
Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infect Human Intestinal Epithelial Cells and Induce Moderate Inflammation
 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
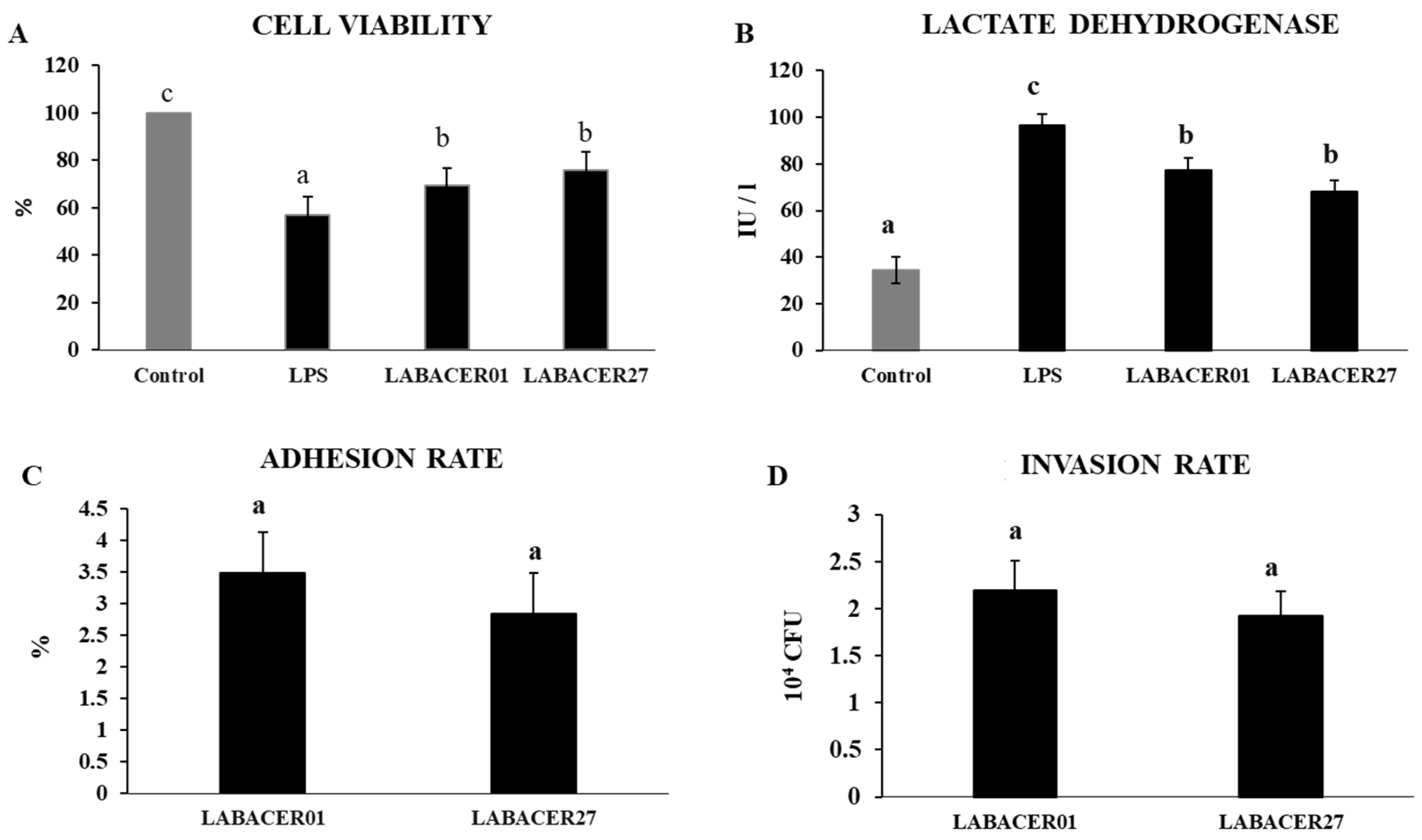
2.1. K. pneumoniae ST25 Adhere and Infect Caco-2 Cells
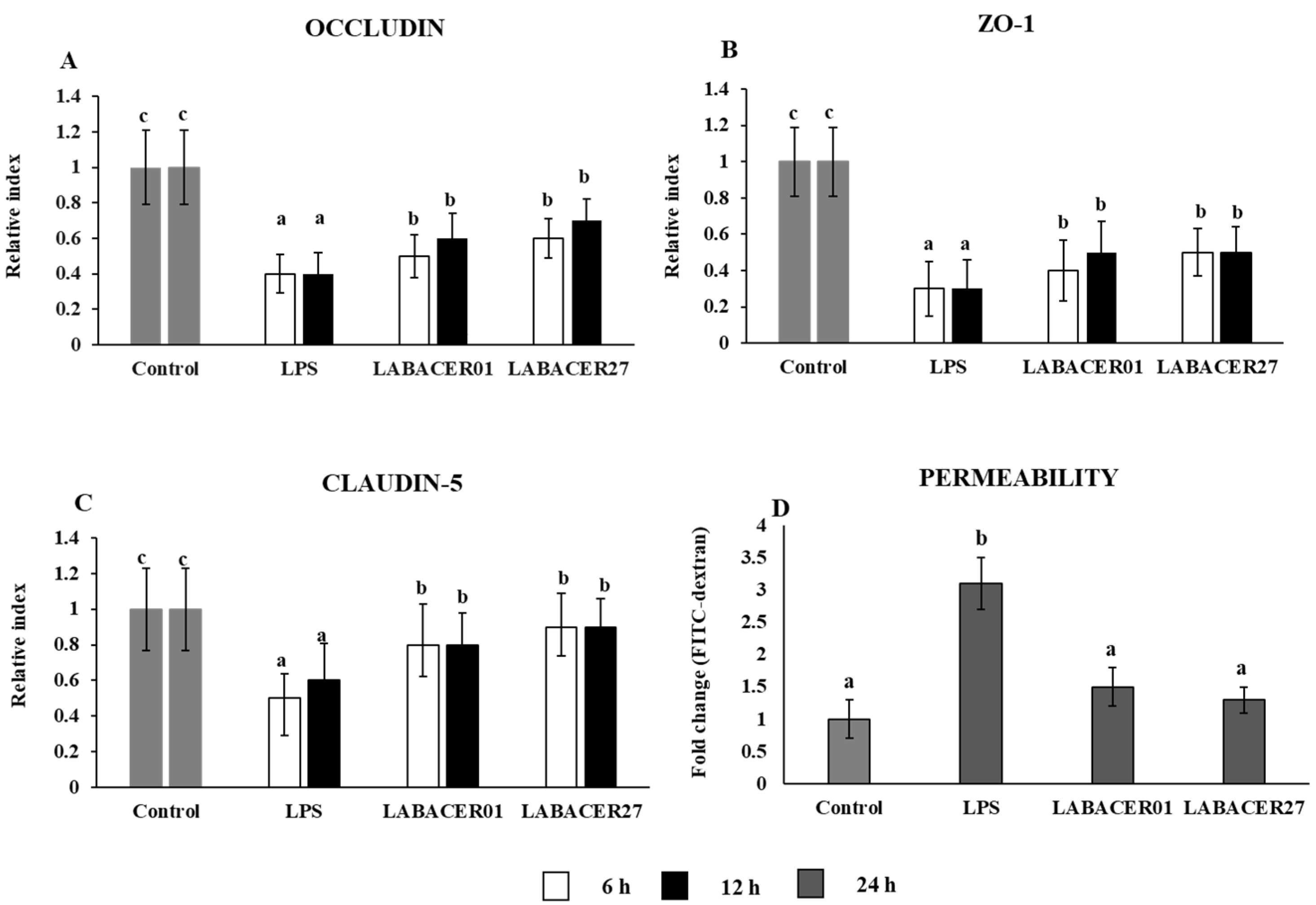
2.2. K. pneumoniae ST25 Infection Affects Tight Junction Proteins Expression in Caco-2 Cells
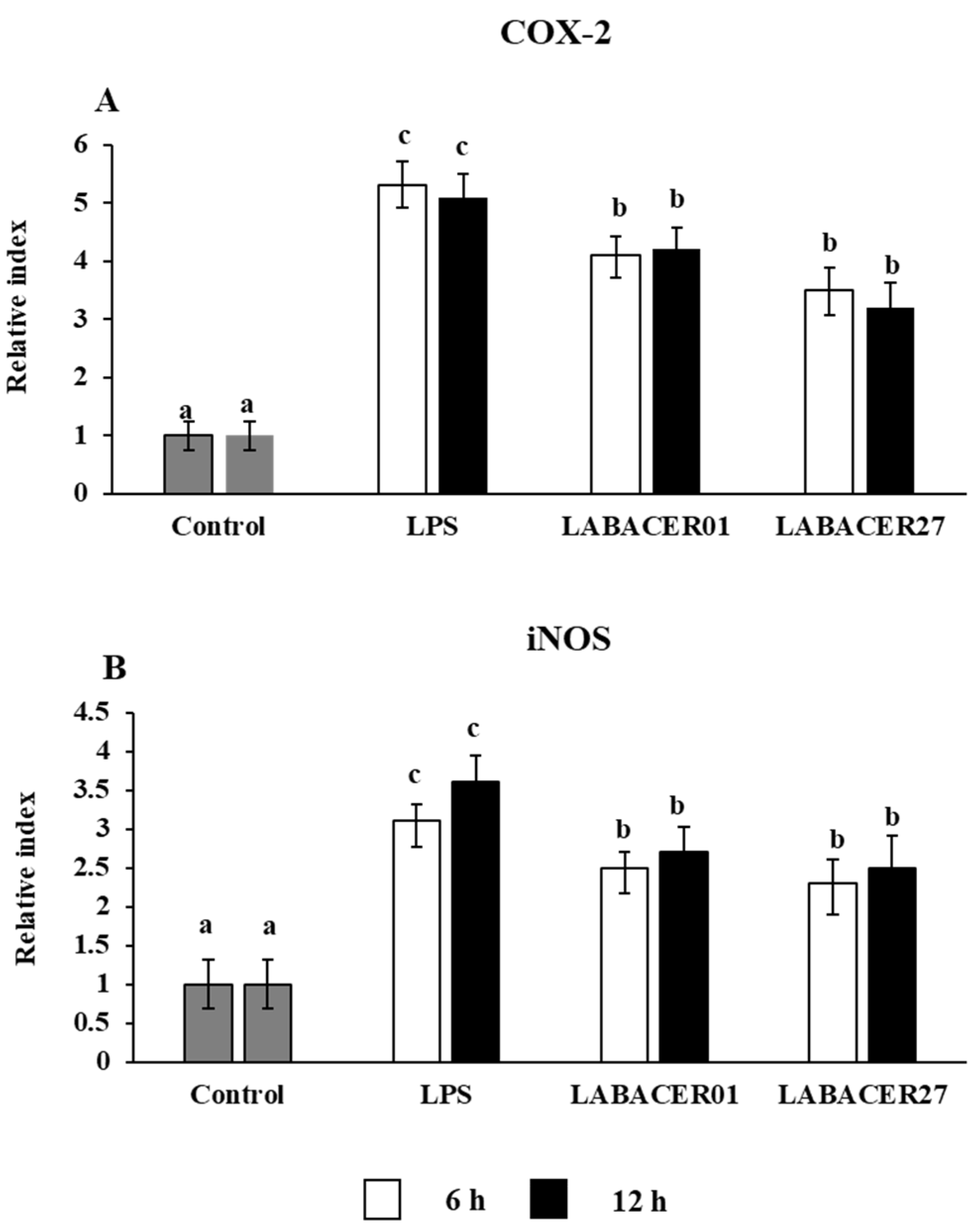
2.3. K. pneumoniae ST25 Infection Affect COX-2 and iNOS Expression in Caco-2 Cells
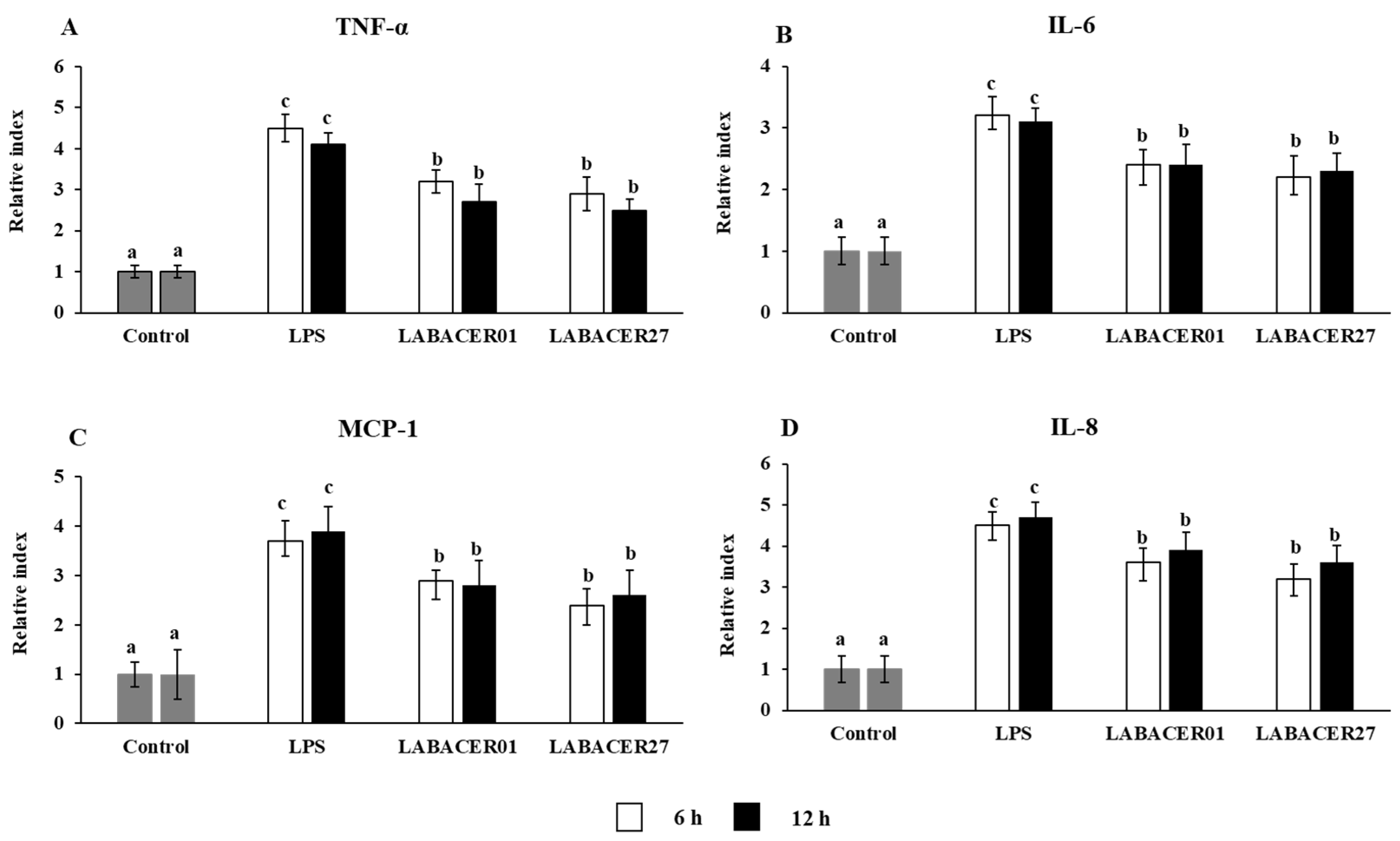
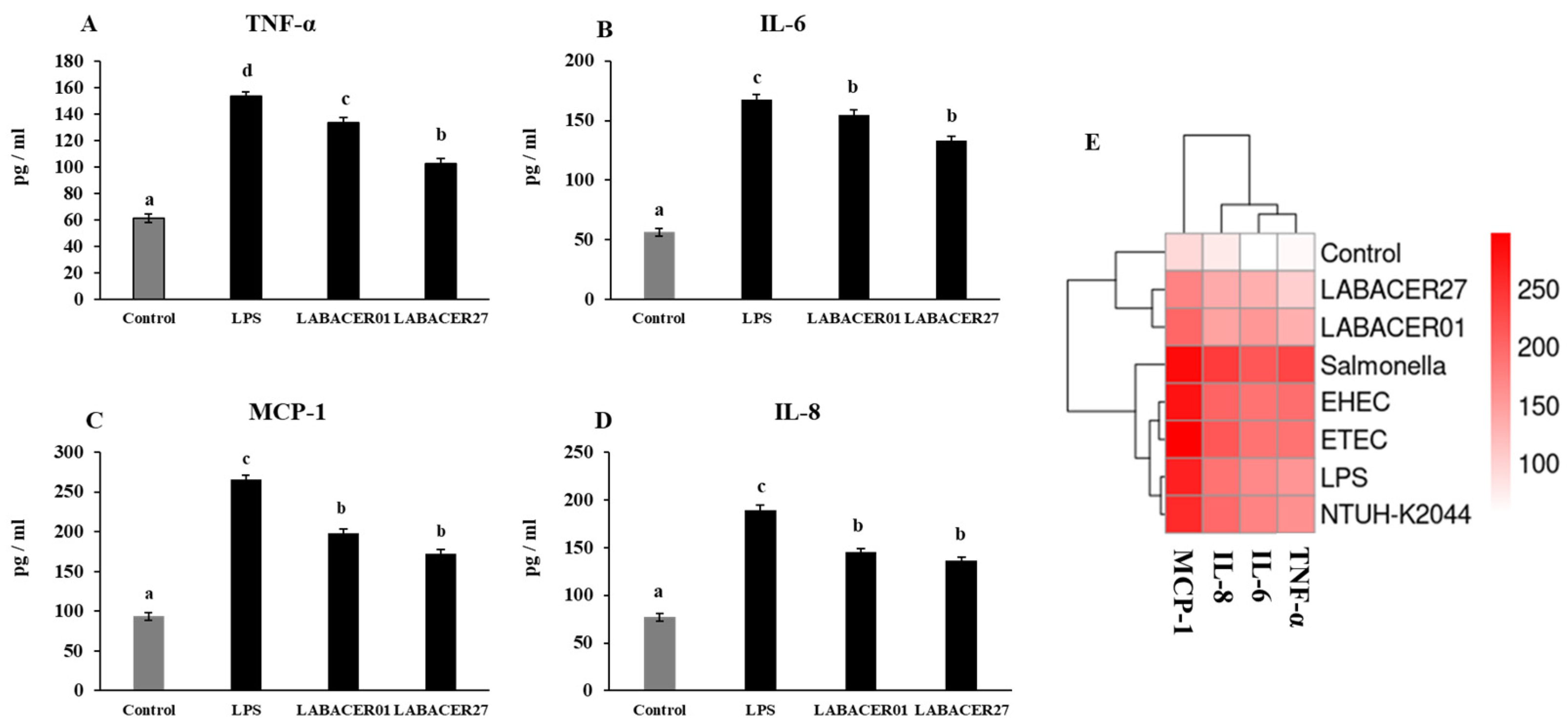
2.4. K. pneumoniae ST25 Trigger Inflammatory Factors Production in Caco-2 Cells
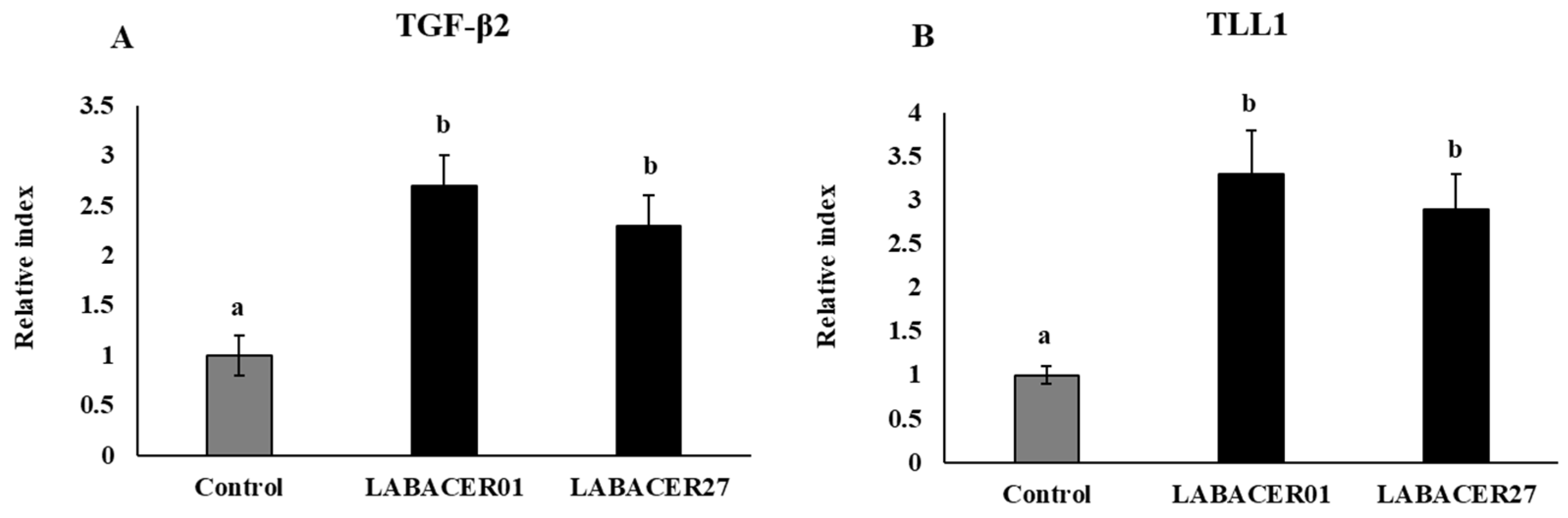
2.5. K. pneumoniae ST25 Modulate TGF-β and TLL1 Expression in Caco-2 Cells
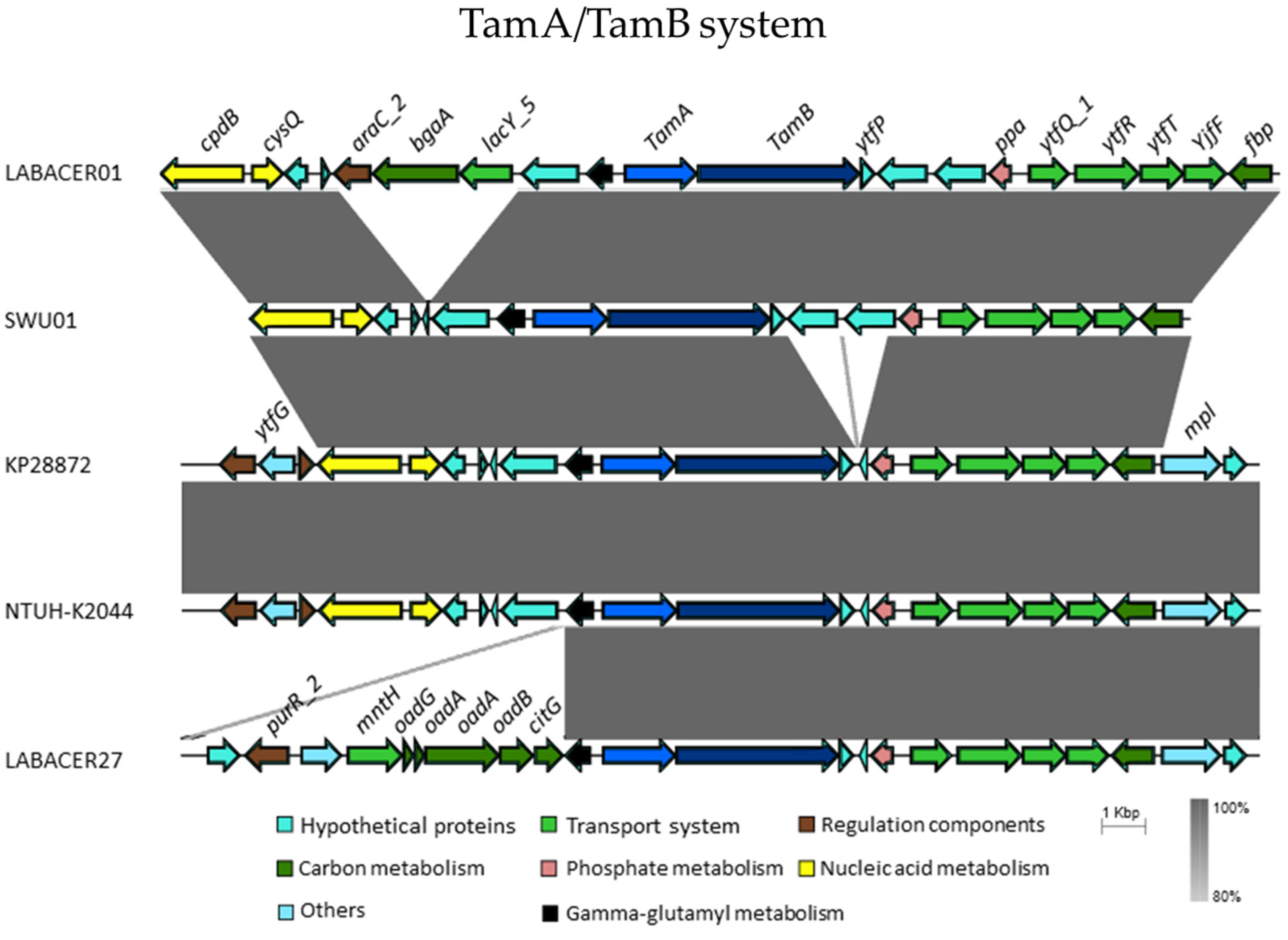
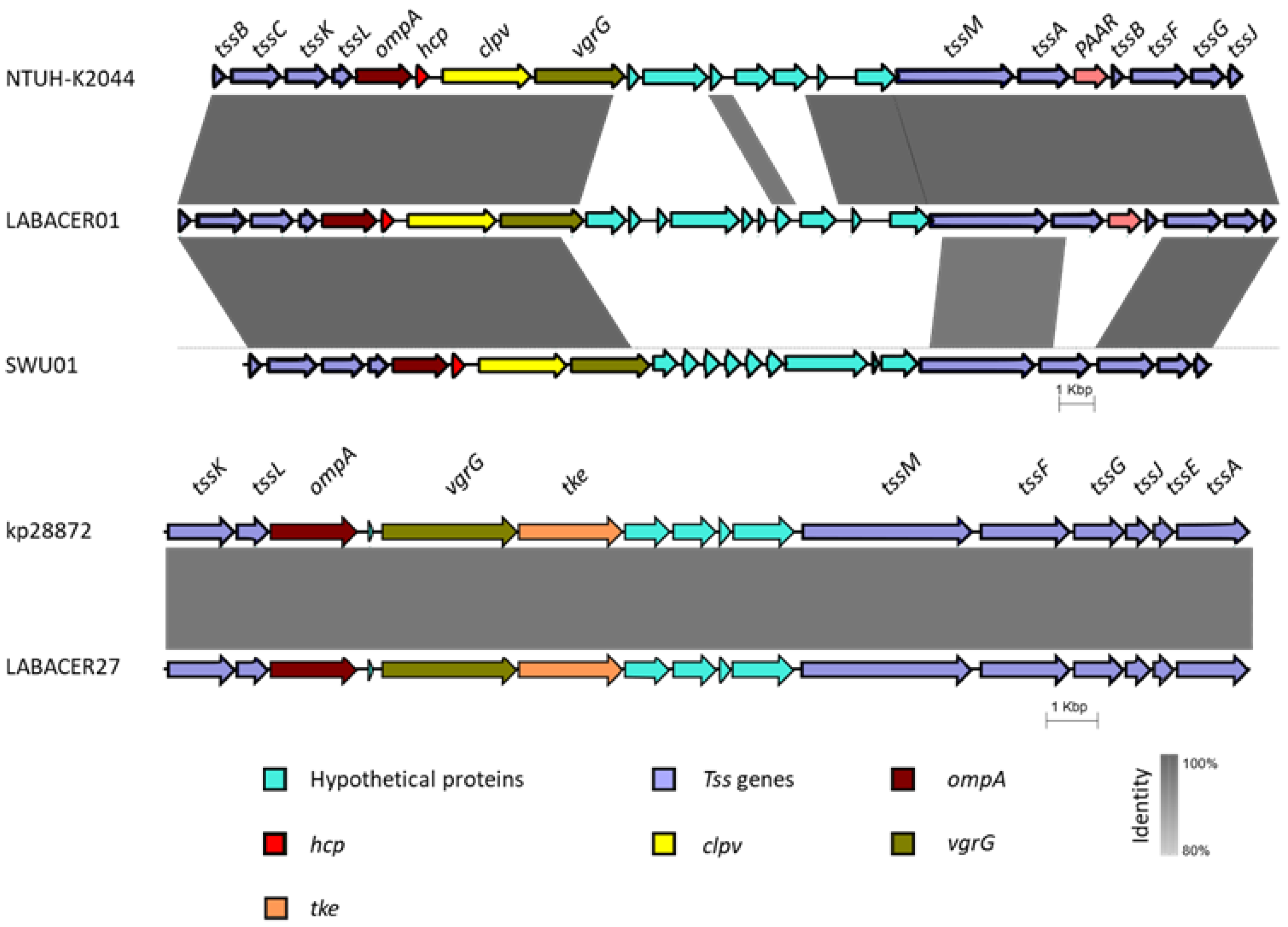
2.6. In Silico Analysis of Virulence Genes from K. pneumoniae ST25
3. Discussion
4. Materials and Methods
4.1. Strains
4.2. Cell Culture and Viability Assays
4.3. Lactate Dehydrogenase (LDH) Activity Assay
4.4. Adhesion, Invasion, and Permeability Experiments in Caco-2 Cells
4.5. RT-PCR
4.6. Cytokine Concentrations
4.7. Genomic Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Keesler, R.I.; Colagross-Schouten, A.; Reader, J.R. Clinical and pathologic features of spontaneous Klebsiella pneumoniae infection in 9 rhesus macaques (Macaca mulatta). Comp. Med. 2020, 70, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef]
- Broberg, C.A.; Palacios, M.; Miller, V.L. Klebsiella: A long way to go towards understanding this enigmatic jet-setter. F1000Prime Rep. 2014, 6, 64. [Google Scholar] [CrossRef]
- Lin, Y.T.; Siu, L.K.; Lin, J.C.; Chen, T.L.; Tseng, C.P.; Yeh, K.M.; Chang, F.Y.; Fung, C.P. Seroepidemiology of Klebsiella pneumoniae colonizing the intestinal tract of healthy Chinese and overseas Chinese adults in Asian countries. BMC Microbiol. 2012, 12, 13. [Google Scholar] [CrossRef]
- Russo, T.A.; Marr, C.M. Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 2019, 32, e00001-19. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wu, C.; Bao, D.; Jia, H.; Draz, M.S.; He, F.; Ruan, Z. Global evolution and geographic diversity of hypervirulent carbapenem-resistant Klebsiella pneumoniae. Lancet Infect. Dis. 2022, 22, 761–762. [Google Scholar] [CrossRef]
- Young, T.M.; Bray, A.S.; Nagpal, R.K.; Caudell, D.L.; Yadav, H.; Zafar, M.A. Animal model to study Klebsiella pneumoniae gastrointestinal colonization and host-to-host transmission. Infect. Immun. 2020, 88, e00071-20. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, S.; Pei, H.; Tu, F.; Liu, B.; Yan, J.; Lin, X. Klebsiella pneumoniae activates the TGF-beta signaling pathway to adhere to and invade intestinal epithelial cells via enhancing TLL1 expression. Int. J. Med. Microbiol. 2022, 312, 151561. [Google Scholar] [CrossRef]
- Struve, C.; Bojer, M.; Nielsen, E.M.; Hansen, D.S.; Krogfelt, K.A. Investigation of the putative virulence gene magA in a worldwide collection of 495 Klebsiella isolates: magA is restricted to the gene cluster of Klebsiella pneumoniae capsule serotype K1. J. Med. Microbiol. 2005, 54, 1111–1113. [Google Scholar] [CrossRef]
- Huang, Y.; Li, J.; Gu, D.; Fang, Y.; Chan, E.W.; Chen, S.; Zhang, R. Rapid Detection of K1 Hypervirulent Klebsiella pneumoniae by MALDI-TOF MS. Front. Microbiol. 2015, 6, 1435. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence factors in hypervirulent Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 642484. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.C.; Peng, H.L.; Chang, H.Y. RmpA2, an activator of capsule biosynthesis in Klebsiella pneumoniae CG43, regulates K2 cps gene expression at the transcriptional level. J. Bacteriol. 2003, 185, 788–800. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Chen, Y.S.; Wu, C.Y.; Chang, H.Y.; Lai, Y.C.; Peng, H.L. RmpA regulation of capsular polysaccharide biosynthesis in Klebsiella pneumoniae CG43. J. Bacteriol. 2010, 192, 3144–3158. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.J.; Littmann, E.R.; Seok, R.; Leiner, I.M.; Taur, Y.; Peled, J.; van den Brink, M.; Ling, L.; Chen, L.; Kreiswirth, B.N.; et al. Genome-wide screening for enteric colonization factors in carbapenem-resistant ST258 Klebsiella pneumoniae. mBio 2019, 10, e02663-18. [Google Scholar] [CrossRef]
- Hsieh, P.F.; Lu, Y.R.; Lin, T.L.; Lai, L.Y.; Wang, J.T. Klebsiella pneumoniae type VI secretion system contributes to bacterial competition, cell invasion, type-1 fimbriae expression, and in vivo colonization. J. Infect. Dis. 2019, 219, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.R.; Pan, Y.J.; Liu, J.Y.; Chen, C.T.; Lin, T.L.; Wang, J.T. Klebsiella pneumoniae translocates across the intestinal epithelium via Rho GTPase- and phosphatidylinositol 3-kinase/Akt-dependent cell invasion. Infect. Immun. 2015, 83, 769–779. [Google Scholar] [CrossRef]
- Vargas, J.M.; Moreno Mochi, M.P.; Nunez, J.M.; Caceres, M.; Mochi, S.; Del Campo Moreno, R.; Jure, M.A. Virulence factors and clinical patterns of multiple-clone hypermucoviscous KPC-2 producing K. pneumoniae. Heliyon 2019, 5, e01829. [Google Scholar] [CrossRef]
- Dentice Maidana, S.; Ortiz Moyano, R.; Vargas, J.M.; Fukuyama, K.; Kurata, S.; Melnikov, V.; Jure, M.A.; Kitazawa, H.; Villena, J. Respiratory Commensal bacteria increase protection against hypermucoviscous carbapenem-resistant Klebsiella pneumoniae ST25 infection. Pathogens 2022, 11, 1063. [Google Scholar] [CrossRef]
- Albarracin, L.; Ortiz Moyano, R.; Vargas, J.M.; Andrade, B.G.N.; Cortez Zamar, J.; Dentice Maidana, S.; Fukuyama, K.; Kurata, S.; Jure, M.A.; Kitazawa, H.; et al. Genomic and immunological characterization of hypermucoviscous carbapenem-resistant Klebsiella pneumoniae ST25 isolates from Northwest Argentina. Int. J. Mol. Sci. 2022, 23, 7361. [Google Scholar] [CrossRef]
- Jure, M.A.; Albarracin, L.; Vargas, J.M.; Maidana, S.D.; Zamar, J.C.; Kitazawa, H.; Villena, J. Draft genome sequences of two hypermucoviscous carbapenem-resistant ST25 Klebsiella pneumoniae strains causing respiratory and systemic infections. J. Glob. Antimicrob. Resist. 2021, 26, 174–176. [Google Scholar] [CrossRef]
- Cuevas-Ramos, G.; Petit, C.R.; Marcq, I.; Boury, M.; Oswald, E.; Nougayrede, J.P. Escherichia coli induces DNA damage in vivo and triggers genomic instability in mammalian cells. Proc. Natl. Acad. Sci. USA 2010, 107, 11537–11542. [Google Scholar] [CrossRef] [PubMed]
- Grasso, F.; Frisan, T. Bacterial Genotoxins: Merging the DNA damage response into infection biology. Biomolecules 2015, 5, 1762–1782. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.C.; Chen, Y.T.; Chiang, M.K.; Wang, Y.C.; Hsiao, P.Y.; Huang, Y.J.; Lin, C.T.; Cheng, C.C.; Liang, C.L.; Lai, Y.C. Colibactin Contributes to the hypervirulence of pks(+) K1 CC23 Klebsiella pneumoniae in mouse meningitis infections. Front. Cell. Infect. Microbiol. 2017, 7, 103. [Google Scholar] [CrossRef] [PubMed]
- Xenofontos, E.; Renieris, G.; Kalogridi, M.; Droggiti, D.E.; Synodinou, K.; Damoraki, G.; Koufargyris, P.; Sabracos, L.; Giamarellos-Bourboulis, E.J. An animal model of limitation of gut colonization by carbapenemase-producing Klebsiella pneumoniae using rifaximin. Sci. Rep. 2022, 12, 3789. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Suda, W.; Luo, C.; Kawaguchi, T.; Motoo, I.; Narushima, S.; Kiguchi, Y.; Yasuma, K.; Watanabe, E.; Tanoue, T.; et al. Ectopic colonization of oral bacteria in the intestine drives T(H)1 cell induction and inflammation. Science 2017, 358, 359–365. [Google Scholar] [CrossRef]
- Pope, J.L.; Yang, Y.; Newsome, R.C.; Sun, W.; Sun, X.; Ukhanova, M.; Neu, J.; Issa, J.P.; Mai, V.; Jobin, C. Microbial colonization coordinates the pathogenesis of a Klebsiella pneumoniae infant isolate. Sci. Rep. 2019, 9, 3380. [Google Scholar] [CrossRef] [PubMed]
- Caballero, S.; Carter, R.; Ke, X.; Susac, B.; Leiner, I.M.; Kim, G.J.; Miller, L.; Ling, L.; Manova, K.; Pamer, E.G. Distinct but spatially overlapping intestinal niches for vancomycin-resistant Enterococcus faecium and carbapenem-resistant Klebsiella pneumoniae. PLoS Pathog. 2015, 11, e1005132. [Google Scholar] [CrossRef]
- Lee, I.A.; Kim, D.H. Klebsiella pneumoniae increases the risk of inflammation and colitis in a murine model of intestinal bowel disease. Scand. J. Gastroenterol. 2011, 46, 684–693. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhao, W.; Liu, G.; Ali, T.; Chen, P.; Liu, Y.; Kastelic, J.P.; Han, B.; Gao, J. Bacteriophages isolated from dairy farm mitigated Klebsiella pneumoniae-induced inflammation in bovine mammary epithelial cells cultured in vitro. BMC Vet. Res. 2021, 17, 37. [Google Scholar] [CrossRef]
- Motta, C.; Salmeri, M.; Anfuso, C.D.; Amodeo, A.; Scalia, M.; Toscano, M.A.; Giurdanella, G.; Alberghina, M.; Lupo, G. Klebsiella pneumoniae induces an inflammatory response in an in vitro model of blood-retinal barrier. Infect. Immun. 2014, 82, 851–863. [Google Scholar] [CrossRef]
- Fodah, R.A.; Scott, J.B.; Tam, H.H.; Yan, P.; Pfeffer, T.L.; Bundschuh, R.; Warawa, J.M. Correlation of Klebsiella pneumoniae comparative genetic analyses with virulence profiles in a murine respiratory disease model. PLoS ONE 2014, 9, e107394. [Google Scholar] [CrossRef]
- Clarke, T.B. Early innate immunity to bacterial infection in the lung is regulated systemically by the commensal microbiota via nod-like receptor ligands. Infect. Immun. 2014, 82, 4596–4606. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.R.; Lin, T.L.; Chen, Y.C.; Chou, H.C.; Wang, J.T. The role of Klebsiella pneumoniae rmpA in capsular polysaccharide synthesis and virulence revisited. Microbiology 2011, 157, 3446–3457. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.M.; Li, L.H.; Yan, J.J.; Tsao, N.; Liao, T.L.; Tsai, H.C.; Fung, C.P.; Chen, H.J.; Liu, Y.M.; Wang, J.T.; et al. Genome sequencing and comparative analysis of Klebsiella pneumoniae NTUH-K2044, a strain causing liver abscess and meningitis. J. Bacteriol. 2009, 191, 4492–4501. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.T.; Chuang, Y.P.; Shun, C.T.; Chang, S.C.; Wang, J.T. A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J. Exp. Med. 2004, 199, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Storey, D.; McNally, A.; Astrand, M.; Sa-Pessoa Graca Santos, J.; Rodriguez-Escudero, I.; Elmore, B.; Palacios, L.; Marshall, H.; Hobley, L.; Molina, M.; et al. Klebsiella pneumoniae type VI secretion system-mediated microbial competition is PhoPQ controlled and reactive oxygen species dependent. PLoS Pathog. 2020, 16, e1007969. [Google Scholar] [CrossRef] [PubMed]
- Merciecca, T.; Bornes, S.; Nakusi, L.; Theil, S.; Rendueles, O.; Forestier, C.; Miquel, S. Role of Klebsiella pneumoniae Type VI secretion system (T6SS) in long-term gastrointestinal colonization. Sci. Rep. 2022, 12, 16968. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Colbactin | Mucoviscosity Associated Protein | RmpA | TamA/TamB System | Type VI Secretion System Main Cluster (Genes ≥ 8) |
|---|---|---|---|---|---|
| KP28872 | - | - | - | + | tssA, tssE, tssF, tssG, tssI (vgrG), tssJ, tssk, tssL, tssM. |
| LABACER01 | - | - | - | + | tssA, tssB (2 copies), tssC, tssD (hcp), tssF, tssG, tssH (clpv), tssI (vgrG), tssJ, tssk, tssL, tssM, evpJ (PAAR). |
| LABACER27 | - | - | - | + | tssA, tssE, tssF, tssG, tssI (vgrG), tssJ, tssk, tssL, tssM. |
| NTUH-K2044 | - | + | + | + | tssA, tssB (2 copies), tssC, tssD (hcp), tssF, tssG, tssH (clpv), tssI (vgrG), tssJ, tssk, tssL, tssM, evpJ (PAAR). |
| SWU01 | - | - | - | + | tssA, tssB, tssC, tssD (hcp), tssF, tssG, tssH (clpv), tssI (vgrG), tssJ, tssk, tssL, tssM, evpJ (PAAR). |
| Gene | Amplicon Size (pb) | Forward Primer (5′→3′) | Reverse Primer (5′→3′) |
|---|---|---|---|
| Occludin | 114 | GAGTTGTATCTGTTGTTGT | TTCGTGGTATAGCATTCT |
| ZO-1 | 118 | GGTGAAGTGAAGACAATG | GGTAATATGGTGAAGTTAGAG |
| Claudin-5 | 84 | TTAACAGACGGAATGAAGT | GAAGCGAAATCCTCAGTC |
| COX-2 | 117 | GAGAGATGTATCCTCCCACAGTCA | GACCAGGCACCAGACCAAAG |
| iNOS | 278 | CCTTACGAGGCGAAGAAGGACAG | CAGTTTGAGAGAGGAGGCTCCG |
| TNF-α | 360 | GTCAGATCATCTTCTCGAACC | CAGATAGATGGGCTCATACC |
| IL-6 | 211 | GACAGCCACTCACCTCTTCA | TTCACCAGGCAAGTCTCCTC |
| MCP-1 | 93 | AGTCTCTGCCGCCCTTCT | GTGACTGGGGCATTGATTG |
| IL-8 | 168 | ACTCCAAACCTTTCCACCC | CCCTCTTCAAAAACTTCTCCAC |
| Strains | Assembly | Size | GC% | Scaffolds | CDS |
|---|---|---|---|---|---|
| KP28872 | GCA_013623415.1 | 5.9 | 56.9 | 364 | 5553 |
| LABACER01 | GCA_013375175.1 | 5.6 | 57.1 | 61 | 5306 |
| LABACER27 | GCA_013375185.1 | 5.6 | 57.1 | 51 | 5170 |
| NTUH-K2044 | GCA_000009885.1 | 5.5 | 57.4 | 2 | 5028 |
| SWU01 | GCA_001902475.1 | 5.7 | 57.4 | 2 | 5420 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dentice Maidana, S.; Elean, M.; Fukuyama, K.; Imamura, Y.; Albarracín, L.; Saha, S.; Suda, Y.; Kurata, S.; Jure, M.Á.; Kitazawa, H.; et al. Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infect Human Intestinal Epithelial Cells and Induce Moderate Inflammation. Int. J. Mol. Sci. 2023, 24, 8804. https://doi.org/10.3390/ijms24108804
Dentice Maidana S, Elean M, Fukuyama K, Imamura Y, Albarracín L, Saha S, Suda Y, Kurata S, Jure MÁ, Kitazawa H, et al. Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infect Human Intestinal Epithelial Cells and Induce Moderate Inflammation. International Journal of Molecular Sciences. 2023; 24(10):8804. https://doi.org/10.3390/ijms24108804
Chicago/Turabian StyleDentice Maidana, Stefania, Mariano Elean, Kohtaro Fukuyama, Yoshiya Imamura, Leonardo Albarracín, Sudeb Saha, Yoshihito Suda, Shoichiro Kurata, María Ángela Jure, Haruki Kitazawa, and et al. 2023. "Hypermucoviscous Carbapenem-Resistant Klebsiella pneumoniae ST25 Infect Human Intestinal Epithelial Cells and Induce Moderate Inflammation" International Journal of Molecular Sciences 24, no. 10: 8804. https://doi.org/10.3390/ijms24108804