
PTEN Inhibitor Treatment Lowers Muscle Plasma Membrane Damage and Enhances Muscle ECM Homeostasis after High-Intensity Eccentric Exercise in Mice

Abstract
:1. Introduction
2. Results
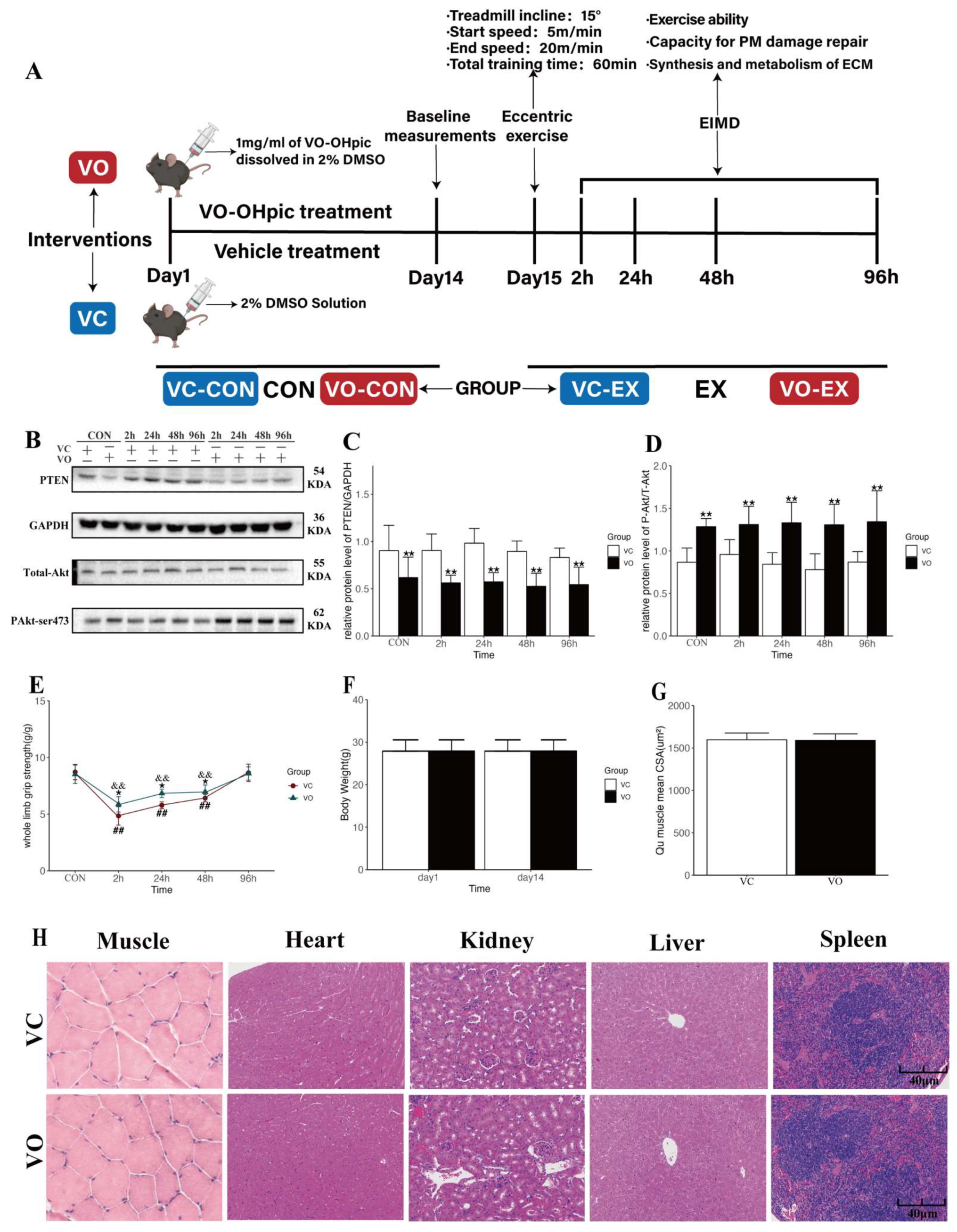
2.1. Two-Week VO-OHpic Treatment Improves Motor Function during EIMD in Mice
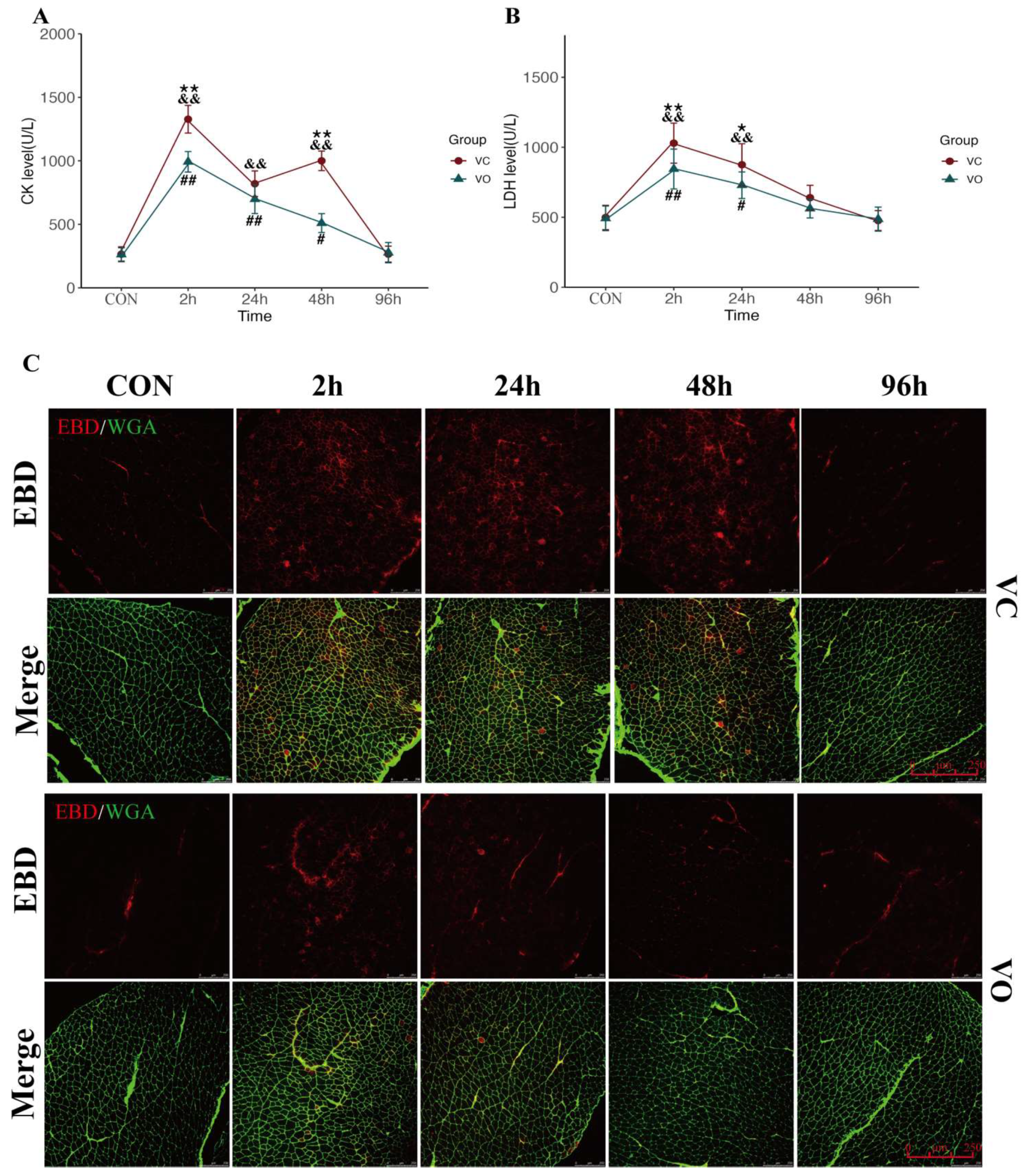
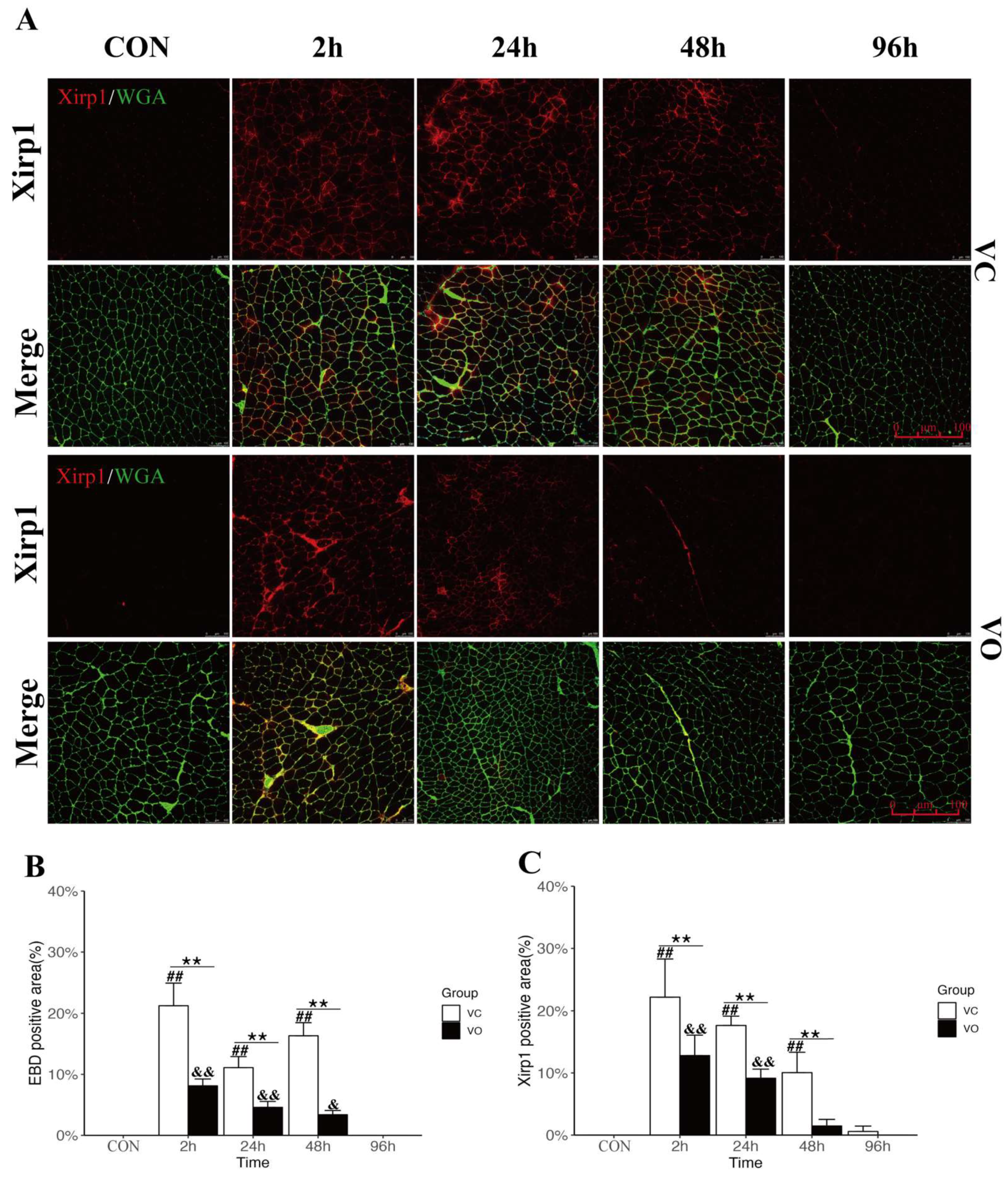
2.2. PTEN Inhibition Enhances Myofibrillar Membrane Repair during EIMD
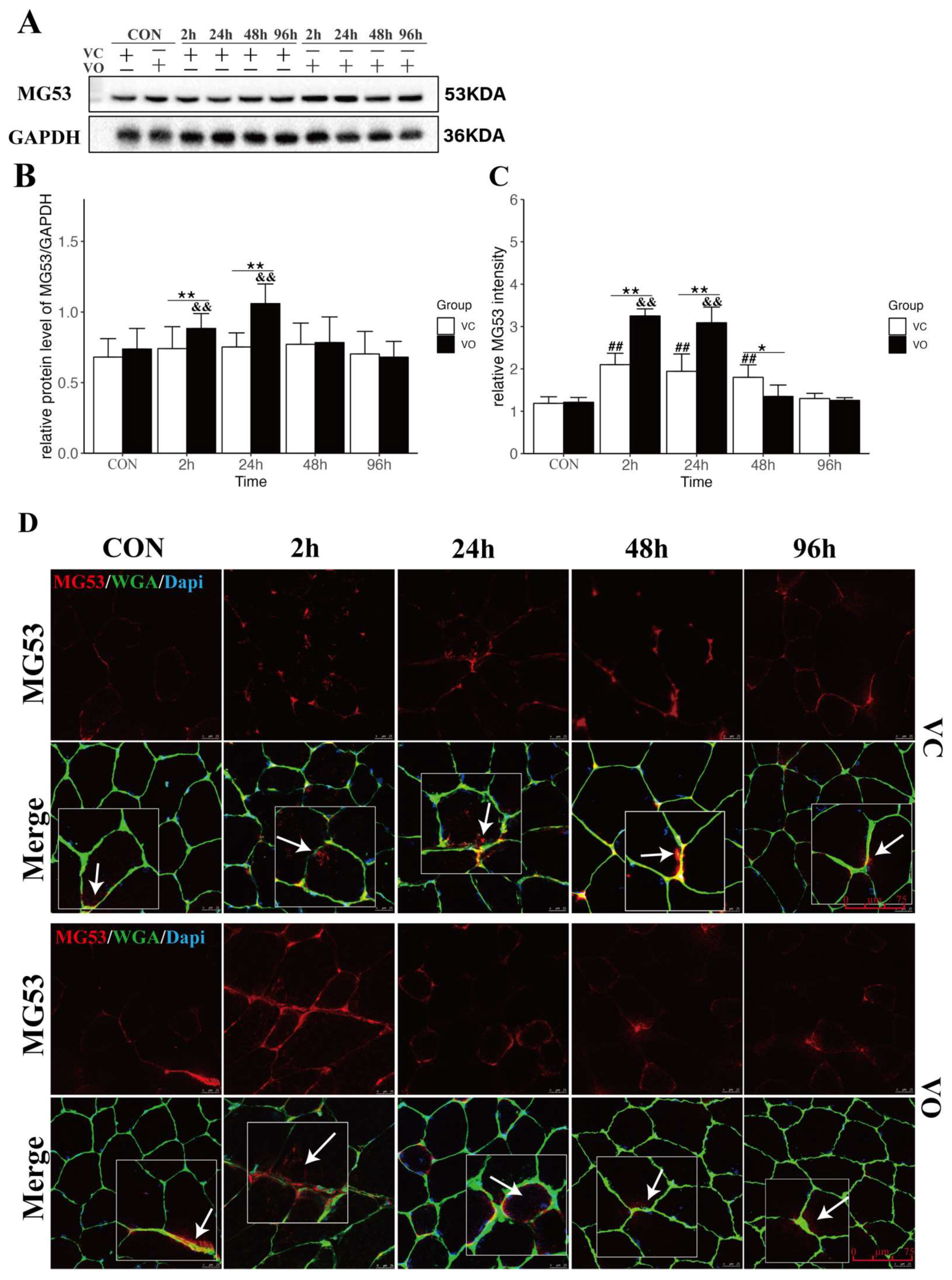
2.3. PTEN Inhibition Upregulates MG53 to Promote Muscle Membrane Repair
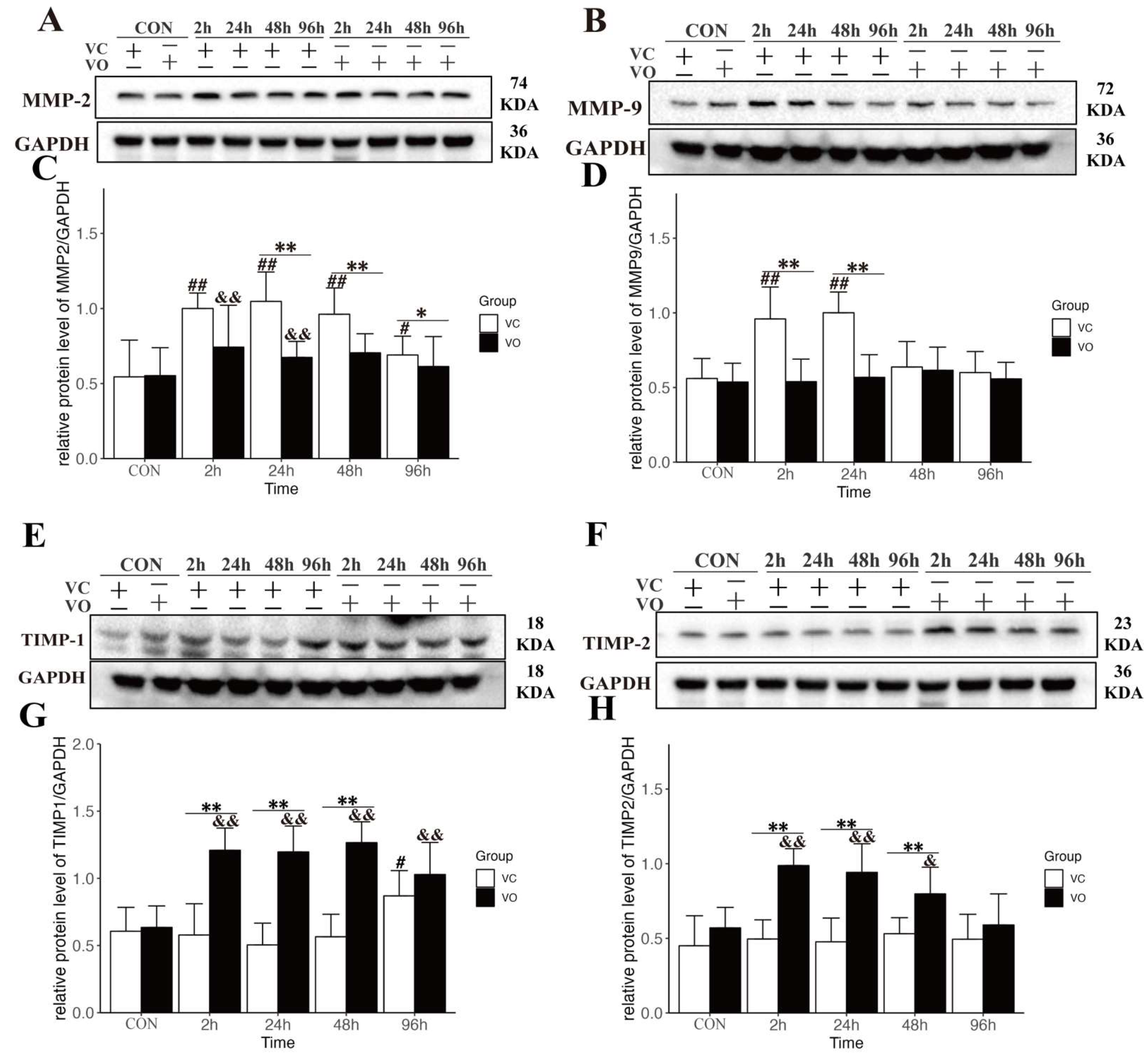
2.4. PTEN Inhibition Regulates the Synthesis and Catabolism of ECM during EIMD
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. EIMD Protocol
4.3. PTEN Inhibitor Treatment
4.4. Animals Experiments
4.5. Whole-Limb Grip Strength Test
4.6. CK and LDH Level Test
4.7. Evans Blue Uptakes
4.8. Histology and Immunofluorescence Staining
4.9. Skeletal Muscle’s ECM Structure as Observed by SEM
4.10. Protein Extraction and Western Blot Analysis
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cheung, K.; Hume, P.; Maxwell, L. Delayed onset muscle soreness: Treatment strategies and performance factors. Sport. Med. 2003, 33, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Buchwald-Werner, S.; Naka, I.; Wilhelm, M.; Schutz, E.; Schoen, C.; Reule, C. Effects of lemon verbena extract (Recoverben(R)) supplementation on muscle strength and recovery after exhaustive exercise: A randomized, placebo-controlled trial. J. Int. Soc. Sport. Nutr. 2018, 15, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hody, S.; Croisier, J.L.; Bury, T.; Rogister, B.; Leprince, P. Eccentric Muscle Contractions: Risks and Benefits. Front. Physiol. 2019, 10, 536. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, G.; Mikkelsen, U.R.; Raastad, T.; Peake, J.M. Leucocytes, cytokines and satellite cells: What role do they play in muscle damage and regeneration following eccentric exercise? Exerc. Immunol. Rev. 2012, 18, 42–97. [Google Scholar] [PubMed]
- Close, G.L.; Ashton, T.; Cable, T.; Doran, D.; Holloway, C.; McArdle, F.; MacLaren, D.P.M. Ascorbic acid supplementation does not attenuate post-exercise muscle soreness following muscle-damaging exercise but may delay the recovery process. Br. J. Nutr. 2006, 95, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Newham, D.J.; Jones, D.A.; Edwards, R.H. Plasma creatine kinase changes after eccentric and concentric contractions. Muscle Nerve 1986, 9, 59–63. [Google Scholar] [CrossRef]
- MacIntyre, D.L.; Reid, W.D.; McKenzie, D.C. Delayed muscle soreness. The inflammatory response to muscle injury and its clinical implications. Sport. Med. 1995, 20, 24–40. [Google Scholar] [CrossRef]
- Stožer, A.; Vodopivc, P.; Križančić Bombek, L. Pathophysiology of exercise-induced muscle damage and its structural, functional, metabolic, and clinical consequences. Physiol. Res. 2020, 69, 565–598. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J. Matrix metalloproteinase and tissue inhibitor of metalloproteinase responses to muscle damage after eccentric exercise. J. Exerc. Rehabil. 2016, 12, 260–265. [Google Scholar] [CrossRef]
- Kjaer, M.; Magnusson, P.; Krogsgaard, M.; Boysen Møller, J.; Olesen, J.; Heinemeier, K.; Hansen, M.; Haraldsson, B.; Koskinen, S.; Esmarck, B.; et al. Extracellular matrix adaptation of tendon and skeletal muscle to exercise. J. Anat. 2006, 208, 445–450. [Google Scholar] [CrossRef]
- Stauber, W.T.; Clarkson, P.M.; Fritz, V.K.; Evans, W.J. Extracellular matrix disruption and pain after eccentric muscle action. J. Appl. Physiol. (Bethesda, Md. 1985) 1990, 69, 868–874. [Google Scholar] [CrossRef]
- Gao, Y.; Kostrominova, T.Y.; Faulkner, J.A.; Wineman, A.S. Age-related changes in the mechanical properties of the epimysium in skeletal muscles of rats. J. Biomech. 2008, 41, 465–469. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Li, Y. Role of matrix metalloproteinases in skeletal muscle: Migration, differentiation, regeneration and fibrosis. Cell Adh. Migr. 2009, 3, 337–341. [Google Scholar] [CrossRef]
- Andrews, N.W.; Corrotte, M. Plasma membrane repair. Curr. Biol. 2018, 28, R392–R397. [Google Scholar] [CrossRef] [Green Version]
- Koskinen, S.O.A.; Ahtikoski, A.M.; Komulainen, J.; Hesselink, M.K.C.; Drost, M.R.; Takala, T.E.S. Short-term effects of forced eccentric contractions on collagen synthesis and degradation in rat skeletal muscle. Pflug. Arch. 2002, 444, 59–72. [Google Scholar] [CrossRef]
- Cooper, S.T.; McNeil, P.L. Membrane Repair: Mechanisms and Pathophysiology. Physiol. Rev. 2015, 95, 1205–1240. [Google Scholar] [CrossRef] [Green Version]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Sun, J.; Wang, Q.; Du, Y.; Cheng, J.; Yi, J.; Xie, B.; Jin, S.; Chen, G.; Wang, L.; et al. Systemic Deficiency of PTEN Accelerates Breast Cancer Growth and Metastasis. Front. Oncol. 2022, 12, 825484. [Google Scholar] [CrossRef]
- Yue, F.; Bi, P.; Wang, C.; Li, J.; Liu, X.; Kuang, S. Conditional Loss of Pten in Myogenic Progenitors Leads to Postnatal Skeletal Muscle Hypertrophy but Age-Dependent Exhaustion of Satellite Cells. Cell Rep. 2016, 17, 2340–2353. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Zhou, L.; Wu, X.; Li, R.; Wen, J.; Sha, J.; Wen, X. The PI3K/AKT pathway in the pathogenesis of prostate cancer. Front. Biosci. (Landmark Ed.) 2016, 21, 1084–1091. [Google Scholar]
- Hameed, J.S.F.; Devarajan, A.; M, S.D.; Bhattacharyya, A.; Shirude, M.B.; Dutta, D.; Karmakar, P.; Mukherjee, A. PTEN-negative endometrial cancer cells protect their genome through enhanced DDB2 expression associated with augmented nucleotide excision repair. BMC Cancer 2023, 23, 399. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Liu, L.; Tian, Y. miR-19-3p Targets PTEN to Regulate Cervical Cancer Cell Proliferation, Invasion, and Autophagy. Genet. Res. 2023, 2023, 4784500. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cheng, Z.; Haifeng, Y.; Chen, M.; Li, L. PTEN signaling inhibitor VO-OHpic improves cardiac myocyte survival by mediating apoptosis resistance in vitro. Biomed. Pharmacother. 2018, 103, 1217–1222. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Song, C.; Huang, D.; Narayanan, N.; Qiu, J.; Jia, Z.; Yuan, Z.; Oprescu, S.N.; Roseguini, B.T.; Deng, M.; et al. PTEN Inhibition Ameliorates Muscle Degeneration and Improves Muscle Function in a Mouse Model of Duchenne Muscular Dystrophy. Mol. Ther. J. Am. Soc. Gene Ther. 2021, 29, 132–148. [Google Scholar] [CrossRef] [PubMed]
- Suntar, I.; Sureda, A.; Belwal, T.; Sanches Silva, A.; Vacca, R.A.; Tewari, D.; Sobarzo-Sánchez, E.; Nabavi, S.F.; Shirooie, S.; Dehpour, A.R.; et al. Natural products, PGC-1 α, and Duchenne muscular dystrophy. Acta Pharm. Sinica. B 2020, 10, 734–745. [Google Scholar] [CrossRef]
- Tidball, J.G. Mechanisms of muscle injury, repair, and regeneration. Compr. Physiol. 2011, 1, 2029–2062. [Google Scholar] [CrossRef]
- Pulido, R. PTEN Inhibition in Human Disease Therapy. Molecules 2018, 23, 285. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, N.M.; Mueller, C.; Grossmann, T.N. The Therapeutic Potential of PTEN Modulation: Targeting Strategies from Gene to Protein. Cell Chem. Biol. 2018, 25, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Zeng, J.; Liu, X.; Lai, J.; Xu, J. Exercise-Induced Muscle Damage and Protein Intake: A Bibliometric and Visual Analysis. Nutrients 2022, 14, 4288. [Google Scholar] [CrossRef]
- Ebstrup, M.L.; Dias, C.; Heitmann, A.S.B.; Sønder, S.L.; Nylandsted, J. Actin Cytoskeletal Dynamics in Single-Cell Wound Repair. Int. J. Mol. Sci. 2021, 22, 10886. [Google Scholar] [CrossRef]
- Nascimento, D.d.C.; Navalta, J.W.; Durigan, J.L.Q.; Marqueti, R.d.C.; Tibana, R.A.; Luiz Franco, O.; de Almeida, J.A.; Camarço, N.F.; Neto, I.V.d.S.; Prestes, J. Acute eccentric resistance exercise decreases matrix metalloproteinase activity in obese elderly women. Clin. Physiol. Funct. Imaging 2016, 36, 139–145. [Google Scholar] [CrossRef]
- Bi, X.; Lv, X.; Liu, D.; Guo, H.; Yao, G.; Wang, L.; Liang, X.; Yang, Y. METTL3-mediated maturation of miR-126-5p promotes ovarian cancer progression via PTEN-mediated PI3K/Akt/mTOR pathway. Cancer Gene 2021, 28, 335–349. [Google Scholar] [CrossRef]
- Alzahrani, A.S. PI3K/Akt/mTOR inhibitors in cancer: At the bench and bedside. Semin Cancer Biol. 2019, 59, 125–132. [Google Scholar] [CrossRef]
- Yao, X.; Yu, S.; Jing, X.; Guo, J.; Sun, K.; Guo, F.; Ye, Y. PTEN inhibitor VO-OHpic attenuates GC-associated endothelial progenitor cell dysfunction and osteonecrosis of the femoral head via activating Nrf2 signaling and inhibiting mitochondrial apoptosis pathway. Stem Cell Res. Ther. 2020, 11, 140. [Google Scholar] [CrossRef]
- Lin, Y.; Guo, W.; Chen, K.W.; Xiao, Z.M. VO-OHpic attenuates intervertebral disc degeneration via PTEN/Akt pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 2811–2819. [Google Scholar] [CrossRef]
- Zhuang, C.; Guo, Z.; Zhu, J.; Wang, W.; Sun, R.; Qi, M.; Wang, Q.; Fan, X.; Ma, R.; Yu, J. PTEN inhibitor attenuates cardiac fibrosis by regulating the M2 macrophage phenotype via the PI3K/AKT/TGF-β/Smad 2/3 signaling pathway. Int. J. Cardiol. 2022, 356, 88–96. [Google Scholar] [CrossRef]
- Mocanu, M.M.; Yellon, D.M. PTEN, the Achilles’ heel of myocardial ischaemia/reperfusion injury? Br. J. Pharmacol. 2007, 150, 833–838. [Google Scholar] [CrossRef]
- Schwartzbauer, G.; Robbins, J. The tumor suppressor gene PTEN can regulate cardiac hypertrophy and survival. J. Biol. Chem. 2001, 276, 35786–35793. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.A.; Singla, D.K. PTEN inhibitor VO-OHpic attenuates inflammatory M1 macrophages and cardiac remodeling in doxorubicin-induced cardiomyopathy. Am. J. Physiology. Heart Circ. Physiol. 2018, 315, H1236–H1249. [Google Scholar] [CrossRef]
- Mak, L.H.; Vilar, R.; Woscholski, R. Characterisation of the PTEN inhibitor VO-OHpic. J. Chem. Biol. 2010, 3, 157–163. [Google Scholar] [CrossRef] [Green Version]
- Cai, C.; Masumiya, H.; Weisleder, N.; Matsuda, N.; Nishi, M.; Hwang, M.; Ko, J.K.; Lin, P.; Thornton, A.; Zhao, X.; et al. MG53 nucleates assembly of cell membrane repair machinery. Nat. Cell Biol. 2009, 11, 56–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, C.; Weisleder, N.; Ko, J.-K.; Komazaki, S.; Sunada, Y.; Nishi, M.; Takeshima, H.; Ma, J. Membrane repair defects in muscular dystrophy are linked to altered interaction between MG53, caveolin-3, and dysferlin. J. Biol. Chem. 2009, 284, 15894–15902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gushchina, L.V.; Bhattacharya, S.; McElhanon, K.E.; Choi, J.H.; Manring, H.; Beck, E.X.; Alloush, J.; Weisleder, N. Treatment with Recombinant Human MG53 Protein Increases Membrane Integrity in a Mouse Model of Limb Girdle Muscular Dystrophy 2B. Mol. Ther. 2017, 25, 2360–2371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gumpper-Fedus, K.; Park, K.H.; Ma, H.; Zhou, X.; Bian, Z.; Krishnamurthy, K.; Sermersheim, M.; Zhou, J.; Tan, T.; Li, L.; et al. MG53 preserves mitochondrial integrity of cardiomyocytes during ischemia reperfusion-induced oxidative stress. Redox Biol. 2022, 54, 102357. [Google Scholar] [CrossRef]
- Han, Y.; Black, S.; Gong, Z.; Chen, Z.; Ko, J.K.; Zhou, Z.; Xia, T.; Fang, D.; Yang, D.; Gu, D.; et al. Membrane-delimited signaling and cytosolic action of MG53 preserve hepatocyte integrity during drug-induced liver injury. J. Hepatol. 2022, 76, 558–567. [Google Scholar] [CrossRef]
- Jia, Y.; Chen, K.; Lin, P.; Lieber, G.; Nishi, M.; Yan, R.; Wang, Z.; Yao, Y.; Li, Y.; Whitson, B.A.; et al. Treatment of acute lung injury by targeting MG53-mediated cell membrane repair. Nat. Commun. 2014, 5, 4387. [Google Scholar] [CrossRef] [Green Version]
- Duann, P.; Li, H.; Lin, P.; Tan, T.; Wang, Z.; Chen, K.; Zhou, X.; Gumpper, K.; Zhu, H.; Ludwig, T.; et al. MG53-mediated cell membrane repair protects against acute kidney injury. Sci. Transl. Med. 2015, 7, 279ra236. [Google Scholar] [CrossRef] [Green Version]
- Rønning, S.B.; Carlson, C.R.; Aronsen, J.M.; Pisconti, A.; Høst, V.; Lunde, M.; Liland, K.H.; Sjaastad, I.; Kolset, S.O.; Christensen, G.; et al. Syndecan-4-/- Mice Have Smaller Muscle Fibers, Increased Akt/mTOR/S6K1 and Notch/HES-1 Pathways, and Alterations in Extracellular Matrix Components. Front. Cell Dev. Biol. 2020, 8, 730. [Google Scholar] [CrossRef]
- Csapo, R.; Gumpenberger, M.; Wessner, B. Skeletal Muscle Extracellular Matrix—What Do We Know About Its Composition, Regulation, and Physiological Roles? A Narrative Review. Front. Physiol. 2020, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Hotfiel, T.; Freiwald, J.; Hoppe, M.W.; Lutter, C.; Forst, R.; Grim, C.; Bloch, W.; Hüttel, M.; Heiss, R. Advances in Delayed-Onset Muscle Soreness (DOMS): Part I: Pathogenesis and Diagnostics. Sport. Sportschaden 2018, 32, 243–250. [Google Scholar] [CrossRef]
- Alameddine, H.S. Matrix metalloproteinases in skeletal muscles: Friends or foes? Neurobiol. Dis. 2012, 48, 508–518. [Google Scholar] [CrossRef]
- Armstrong, R.B.; Ogilvie, R.W.; Schwane, J.A. Eccentric exercise-induced injury to rat skeletal muscle. J. Appl. Physiol. Respir. Env. Exerc. Physiol. 1983, 54, 80–93. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Heart | Left Kidney | Right Kidney | Liver | Spleen |
|---|---|---|---|---|---|
| VO | 0.167 ± 0.013 | 0.174 ± 0.01 | 0.174 ± 0.011 | 1.469 ± 0.094 | 0.081 ± 0.018 |
| VC | 0.169 ± 0.01 | 0.173 ± 0.01 | 0.172 ± 0.011 | 1.475 ± 0.062 | 0.079 ± 0.018 |
| Pancreas | Body weight | Soleus | Gastrocnemius | Quadriceps | |
| VO | 0.155 ± 0.33 | 27.94 ± 2.68 | 0.017 ± 0.003 | 0.156 ± 0.013 | 0.239 ± 0.17 |
| VC | 0.156 ± 0.21 | 27.81 ± 2.65 | 0.016 ± 0.004 | 0.153 ± 0.077 | 0.24 ± 0.023 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, B.; Shi, L.; Wu, Y. PTEN Inhibitor Treatment Lowers Muscle Plasma Membrane Damage and Enhances Muscle ECM Homeostasis after High-Intensity Eccentric Exercise in Mice. Int. J. Mol. Sci. 2023, 24, 9954. https://doi.org/10.3390/ijms24129954
Wu B, Shi L, Wu Y. PTEN Inhibitor Treatment Lowers Muscle Plasma Membrane Damage and Enhances Muscle ECM Homeostasis after High-Intensity Eccentric Exercise in Mice. International Journal of Molecular Sciences. 2023; 24(12):9954. https://doi.org/10.3390/ijms24129954
Chicago/Turabian StyleWu, Baile, Lijun Shi, and Ying Wu. 2023. "PTEN Inhibitor Treatment Lowers Muscle Plasma Membrane Damage and Enhances Muscle ECM Homeostasis after High-Intensity Eccentric Exercise in Mice" International Journal of Molecular Sciences 24, no. 12: 9954. https://doi.org/10.3390/ijms24129954