

Genome-Wide Analysis and Identification of 1-Aminocyclopropane-1-Carboxylate Synthase (ACS) Gene Family in Wheat (Triticum aestivum L.)

Abstract
:1. Introduction
2. Results
2.1. Identification of the ACS Gene Family in Wheat
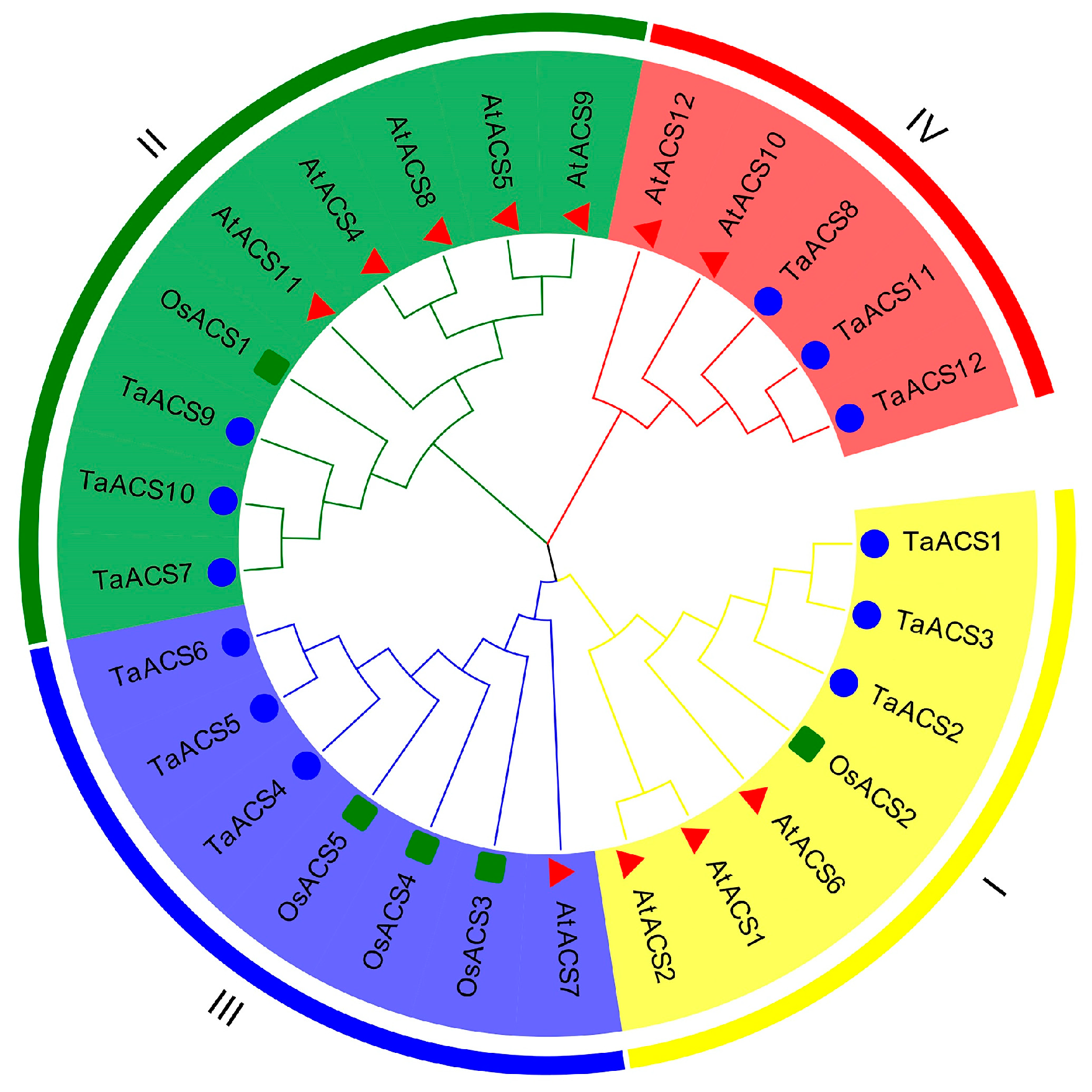
2.2. Phylogenetic Analysis of the ACS Gene Family in Wheat
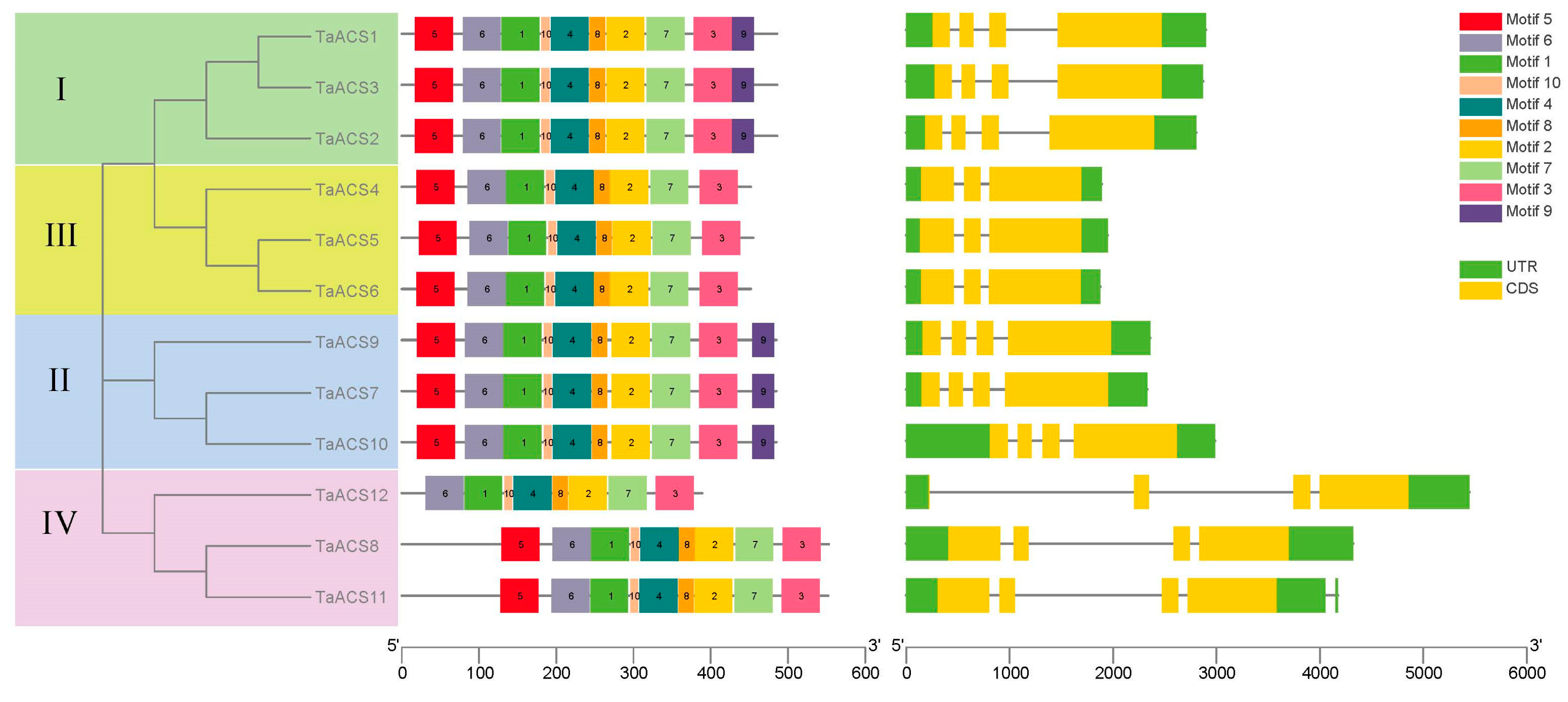
2.3. Analysis of Protein Conserved Motif and Gene Structure of ACS in Wheat
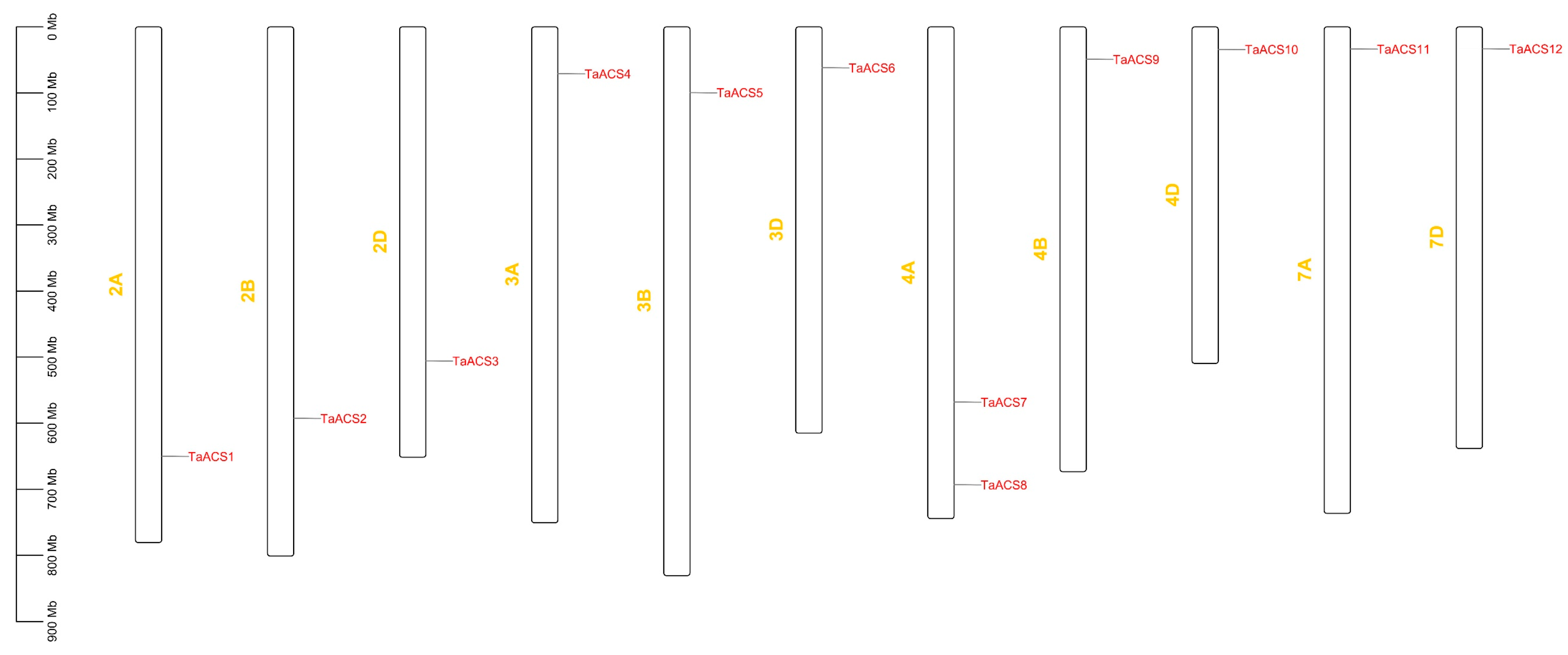
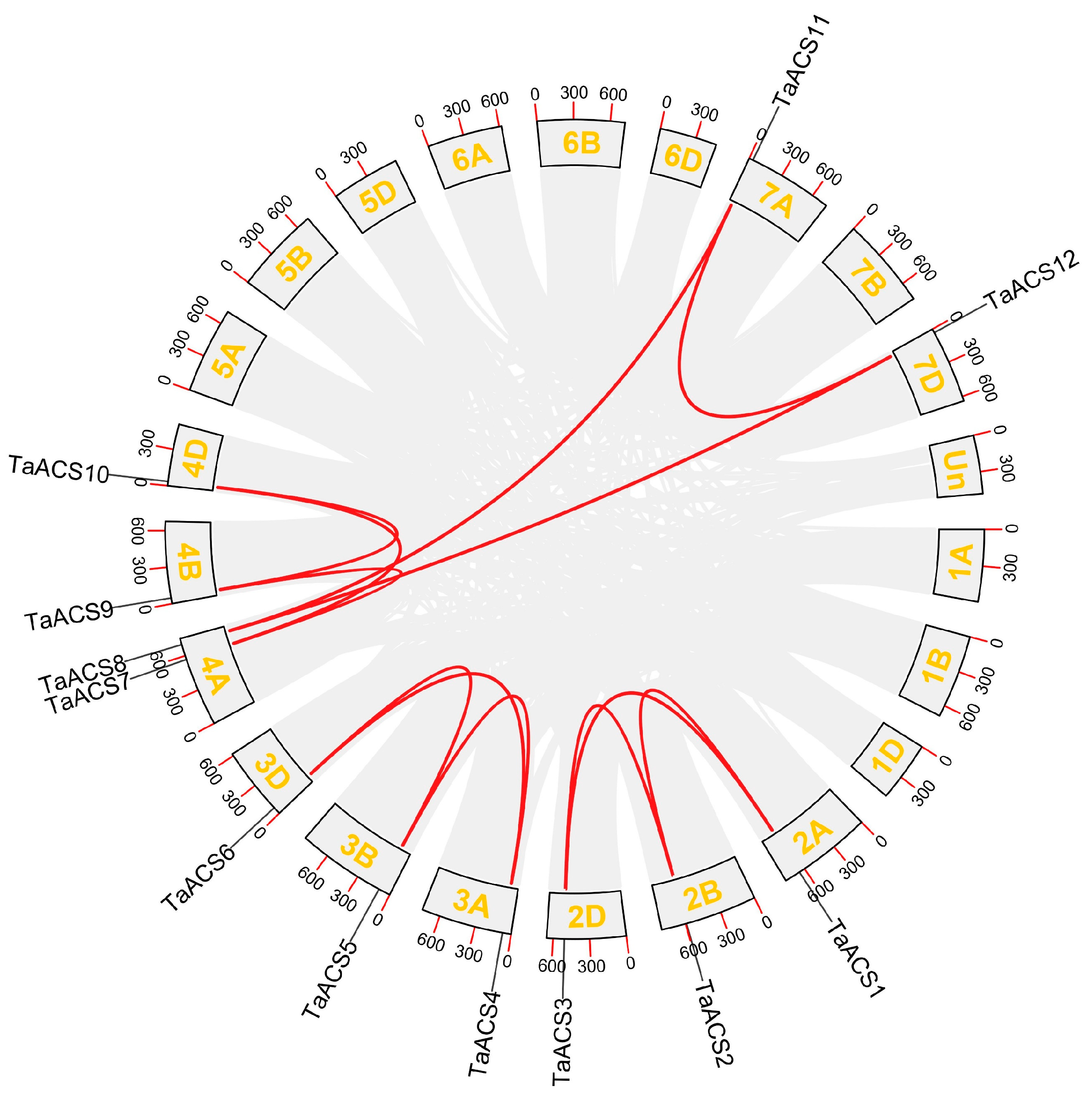
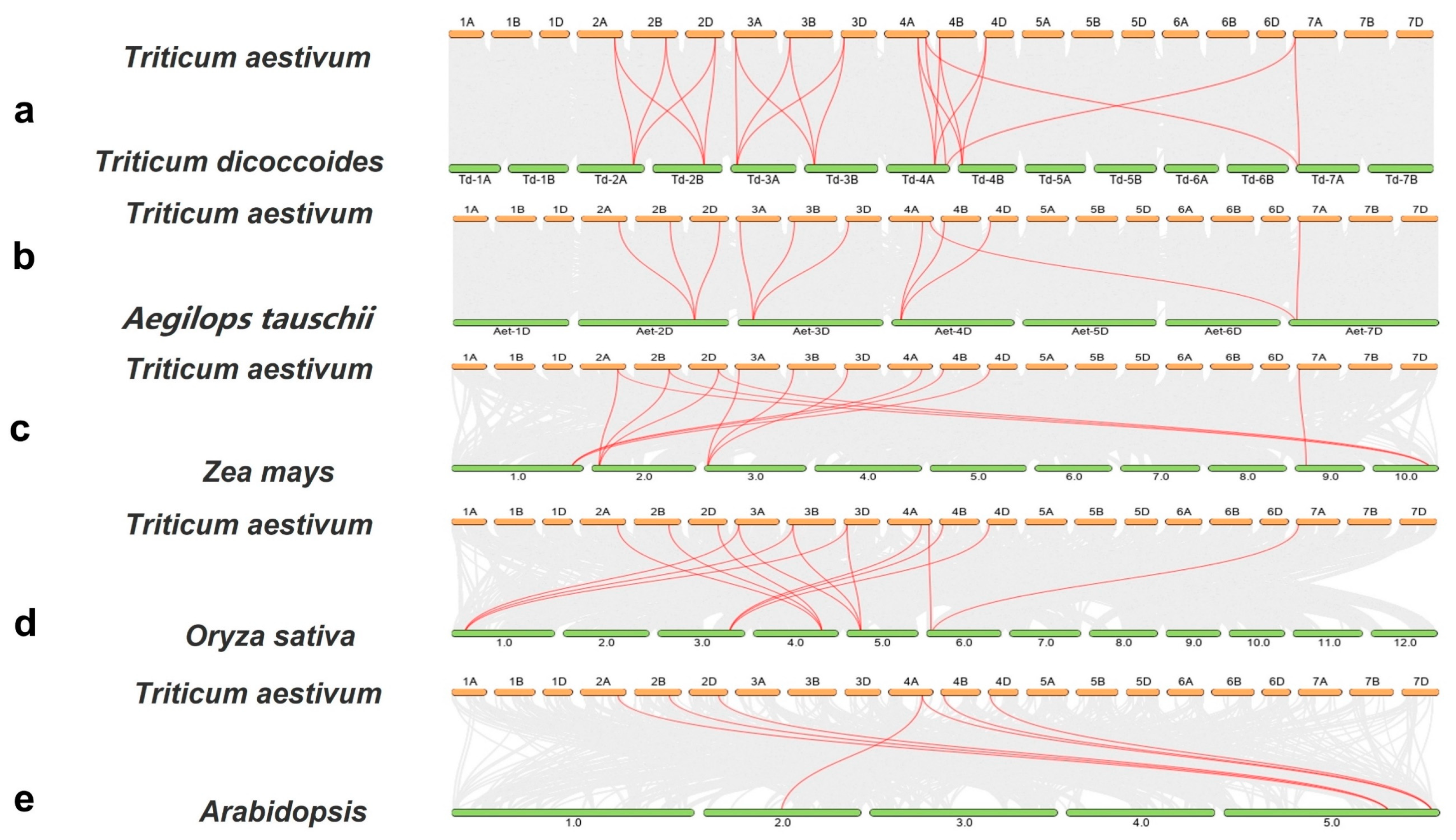
2.4. Chromosome Distribution and Collinearity Analysis of ACS Genes in Wheat
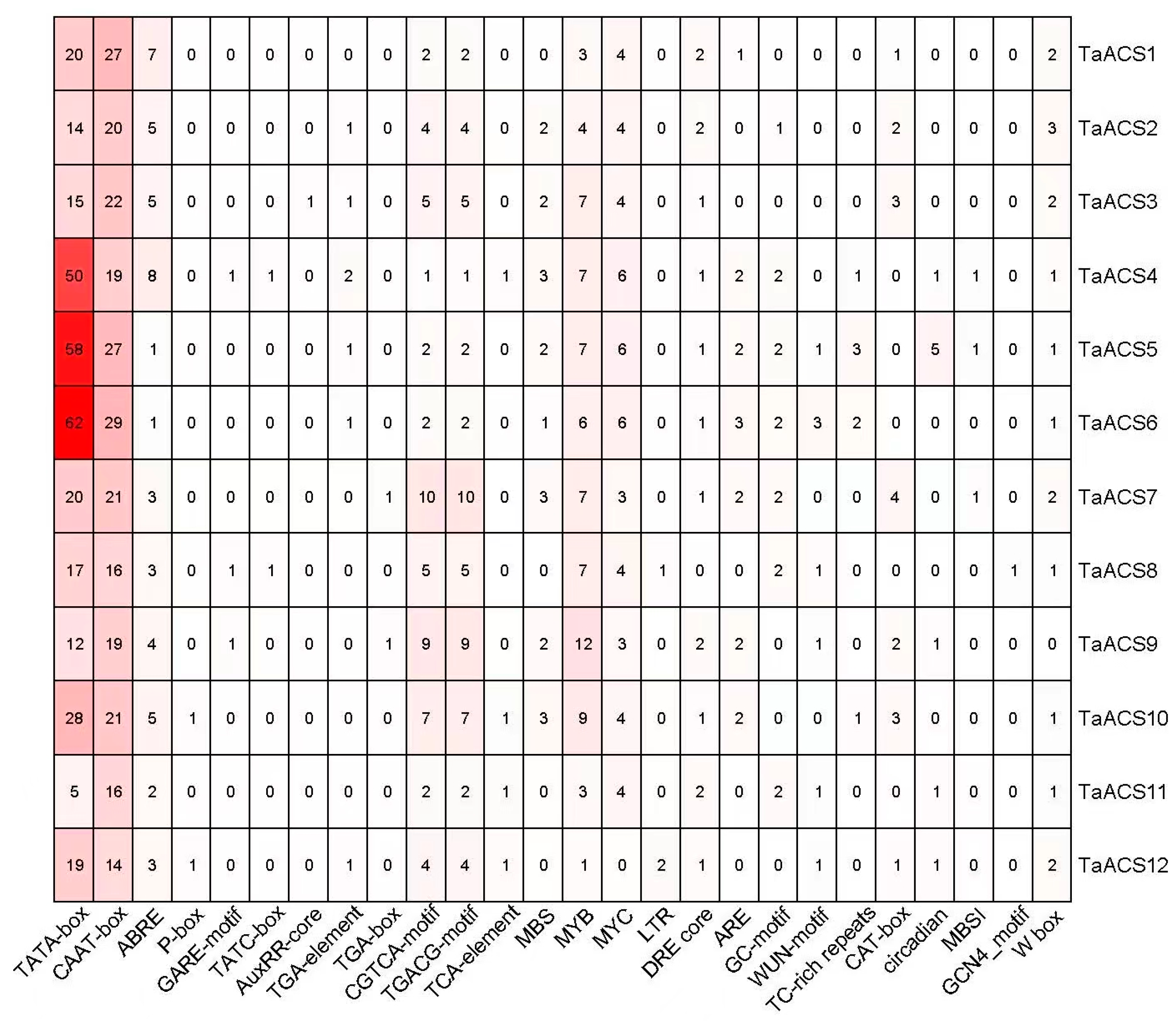
2.5. Analysis of Cis Elements in TaACS Gene Promoter
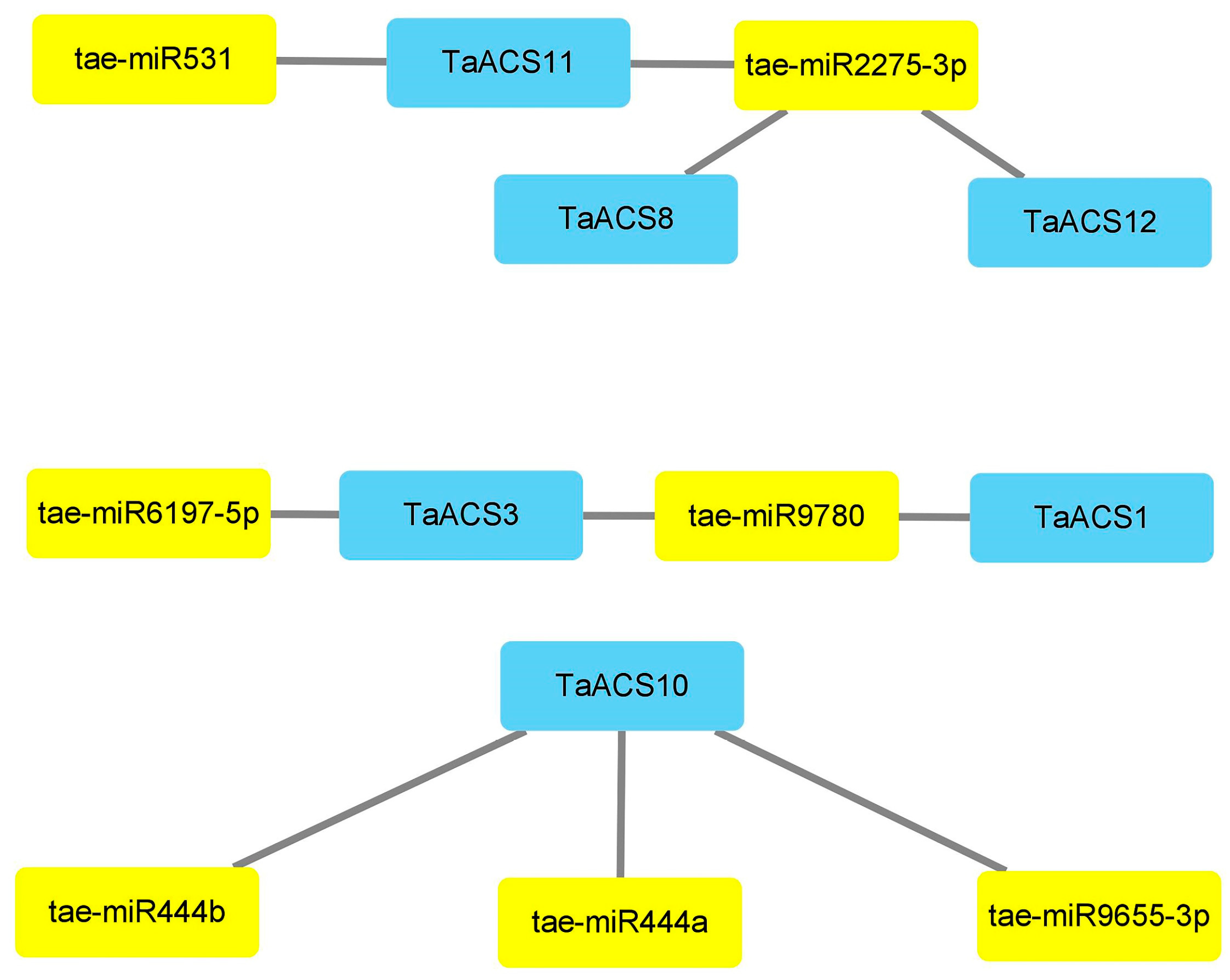
2.6. Analysis of miRNA Targeting TaACS Gene
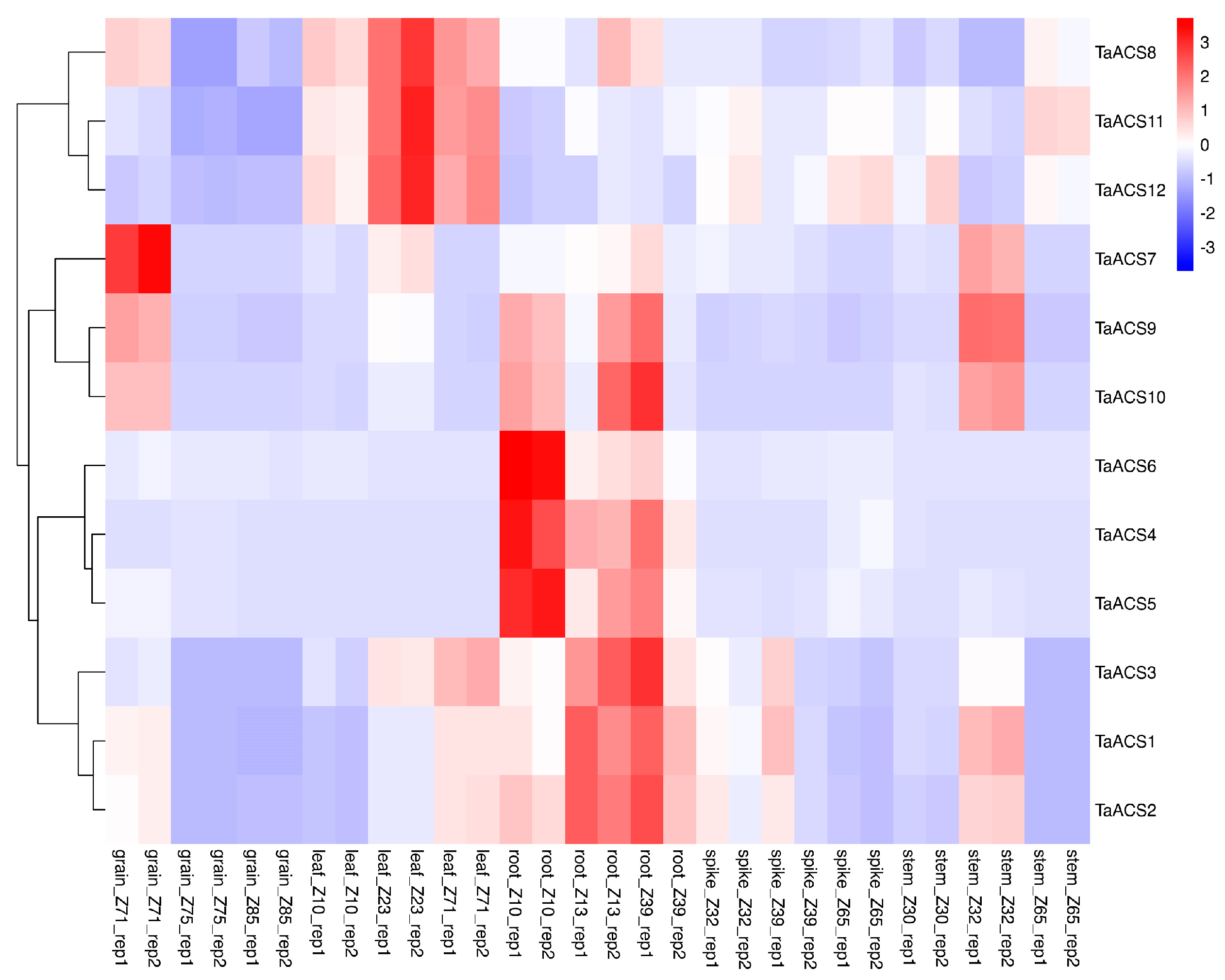
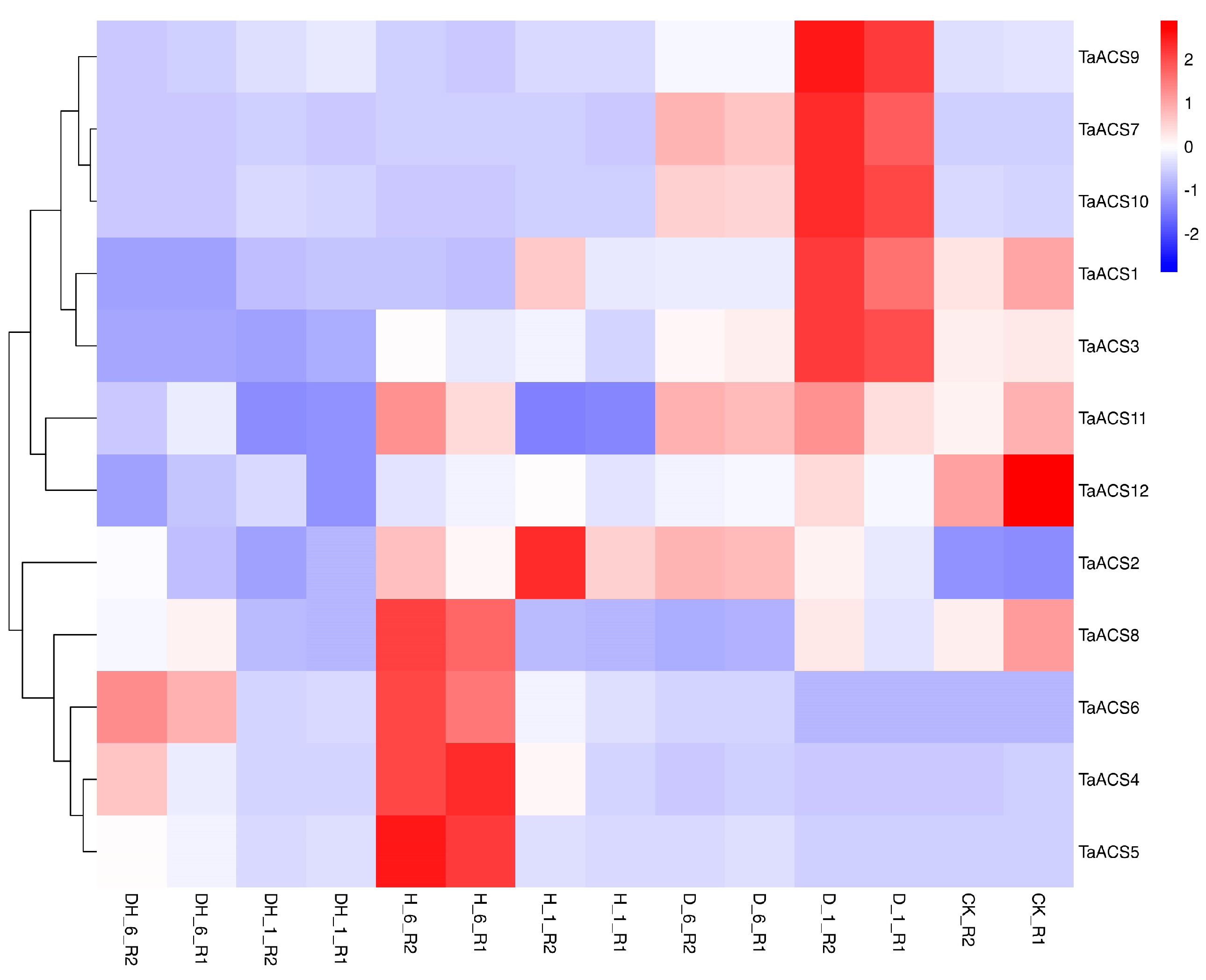
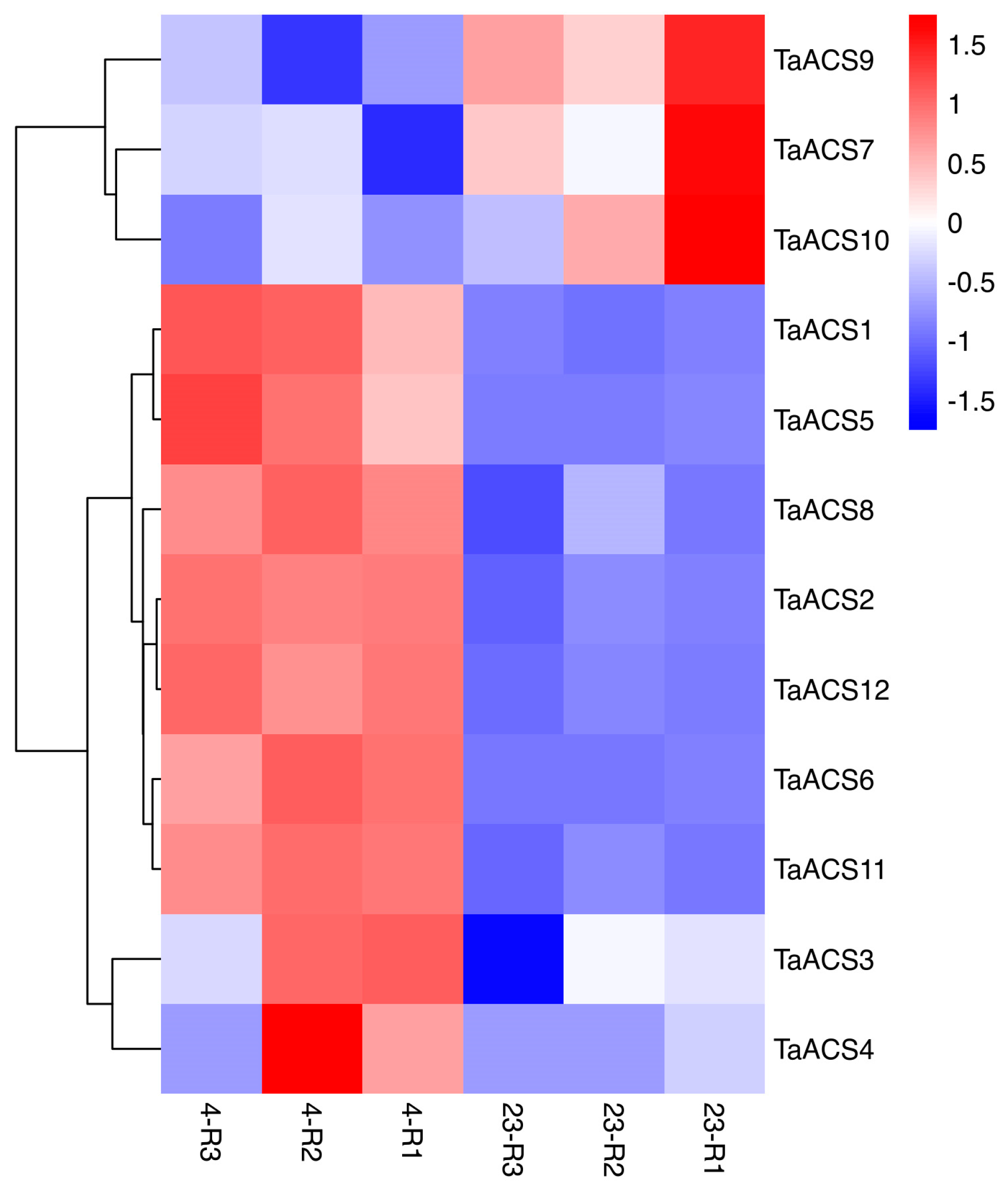
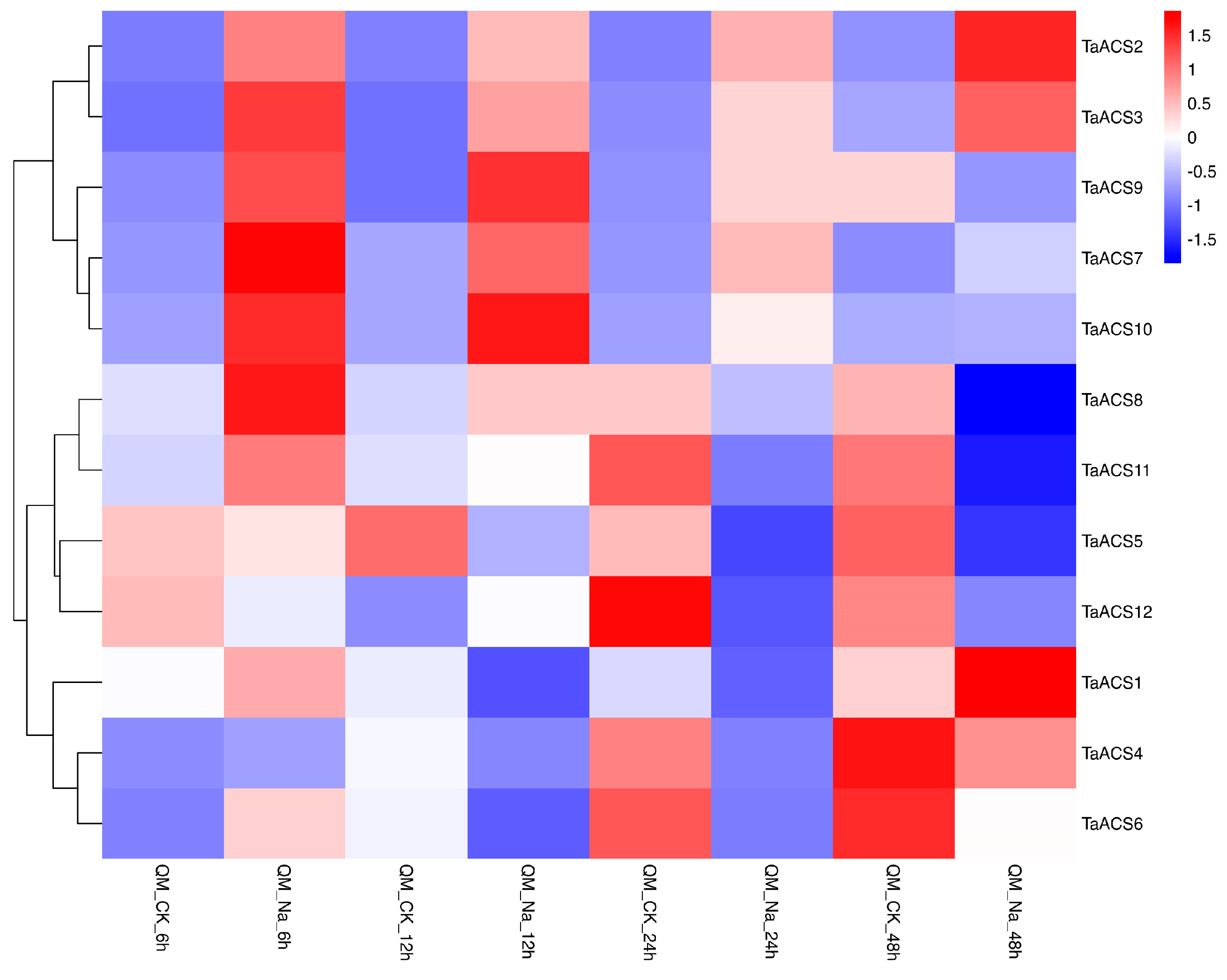
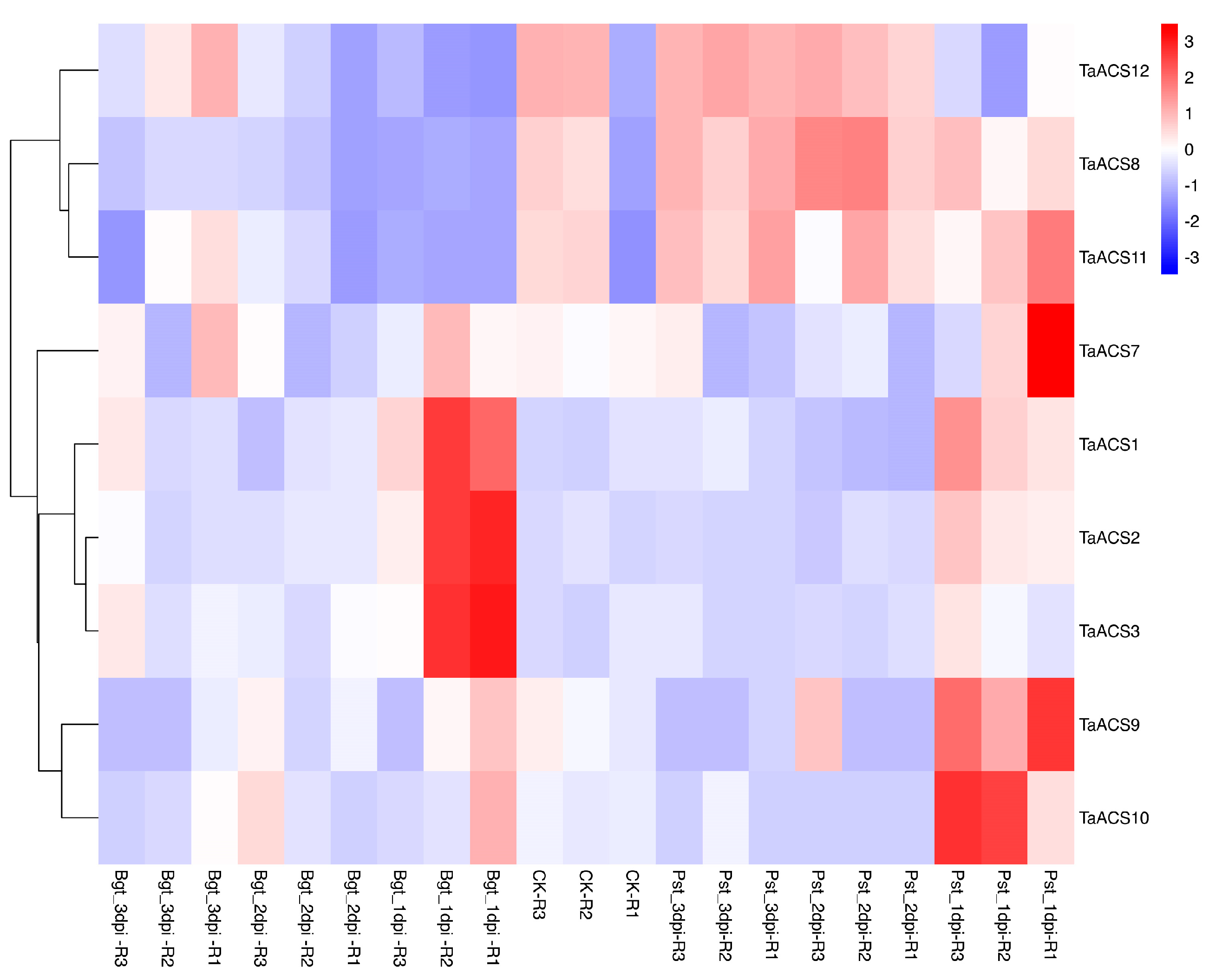
2.7. Expression Profile of ACS Gene in Wheat
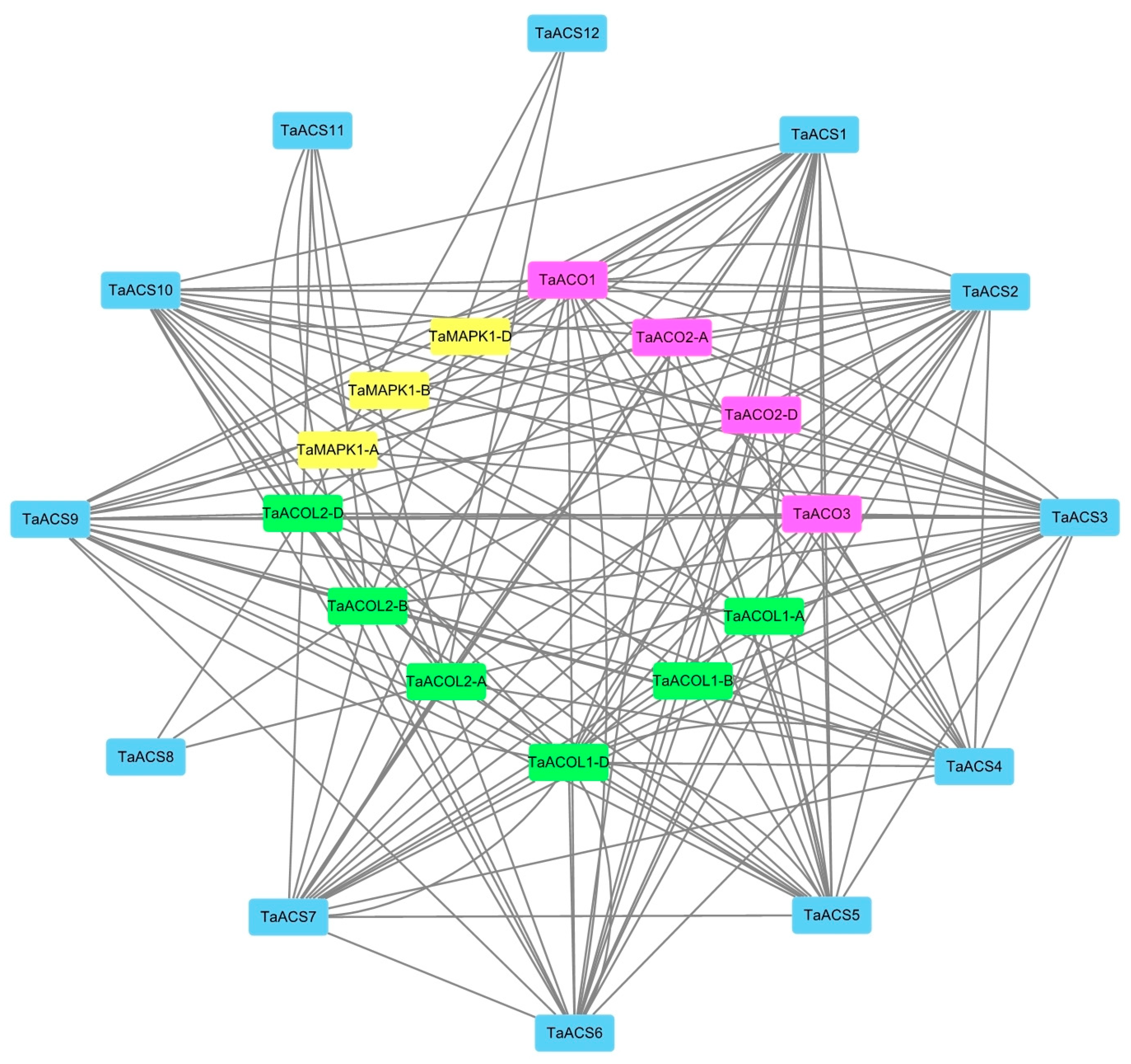
2.8. Analysis of Wheat ACS Protein Interaction Network
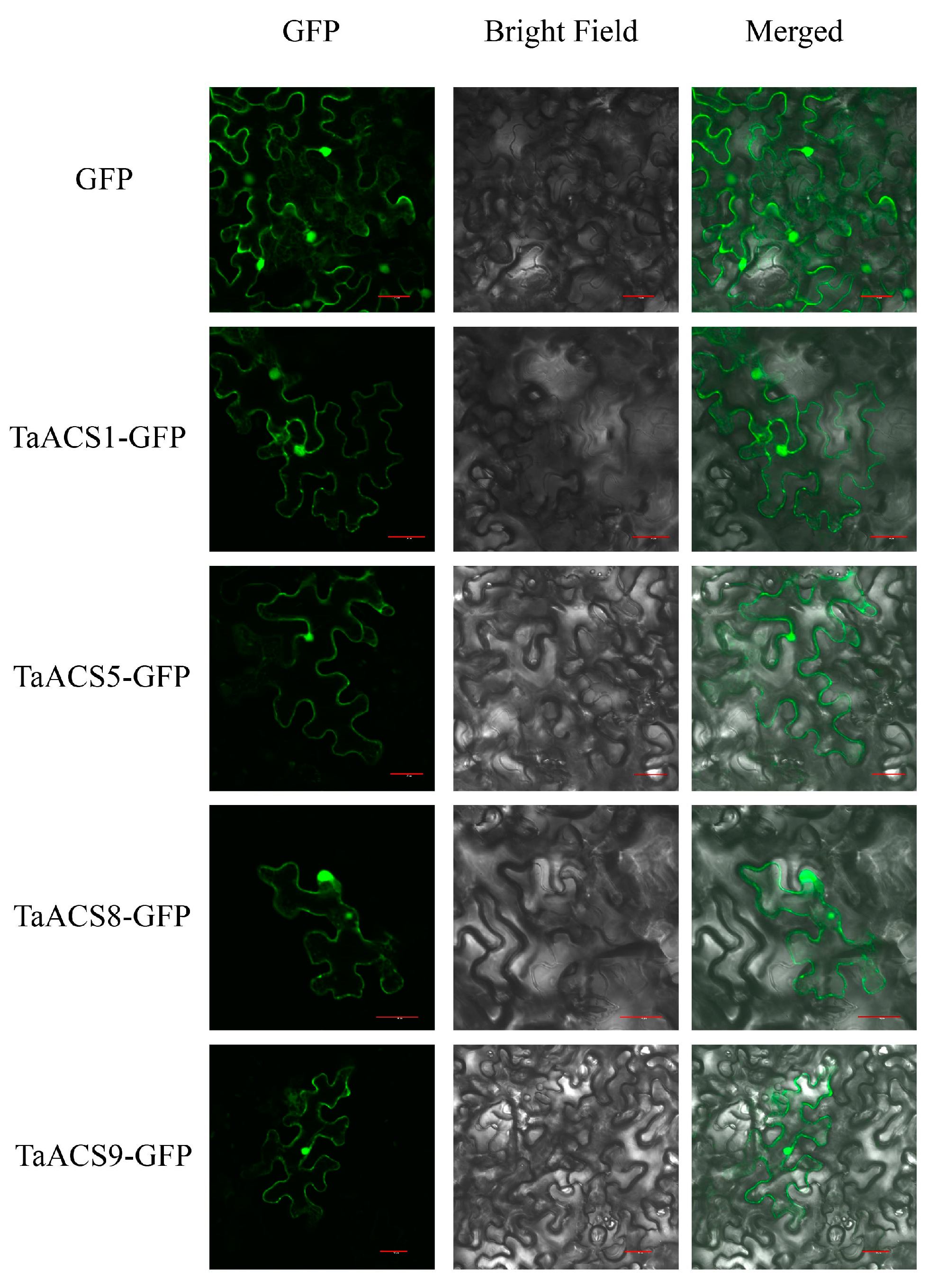
2.9. Subcellular Localization Analysis of TaACS Genes
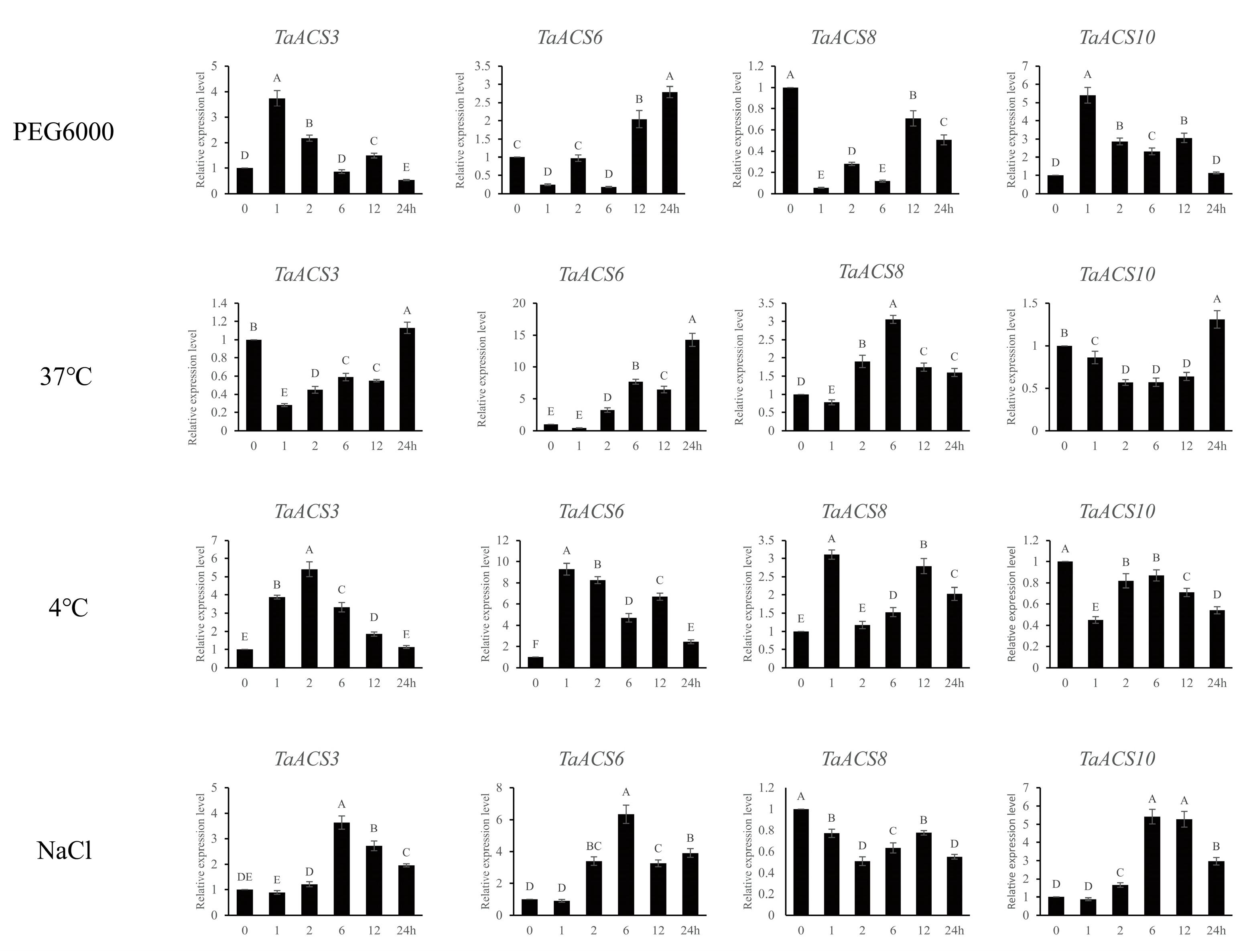
2.10. Validations of the Expressions of TaACS Genes Using RT-qPCR Analyses
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of TaACS Genes
4.2. Phylogenetic Analysis
4.3. Gene Structure and Conserved Motif Analysis of the TaACS Gene Family
4.4. Analysis of Chromosomal Localization and Gene Duplication Events in the TaACS Gene Family
4.5. Analysis of Cis-Regulatory Elements in Promoters
4.6. Prediction of miRNA Target Site of TaACS
4.7. Expression Pattern Analysis
4.8. Protein Interaction Network Analysis of TaACS
4.9. Subcellular Localization
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thao, N.; Khan, M.; Thu, N.; Hoang, X.; Asgher, D.M.; Khan, N.; Tran, L.-S. Role of Ethylene and Its Cross Talk with Other Signaling Molecules in Plant Responses to Heavy Metal Stress. Plant Physiol. 2015, 169, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Kamiyoshihara, Y.; Iwata, M.; Fukaya, T.; Tatsuki, M.; Mori, H. Turnover of LeACS2, a Wound-Inducible 1-Aminocyclopropane-1-Carboxylic Acid Synthase in Tomato, Is Regulated by Phosphorylation/Dephosphorylation. Plant J. 2010, 64, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Marhavy, P.; Kurenda, A.; Siddique, S.; Denervaud Tendon, V.; Zhou, F.; Holbein, J.; Hasan, S.; Grundler, F.; Farmer, E.; Geldner, N. Single-cell Damage Elicits Regional, Nematode-restricting Ethylene Responses in Roots. EMBO J. 2019, 38, e100972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waadt, R.; Seller, C.; Hsu, P.-K.; Takahashi, Y.; Munemasa, S.; Schroeder, J. Plant Hormone Regulation of Abiotic Stress Responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Mattoo, A.; Goyal, R. Multitasking Antimicrobial Peptides in Plant Development and Host Defense against Biotic/Abiotic Stres. Plant Sci. 2014, 228, 135–149. [Google Scholar]
- Trusov, Y.; Botella, J. Silencing of the ACC Synthase Gene ACACS2 Causes Delayed Flowering in Pineapple [Ananas Comosus (L.) Merr.]. J. Exp. Bot. 2006, 57, 3953–3960. [Google Scholar] [CrossRef] [Green Version]
- Frankowski, K.; Kesy, J.; Wojciechowski, W.; Kopcewicz, J. Light- and IAA-Regulated ACC Synthase Gene (PnACS) from Pharbitis Nil and Its Possible Role in IAA-Mediated Flower Inhibition. J. Plant Physiol. 2008, 166, 192–202. [Google Scholar] [CrossRef]
- Tatsuki, M.; Haji, T.; Yamaguchi, M. The Involvement of 1-Aminocyclopropane-1-Carboxylic Acid Synthase Isogene, Pp-ACS1, in Peach Fruit Softening. J. Exp. Bot. 2006, 57, 1281–1289. [Google Scholar] [CrossRef]
- Fenn, M.; Giovannoni, J. Phytohormones in Fruit Development and Maturation. Plant J. 2020, 105, 446–458. [Google Scholar] [CrossRef]
- Numata, T.; Sugita, K.; Rahman, A.A.; Rahman, A. Actin Isovariant ACT7 Controls Root Meristem Development in Arabidopsis through Modulating Auxin and Ethylene Responses. J. Exp. Bot. 2022, 73, 6255–6271. [Google Scholar] [CrossRef]
- Schneider, H.; Wojciechowski, T.; Postma, J.; Brown, K.; Lynch, J. Ethylene Modulates Root Cortical Senescence in Barley. Ann. Bot. 2018, 122, 95–105. [Google Scholar] [CrossRef] [Green Version]
- Parankusam, S.; Nongmaithem, S.; Sreelakshmi, Y.; Ivanchenko, M.; Sharma, R. The Root as a Drill: An Ethylene-Auxin Interaction Facilitates Root Penetration in Soil. Plant Signal. Behav. 2012, 7, 151–156. [Google Scholar]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The Regulation of Ethylene Biosynthesis: A Complex Multilevel Control Circuitry. New Phytol. 2020, 229, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O.; Yang, S.-F. Ethylene Biosynthesis: Identification of 1-Aminocyclopropane-1-Carboxylic Acid as an Intermediate in the Conversion of Methionine to Ethylene. Proc. Natl. Acad. Sci. USA 1979, 76, 170–174. [Google Scholar] [CrossRef]
- DeLong, A.; Booker, M. Producing the Ethylene Signal: Regulation and Diversification of Ethylene Biosynthetic Enzymes. Plant Physiol. 2015, 169, 42–50. [Google Scholar]
- Dong, J.; Kim, W.T.; Yip, W.; Thompson, G.; Li, L.; Bennett, A.; Yang, S.-F. Cloning of a CDNA Encoding 1-Aminocyclopropane-1-Carboxylate Synthase and Expression of Its MRNA in Ripening Apple Fruit. Planta 1991, 185, 38–45. [Google Scholar] [CrossRef]
- Rottmann, W.; Peter, G.; Oeller, P.; Keller, J.; Shen, N.; Nagy, B.; Taylor, L.; Campbell, A.; Theologis, A. 1-Aminocyclopropane-1-Carboxylate Synthase in Tomato Is Encoded by a Multigene Family Whose Transcription Is Induced during Fruit and Floral Senescence. J. Mol. Biol. 1992, 222, 937–961. [Google Scholar] [CrossRef]
- Jakubowicz, M. Structure, Catalytic Activity and Evolutionary Relationships of 1-Aminocyclopropane-1-Carboxylate Synthase, the Key Enzyme of Ethylene Synthesis in Higher Plants. Acta Biochim. Pol. 2002, 49, 757–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamagami, T.; Tsuchisaka, A.; Yamada, K.; Haddon, W.F.; Harden, L.A.; Theologis, A. Biochemical Diversity among the 1-Amino-Cyclopropane-1-Carboxylate Synthase Isozymes Encoded by the Arabidopsis Gene Family. J. Biol. Chem. 2003, 278, 49102–49112. [Google Scholar] [CrossRef] [Green Version]
- Barry, C.S.; Llop-Tous, M.I.; Grierson, D. The Regulation of 1-Aminocyclopropane-1-Carboxylic Acid Synthase Gene Expression during the Transition from System-1 to System-2 Ethylene Synthesis in Tomato1. Society 2000, 123, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.H.; Xu, M. Genome-Wide Analysis of 1-Amino-Cyclopropane-1-Carboxylate Synthase Gene Family in Arabidopsis, Rice, Grapevine and Poplar. Afr. J. Biotechnol. 2012, 11, 1106–1118. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Chen, F.; Cheng, Y.; Huang, X.; Zou, B.; Wang, Y.; Xu, W.; Qu, S. Genome-Wide Identification and Characterization of Members of the ACS Gene Family in Cucurbita Maxima and Their Transcriptional Responses to the Specific Treatments. Int. J. Mol. Sci. 2022, 23, 8476. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Lee, H.Y.; Yoon, G.M. The Regulation of ACC Synthase Protein Turnover: A Rapid Route for Modulating Plant Development and Stress Responses. Curr. Opin. Plant Biol. 2021, 63, 102046. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, R.; Zheng, M.; Liu, X.; Meng, F.; Wu, H.; Yao, Y.; Xin, M.; Peng, H.; Ni, Z.; et al. TaWRKY51 Promotes Lateral Root Formation through Negative Regulation of Ethylene Biosynthesis in Wheat (Triticum Aestivum L.). Plant J. 2018, 96, 372–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Meng, X.; Wang, R.; Mao, G.; Han, L.; Liu, Y.; Zhang, S. Dual-Level Regulation of ACC Synthase Activity by MPK3/MPK6 Cascade and Its Downstream WRKY Transcription Factor during Ethylene Induction in Arabidopsis. PLoS Genet. 2012, 8, e1002767. [Google Scholar] [CrossRef]
- Arteca, J.; Arteca, R. A Multi-Responsive Gene Encoding 1-Aminocyclopropane-1-Carboxylate Synthase (ACS6) in Mature Arabidopsis Leaves. Plant Mol. Biol. 1999, 39, 209–219. [Google Scholar] [CrossRef]
- Young, T.; Meeley, R.; Gallie, D. ACC Synthase Expression Regulates Leaf Performance and Drought Tolerance in Maize. Plant J. 2005, 40, 813–825. [Google Scholar] [CrossRef]
- Zhou, Z.; Vriezen, W.; Van Caeneghem, W.; Van Montagu, M.; Van Der Straeten, D. Rapid Induction of a Novel ACC Synthase Gene in Deepwater Rice Seedlings upon Complete Submergence. Euphytica 2001, 121, 137–143. [Google Scholar] [CrossRef]
- Wi, S.J.; Park, K.Y. Antisense Expression of Carnation CDNA Encoding ACC Synthase or ACC Oxidase Enhances Polyamine Content and Abiotic Stress Tolerance in Transgenic Tobacco Plants. Mol. Cells 2002, 13, 209–220. [Google Scholar]
- Boualem, A.; Troadec, C.; Kovalski, I.; Sari, M.-A.; Perl-Treves, R.; Bendahmane, A. A Conserved Ethylene Biosynthesis Enzyme Leads to Andromonoecy in Two Cucumis Species. PLoS ONE 2009, 4, e6144. [Google Scholar] [CrossRef]
- Manzano, S.; Aguado, E.; Martínez, C.; Megías, Z.; García Fuentes, A.; Jamilena, M. The Ethylene Biosynthesis Gene CitACS4 Regulates Monoecy/Andromonoecy in Watermelon (Citrullus Lanatus). PLoS ONE 2016, 11, e0154362. [Google Scholar] [CrossRef] [Green Version]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N. Current Understanding on Ethylene Signaling in Plants: The Influence of Nutrient Availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Atkinson, M.; Chinoy, C.N.; Devos, K.M.; Gale, M. Nonhomoeologous Translocations between Group 4, 5 and 7 Chromosomes within Wheat and Rye. Theor. Appl. Genet. 1992, 83, 305–312. [Google Scholar] [CrossRef] [PubMed]
- King, I.; Purdie, K.; Liu, C.; Reader, S.; Pittaway, T.; Orford, S.; Miller, T. Detection of Interchromosomal Translocations within the Triticeae by RFLP Analysis. Genome 1994, 37, 882–887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, L.; Yun, L.; Ji, L.; Li, G.; Ji, M.; Shi, Y.; Zheng, X. Catalase (CAT) Gene Family in Wheat (Triticum Aestivum L.): Evolution, Expression Pattern and Function Analysis. Int. J. Mol. Sci. 2022, 23, 542. [Google Scholar] [CrossRef]
- Levy, A.; Feldman, M. Evolution and Origin of Bread Wheat. Plant Cell 2022, 34, 2549–2567. [Google Scholar] [CrossRef]
- Adeleke, D.; Millas, R.; McNeal, W.; Faris, J.; Taheri, A. Variation Analysis of Root System Development in Wheat Seedlings Using Root Phenotyping System. Agronomy 2020, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Iwasa, T.; Shinkawa, T.; Kawahara, A.; Kurusu, T.; Inoue, Y. Isolation and Characterization of the ACC Synthase Genes from Lettuce (Lactuca Sativa L.), and the Involvement in Low PH-Induced Root Hair Initiation. Plant Cell Physiol. 2003, 44, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Sarwar, R.; Jiang, T.; Ding, P.; Gao, Y.; Tan, X.-L.; Zhu, K. Genome-Wide Analysis and Functional Characterization of the DELLA Gene Family Associated with Stress Tolerance in B. Napus. BMC Plant Biol. 2021, 21, 286. [Google Scholar] [CrossRef]
- Jeong, J.; Jung, W.J.; Seo, Y. Genome-Wide Identification and Expression Analysis of the Annexin Gene Family in Rye (Secale Cereale L.). Gene 2022, 838, 146704. [Google Scholar] [CrossRef]
- Wang, N.-N.; Shih, M.-H.; Li, N. The GUS Reporter-Aided Analysis of the Promoter Activities of Arabidopsis ACC Synthase Genes AtACS4, AtACS5, and AtACS7 Induced by Hormones and Stresses. J. Exp. Bot. 2005, 56, 909–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.; Song, T.; Zhou, H.; Zhang, M.; Li, N.; Xiang, J.; Zhang, X. Genome-Wide Identification, Characterization, Evolution, and Expression Pattern Analyses of the Typical Thioredoxin Gene Family in Wheat (Triticum Aestivum L.). Front. Plant Sci. 2022, 13, 1020584. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.Q.; Wang, Z.Z.; Yang, L.T.; Wei, Y.T.; Li, Y. Research Progress of Two Key Enzyme Genes on the Ethylene Biosynthesis. J. Guangxi Agric. Biol. Sci. 2004, 23, 164–169. [Google Scholar]
- Zhao, Y.; Song, C.; Brummell, D.; Qi, S.; Lin, Q.; Duan, Y. Jasmonic Acid Treatment Alleviates Chilling Injury in Peach Fruit by Promoting Sugar and Ethylene Metabolism. Food Chem. 2020, 338, 128005. [Google Scholar] [CrossRef] [PubMed]
- Van Der Straeten, D.; Anuntalabhochai, S.; Van Caeneghem, W.; Zhou, Z.; Gielen, J.; Van Montagu, M. Expression of Three Members of the ACC Synthase Gene Family in Deepwater Rice by Submergence, Wounding and Hormonal Treatments. Plant Sci.–PLANT SCI 1997, 124, 79–87. [Google Scholar] [CrossRef]
- Sousa, M.; Dolicka, D.; Gjorgjieva, M.; Sobolewski, C.; Foti, M. Deciphering MiRNAs’ Action through MiRNA Editing. Int. J. Mol. Sci. 2019, 20, 6249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach, S.; Kretz, M. Non-Coding RNAs: Classification, Biology and Functioning. In Advances in Experimental Medicine and Biology; Springer: Heidelberg, Germany, 2016; pp. 3–17. [Google Scholar]
- Kim, C.; Pak, T. MiRNA Degradation in the Mammalian Brain. Am. J. Physiol. Cell Physiol. 2020, 319, 624–629. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, H.; Hamera, S.; Chen, X.; Fang, R.-X. MiR444a Has Multiple Functions in the Rice Nitrate-Signaling Pathway. Plant J. 2014, 78, 44–55. [Google Scholar] [CrossRef]
- Jin, X.; Jia, L.; Wang, Y.; Li, B.; Sun, D.; Chen, X. Identification of Fusarium Graminearum-Responsive MiRNAs and Their Targets in Wheat by SRNA Sequencing and Degradome Analysis. Funct. Integr. Genom. 2020, 20, 51–61. [Google Scholar] [CrossRef]
- Yue, H.; Zhang, H.; Su, N.; Sun, X.; Zhao, Q.; Weining, S.; Nie, X.; Yue, W. Integrate Small RNA and Degradome Sequencing to Reveal Drought Memory Response in Wheat (Triticum Aestivum L.). Int. J. Mol. Sci. 2022, 23, 5917. [Google Scholar] [CrossRef]
- Sun, L.; Sun, G.; Shi, C.; Sun, D. Transcriptome Analysis Reveals New MicroRNAs-Mediated Pathway Involved in Anther Development in Male Sterile Wheat. BMC Genom. 2018, 19, 333. [Google Scholar] [CrossRef] [Green Version]
- Tariq, R.; Hussain, A.; Tariq, A.; Khalid, M.H.; Khan, I.; Basim, H.; Ingvarsson, P. Genome-Wide Analyses of the Mung Bean NAC Gene Family Reveals Orthologs, Co-Expression Networking and Expression Profiling under Abiotic and Biotic Stresses. BMC Plant Biol. 2022, 22, 343. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Wang, L.; Gao, J. Bioinformatics of 1-Aminocyclopropane-1-Carboxylic Acid Synthase Gene from Eggplant and Its Expression Analysis in Response to Adversity Stresses. J. Zhejiang Univ. 2021, 47, 325–334. [Google Scholar]
- Wang, X.; Meng, H.; Tang, Y.; Zhang, Y.; He, Y.; Zhou, J.; Meng, X. Phosphorylation of an Ethylene Response Factor by MPK3/MPK6 Mediates Negative Feedback Regulation of Pathogen-Induced Ethylene Biosynthesis in Arabidopsis. J. Genet. Genom. 2022, 49, 810–822. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ka | Ks | Ka/Ks | ||
|---|---|---|---|---|
| TaACS1 | TaACS2 | 0.007273 | 0.06513 | 0.111666 |
| TaACS1 | TaACS3 | 0.006363 | 0.071235 | 0.08932 |
| TaACS2 | TaACS3 | 0.005905 | 0.057409 | 0.102852 |
| TaACS4 | TaACS5 | 0.008461 | 0.103747 | 0.081557 |
| TaACS4 | TaACS6 | 0.004963 | 0.063434 | 0.078242 |
| TaACS5 | TaACS6 | 0.007459 | 0.064963 | 0.114821 |
| TaACS7 | TaACS9 | 0.014696 | 0.055432 | 0.26512 |
| TaACS7 | TaACS10 | 0.007325 | 0.06421 | 0.114077 |
| TaACS8 | TaACS11 | 0.018778 | 0.097469 | 0.192654 |
| TaACS8 | TaACS12 | 0.011384 | 0.096819 | 0.117577 |
| TaACS9 | TaACS10 | 0.01471 | 0.073606 | 0.199843 |
| TaACS11 | TaACS12 | 0.008741 | 0.093894 | 0.093091 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Lei, C.; Zhu, Z.; Li, M.; Chen, Z.; He, W.; Liu, B.; Chen, L.; Li, X.; Xie, Y. Genome-Wide Analysis and Identification of 1-Aminocyclopropane-1-Carboxylate Synthase (ACS) Gene Family in Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2023, 24, 11158. https://doi.org/10.3390/ijms241311158
Liu S, Lei C, Zhu Z, Li M, Chen Z, He W, Liu B, Chen L, Li X, Xie Y. Genome-Wide Analysis and Identification of 1-Aminocyclopropane-1-Carboxylate Synthase (ACS) Gene Family in Wheat (Triticum aestivum L.). International Journal of Molecular Sciences. 2023; 24(13):11158. https://doi.org/10.3390/ijms241311158
Chicago/Turabian StyleLiu, Shuqing, Chao Lei, Zhanhua Zhu, Mingzhen Li, Zhaopeng Chen, Wei He, Bin Liu, Liuping Chen, Xuejun Li, and Yanzhou Xie. 2023. "Genome-Wide Analysis and Identification of 1-Aminocyclopropane-1-Carboxylate Synthase (ACS) Gene Family in Wheat (Triticum aestivum L.)" International Journal of Molecular Sciences 24, no. 13: 11158. https://doi.org/10.3390/ijms241311158