
Genome-Wide Analysis of the PHT Gene Family and Its Response to Mycorrhizal Symbiosis in Tomatoes under Phosphate Starvation Conditions

Abstract
:1. Introduction
2. Results
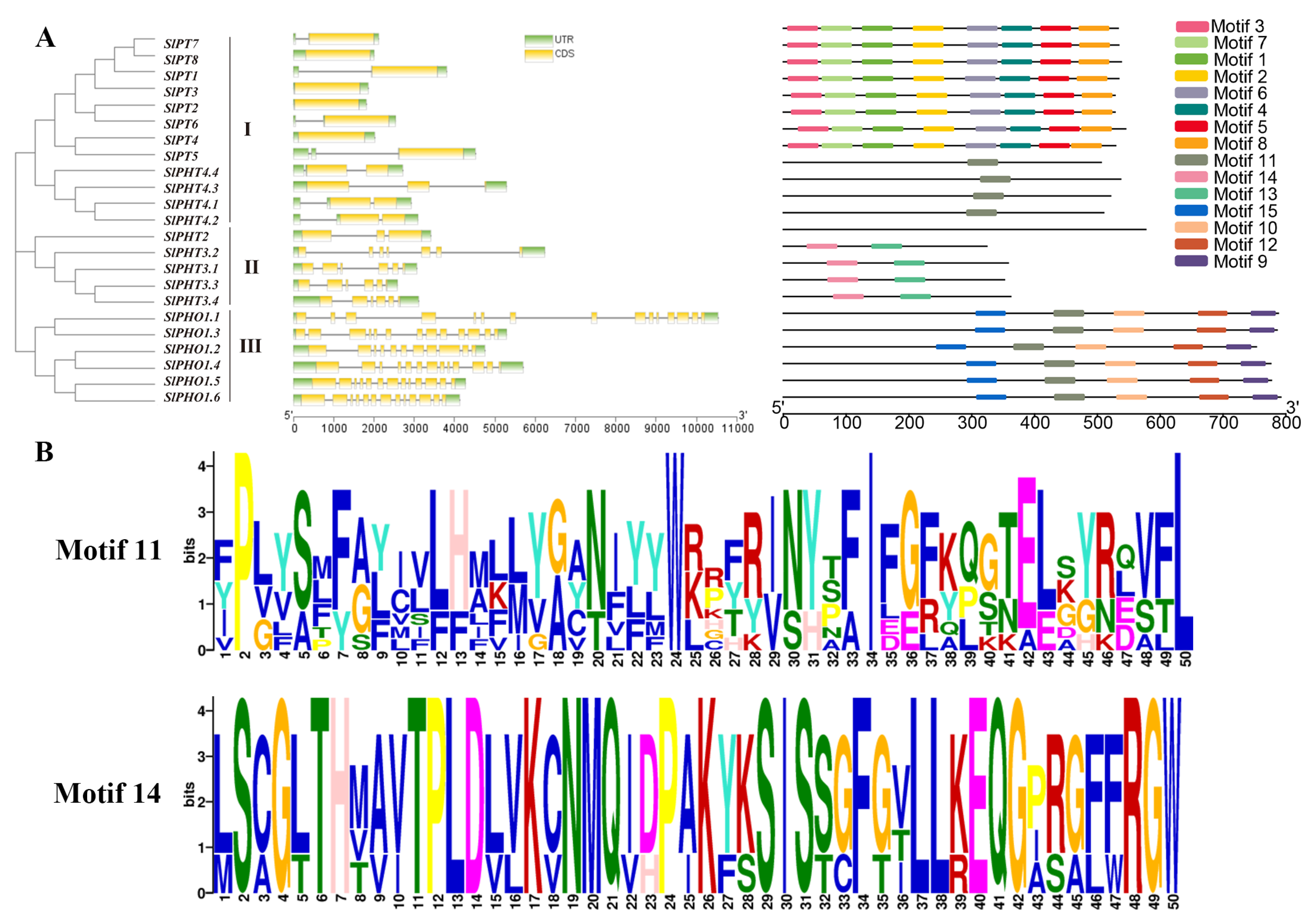
2.1. Identification and Phylogenetic Classification of the SlPHT Gene Family
2.2. Gene Structure and Protein Domain Analysis of SlPHT Members
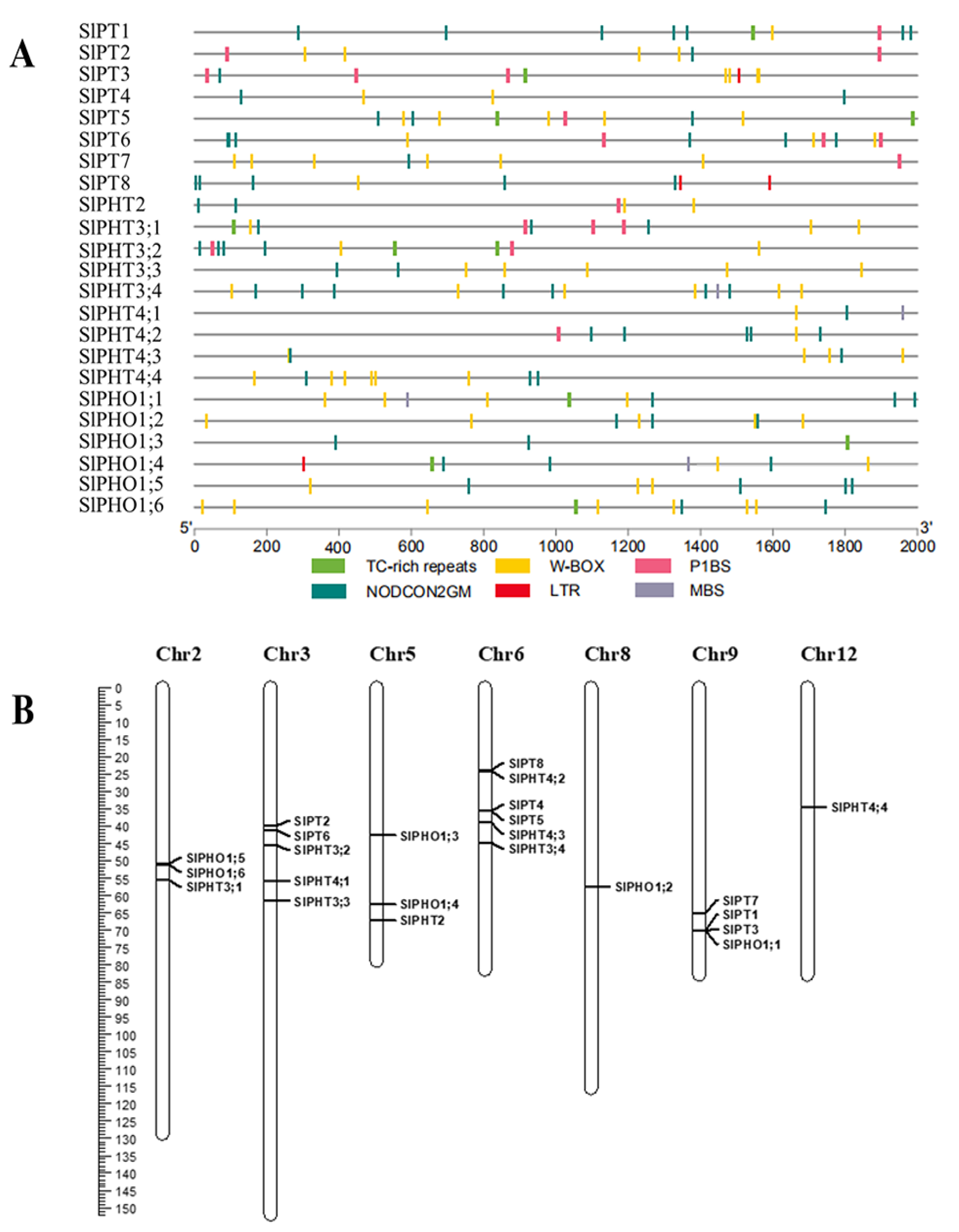
2.3. Cis-Acting Element and Chromosomal Distribution
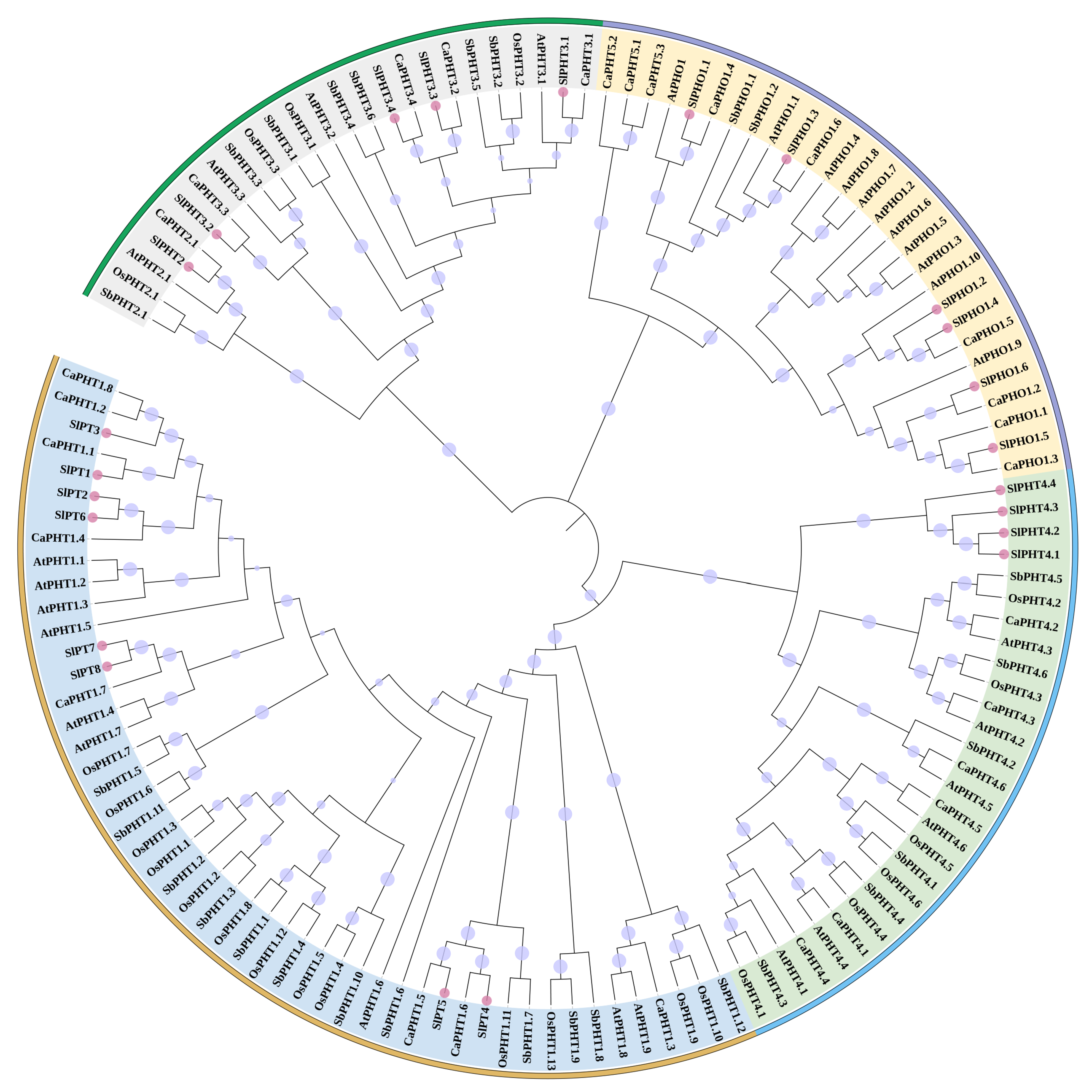
2.4. Phylogenetic Analysis
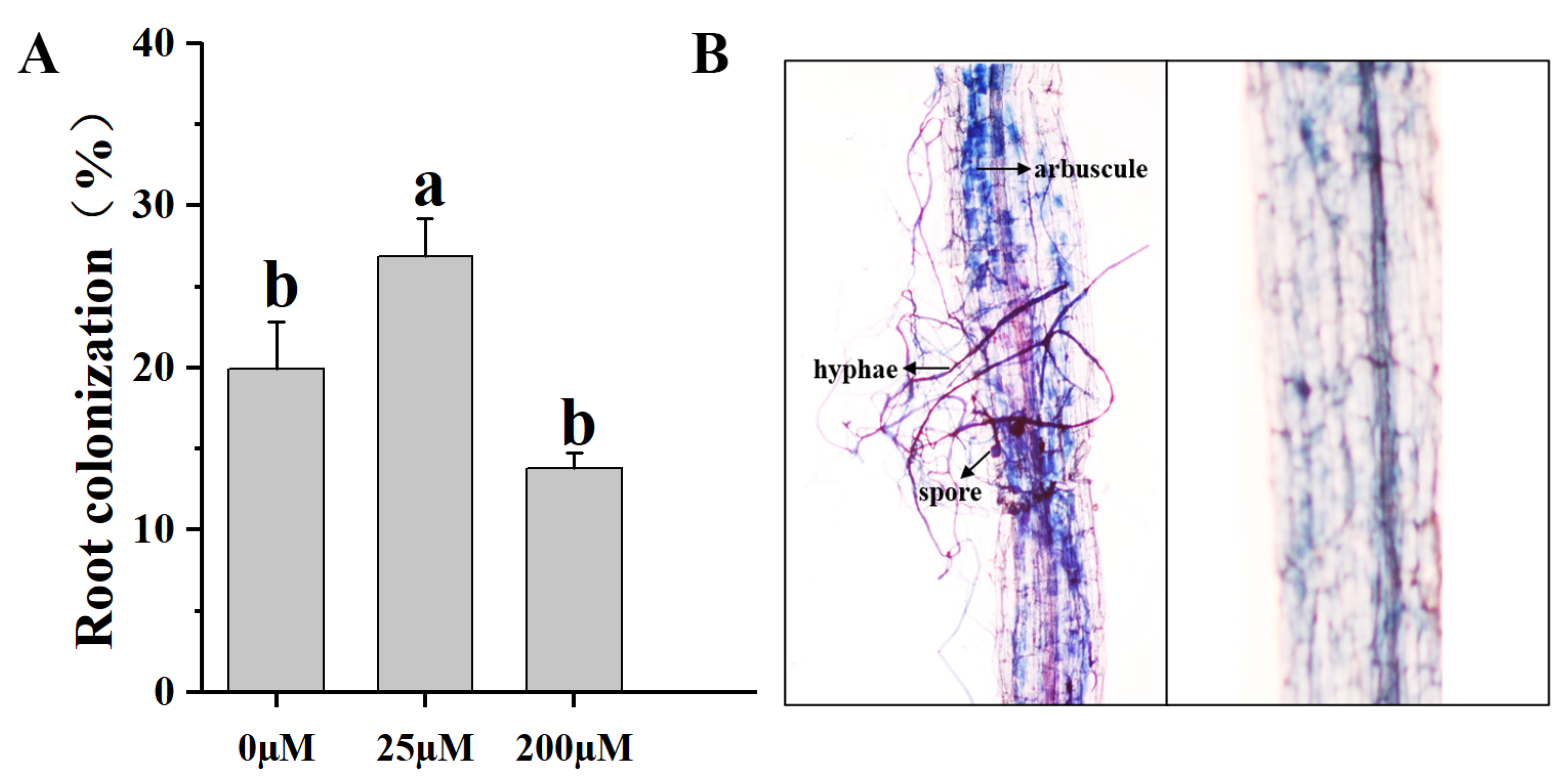
2.5. Mycorrhizal Colonization
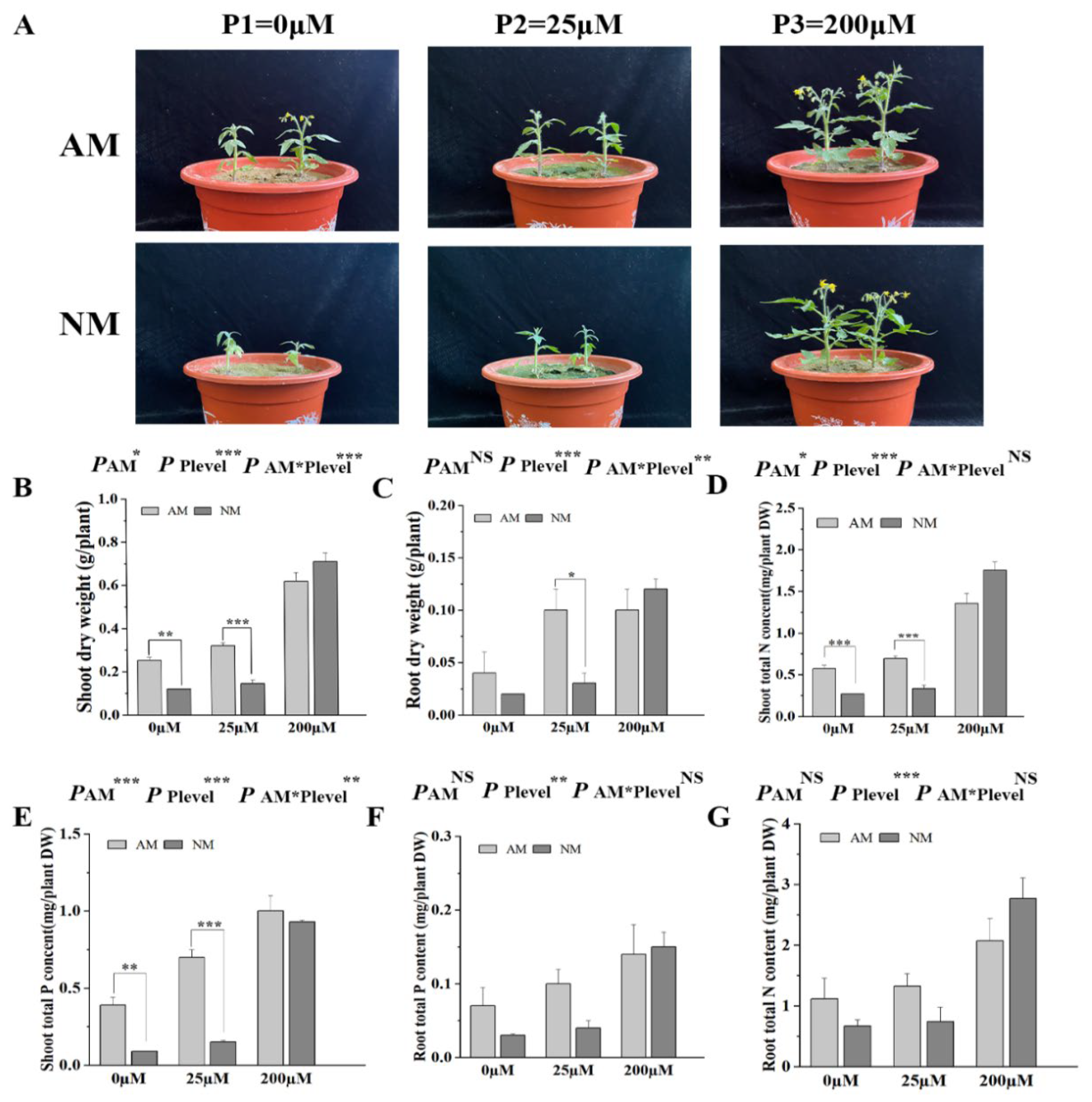
2.6. AM Fungal Colonization Promotes Tomato Growth and Nutrient Uptake
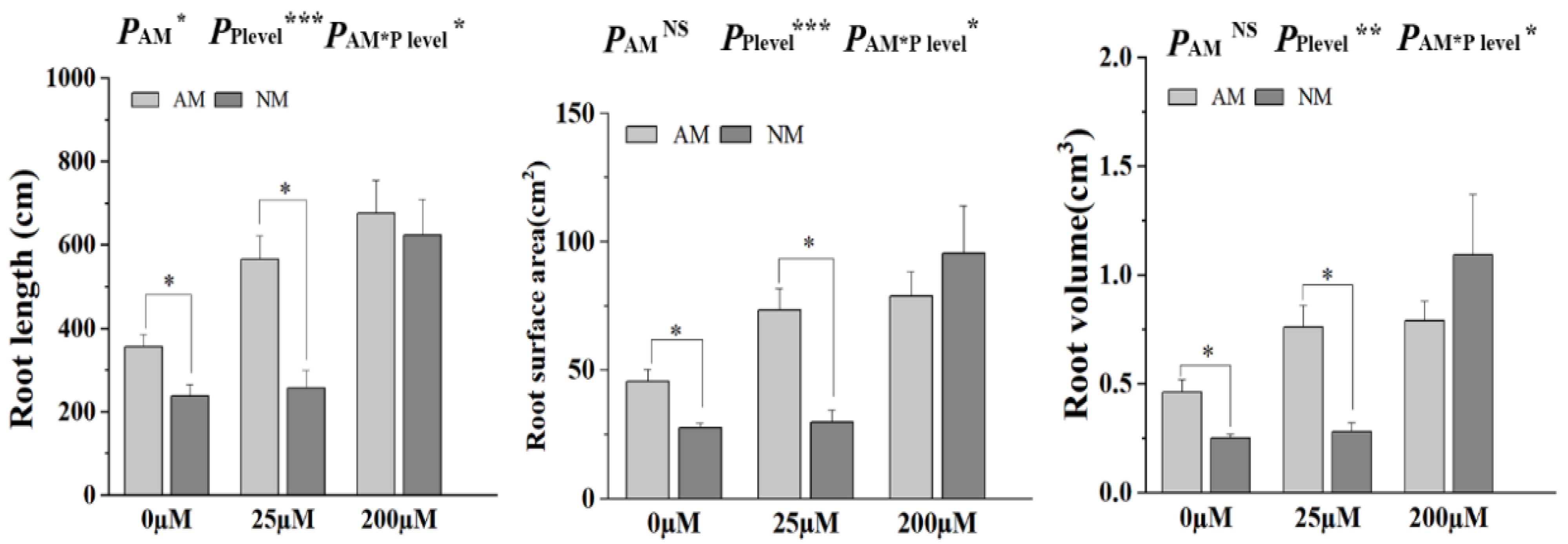
2.7. AM Fungal Colonization Promotes Tomato Root Morphological Plasticity
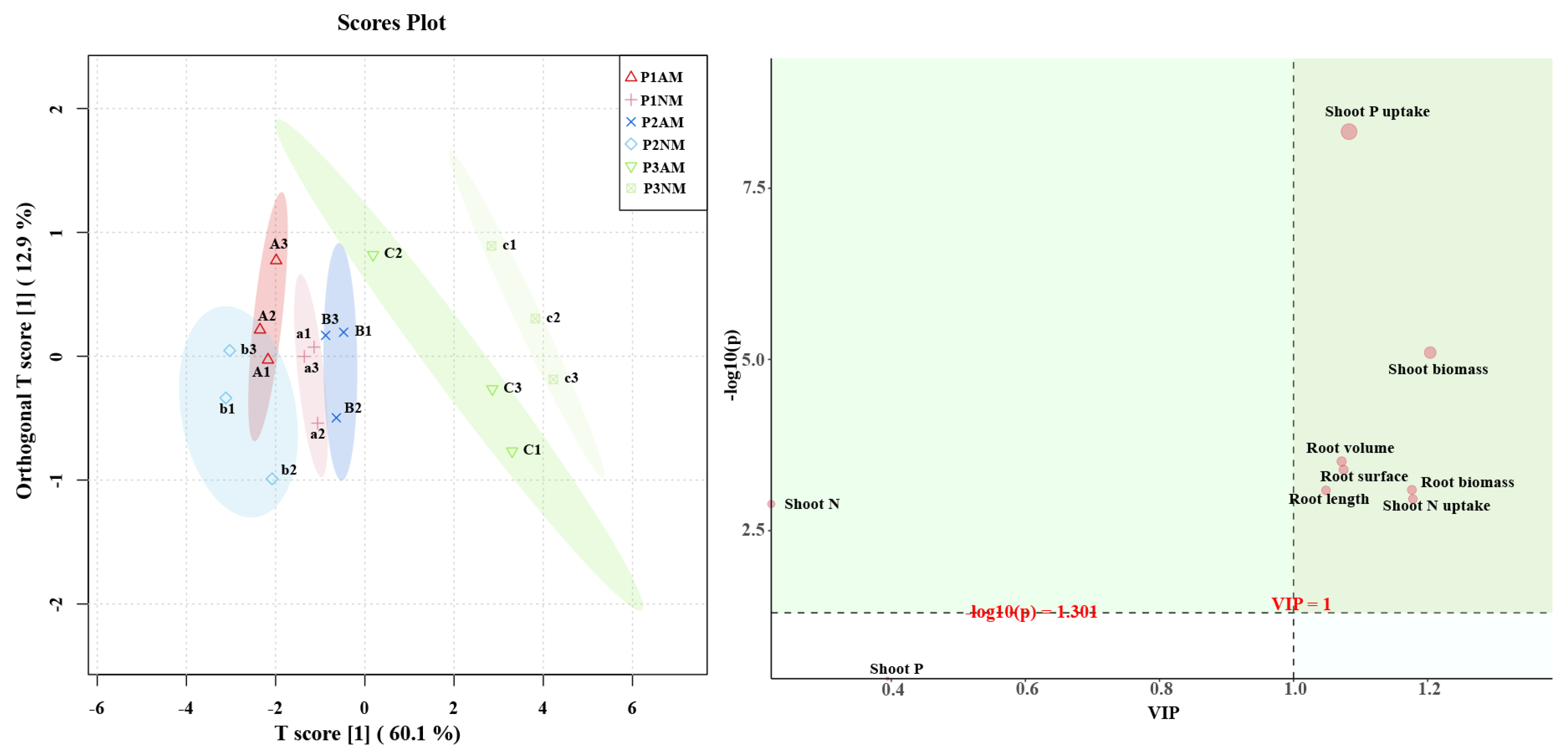
2.8. Orthogonal Partial Least-Squares Discrimination Analysis (OPLS-DA) of Physiological Indicators
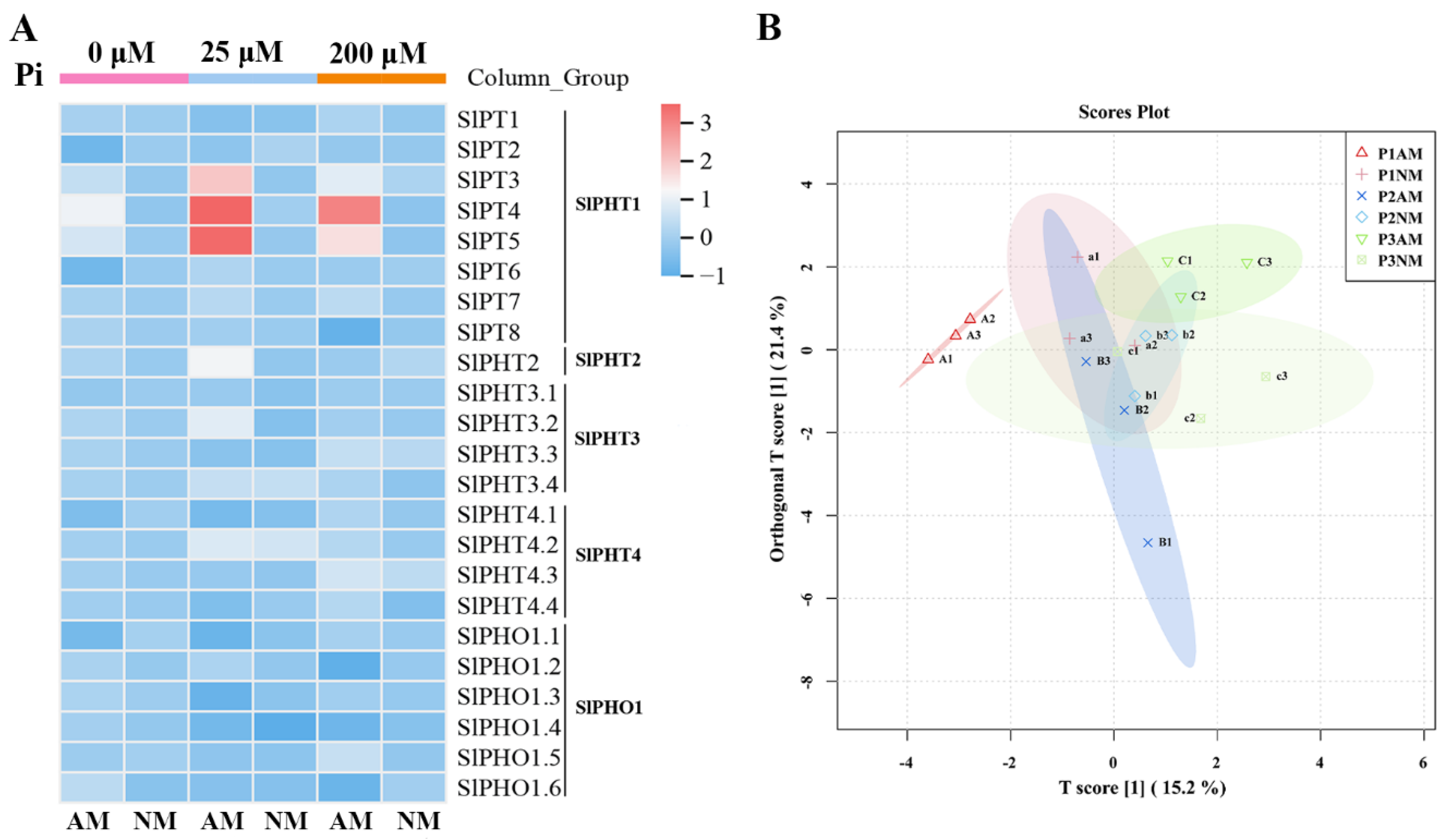
2.9. The Expression Patterns of SlPHT Genes in Response to AM and Pi Concentrations
3. Discussion
3.1. Identification and Characterization of SlPHT Genes in Tomato
3.2. Choice and Validation of P Supply Conditions Affecting AM Symbiosis
3.3. SlPHT Responses to Pi Availability and AM Fungi Inoculation
4. Materials and Methods
4.1. Plant Material and AM Fungi Inoculation
4.2. Experimental Design
4.3. Identification of Phosphate Transporter (PHT) Genes in the Tomato Genome
4.4. Conserved Motif, Gene Structure, and Chromosomal Mapping
4.5. Phylogenetic and Cis-Element Analysis
4.6. Real-Time Quantitative Polymerase Chain Reaction (RT qPCR)
4.7. Mycorrhizal Colonization Rates
4.8. Determination of the Concentrations of N and P and Root System Parameters
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wu, P.; Shou, H.X.; Xu, G.H.; Lian, X.M. Improvement of phosphorus efficiency in rice on the basis of understanding phosphate signaling and homeostasis. Curr. Opin. Plant Biol. 2013, 16, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo-Orts, L.; Couto, D.; Hothorn, M. Identity and functions of inorganic and inositol polyphosphates in plants. New Phytol. 2020, 225, 637–652. [Google Scholar] [CrossRef] [Green Version]
- Lhamo, D.; Shao, Q.L.; Tang, R.J.; Luan, S. Genome-Wide Analysis of the Five Phosphate Transporter Families in Camelina sativa and Their Expressions in Response to Low-P. Int. J. Mol. Sci. 2020, 21, 8365. [Google Scholar] [CrossRef]
- Fan, X.N.; Che, X.R.; Lai, W.Z.; Wang, S.J.; Hu, W.T.; Chen, H.; Zhao, B.; Tang, M.; Xie, X.A. The auxin-inducible phosphate transporter AsPT5 mediates phosphate transport and is indispensable for arbuscule formation in Chinese milk vetch at moderately high phosphate supply. Environ. Microbiol. 2020, 22, 2053–2079. [Google Scholar] [CrossRef]
- Shu, B.; Xia, R.X.; Wang, P. Differential regulation of Pht1 phosphate transporters from trifoliate orange (Poncirus trifoliata L. Raf) seedlings. Sci. Hortic. 2012, 146, 115–123. [Google Scholar] [CrossRef]
- Lang, M.; Li, X.; Zheng, C.Y.; Li, H.G.; Zhang, J.L. Shading mediates the response of mycorrhizal maize (Zea mays L.) seedlings under varying levels of phosphorus. Appl. Soil Ecol. 2021, 166, 104060. [Google Scholar] [CrossRef]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2008; pp. 1–787. [Google Scholar]
- Nagy, R.; Karandashov, V.; Chague, W.; Kalinkevich, K.; Tamasloukht, M.; Xu, G.H.; Jakobsen, I.; Levy, A.A.; Amrhein, N.; Bucher, M. The characterization of novel mycorrhiza-specific phosphate transporters from Lycopersicon esculentum and Solanum tuberosum uncovers functional redundancy in symbiotic phosphate transport in solanaceous species. Plant J. 2005, 42, 236–250. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.H.; Huang, S.C.; He, R.; Li, Y.Z.; Cheng, X.G. Transgenic Rice Overexperessing a Tomato Mitochondrial Phosphate Transporter, SlMPT3;1, Promotes Phosphate Uptake and Increases Grain Yield. J. Plant Biol. 2018, 61, 383–400. [Google Scholar] [CrossRef]
- Nagy, R.; Vasconcelos, M.J.V.; Zhao, S.; McElver, J.; Bruce, W.; Amrhein, N.; Raghothama, K.G.; Bucher, M. Differential regulation of five Pht1 phosphate transporters from maize (Zea mays L.). Plant Biol. 2006, 8, 186–197. [Google Scholar] [CrossRef]
- Bunya, M.; Nishimura, M.; Harashima, S.; Oshima, Y. The PHO84 gene of Saccharomyces-cerevisiae encodes an inorganic-phosphate transporter. Mol. Cell. Biol. 1991, 11, 3229–3238. [Google Scholar] [CrossRef] [Green Version]
- Guo, B.; Jin, Y.; Wussler, C.; Blancaflor, E.B.; Motes, C.M.; Versaw, W.K. Functional analysis of the Arabidopsis PHT4 family of intracellular phosphate transporters. New Phytol. 2008, 177, 889–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muchhal, U.S.; Pardo, J.M.; Raghothama, K.G. Phosphate transporters from the higher plant Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 10519–10523. [Google Scholar] [CrossRef] [Green Version]
- Chen, A.Q.; Chen, X.; Wang, H.M.; Liao, D.H.; Gu, M.; Qu, H.Y.; Sun, S.B.; Xu, G.H. Genome-wide investigation and expression analysis suggest diverse roles and genetic redundancy of Pht1 family genes in response to Pi deficiency in tomato. BMC Plant Biol. 2014, 14, 15. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Xu, Y.; Jiang, H.; Jiang, C.; Du, Y.; Gong, C.; Wang, W.; Zhu, S.; Han, G.; Cheng, B. Systematic Identification, Evolution and Expression Analysis of the Zea mays PHT1 Gene Family Reveals Several New Members Involved in Root Colonization by Arbuscular Mycorrhizal Fungi. Int. J. Mol. Sci. 2016, 17, 930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Hu, L.Z.; Yu, D.S.; Xu, K.D.; Zhang, J.; Li, X.L.; Wang, P.F.; Chen, G.; Liu, Z.H.; Peng, C.F.; et al. Integrative Analysis of the Wheat PHT1 Gene Family Reveals a Novel Member Involved in Arbuscular Mycorrhizal Phosphate Transport and Immunity. Cells 2019, 8, 490. [Google Scholar] [CrossRef] [Green Version]
- Teng, W.; Zhao, Y.Y.; Zhao, X.Q.; He, X.; Ma, W.Y.; Deng, Y.; Chen, X.P.; Tong, Y.P. Genome-wide Identification, Characterization, and Expression Analysis of PHT1 Phosphate Transporters in Wheat. Front. Plant Sci. 2017, 8, 543. [Google Scholar] [CrossRef] [Green Version]
- Ceasar, S.A.; Hodge, A.; Baker, A.; Baldwin, S.A. Phosphate Concentration and Arbuscular Mycorrhizal Colonisation Influence the Growth, Yield and Expression of Twelve PHT1 Family Phosphate Transporters in Foxtail Millet (Setaria italica). PLoS ONE 2014, 9, e108459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faraji, S.; Ahmadizadeh, M.; Heidari, P. Genome-wide comparative analysis of Mg transporter gene family between Triticum turgidum and Camelina sativa. Biometals 2021, 34, 639–660. [Google Scholar] [CrossRef]
- Paszkowski, U.; Kroken, S.; Roux, C.; Briggs, S.P. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 13324–13329. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, I.; Rawoof, A.; Islam, K.; Momo, J.; Ramchiary, N. Identification and expression analysis of phosphate transporter genes and metabolites in response to phosphate stress in Capsicum annuum. Environ. Exp. Bot. 2021, 190, 104597. [Google Scholar] [CrossRef]
- Li, Y.; Wang, X.; Zhang, H.; Wang, S.L.; Ye, X.S.; Shi, L.; Xu, F.S.; Ding, G.D. Molecular identification of the phosphate transporter family 1 (PHT1) genes and their expression profiles in response to phosphorus deprivation and other abiotic stresses in Brassica napus. PLoS ONE 2019, 14, e0220374. [Google Scholar] [CrossRef] [Green Version]
- Murugan, N.; Palanisamy, V.; Channappa, M.; Ramanathan, V.; Ramaswamy, M.; Govindakurup, H.; Chinnaswamy, A. Genome-Wide In Silico Identification, Structural Analysis, Promoter Analysis, and Expression Profiling of PHT Gene Family in Sugarcane Root under Salinity Stress. Sustainability 2022, 14, 15893. [Google Scholar] [CrossRef]
- Rausch, C.; Daram, P.; Brunner, S.; Jansa, J.; Laloi, M.; Leggewie, G.; Amrhein, N.; Bucher, M. A phosphate transporter expressed in arbuscule-containing cells in potato. Nature 2001, 414, 462–466. [Google Scholar] [CrossRef]
- Harrison, M.J.; Dewbre, G.R.; Liu, J.Y. A phosphate transporter from Medicago truncatula involved in the acquisiton of phosphate released by arbuscular mycorrhizal fungi. Plant Cell 2002, 14, 2413–2429. [Google Scholar] [CrossRef] [Green Version]
- Kobae, Y.; Hata, S. Dynamics of Periarbuscular Membranes Visualized with a Fluorescent Phosphate Transporter in Arbuscular Mycorrhizal Roots of Rice. Plant Cell Physiol. 2010, 51, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Volpe, V.; Giovannetti, M.; Sun, X.G.; Fiorilli, V.; Bonfante, P. The phosphate transporters LjPT4 and MtPT4 mediate early root responses to phosphate status in non mycorrhizal roots. Plant Cell Environ. 2016, 39, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Wegmueller, S.; Svistoonoff, S.; Reinhardt, D.; Stuurman, J.; Amrhein, N.; Bucher, M. A transgenic dTph1 insertional mutagenesis system for forward genetics in mycorrhizal phosphate transport of Petunia. Plant J. 2008, 54, 1115–1127. [Google Scholar] [CrossRef] [Green Version]
- Versaw, W.K.; Harrison, M.J. A chloroplast phosphate transporter, PHT2;1, influences allocation of phosphate within the plant and phosphate-starvation responses. Plant Cell 2002, 14, 1751–1766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javot, H.; Pumplin, N.; Harrison, M.J. Phosphate in the arbuscular mycorrhizal symbiosis: Transport properties and regulatory roles. Plant Cell Environ. 2007, 30, 310–322. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.Y.; Wang, Z.; Xia, J.C.; Shen, S.L.; Guan, M.W.; Zhu, M.C.; Qiao, C.L.; Sun, F.J.; Liang, Y.; Li, J.; et al. Genome-Wide Analysis of Phosphorus Transporter Genes in Brassica and Their Roles in Heavy Metal Stress Tolerance. Int. J. Mol. Sci. 2020, 21, 2209. [Google Scholar] [CrossRef] [Green Version]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar]
- Szentpeteri, V.; Mayer, Z.; Posta, K. Mycorrhizal symbiosis-induced abiotic stress mitigation through phosphate transporters in Solanum lycopersicum L. Plant Growth Regul. 2022, 99, 265–281. [Google Scholar] [CrossRef]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with trifolium-subterraneum L.1. spread of hyphae and phosphorus inflow into roots. New Phytol. 1992, 120, 371–380. [Google Scholar] [CrossRef]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular arbuscular mycorrhizal fungi associated with trifolium-subterraneum L. 2. Hyphal transport of P-32 over defined distances. New Phytol. 1992, 120, 509–516. [Google Scholar] [CrossRef]
- Karandashov, V.; Bucher, M. Symbiotic phosphate transport in arbuscular mycorrhizas. Trends Plant Sci. 2005, 10, 22–29. [Google Scholar] [CrossRef]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Pochon, N.; Ayadi, A.; Nakanishi, T.M.; Thibaud, M.C. Phosphate import in plants: Focus on the PHT1 transporters. Front. Plant Sci. 2011, 2, 83. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.Y.; Gronlund, M.; Jakobsen, I.; Grotemeyer, M.S.; Rentsch, D.; Miyao, A.; Hirochika, H.; Kumar, C.S.; Sundaresan, V.; Salamin, N.; et al. Nonredundant Regulation of Rice Arbuscular Mycorrhizal Symbiosis by Two Members of the PHOSPHATE TRANSPORTER1 Gene Family. Plant Cell 2012, 24, 4236–4251. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.H.; Chague, V.; Melamed-Bessudo, C.; Kapulnik, Y.; Jain, A.; Raghothama, K.G.; Levy, A.A.; Silber, A. Functional characterization of LePT4: A phosphate transporter in tomato with mycorrhiza-enhanced expression. J. Exp. Bot. 2007, 58, 2491–2501. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.L.; Wang, D.L.; Jiang, P.; Jia, W.T.; Li, Y.X. Variation of PHT families adapts salt cress to phosphate limitation under salinity. Plant Cell Environ. 2021, 44, 1549–1564. [Google Scholar] [CrossRef]
- Wang, J.H.; Yang, Y.; Liao, L.Z.; Xu, J.W.; Liang, X.; Liu, W. Genome-Wide Identification and Functional Characterization of the Phosphate Transporter Gene Family in Sorghum. Biomolecules 2019, 9, 670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.L.; Zhao, S.; Wu, X.F.; Wang, X.Y.; Nan, Y.Y.; Wang, D.D.; Chen, Q. Identification and characterization of phosphate transporter genes in potato. J. Biotechnol. 2017, 264, 17–28. [Google Scholar] [CrossRef]
- Rausch, C.; Bucher, M. Molecular mechanisms of phosphate transport in plants. Planta 2002, 216, 23–37. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Kalay, G. Cis-regulatory elements: Molecular mechanisms and evolutionary processes underlying divergence. Nat. Rev. Genet. 2012, 13, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Raphael, B.; Nicolas, M.; Martina, J.; Daphnee, B.; Daniel, W.; Pierre-Emmanuel, C. The fine-tuning of mycorrhizal pathway in sorghum depends on both nitrogen-phosphorus availability and the identity of the fungal partner. Plant Cell Environ. 2022, 45, 3354–3366. [Google Scholar] [CrossRef] [PubMed]
- Bonneau, L.; Huguet, S.; Wipf, D.; Pauly, N.; Truong, H.N. Combined phosphate and nitrogen limitation generates a nutrient stress transcriptome favorable for arbuscular mycorrhizal symbiosis in Medicago truncatula. New Phytol. 2013, 199, 188–202. [Google Scholar] [CrossRef]
- Duan, J.F.; Tian, H.; Drijber, R.A.; Gao, Y.J. Systemic and local regulation of phosphate and nitrogen transporter genes by arbuscular mycorrhizal fungi in roots of winter wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2015, 96, 199–208. [Google Scholar] [CrossRef]
- Nouri, E.; Surve, R.; Bapaume, L.; Stumpe, M.; Chen, M.; Zhang, Y.; Ruyter-Spira, C.; Bouwmeester, H.; Glauser, G.; Bruisson, S.; et al. Phosphate Suppression of Arbuscular Mycorrhizal Symbiosis Involves Gibberellic Acid Signalling. Plant Cell Physiol. 2021, 62, 959–970. [Google Scholar] [CrossRef]
- Breuillin, F.; Schramm, J.; Hajirezaei, M.; Ahkami, A.; Favre, P.; Druege, U.; Hause, B.; Bucher, M.; Kretzschmar, T.; Bossolini, E.; et al. Phosphate systemically inhibits development of arbuscular mycorrhiza in Petunia hybrida and represses genes involved in mycorrhizal functioning. Plant J. 2010, 64, 1002–1017. [Google Scholar] [CrossRef]
- Balzergue, C.; Puech-Pages, V.; Becard, G.; Rochange, S.F. The regulation of arbuscular mycorrhizal symbiosis by phosphate in pea involves early and systemic signalling events. J. Exp. Bot. 2011, 62, 1049–1060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, R.; Drissner, D.; Amrhein, N.; Jakobsen, I.; Bucher, M. Mycorrhizal phosphate uptake pathway in tomato is phosphorus-repressible and transcriptionally regulated. New Phytol. 2009, 181, 950–959. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Smith, F.A.; Jakobsen, I. Mycorrhizal fungi can dominate phosphate supply to plants irrespective of growth responses. Plant Physiol. 2003, 133, 16–20. [Google Scholar] [CrossRef] [Green Version]
- Begum, N.; Ahanger, M.A.; Zhang, L.X. AMF inoculation and phosphorus supplementation alleviates drought induced growth and photosynthetic decline in Nicotiana tabacum by up regulating antioxidant metabolism and osmolyte accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Xiao, D.; Che, R.X.; Liu, X.; Tan, Y.J.; Yang, R.; Zhang, W.; He, X.Y.; Xu, Z.H.; Wang, K.L. Arbuscular mycorrhizal fungi abundance was sensitive to nitrogen addition but diversity was sensitive to phosphorus addition in karst ecosystems. Biol. Fertil. Soils 2019, 55, 457–469. [Google Scholar] [CrossRef]
- Essahibi, A.; Benhiba, L.; Fouad, M.O.; Babram, M.A.; Ghoulam, C.; Qaddoury, A. Responsiveness of Carob (Ceratonia siliqua L.) Plants to Arbuscular Mycorrhizal Symbiosis Under Different Phosphate Fertilization Levels. J. Plant Growth Regul. 2019, 38, 1243–1254. [Google Scholar] [CrossRef]
- Calvo-Polanco, M.; Molina, S.; Zamarreno, A.M.; Garcia-Mina, J.M.; Aroca, R. The Symbiosis with the Arbuscular Mycorrhizal Fungus Rhizophagus irregularis Drives Root Water Transport in Flooded Tomato Plants. Plant Cell Physiol. 2014, 55, 1017–1029. [Google Scholar] [CrossRef]
- Martin, S.L.; Mooney, S.J.; Dickinson, M.J.; West, H.M. The effects of simultaneous root colonisation by three Glomus species on soil pore characteristics. Soil Biol. Biochem. 2012, 49, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.; Fang, F.R.; Wu, N.; Li, L.; Tang, M. Nitrate Transporter Gene Expression and Kinetics of Nitrate Uptake by Populus x canadensis ‘Neva’ in Relation to Arbuscular Mycorrhizal Fungi and Nitrogen Availability. Front. Microbiol. 2020, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Nouri, E.; Breuillin-Sessoms, F.; Feller, U.; Reinhardt, D. Phosphorus and Nitrogen Regulate Arbuscular Mycorrhizal Symbiosis in Petunia hybrida. PLoS ONE 2014, 9, 14. [Google Scholar] [CrossRef]
- Balzergue, C.; Chabaud, M.; Barker, D.G.; Becard, G.; Rochange, S.F. High phosphate reduces host ability to develop arbuscular mycorrhizal symbiosis without affecting root calcium spiking responses to the fungus. Front. Plant Sci. 2013, 4, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, L.; Guo, Y.X.; Chen, L.Y.; Liang, R.K.; Gu, M.A.; Xu, G.H.; Zhao, J.; Walk, T.; Liao, H. Functional Characterization of 14 Pht1 Family Genes in Yeast and Their Expressions in Response to Nutrient Starvation in Soybean. PLoS ONE 2012, 7, e47726. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Chang, X.J.; Ye, Y.; Xie, W.B.; Wu, P.; Lian, X.M. Comprehensive Sequence and Whole-Life-Cycle Expression Profile Analysis of the Phosphate Transporter Gene Family in Rice. Mol. Plant 2011, 4, 1105–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daram, P.; Brunner, S.; Rausch, C.; Steiner, C.; Amrhein, N.; Bucher, M. Pht2;1 encodes a low-affinity phosphate transporter from Arabidopsis. Plant Cell 1999, 11, 2153–2166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.M.; Versaw, W.K.; Liu, J.Y.; Harrison, M.J. A phosphate transporter from Medicago truncatula is expressed in the photosynthetic tissues of the plant and located in the chloroplast envelope. New Phytol. 2003, 157, 291–302. [Google Scholar] [CrossRef] [PubMed]
- Rausch, C.; Zimmermann, P.; Amrhein, N.; Bucher, M. Expression analysis suggests novel roles for the plastidic phosphate transporter Pht2;1 in auto- and heterotrophic tissues in potato and Arabidopsis. Plant J. 2004, 39, 13–28. [Google Scholar] [CrossRef]
- Aung, K.; Lin, S.I.; Wu, C.C.; Huang, Y.T.; Su, C.L.; Chiou, T.J. pho2, a phosphate overaccumulator, is caused by a nonsense mutation in a MicroRNA399 target gene. Plant Physiol. 2006, 141, 1000–1011. [Google Scholar] [CrossRef] [Green Version]
- Fester, T.; Wray, V.; Nimtz, M.; Strack, D. Is stimulation of carotenoid biosynthesis in arbuscular mycorrhizal roots a general phenomenon? Phytochemistry 2005, 66, 1781–1786. [Google Scholar] [CrossRef]
- Chen, C.J.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.H.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Zhao, Y.; Ma, R.Y.; Xu, D.L.; Bi, H.H.; Xia, Z.L.; Peng, H.R. Genome-Wide Identification and Analysis of the AP2 Transcription Factor Gene Family in Wheat (Triticum aestivum L.). Front. Plant Sci. 2019, 10, 1286. [Google Scholar] [CrossRef] [Green Version]
- Shu, B.; Xie, Y.C.; Zhang, F.; Zhang, D.J.; Liu, C.Y.; Wu, Q.S.; Luo, C. Genome-wide identification of citrus histone acetyltransferase and deacetylase families and their expression in response to arbuscular mycorrhizal fungi and drought. J. Plant Interact. 2021, 16, 367–376. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158-IN18. [Google Scholar] [CrossRef]
- Veneklaas, E.J.; Stevens, J.; Cawthray, G.R.; Turner, S.; Grigg, A.M.; Lambers, H. Chickpea and white lupin rhizosphere carboxylates vary with soil properties and enhance phosphorus uptake. Plant Soil 2003, 248, 187–197. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Accession Number | Location | ORF Length (bp) | Size (bp) | Molecular Weight | PI |
|---|---|---|---|---|---|---|
| SlPT1 | NP_001234361.2 | 9:70098389_70102955 | 538 | 2002 | 58,699.54 | 8.51 |
| SlPT2 | NP_001234043.1 | 3:398936_401108 | 528 | 1826 | 57,762.29 | 8.61 |
| SlPT3 | NP_001318089.1 | 9:70109008_70111237 | 534 | 1867 | 58,536.25 | 8.67 |
| SlPT4 | NP_001234674.2 | 6:35656579_35659001 | 545 | 2019 | 60,502.44 | 8.49 |
| SlPT5 | XP_004240951.1 | 6:35659554_35664968 | 529 | 2384 | 59,037.82 | 8.73 |
| SlPT6 | XP_004234039.1 | 3:411956_415011 | 528 | 1857 | 57,762.29 | 8.61 |
| SlPT7 | XP_004247235.1 | 9:65061109_65063652 | 533 | 1793 | 58,300.12 | 8.90 |
| SlPT8 | XP_004240760.1 | 6:23920599_23923011 | 534 | 2011 | 58,486.39 | 8.90 |
| SlPHT2 | XP_004239128.1 | 5:6724597_6728688 | 577 | 2180 | 61,077.15 | 9.28 |
| SlPHT3.1 | NP_001266267.1 | 2:55614122_55617796 | 358 | 1593 | 38,440.82 | 9.22 |
| SlPHT3.2 | XP_004234502.1 | 3:4560513_4568003 | 324 | 1497 | 35,672.47 | 9.41 |
| SlPHT3.3 | XP_004235310.1 | 3:61601724_61604831 | 352 | 1446 | 37,656.50 | 9.18 |
| SlPHT3.4 | XP_004242142.1 | 6:44858180_44861909 | 362 | 2213 | 38,663.85 | 9.25 |
| SlPHT4.1 | XP_004235159.1 | 3:55746222_55749729 | 521 | 2176 | 56,581.50 | 6.40 |
| SlPHT4.2 | XP_004240757.1 | 6:24109636_24113354 | 510 | 2109 | 55,395.36 | 8.50 |
| SlPHT4.3 | XP_004242279.1 | 6:39021711_39028059 | 537 | 2465 | 58,269.86 | 8.36 |
| SlPHT4.4 | XP_010313844.1 | 12:3467625_3470879 | 506 | 2183 | 54,221.91 | 6.72 |
| SlPHO1.1 | XP_004247746.1 | 9:70338431_70351065 | 788 | 2766 | 90,815.17 | 9.21 |
| SlPHO1.2 | XP_019070605.2 | 8:57407221_57412919 | 753 | 2950 | 87,205.56 | 9.10 |
| SlPHO1.3 | XP_004238979.1 | 5:4267987_4274342 | 786 | 2646 | 92,036.01 | 9.17 |
| SlPHO1.4 | XP_010320919.1 | 5:6256934_6264807 | 776 | 3394 | 91,158.92 | 9.36 |
| SlPHO1.5 | XP_004232209.1 | 2:51014310_51019443 | 777 | 3063 | 89,979.97 | 9.35 |
| SlPHO1.6 | XP_004232205.1 | 2:51026560_51031505 | 792 | 2917 | 92,266.21 | 8.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rui, W.; Ma, J.; Wei, N.; Zhu, X.; Li, Z. Genome-Wide Analysis of the PHT Gene Family and Its Response to Mycorrhizal Symbiosis in Tomatoes under Phosphate Starvation Conditions. Int. J. Mol. Sci. 2023, 24, 10246. https://doi.org/10.3390/ijms241210246
Rui W, Ma J, Wei N, Zhu X, Li Z. Genome-Wide Analysis of the PHT Gene Family and Its Response to Mycorrhizal Symbiosis in Tomatoes under Phosphate Starvation Conditions. International Journal of Molecular Sciences. 2023; 24(12):10246. https://doi.org/10.3390/ijms241210246
Chicago/Turabian StyleRui, Wenjing, Jing Ma, Ning Wei, Xiaoya Zhu, and Zhifang Li. 2023. "Genome-Wide Analysis of the PHT Gene Family and Its Response to Mycorrhizal Symbiosis in Tomatoes under Phosphate Starvation Conditions" International Journal of Molecular Sciences 24, no. 12: 10246. https://doi.org/10.3390/ijms241210246