
Hypoxia Preconditioned Serum (HPS) Promotes Proliferation and Chondrogenic Phenotype of Chondrocytes In Vitro
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
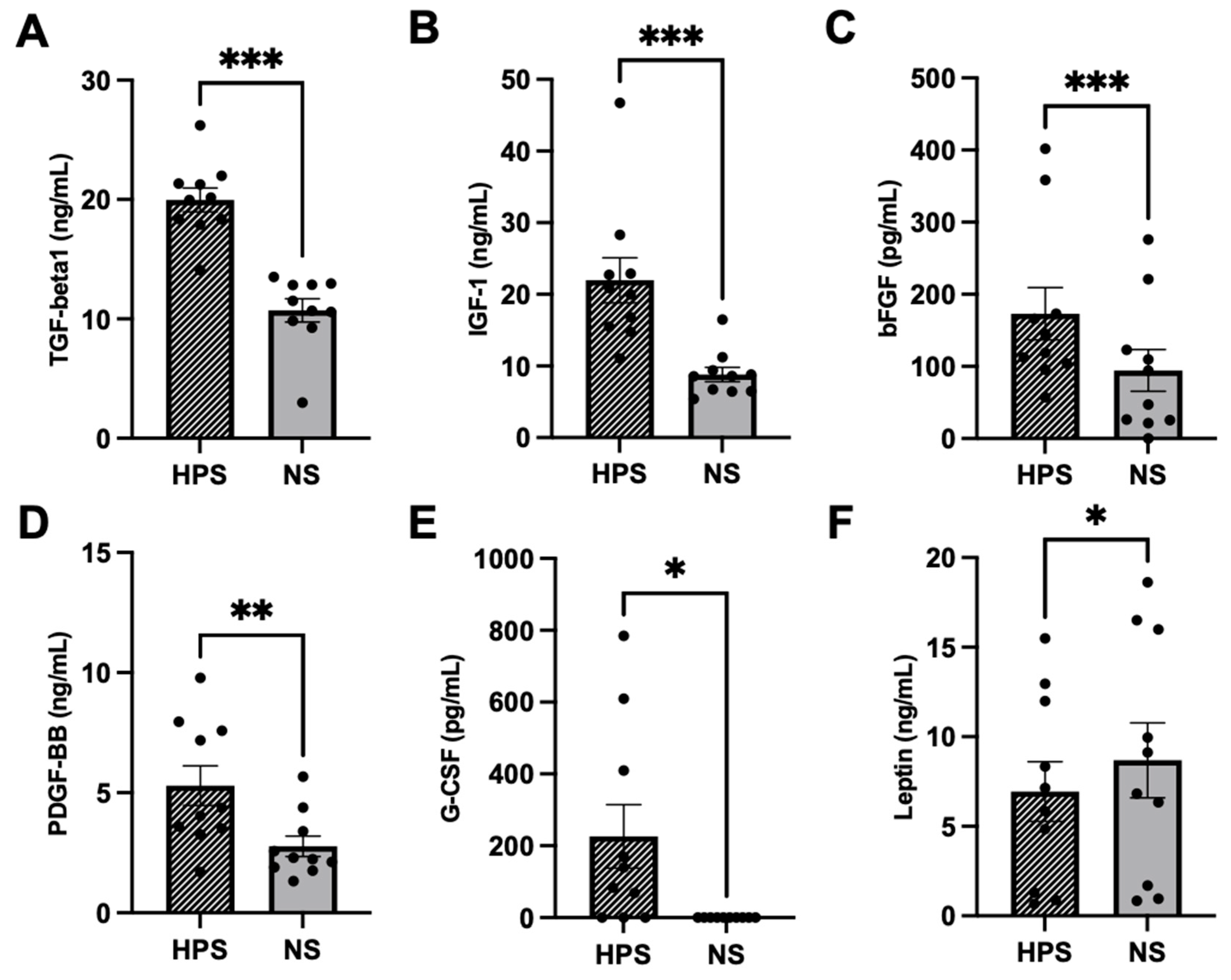
2.1. Quantitative Analysis of Chondrogenic Growth Factors in HPS
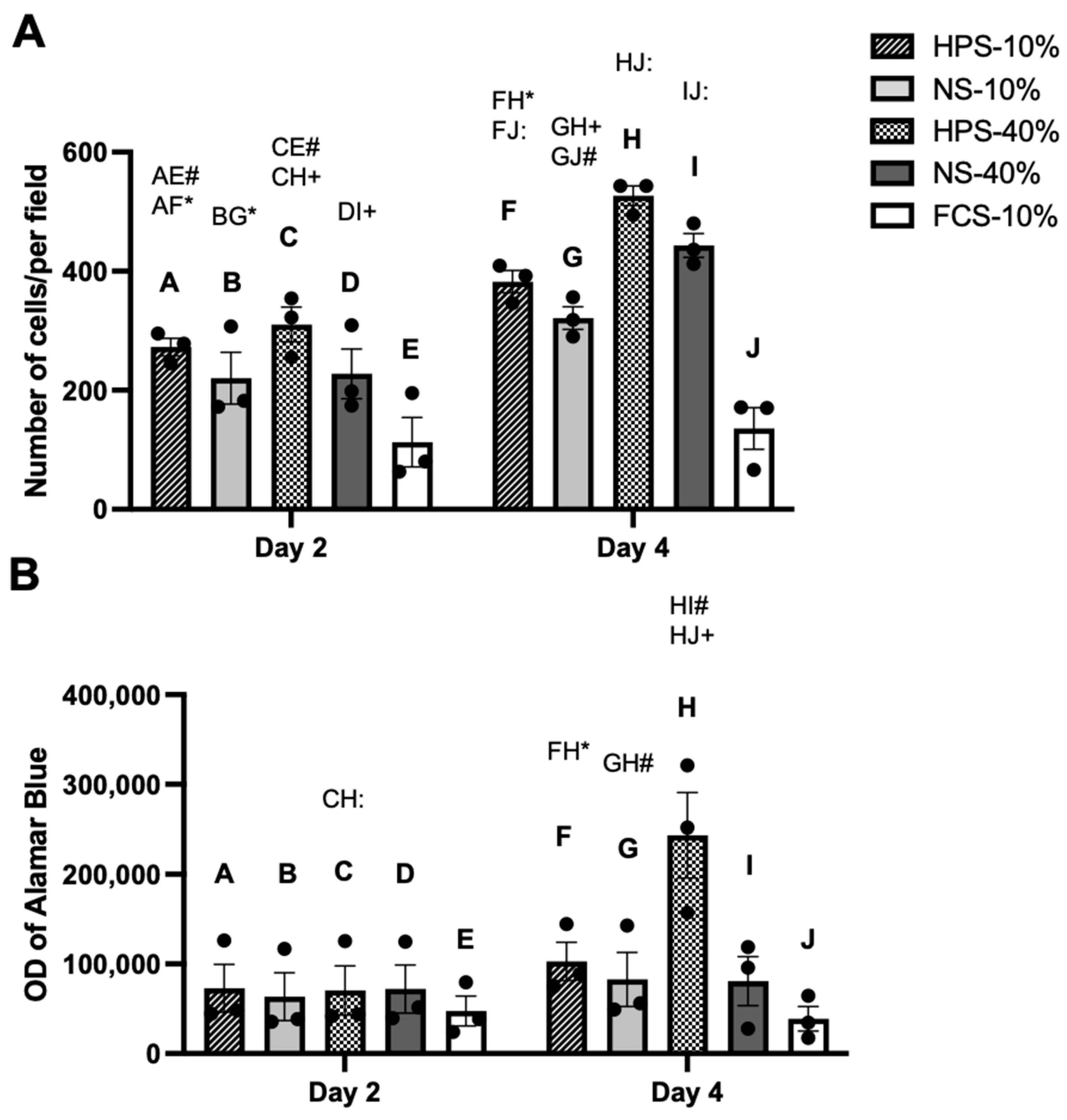
2.2. The Effect of HPS on Chondrocyte Proliferation and Metabolic Activity
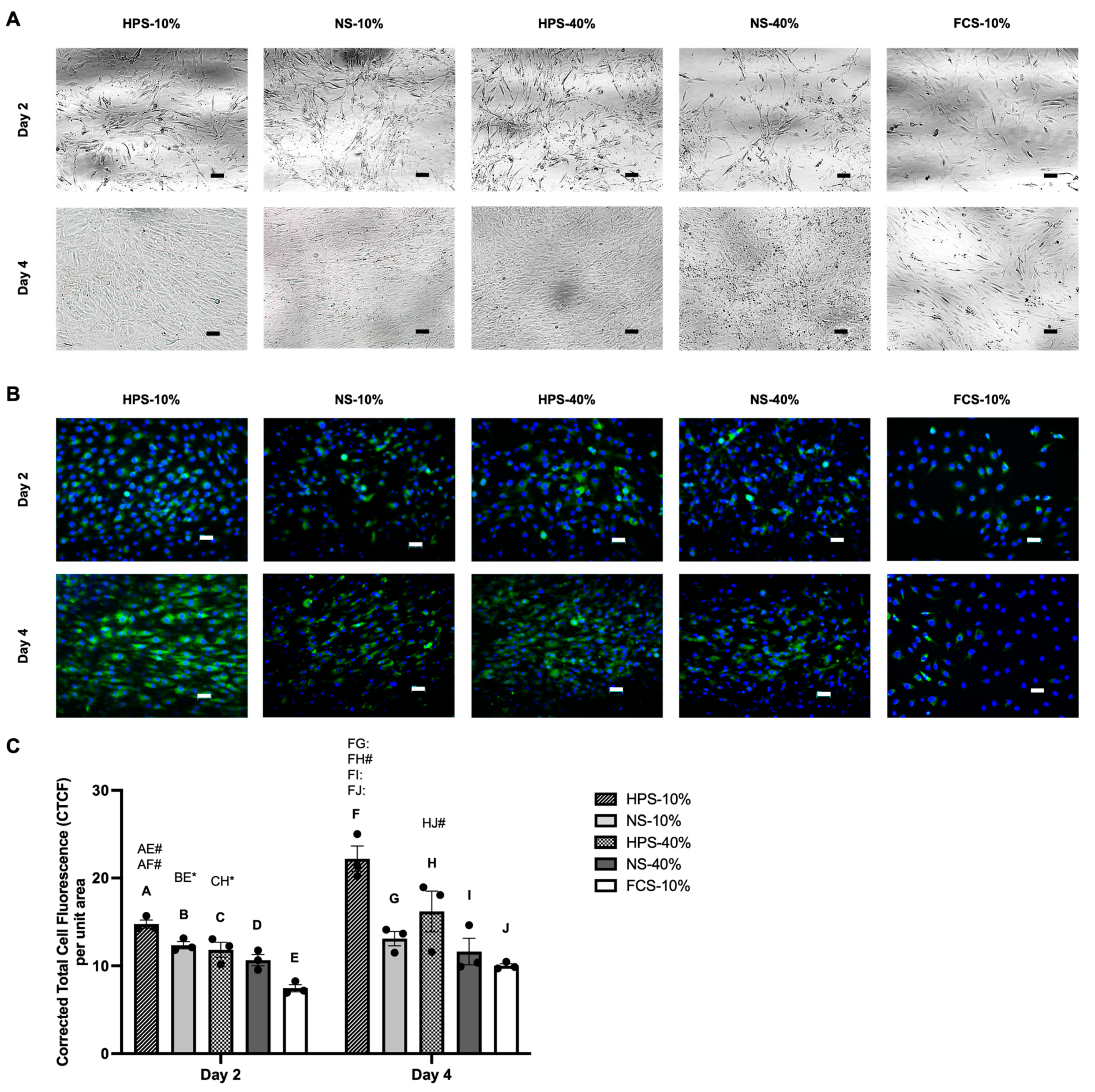
2.3. Macroscopic Cell Appearance and Immunofluorescence Analysis of Collagen Type II
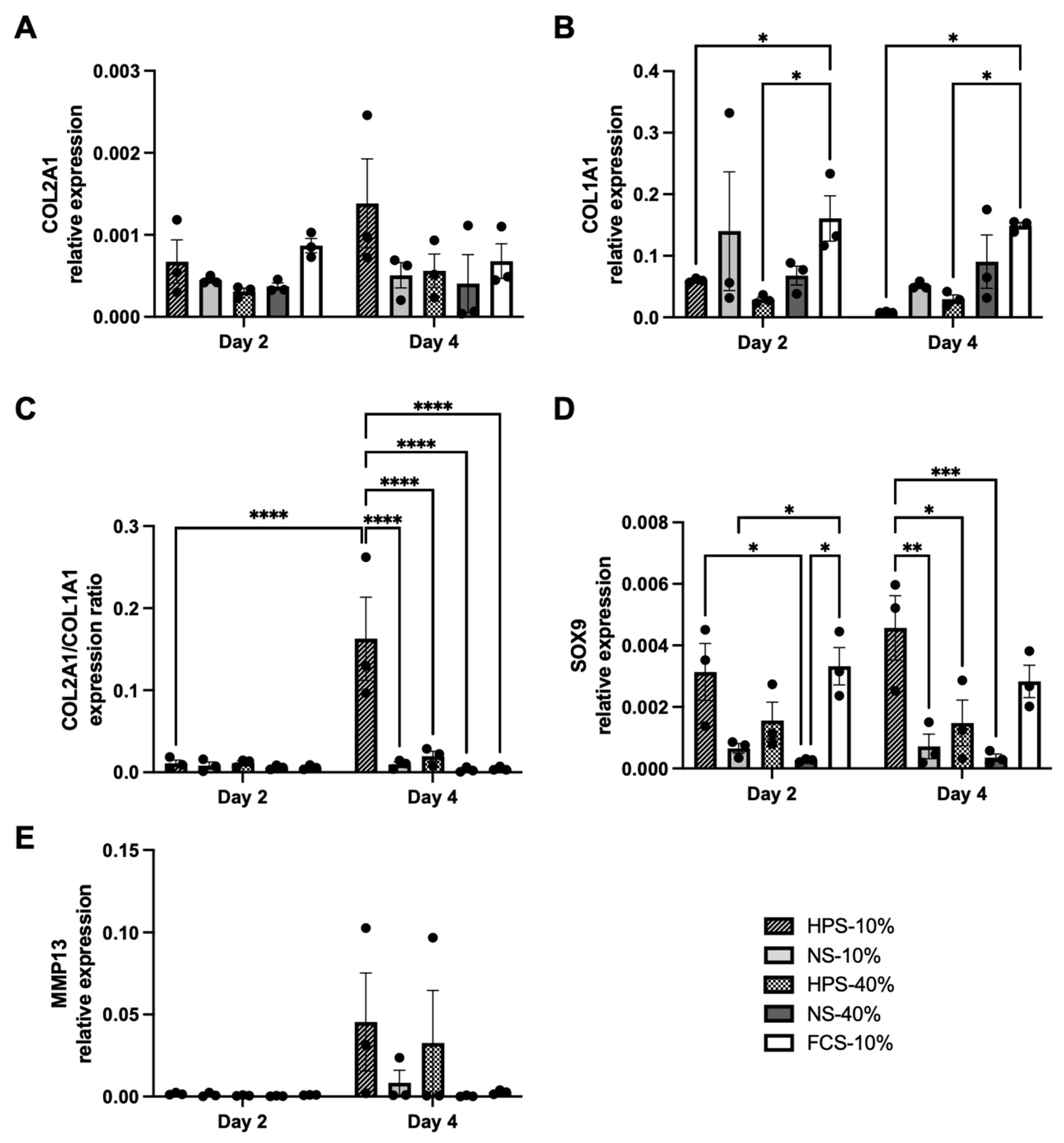
2.4. The Effect of HPS on Chondrocyte Differentiation
3. Discussion
4. Materials and Methods
4.1. Ethical Approval
4.2. Production of Hypoxia Preconditioned Serum (HPS)
4.3. Production of Fresh Normal Serum (NS)
4.4. Cell Culture
4.5. Quantification of the Chondrogenic Cytokines
4.6. Alamar Blue Metabolic Assay
4.7. Immunofluorescence Staining
4.8. Quantification of Cell Number and Fluorescence Intensity
4.9. Analysis of Gene Expression
4.10. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACI | Autologous chondrocyte implantation |
| bFGF | Basic fibroblast growth factor |
| COL1A1 | Collagen type 1 alpha 1 |
| COL2A1 | Collagen type 2 alpha 1 |
| G-CSF | Granulocyte colony-stimulating factor |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HPS | Hypoxia Preconditioned Serum |
| IGF-1 | Insulin-like growth factor-1 |
| IL-1 | Interleukin-1 |
| MMP-13 | Matrix metalloproteinase-13 |
| PBC | Peripheral blood cells |
| PBS | Phosphate-buffered saline |
| PDGF-BB | Platelet-derived growth factor-beta polypeptide |
| PRP | Platelet-rich plasma |
| SOX9 | Sex-determining region Y-box 9 |
| RT-qPCR | Real-time quantitative reverse transcription-polymerase chain reaction |
| TGF-beta1 | Transforming growth factor-beta 1 |
| TNF-alpha | Tumour necrosis factor-alpha |
References
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, H.; Jiang, C.-C. Repair of Articular Cartilage Defects: Review and Perspectives. J. Formos. Med. Assoc. 2009, 108, 87–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedi, A.; Feeley, B.T.; Williams, R.J., 3rd. Management of articular cartilage defects of the knee. J. Bone Joint Surg. Am. 2010, 92, 994–1009. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Liu, Y.; Shah, K.M.; Luo, J. Strategies for Articular Cartilage Repair and Regeneration. Front. Bioeng. Biotechnol. 2021, 9, 1328. [Google Scholar] [CrossRef]
- Ogura, T.; Bryant, T.; Minas, T. Long-term Outcomes of Autologous Chondrocyte Implantation in Adolescent Patients. Am. J. Sports Med. 2017, 45, 1066–1074. [Google Scholar] [CrossRef]
- Davies, R.L.; Kuiper, N.J. Regenerative Medicine: A Review of the Evolution of Autologous Chondrocyte Implantation (ACI) Therapy. Bioengineering 2019, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Schnabel, M.; Marlovits, S.; Eckhoff, G.; Fichtel, I.; Gotzen, L.; Vécsei, V.; Schlegel, J. Dedifferentiation-associated changes in morphology and gene expression in primary human articular chondrocytes in cell culture. Osteoarthr. Cartil. 2002, 10, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Jeyakumar, V.; Niculescu-Morzsa, E.; Bauer, C.; Lacza, Z.; Nehrer, S. Platelet-Rich Plasma Supports Proliferation and Redifferentiation of Chondrocytes during In Vitro Expansion. Front. Bioeng. Biotechnol. 2017, 5, 75. [Google Scholar] [CrossRef]
- Pei, M.; Seidel, J.; Vunjak-Novakovic, G.; Freed, L.E. Growth factors for sequential cellular de- and re-differentiation in tissue engineering. Biochem. Biophys. Res. Commun. 2002, 294, 149–154. [Google Scholar] [CrossRef]
- Aurich, M.; Hofmann, G.O.; Best, N.; Rolauffs, B. Induced Redifferentiation of Human Chondrocytes from Articular Cartilage Lesion in Alginate Bead Culture After Monolayer Dedifferentiation: An Alternative Cell Source for Cell-Based Therapies? Tissue Eng. A 2018, 24, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Zhang, W.; Li, X.; Zhong, D.; Li, Y.; Li, J.; Jin, R. Strategies to Modulate the Redifferentiation of Chondrocytes. Front. Bioeng. Biotechnol. 2021, 9, 764193. [Google Scholar] [CrossRef] [PubMed]
- Duval, E.; Leclercq, S.; Elissalde, J.M.; Demoor, M.; Galéra, P.; Boumédiene, K. Hypoxia-inducible factor 1alpha inhibits the fibroblast-like markers type I and type III collagen during hypoxia-induced chondrocyte redifferentiation: Hypoxia not only induces type II collagen and aggrecan, but it also inhibits type I and type III collagen in the hypoxia-inducible factor 1alpha-dependent redifferentiation of chondrocytes. Arthritis Rheum. 2009, 60, 3038–3048. [Google Scholar] [CrossRef]
- Pötter, N.; Westbrock, F.; Grad, S.; Alini, M.; Stoddart, M.J.; Schmal, H.; Kubosch, D.; Salzmann, G.; Kubosch, E.J. Evaluation of the influence of platelet-rich plasma (PRP), platelet lysate (PL) and mechanical loading on chondrogenesis in vitro. Sci. Rep. 2021, 11, 20188. [Google Scholar] [CrossRef] [PubMed]
- Brandl, A.; Angele, P.; Roll, C.; Prantl, L.; Kujat, R.; Kinner, B. Influence of the growth factors PDGF-BB, TGF-beta1 and bFGF on the replicative aging of human articular chondrocytes during in vitro expansion. J. Orthop. Res. 2010, 28, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Feng, J.; Wu, W.K.; Xiao, J.; Wu, Z.; Han, D.; Zhu, Y.; Qiu, G. Leptin-mediated cytoskeletal remodeling in chondrocytes occurs via the RhoA/ROCK pathway. J. Orthop. Res. 2011, 29, 369–374. [Google Scholar] [CrossRef]
- Marmotti, A.; Bonasia, D.E.; Bruzzone, M.; Rossi, R.; Castoldi, F.; Collo, G.; Realmuto, C.; Tarella, C.; Peretti, G.M. Human cartilage fragments in a composite scaffold for single-stage cartilage repair: An in vitro study of the chondrocyte migration and the influence of TGF-β1 and G-CSF. Knee Surg. Sports Traumatol. Arthrosc. 2013, 21, 1819–1833. [Google Scholar] [CrossRef]
- Jakobsen, R.B.; Østrup, E.; Zhang, X.; Mikkelsen, T.S.; Brinchmann, J.E. Analysis of the effects of five factors relevant to in vitro chondrogenesis of human mesenchymal stem cells using factorial design and high throughput mRNA-profiling. PLoS ONE 2014, 9, e96615. [Google Scholar] [CrossRef]
- Mullen, L.M.; Best, S.M.; Ghose, S.; Wardale, J.; Rushton, N.; Cameron, R.E. Bioactive IGF-1 release from collagen-GAG scaffold to enhance cartilage repair in vitro. J. Mater. Sci. Mater. Med. 2015, 26, 5325. [Google Scholar] [CrossRef] [Green Version]
- Vonk, L.A.; Roël, G.; Hernigou, J.; Kaps, C.; Hernigou, P. Role of Matrix-Associated Autologous Chondrocyte Implantation with Spheroids in the Treatment of Large Chondral Defects in the Knee: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 7149. [Google Scholar] [CrossRef]
- Drengk, A.; Zapf, A.; Stürmer, E.K.; Stürmer, K.M.; Frosch, K.H. Influence of platelet-rich plasma on chondrogenic differentiation and proliferation of chondrocytes and mesenchymal stem cells. Cells Tissues Organs 2009, 189, 317–326. [Google Scholar] [CrossRef]
- Okuda, K.; Kawase, T.; Momose, M.; Murata, M.; Saito, Y.; Suzuki, H.; Wolff, L.F.; Yoshie, H. Platelet-rich plasma contains high levels of platelet-derived growth factor and transforming growth factor-beta and modulates the proliferation of periodontally related cells in vitro. J. Periodontol. 2003, 74, 849–857. [Google Scholar] [CrossRef]
- Spreafico, A.; Chellini, F.; Frediani, B.; Bernardini, G.; Niccolini, S.; Serchi, T.; Collodel, G.; Paffetti, A.; Fossombroni, V.; Galeazzi, M.; et al. Biochemical investigation of the effects of human platelet releasates on human articular chondrocytes. J. Cell. Biochem. 2009, 108, 1153–1165. [Google Scholar] [CrossRef] [PubMed]
- Akeda, K.; An, H.S.; Okuma, M.; Attawia, M.; Miyamoto, K.; Thonar, E.J.; Lenz, M.E.; Sah, R.L.; Masuda, K. Platelet-rich plasma stimulates porcine articular chondrocyte proliferation and matrix biosynthesis. Osteoarthr. Cartil. 2006, 14, 1272–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gobbi, A.; Lad, D.; Karnatzikos, G. The effects of repeated intra-articular PRP injections on clinical outcomes of early osteoarthritis of the knee. Knee Surg. Sports Traumatol. Arthrosc. 2015, 23, 2170–2177. [Google Scholar] [CrossRef] [PubMed]
- Alsousou, J.; Ali, A.; Willett, K.; Harrison, P. The role of platelet-rich plasma in tissue regeneration. Platelets 2013, 24, 173–182. [Google Scholar] [CrossRef]
- Lang, S.; Loibl, M.; Herrmann, M. Platelet-Rich Plasma in Tissue Engineering: Hype and Hope. Eur. Surg. Res. 2018, 59, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayi, E.; Moog, P.; Bekeran, S.; Kirchhoff, K.; Berezhnoi, A.; Aguirre, J.; Bauer, A.T.; Kukrek, H.; Schmauss, D.; Hopfner, U.; et al. In Vitro Characterization of Hypoxia Preconditioned Serum (HPS)-Fibrin Hydrogels: Basis for an Injectable Biomimetic Tissue Regeneration Therapy. J. Funct. Biomater. 2019, 10, 22. [Google Scholar] [CrossRef] [Green Version]
- Hadjipanayi, E.; Bauer, A.T.; Moog, P.; Salgin, B.; Kuekrek, H.; Fersch, B.; Hopfner, U.; Meissner, T.; Schluter, A.; Ninkovic, M.; et al. Cell-free carrier system for localized delivery of peripheral blood cell-derived engineered factor signaling: Towards development of a one-step device for autologous angiogenic therapy. J. Control. Release 2013, 169, 91–102. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Cheema, U.; Hopfner, U.; Bauer, A.; Machens, H.G.; Schilling, A.F. Injectable system for spatio-temporally controlled delivery of hypoxia-induced angiogenic signalling. J. Control. Release 2012, 161, 852–860. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Cheema, U.; Mudera, V.; Deng, D.; Liu, W.; Brown, R.A. First implantable device for hypoxia-mediated angiogenic induction. J. Control. Release 2011, 153, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Hadjipanayi, E.; Kuhn, P.H.; Moog, P.; Bauer, A.T.; Kuekrek, H.; Mirzoyan, L.; Hummel, A.; Kirchhoff, K.; Salgin, B.; Isenburg, S.; et al. The Fibrin Matrix Regulates Angiogenic Responses within the Hemostatic Microenvironment through Biochemical Control. PLoS ONE 2015, 10, e0135618. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Bekeran, S.; Moog, P. Extracorporeal Wound Simulation as a Foundation for Tissue Repair and Regeneration Therapies. Int. J. Transplant. Plast. Surg. 2018, 2, 1–10. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Schilling, A.F. Regeneration through autologous hypoxia preconditioned plasma. Organogenesis 2014, 10, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Moog, P.; Kirchhoff, K.; Bekeran, S.; Bauer, A.T.; von Isenburg, S.; Dornseifer, U.; Machens, H.G.; Schilling, A.F.; Hadjipanayi, E. Comparative Evaluation of the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes and Platelet-Rich Plasma: An In Vitro Analysis. Biomedicines 2020, 8, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moog, P.; Hughes, J.; Jiang, J.; Röper, L.; Dornseifer, U.; Schilling, A.F.; Machens, H.-G.; Hadjipanayi, E. Comparison of the Effect of Different Conditioning Media on the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes: Towards Engineering Next-Generation Autologous Growth Factor Cocktails. Int. J. Mol. Sci. 2023, 24, 5485. [Google Scholar] [CrossRef]
- Jiang, J.; Cong, X.; Alageel, S.; Dornseifer, U.; Schilling, A.F.; Hadjipanayi, E.; Machens, H.-G.; Moog, P. In Vitro Comparison of Lymphangiogenic Potential of Hypoxia Preconditioned Serum (HPS) and Platelet-Rich Plasma (PRP). Int. J. Mol. Sci. 2023, 24, 1961. [Google Scholar] [CrossRef] [PubMed]
- Moog, P.; Schams, R.; Schneidinger, A.; Schilling, A.F.; Machens, H.G.; Hadjipanayi, E.; Dornseifer, U. Effect of Hypoxia Preconditioned Secretomes on Lymphangiogenic and Angiogenic Sprouting: An in Vitro Analysis. Biomedicines 2020, 8, 365. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Brown, R.A.; Mudera, V.; Deng, D.; Liu, W.; Cheema, U. Controlling physiological angiogenesis by hypoxia-induced signaling. J. Control. Release 2010, 146, 309–317. [Google Scholar] [CrossRef]
- Hadjipanayi, E.; Schilling, A.F. Hypoxia-based strategies for angiogenic induction: The dawn of a new era for ischemia therapy and tissue regeneration. Organogenesis 2013, 9, 261–272. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Kraneburg, U.; Dornseifer, U.; Schilling, A.F.; Hadjipanayi, E.; Machens, H.G.; Moog, P. Hypoxia Preconditioned Serum (HPS)-Hydrogel Can Accelerate Dermal Wound Healing in Mice-An In Vivo Pilot Study. Biomedicines 2022, 10, 176. [Google Scholar] [CrossRef] [PubMed]
- Moog, P.; Jensch, M.; Hughes, J.; Salgin, B.; Dornseifer, U.; Machens, H.G.; Schilling, A.F.; Hadjipanayi, E. Use of Oral Anticoagulation and Diabetes Do Not Inhibit the Angiogenic Potential of Hypoxia Preconditioned Blood-Derived Secretomes. Biomedicines 2020, 8, 283. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Röper, L.; Alageel, S.; Dornseifer, U.; Schilling, A.F.; Hadjipanayi, E.; Machens, H.-G.; Moog, P. Hypoxia Preconditioned Serum (HPS) Promotes Osteoblast Proliferation, Migration and Matrix Deposition. Biomedicines 2022, 10, 1631. [Google Scholar] [CrossRef] [PubMed]
- Colombini, A.; Libonati, F.; Lopa, S.; Peretti, G.M.; Moretti, M.; de Girolamo, L. Autologous chondrocyte implantation provides good long-term clinical results in the treatment of knee osteoarthritis: A systematic review. Knee Surg. Sports Traumatol. Arthrosc. 2022, 31, 2338–2348. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Bunpetch, V.; Zhou, W.; Ouyang, H. Optimization strategies for ACI: A step-chronicle review. J. Orthop. Transl. 2019, 17, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Chow, V.T.; Phoon, M.C. Measurement of serum leptin concentrations in university undergraduates by competitive ELISA reveals correlations with body mass index and sex. Adv. Physiol. Educ. 2003, 27, 70–77. [Google Scholar] [CrossRef]
- Otero, M.; Lago, R.; Lago, F.; Reino, J.J.G.; Gualillo, O. Signalling pathway involved in nitric oxide synthase type II activation in chondrocytes: Synergistic effect of leptin with interleukin-1. Arthritis Res. Ther. 2005, 7, R581. [Google Scholar] [CrossRef] [Green Version]
- Quintero, M.; Colantuoni, G.; Khatib, A.M.; Panasyuk, A.; Lomri, A.; Mitrovic, D.R. Granulocyte-macrophage colony stimulating factor activates proteoglycan, type II collagen, and cAMP production by rat articular chondrocytes through specific binding sites. J. Rheumatol. 2001, 28, 2075–2084. [Google Scholar]
- Montaseri, A.; Busch, F.; Mobasheri, A.; Buhrmann, C.; Aldinger, C.; Rad, J.S.; Shakibaei, M. IGF-1 and PDGF-bb suppress IL-1β-induced cartilage degradation through down-regulation of NF-κB signaling: Involvement of Src/PI-3K/AKT pathway. PLoS ONE 2011, 6, e28663. [Google Scholar] [CrossRef] [Green Version]
- Vuolteenaho, K.; Moilanen, T.; Jalonen, U.; Lahti, A.; Nieminen, R.; van Beuningen, H.M.; van der Kraan, P.M.; Moilanen, E. TGFbeta inhibits IL-1-induced iNOS expression and NO production in immortalized chondrocytes. Inflamm. Res. 2005, 54, 420–427. [Google Scholar] [CrossRef]
- El-Sharkawy, H.; Kantarci, A.; Deady, J.; Hasturk, H.; Liu, H.; Alshahat, M.; Van Dyke, T.E. Platelet-rich plasma: Growth factors and pro- and anti-inflammatory properties. J. Periodontol. 2007, 78, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qian, Y.; Han, Q.; Chen, W.; Song, J.; Zhao, X.; Ouyang, Y.; Yuan, W.; Fan, C. Platelet-Rich Plasma Derived Growth Factors Contribute to Stem Cell Differentiation in Musculoskeletal Regeneration. Front. Chem. 2017, 5, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moussa, M.; Lajeunesse, D.; Hilal, G.; El Atat, O.; Haykal, G.; Serhal, R.; Chalhoub, A.; Khalil, C.; Alaaeddine, N. Platelet rich plasma (PRP) induces chondroprotection via increasing autophagy, anti-inflammatory markers, and decreasing apoptosis in human osteoarthritic cartilage. Exp. Cell Res. 2017, 352, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Jeyakumar, V.; Niculescu-Morzsa, E.; Bauer, C.; Lacza, Z.; Nehrer, S. Redifferentiation of Articular Chondrocytes by Hyperacute Serum and Platelet Rich Plasma in Collagen Type I Hydrogels. Int. J. Mol. Sci. 2019, 20, 316. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Zhang, Z.; Sheng, P.; Mobasheri, A. The role of metabolism in chondrocyte dysfunction and the progression of osteoarthritis. Ageing Res. Rev. 2021, 66, 101249. [Google Scholar] [CrossRef]
- Yamamoto, K.; Okano, H.; Miyagawa, W.; Visse, R.; Shitomi, Y.; Santamaria, S.; Dudhia, J.; Troeberg, L.; Strickland, D.K.; Hirohata, S.; et al. MMP-13 is constitutively produced in human chondrocytes and co-endocytosed with ADAMTS-5 and TIMP-3 by the endocytic receptor LRP1. Matrix Biol. 2016, 56, 57–73. [Google Scholar] [CrossRef]
- Borzí, R.M.; Olivotto, E.; Pagani, S.; Vitellozzi, R.; Neri, S.; Battistelli, M.; Falcieri, E.; Facchini, A.; Flamigni, F.; Penzo, M.; et al. Matrix metalloproteinase 13 loss associated with impaired extracellular matrix remodeling disrupts chondrocyte differentiation by concerted effects on multiple regulatory factors. Arthritis Rheum. 2010, 62, 2370–2381. [Google Scholar] [CrossRef] [Green Version]
- Murphy, C.L.; Thoms, B.L.; Vaghjiani, R.J.; Lafont, J.E. Hypoxia. HIF-mediated articular chondrocyte function: Prospects for cartilage repair. Arthritis Res. Ther. 2009, 11, 213. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Li, X.; Jing, X.; Li, M.; Ren, Y.; Chen, J.; Yang, C.; Wu, H.; Guo, F. Hypoxia promotes maintenance of the chondrogenic phenotype in rat growth plate chondrocytes through the HIF-1α/YAP signaling pathway. Int. J. Mol. Med. 2018, 42, 3181–3192. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wu, Y.; Yang, D.; Neo, S.H.; Kadir, N.D.; Goh, D.; Tan, J.X.; Denslin, V.; Lee, E.H.; Yang, Z. Secretive derived from hypoxia preconditioned mesenchymal stem cells promote cartilage regeneration and mitigate joint inflammation via extracellular vesicles. Bioact. Mater. 2023, 27, 98–112. [Google Scholar] [CrossRef]
- Zhang, B.; Tian, X.; Qu, Z.; Hao, J.; Zhang, W. Hypoxia-Preconditioned Extracellular Vesicles from Mesenchymal Stem Cells Improve Cartilage Repair in Osteoarthritis. Membranes 2022, 12, 225. [Google Scholar] [CrossRef] [PubMed]
- Barbon, S.; Stocco, E.; Macchi, V.; Contran, M.; Grandi, F.; Borean, A.; Parnigotto, P.P.; Porzionato, A.; De Caro, R. Platelet-Rich Fibrin Scaffolds for Cartilage and Tendon Regenerative Medicine: From Bench to Bedside. Int. J. Mol. Sci. 2019, 20, 1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nichele, L.; Persichetti, V.; Lucidi, M.; Cincotti, G. Quantitative evaluation of ImageJ thresholding algorithms for microbial cell counting. OSA Contin. 2020, 3, 1417–1427. [Google Scholar] [CrossRef]
- Bora, P.; Gahurova, L.; Mašek, T.; Hauserova, A.; Potěšil, D.; Jansova, D.; Susor, A.; Zdráhal, Z.; Ajduk, A.; Pospíšek, M.; et al. p38-MAPK-mediated translation regulation during early blastocyst development is required for primitive endoderm differentiation in mice. Commun. Biol. 2021, 4, 788. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Forward | Primer Reverse |
|---|---|---|
| GAPDH | CTCTGCTCCTCCTGTTCGAC | ACGACCAAATCCGTTGACTC |
| COL2A1 | GTGTCAGGGCCAGGATGT | TCCCAGTGTCACAGACACAGAT |
| COL1A1 | GGGATTCCCTGGACCTAAAG | GGAACACCTCGCTCTCCAG |
| SOX9 | TACCCGCACTTGCACAAC | TCTCGCTCTCGTTCAGAAGTC |
| MMP13 | TTTCCTCCTGGGCCAAAT | GCAACAAGAAACAAGTTGTAGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, J.; Altammar, J.; Cong, X.; Ramsauer, L.; Steinbacher, V.; Dornseifer, U.; Schilling, A.F.; Machens, H.-G.; Moog, P. Hypoxia Preconditioned Serum (HPS) Promotes Proliferation and Chondrogenic Phenotype of Chondrocytes In Vitro. Int. J. Mol. Sci. 2023, 24, 10441. https://doi.org/10.3390/ijms241310441
Jiang J, Altammar J, Cong X, Ramsauer L, Steinbacher V, Dornseifer U, Schilling AF, Machens H-G, Moog P. Hypoxia Preconditioned Serum (HPS) Promotes Proliferation and Chondrogenic Phenotype of Chondrocytes In Vitro. International Journal of Molecular Sciences. 2023; 24(13):10441. https://doi.org/10.3390/ijms241310441
Chicago/Turabian StyleJiang, Jun, Jannat Altammar, Xiaobin Cong, Lukas Ramsauer, Vincent Steinbacher, Ulf Dornseifer, Arndt F. Schilling, Hans-Günther Machens, and Philipp Moog. 2023. "Hypoxia Preconditioned Serum (HPS) Promotes Proliferation and Chondrogenic Phenotype of Chondrocytes In Vitro" International Journal of Molecular Sciences 24, no. 13: 10441. https://doi.org/10.3390/ijms241310441