
Increased Interleukin-36β Expression Promotes Angiogenesis in Japanese Atopic Dermatitis
 ,
,
Abstract
:1. Introduction
2. Results
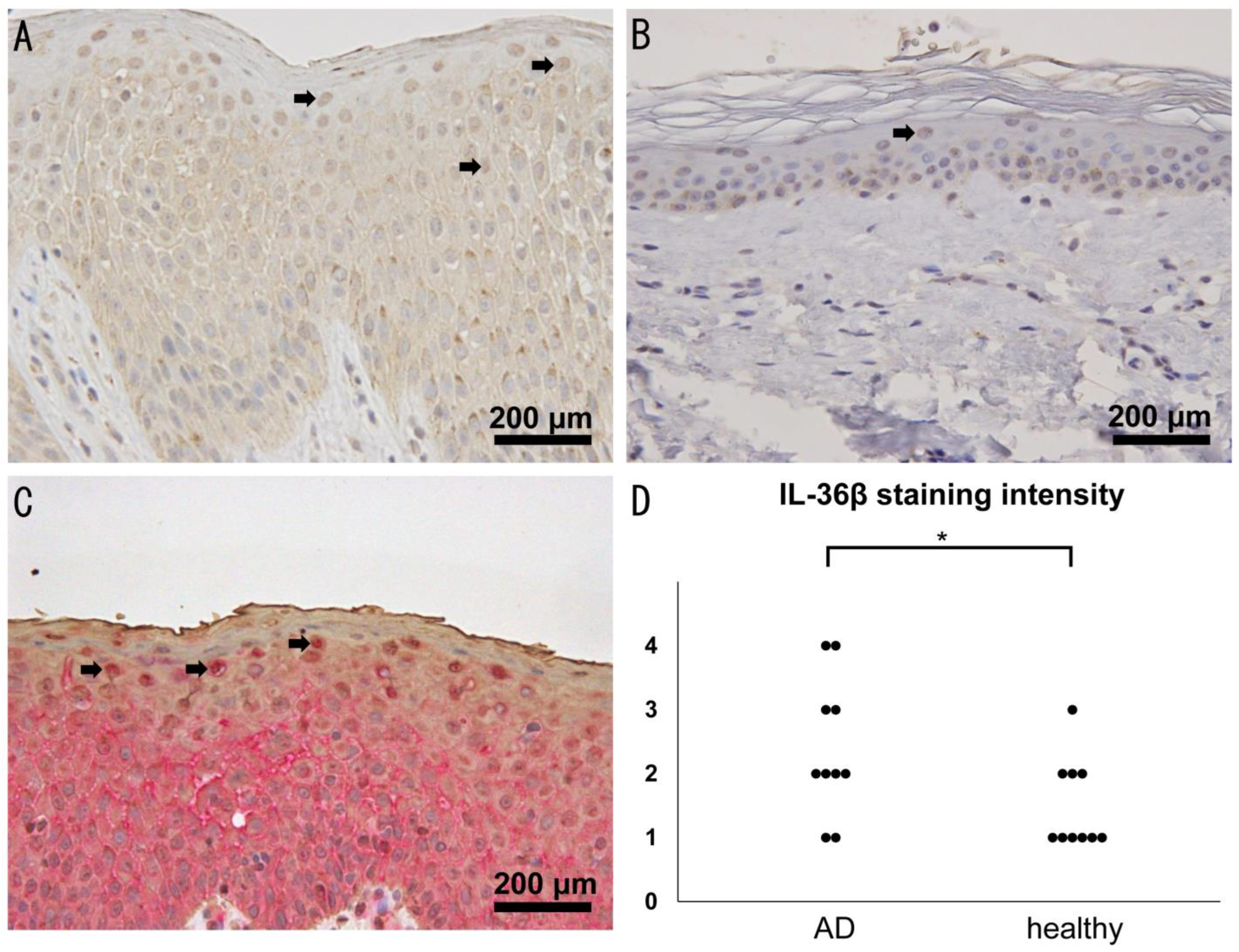
2.1. Increased Expression of IL-36β in Lesional Skin of AD
2.2. Serum IL-36β Levels Are Not Elevated in Patients with AD but Decreased by Dupilumab
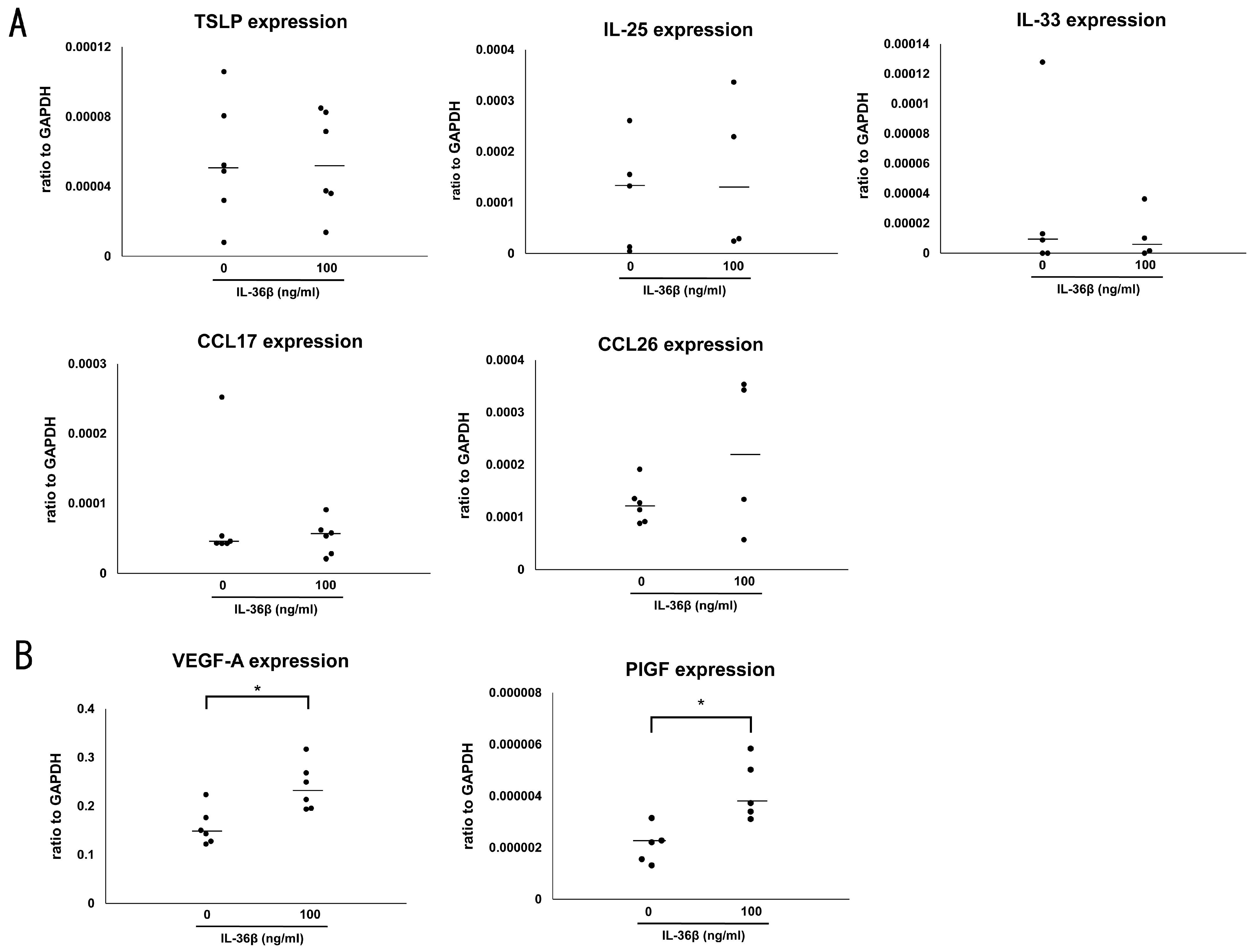
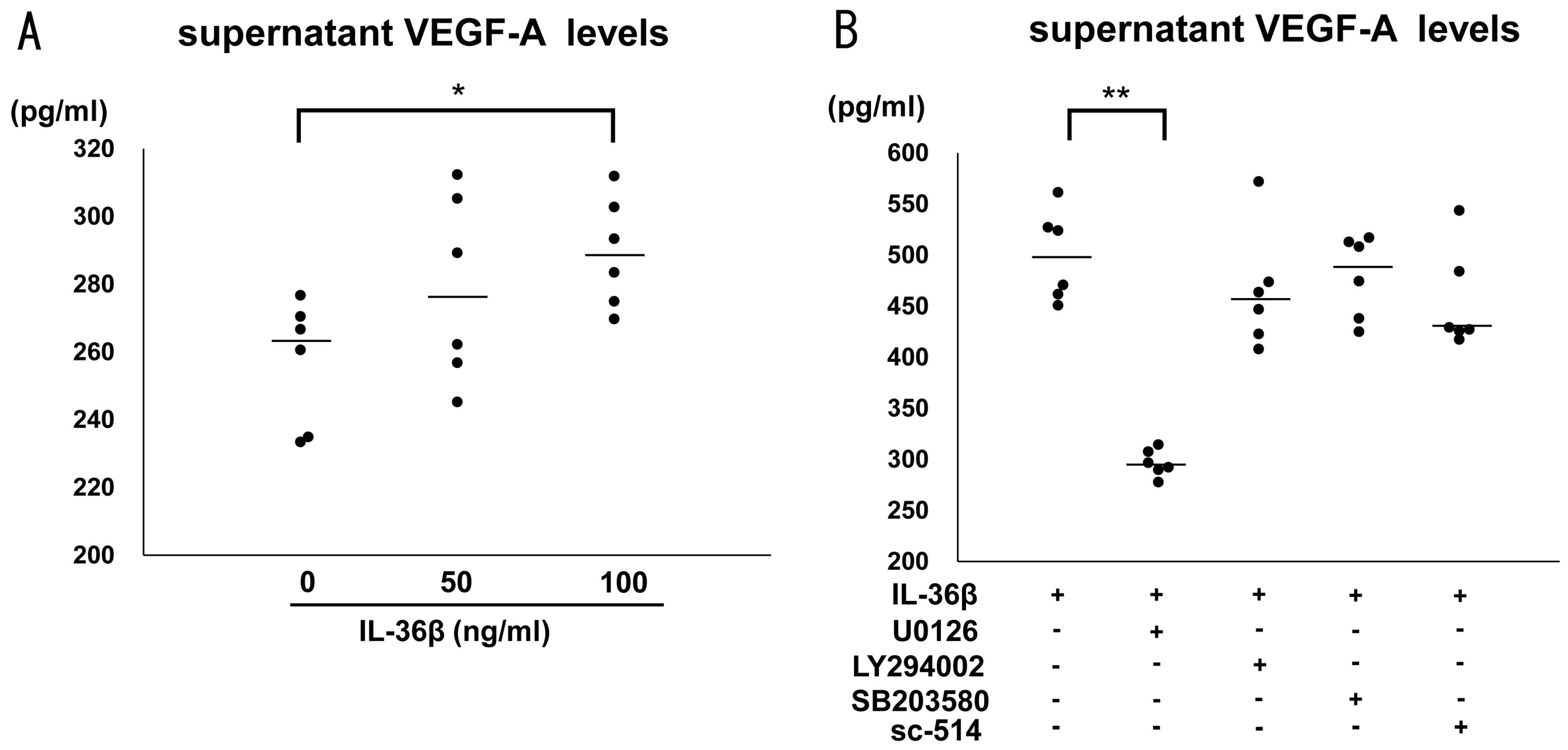
2.3. IL-36β Promotes VEGF-A and PlGF Expression in HaCaT Cells
2.4. IL-36β Enhanced VEGF-A Expression via ERK1/2 Pathway in HaCaT Cells
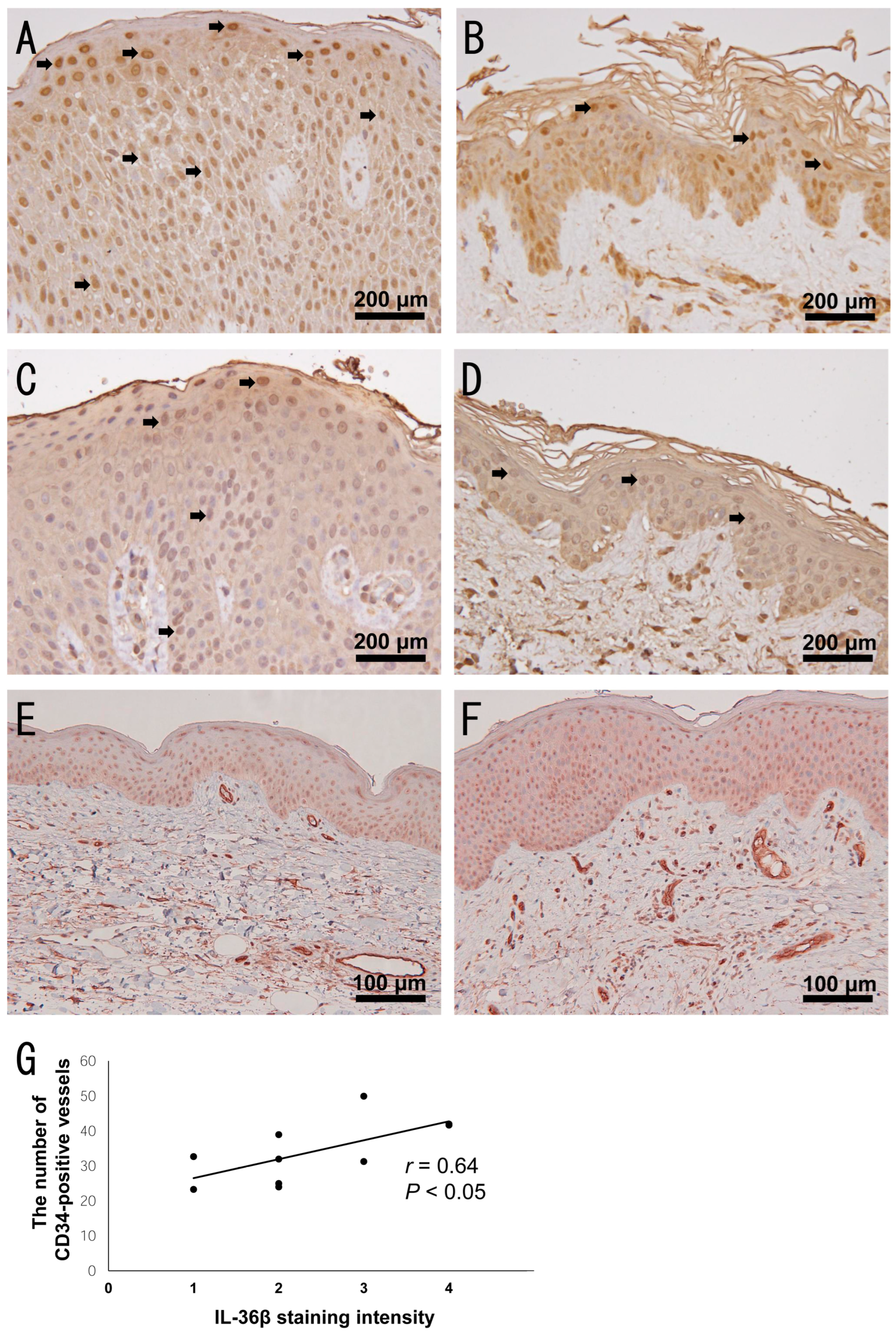
2.5. Increased Dermal Vessels in Highly IL-36β-Expressed AD Skin
3. Discussion
4. Materials and Methods
4.1. Patients and Tissue Samples
4.2. Cell Line
4.3. RNA Isolation and Quantitative Reverse Transcription-PCR
4.4. Immunohistochemistry
4.5. Enzyme-Linked Immunosorbent Assay
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bieber, T. Atopic Dermatitis. N. Engl. J. Med. 2008, 358, 1483–1494. [Google Scholar] [CrossRef]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- Paller, A.S.; Kabashima, K.; Bieber, T. Therapeutic pipeline for atopic dermatitis: End of the drought? J. Allergy Clin. Immunol. 2017, 140, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Oetjen, L.K.; Mack, M.R.; Feng, J.; Whelan, T.M.; Niu, H.; Guo, C.J.; Chen, S.; Trier, A.M.; Xu, A.Z.; Tripathi, S.V.; et al. Sensory Neurons Co-opt Classical Immune Signaling Pathways to Mediate Chronic Itch. Cell 2017, 171, 217–228.e13. [Google Scholar] [CrossRef] [Green Version]
- Howell, M.D.; Fairchild, H.R.; Kim, B.E.; Bin, L.; Boguniewicz, M.; Redzic, J.S.; Hansen, K.C.; Leung, D.Y.M. Th2 cytokines act on S100/A11 to downregulate keratinocyte differentiation. J. Investig. Dermatol. 2008, 128, 2248–2258. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.L.; Bieber, T.; Guttman-Yassky, E.; Beck, L.A.; Blauvelt, A.; Cork, M.J.; Silverberg, J.I.; Deleuran, M.; Kataoka, Y.; Lacour, J.P.; et al. Two Phase 3 Trials of Dupilumab versus Placebo in Atopic Dermatitis. N. Engl. J. Med. 2016, 375, 2335–2348. [Google Scholar] [CrossRef]
- Kamata, M.; Tada, Y. A Literature Review of Real-World Effectiveness and Safety of Dupilumab for Atopic Dermatitis. JID Innov. 2021, 1, 100042. [Google Scholar] [CrossRef]
- Silverberg, J.I.; Pinter, A.; Pulka, G.; Poulin, Y.; Bouaziz, J.D.; Wollenberg, A.; Murrell, D.F.; Alexis, A.; Lindsey, L.; Ahmad, F.; et al. Phase 2B randomized study of nemolizumab in adults with moderate-to-severe atopic dermatitis and severe pruritus. J. Allergy Clin. Immunol. 2020, 145, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Haddad, E.B.; Cyr, S.L.; Arima, K.; McDonald, R.A.; Levit, N.A.; Nestle, F.O. Current and Emerging Strategies to Inhibit Type 2 Inflammation in Atopic Dermatitis. Dermatol. Ther. 2022, 12, 1501–1533. [Google Scholar] [CrossRef]
- Nograles, K.E.; Zaba, L.C.; Shemer, A.; Fuentes-Duculan, J.; Cardinale, I.; Kikuchi, T.; Ramon, M.; Bergman, R.; Krueger, J.G.; Guttman-Yassky, E. IL-22-producing “T22” T cells account for upregulated IL-22 in atopic dermatitis despite reduced IL-17-producing TH17 T cells. J. Allergy Clin. Immunol. 2009, 123, 1244–1252.e2. [Google Scholar] [CrossRef] [Green Version]
- Guttman-Yassky, E.; Brunner, P.M.; Neumann, A.U.; Khattri, S.; Pavel, A.B.; Malik, K.; Singer, G.K.; Baum, D.; Gilleaudeau, P.; Sullivan-Whalen, M.; et al. Efficacy and safety of fezakinumab (an IL-22 monoclonal antibody) in adults with moderate-to-severe atopic dermatitis inadequately controlled by conventional treatments: A randomized, double-blind, phase 2a trial. J. Am. Acad. Dermatol. 2018, 78, 872–881.e6. [Google Scholar] [CrossRef]
- Kamijo, H.; Miyagaki, T.; Hayashi, Y.; Akatsuka, T.; Watanabe-Otobe, S.; Oka, T.; Shishido-Takahashi, N.; Suga, H.; Sugaya, M.; Sato, S. Increased IL-26 Expression Promotes T Helper Type 17- and T Helper Type 2-Associated Cytokine Production by Keratinocytes in Atopic Dermatitis. J. Investig. Dermatol. 2020, 140, 636–644.e2. [Google Scholar] [CrossRef]
- Ding, L.; Wang, X.; Hong, X.; Lu, L.; Liu, D. IL-36 cytokines in autoimmunity and inflammatory disease. Oncotarget 2017, 9, 2895–2901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neurath, M.F. IL-36 in chronic inflammation and cancer. Cytokine Growth Factor Rev. 2020, 55, 70–79. [Google Scholar] [CrossRef]
- Sachen, K.L.; Arnold Greving, C.N.; Towne, J.E. Role of IL-36 cytokines in psoriasis and other inflammatory skin conditions. Cytokine 2022, 156, 155897. [Google Scholar] [CrossRef]
- Carrier, Y.; Ma, H.L.; Ramon, H.E.; Napierata, L.; Small, C.; O’Toole, M.; Young, D.A.; Fouser, L.A.; Nickerson-Nutter, C.; Collins, M.; et al. Inter-regulation of Th17 cytokines and the IL-36 cytokines in vitro and in vivo: Implications in psoriasis pathogenesis. J. Investig. Dermatol. 2011, 131, 2428–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercurio, L.; Morelli, M.; Scarponi, C.; Eisenmesser, E.Z.; Doti, N.; Pagnanelli, G.; Gubinelli, E.; Mazzanti, C.; Cavani, A.; Ruvo, M.; et al. IL-38 has an anti-inflammatory action in psoriasis and its expression correlates with disease severity and therapeutic response to anti-IL-17A treatment. Cell Death Dis. 2018, 9, 1104. [Google Scholar] [CrossRef] [Green Version]
- Suárez-Fariñas, M.; Ungar, B.; Correa da Rosa, J.; Ewald, D.A.; Rozenblit, M.; Gonzalez, J.; Xu, H.; Zheng, X.; Peng, X.; Estrada, Y.D.; et al. RNA sequencing atopic dermatitis transcriptome profiling provides insights into novel disease mechanisms with potential therapeutic implications. J. Allergy Clin. Immunol. 2015, 135, 1218–1227. [Google Scholar] [CrossRef]
- Martel, B.C.; Litman, T.; Hald, A.; Norsgaard, H.; Lovato, P.; Dyring-Andersen, B.; Skov, L.; Thestrup-Pedersen, K.; Skov, S.; Skak, K.; et al. Distinct molecular signatures of mild extrinsic and intrinsic atopic dermatitis. Exp. Dermatol. 2016, 25, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Otobe, S.; Sugaya, M.; Nakajima, R.; Oka, T.; Takahashi, N.; Kabasawa, M.; Miyagaki, T.; Asano, Y.; Sato, S. Increased interleukin-36γ expression in skin and sera of patients with atopic dermatitis and mycosis fungoides/Sézary syndrome. J. Dermatol. 2018, 45, 468–471. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Rodriguez, E.; Stölzl, D.; Wehkamp, U.; Sun, J.; Gerdes, S.; Sarkar, M.K.; Hübenthal, M.; Zeng, C.; Uppala, R.; et al. Progression of acute-to-chronic atopic dermatitis is associated with quantitative rather than qualitative changes in cytokine responses. J. Allergy Clin. Immunol. 2020, 145, 1406–1415. [Google Scholar] [CrossRef]
- Bissonnette, R.; Abramovits, W.; Saint-Cyr Proulx, É.; Lee, P.; Guttman-Yassky, E.; Zovko, E.; Sigmund, R.; Willcox, J.; Bieber, T. Spesolimab, an anti-interleukin-36 receptor antibody, in patients with moderate-to-severe atopic dermatitis: Results from a multicentre, randomized, double-blind, placebo-controlled, phase IIa study. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 549–557. [Google Scholar] [CrossRef]
- Boutet, M.A.; Bart, G.; Penhoat, M.; Amiaud, J.; Brulin, B.; Charrier, C.; Morel, F.; Lecron, J.C.; Rolli-Derkinderen, M.; Bourreille, A.; et al. Distinct expression of interleukin (IL)-36α, β and γ, their antagonist IL-36Ra and IL-38 in psoriasis, rheumatoid arthritis and Crohn’s disease. Clin. Exp. Immunol. 2016, 184, 159–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, A.M.; Baliwag, J.; Chen, C.S.; Guzman, A.M.; Stoll, S.W.; Gudjonsson, J.E.; Ward, N.L.; Johnston, A. IL-36 promotes myeloid cell infiltration, activation, and inflammatory activity in skin. J. Immunol. 2014, 192, 6053–6061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, T.T.; Niyonsaba, F.; Ushio, H.; Akiyama, T.; Kiatsurayanon, C.; Smithrithee, R.; Ikeda, S.; Okumura, K.; Ogawa, H. Interleukin-36 cytokines enhance the production of host defense peptides psoriasin and LL-37 by human keratinocytes through activation of MAPKs and NF-κB. J. Dermatol. Sci. 2012, 68, 63–66. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yin, Y.; Lin, X.; Yan, X.; Xia, Y.; Zhang, L.; Cao, J. IL-36 induces cytokine IL-6 and chemokine CXCL8 expression in human lung tissue cells: Implications for pulmonary inflammatory responses. Cytokine 2017, 99, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lang, C.C.V.; Renert-Yuval, Y.; Del Duca, E.; Pavel, A.B.; Wu, J.; Zhang, N.; Dubin, C.; Obi, A.; Chowdhoury, M.; Kim, M.; et al. Immune and barrier characterization of atopic dermatitis skin phenotype in Tanzanian patients. Ann. Allergy Asthma Immunol. 2021, 127, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Brunner, P.M.; Guttman-Yassky, E. Racial differences in atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 122, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Sanyal, R.D.; Pavel, A.B.; Glickman, J.; Chan, T.C.; Zheng, X.; Zhang, N.; Cueto, I.; Peng, X.; Estrada, Y.; Fuentes-Duculan, J.; et al. Atopic dermatitis in African American patients is TH2/TH22-skewed with TH1/TH17 attenuation. Ann. Allergy Asthma Immunol. 2019, 122, 99–110.e6. [Google Scholar] [CrossRef] [Green Version]
- Noda, S.; Suárez-Fariñas, M.; Ungar, B.; Kim, S.J.; de Guzman Strong, C.; Xu, H.; Peng, X.; Estrada, Y.D.; Nakajima, S.; Honda, T.; et al. The Asian atopic dermatitis phenotype combines features of atopic dermatitis and psoriasis with increased TH17 polarization. J. Allergy Clin. Immunol. 2015, 136, 1254–1264. [Google Scholar] [CrossRef]
- Johnston, A.; Xing, X.; Guzman, A.M.; Riblett, M.; Loyd, C.M.; Ward, N.L.; Wohn, C.; Prens, E.P.; Wang, F.; Maier, L.E.; et al. IL-1F5, -F6, -F8, and -F9: A novel IL-1 family signaling system that is active in psoriasis and promotes keratinocyte antimicrobial peptide expression. J. Immunol. 2011, 186, 2613–2622. [Google Scholar] [CrossRef] [Green Version]
- Thomi, R.; Kakeda, M.; Yawalkar, N.; Schlapbach, C.; Hunger, R.E. Increased expression of the interleukin-36 cytokines in lesions of hidradenitis suppurativa. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 2091–2096. [Google Scholar] [CrossRef]
- Wu, Y.; Gu, C.; Wang, S.; Yin, H.; Qiu, Z.; Luo, Y.; Li, Z.; Wang, C.; Yao, X.; Li, W. Serum biomarker-based endotypes of atopic dermatitis in China and prediction for efficacy of dupilumab. Br. J. Dermatol. 2023, 188, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Yang, H.; Liu, E.; Wang, H. Establishing a Role for Interleukin-17 in Atopic Dermatitis-Related Skin Inflammation. J. Cutan. Med. Surg. 2017, 21, 308–315. [Google Scholar] [CrossRef]
- Ungar, B.; Garcet, S.; Gonzalez, J.; Dhingra, N.; Correa da Rosa, J.; Shemer, A.; Krueger, J.G.; Suarez-Farinas, M.; Guttman-Yassky, E. An Integrated Model of Atopic Dermatitis Biomarkers Highlights the Systemic Nature of the Disease. J. Investig. Dermatol. 2017, 137, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Bridgewood, C.; Fearnley, G.W.; Berekmeri, A.; Laws, P.; Macleod, T.; Ponnambalam, S.; Stacey, M.; Graham, A.; Wittmann, M. IL-36γ Is a Strong Inducer of IL-23 in Psoriatic Cells and Activates Angiogenesis. Front. Immunol. 2018, 9, 200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, W.J.; Elbasiony, E.; Singh, A.; Mittal, S.K.; Chauhan, S.K. IL-36γ Augments Ocular Angiogenesis by Promoting the Vascular Endothelial Growth Factor-Vascular Endothelial Growth Factor Receptor Axis. Am. J. Pathol. 2023; in press. [Google Scholar]
- Samochocki, Z.; Bogaczewicz, J.; Sysa-Jędrzejowska, A.; McCauliffe, D.P.; Kontny, E.; Wozniacka, A. Expression of vascular endothelial growth factor and other cytokines in atopic dermatitis, and correlation with clinical features. Int. J. Dermatol. 2016, 55, e141–e146. [Google Scholar] [CrossRef] [PubMed]
- Zablotna, M.; Sobjanek, M.; Glen, J.; Niedoszytko, M.; Wilkowska, A.; Roszkiewicz, J.; Nedoszytko, B. Association between the—1154 G/A promoter polymorphism of the vascular endothelial growth factor gene and atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 91–92. [Google Scholar] [CrossRef]
- van Aanhold, C.C.L.; Bus, P.; Zandbergen, M.; Bos, M.; Berbée, J.F.P.; Quint, K.D.; Bruijn, J.A.; Baelde, H.J. The Vascular Endothelial Growth Factor Inhibitor Soluble FLT-1 Ameliorates Atopic Dermatitis in APOC1 Transgenic Mice. J. Investig. Dermatol. 2020, 140, 491–494.e4. [Google Scholar] [CrossRef]
- Odorisio, T.; Cianfarani, F.; Failla, C.M.; Zambruno, G. The placenta growth factor in skin angiogenesis. J. Dermatol. Sci. 2006, 41, 11–19. [Google Scholar] [CrossRef]
- Lin, Y.L.; Liang, Y.C.; Chiang, B.L. Placental growth factor down-regulates type 1 T helper immune response by modulating the function of dendritic cells. J. Leukoc. Biol. 2007, 82, 1473–1480. [Google Scholar] [CrossRef] [PubMed]
- Miyagaki, T.; Sugaya, M.; Oka, T.; Takahashi, N.; Kawaguchi, M.; Suga, H.; Fujita, H.; Yoshizaki, A.; Asano, Y.; Sato, S. Placental Growth Factor and Vascular Endothelial Growth Factor Together Regulate Tumour Progression via Increased Vasculature in Cutaneous T-cell Lymphoma. Acta. Derm. Venereol. 2017, 97, 586–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guttman-Yassky, E.; Bissonnette, R.; Ungar, B.; Suárez-Fariñas, M.; Ardeleanu, M.; Esaki, H.; Suprun, M.; Estrada, Y.; Xu, H.; Peng, X.; et al. Dupilumab progressively improves systemic and cutaneous abnormalities in patients with atopic dermatitis. J. Allergy Clin. Immunol. 2019, 143, 155–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yew, Y.W.; Dinish, U.S.; Choi, E.C.E.; Bi, R.; Ho, C.J.H.; Dev, K.; Li, X.; Attia, A.B.E.; Wong, M.K.W.; Balasundaram, G.; et al. Investigation of morphological, vascular and biochemical changes in the skin of an atopic dermatitis (AD) patient in response to dupilumab using raster scanning optoacoustic mesoscopy (RSOM) and handheld confocal Raman spectroscopy (CRS). J. Dermatol. Sci. 2019, 95, 123–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanifin, J.M.; Rajka, G. Diagnostic features of atopic dermatitis. Acta. Derm. Venereol. Suppl. 1980, 92, 44–47. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atopic Dermatitis (n = 36) | Healthy Controls (n = 15) | |
|---|---|---|
| Sex (Male:Female) | 26:10 | 8:7 |
| Age (years) | 38.9 ± 12.4 | 44.3 ± 9.6 |
| Atopic comorbidities | ||
| Allergic rhinitis | 12 (33.3%) | 0 (0%) |
| Allergic conjunctivitis | 1 (2.6%) | 0 (0%) |
| Asthma | 8 (22.2%) | 0 (0%) |
| EASI | 20.6 ± 10.1 | - |
| The number of eosinophils in peripheral blood (/μL) | 484 ± 397 | - |
| Serum LDH levels (U/L) | 259 ± 114 | - |
| Serum IgE levels (IU/mL) | 10,628 ± 9808 | - |
| Serum TARC levels (pg/mL) | 3487 ± 5279 | - |
| Target | Forward/Reverse (5′-3′) |
|---|---|
| IL-25 | CCAGGTGGTTGCATTCTTGG/TGGCTGTAGGTGTGGGTTCC |
| IL-33 | GGAAGAACACAGCAAGCAAAGCCT/ TAAGGCCAGAGCGGAGCTTCATAA |
| TSLP | CCCAGGCTATTCGGAAACTCA/ACGCCACAATCCTTGTAATTGTG |
| CCL17 | GCAAAGCCTTGAGAGGTCTTTGA/CGGTGGAGGTCCCAGGTAGT |
| CCL26 | TCTGTACCCATCCAAGGAAAA/GGGTCCATGTAGCCTTCAGA |
| VEGF-A | AGCCTTGCCTTGCTGCTCTAC/TCCTCCTTCTGCCATGGGT |
| PlGF | GCGATGAGAATCTGCACTCTGT/TCCCCAGAACGGATCTTTAGG |
| GAPDH | ACCCACTCCTCCACCTTTGA/CATACCAGGAAATGAGCTTGACAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Komaki, R.; Miyagaki, T.; Tanaka, M.; Nakajima, K.; Okano, T.; Takeuchi, S.; Kadono, T. Increased Interleukin-36β Expression Promotes Angiogenesis in Japanese Atopic Dermatitis. Int. J. Mol. Sci. 2023, 24, 11104. https://doi.org/10.3390/ijms241311104
Komaki R, Miyagaki T, Tanaka M, Nakajima K, Okano T, Takeuchi S, Kadono T. Increased Interleukin-36β Expression Promotes Angiogenesis in Japanese Atopic Dermatitis. International Journal of Molecular Sciences. 2023; 24(13):11104. https://doi.org/10.3390/ijms241311104
Chicago/Turabian StyleKomaki, Reo, Tomomitsu Miyagaki, Miho Tanaka, Kaori Nakajima, Tatsuro Okano, Sora Takeuchi, and Takafumi Kadono. 2023. "Increased Interleukin-36β Expression Promotes Angiogenesis in Japanese Atopic Dermatitis" International Journal of Molecular Sciences 24, no. 13: 11104. https://doi.org/10.3390/ijms241311104