | [161] |
| BmooPLA2 | B. moojeni | − | [157] | |
| BJ-PLA2 | B. jararaca | − | [136] | |
| BthA-I- PLA2 | B. jararacussu | − | [158] | |
| BE-I-PLA2 | B. erythromelas | − | [68] | |
| BpPLA2-TXI | B. pauloensis | − | [160] | |
| BmooTX-I | B. moojeni | − | [156] | |
| Braziliase-I and II | B. brazili | − | [165] | |
| BJ-PLA2-I | B. jararaca | − | [125] | |
| BaltPLA2 | B. alternatus | − | [166] | |
| SVSP | BpirSP27 | B. pirajai | + | [167] |
| BpirSP41 B | B. pirajai | + | [167] | |
| TLBm | B. marajoensis | + | [168] | |
| PA-BJ | B. jararaca | + | [169] | |
| Thrombocytin | B. jararaca | + | [169] | |
| Cerastotin | B. jararaca | + | [170] | |
| Bothrombin | B. jararaca | + | [171] | |
| Moojase | B. moojeni | + | [80] | |
| SVMP | BmooMPα-II | B. moojeni | − | [172] |
| Jararhagina | B. jararaca | − | [173] | |
| BmooPAi | B. moojeni | − | [174] | |
| Bar-I | B. barnetti | − | [164] | |
| Batroxrhagin | B. atrox | − | [74] | |
| BaltDC | B. alternatus | − | [175] | |
| Moojenactivase | B. moojeni | + | [176] | |
| Atroxlysin-III | B. atrox | − | [177] | |
| r-colombistatins 2,3,4 | B. colombiensis | − | [178] | |
| LAAOs | BpirLAAO-I | B. pirajai | + | [179] |
| Bp-LAAO | B. pauloensis | + | [180] | |
| BmooLAAO-I | B. moojeni | + | [181] | |
| Bl-LAAO | B. leucurus | − | [182] | |
| CTL | Botrocetin | B. jararaca | + | [183] |
| Baltetin | B. alternatus | − | [184] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cavalcante, J.S.; de Almeida, D.E.G.; Santos-Filho, N.A.; Sartim, M.A.; de Almeida Baldo, A.; Brasileiro, L.; Albuquerque, P.L.; Oliveira, S.S.; Sachett, J.A.G.; Monteiro, W.M.; et al. Crosstalk of Inflammation and Coagulation in Bothrops Snakebite Envenoming: Endogenous Signaling Pathways and Pathophysiology. Int. J. Mol. Sci. 2023, 24, 11508. https://doi.org/10.3390/ijms241411508
Cavalcante JS, de Almeida DEG, Santos-Filho NA, Sartim MA, de Almeida Baldo A, Brasileiro L, Albuquerque PL, Oliveira SS, Sachett JAG, Monteiro WM, et al. Crosstalk of Inflammation and Coagulation in Bothrops Snakebite Envenoming: Endogenous Signaling Pathways and Pathophysiology. International Journal of Molecular Sciences. 2023; 24(14):11508. https://doi.org/10.3390/ijms241411508
Chicago/Turabian StyleCavalcante, Joeliton S., Denis Emanuel Garcia de Almeida, Norival A. Santos-Filho, Marco Aurélio Sartim, Amanda de Almeida Baldo, Lisele Brasileiro, Polianna L. Albuquerque, Sâmella S. Oliveira, Jacqueline Almeida Gonçalves Sachett, Wuelton Marcelo Monteiro, and et al. 2023. "Crosstalk of Inflammation and Coagulation in Bothrops Snakebite Envenoming: Endogenous Signaling Pathways and Pathophysiology" International Journal of Molecular Sciences 24, no. 14: 11508. https://doi.org/10.3390/ijms241411508