
Determinants of COVID-19 Disease Severity–Lessons from Primary and Secondary Immune Disorders including Cancer
 , , ,
, , ,
Abstract
:1. Introduction
2. Immunity, Immune Aging and COVID-19
2.1. Immune Response against SARS-CoV-2
2.2. Lessons from HIV Infection
2.3. Vaccination and Immunity
3. Immune Dysregulation and Severity of COVID-19 Infection
3.1. COVID-19 in Patients with Cancer—Clinical Data, Risk Factors and Vaccination Response
3.2. Immune Dysregulation and Severity of COVID-19 Infection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karami, H.; Derakhshani, A.; Ghasemigol, M.; Fereidouni, M.; Miri-Moghaddam, E.; Baradaran, B.; Tabrizi, N.J.; Najafi, S.; Solimando, A.G.; Marsh, L.M.; et al. Weighted Gene Co-Expression Network Analysis Combined with Machine Learning Validation to Identify Key Modules and Hub Genes Associated with SARS-CoV-2 Infection. J. Clin. Med. 2021, 10, 3567. [Google Scholar] [CrossRef] [PubMed]
- Abdelmoaty, M.M.; Yeapuri, P.; Machhi, J.; Olson, K.E.; Shahjin, F.; Kumar, V.; Zhou, Y.; Liang, J.; Pandey, K.; Acharya, A.; et al. Defining the Innate Immune Responses for SARS-CoV-2-Human Macrophage Interactions. Front. Immunol. 2021, 12, 741502. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Chu, H.; Hou, Y.; Chai, Y.; Shuai, H.; Lee, A.C.-Y.; Zhang, X.; Wang, Y.; Hu, B.; Huang, X.; et al. Attenuated Interferon and Proinflammatory Response in SARS-CoV-2-Infected Human Dendritic Cells Is Associated with Viral Antagonism of STAT1 Phosphorylation. J. Infect. Dis. 2020, 222, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Meffre, E.; Iwasaki, A. Interferon Deficiency Can Lead to Severe COVID. Nature 2020, 587, 374–376. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn Errors of Type I IFN Immunity in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.-H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Béziat, V.; et al. Autoantibodies against Type I IFNs in Patients with Life-Threatening COVID-19. Science 2020, 370, eabd4585. [Google Scholar] [CrossRef]
- Su, H.C.; Jing, H.; Zhang, Y.; Casanova, J.-L. Interfering with Interferons: A Critical Mechanism for Critical COVID-19 Pneumonia. Annu. Rev. Immunol. 2023, 41, 561–585. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; Astudillo, M.G.; Yang, D.; Miller, T.E.; Feldman, J.; Hauser, B.M.; Caradonna, T.M.; Clayton, K.L.; Nitido, A.D.; et al. COVID-19-Neutralizing Antibodies Predict Disease Severity and Survival. Cell 2021, 184, 476–488.e11. [Google Scholar] [CrossRef]
- Daamen, A.R.; Bachali, P.; Owen, K.A.; Kingsmore, K.M.; Hubbard, E.L.; Labonte, A.C.; Robl, R.; Shrotri, S.; Grammer, A.C.; Lipsky, P.E. Comprehensive Transcriptomic Analysis of COVID-19 Blood, Lung, and Airway. Sci. Rep. 2021, 11, 7052. [Google Scholar] [CrossRef]
- Gardinassi, L.G.; Souza, C.O.S.; Sales-Campos, H.; Fonseca, S.G. Immune and Metabolic Signatures of COVID-19 Revealed by Transcriptomics Data Reuse. Front. Immunol. 2020, 11, 1636. [Google Scholar] [CrossRef]
- Dwivedi, P.; Alam, S.I.; Tomar, R.S. Secretome, Surfome and Immunome: Emerging Approaches for the Discovery of New Vaccine Candidates against Bacterial Infections. World J. Microbiol. Biotechnol. 2016, 32, 155. [Google Scholar] [CrossRef] [PubMed]
- Kuri-Cervantes, L.; Pampena, M.B.; Meng, W.; Rosenfeld, A.M.; Ittner, C.A.G.; Weisman, A.R.; Agyekum, R.S.; Mathew, D.; Baxter, A.E.; Vella, L.A.; et al. Comprehensive Mapping of Immune Perturbations Associated with Severe COVID-19. Sci. Immunol. 2020, 5, eabd7114. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.Z.; Poh, C.M.; Rénia, L.; MacAry, P.A.; Ng, L.F.P. The Trinity of COVID-19: Immunity, Inflammation and Intervention. Nat. Rev. Immunol. 2020, 20, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Randolph, H.E.; Barreiro, L.B. Herd Immunity: Understanding COVID-19. Immunity 2020, 52, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Nutrition, Immunity and COVID-19. BMJ Nutr. Prev. Health 2020, 3, 74–92. [Google Scholar] [CrossRef] [PubMed]
- Solimando, A.G.; Susca, N.; Borrelli, P.; Prete, M.; Lauletta, G.; Pappagallo, F.; Buono, R.; Inglese, G.; Forina, B.M.; Bochicchio, D.; et al. Short-Term Variations in Neutrophil-to-Lymphocyte and Urea-to-Creatinine Ratios Anticipate Intensive Care Unit Admission of COVID-19 Patients in the Emergency Department. Front. Med. 2020, 7, 625176. [Google Scholar] [CrossRef] [PubMed]
- Bavaro, D.F.; Diella, L.; Solimando, A.G.; Cicco, S.; Buonamico, E.; Stasi, C.; Ciannarella, M.; Marrone, M.; Carpagnano, F.; Resta, O.; et al. Bamlanivimab and Etesevimab Administered in an Outpatient Setting for SARS-CoV-2 Infection. Pathog. Glob. Health 2022, 1–8. [Google Scholar] [CrossRef]
- Chevrier, S.; Zurbuchen, Y.; Cervia, C.; Adamo, S.; Raeber, M.E.; de Souza, N.; Sivapatham, S.; Jacobs, A.; Bachli, E.; Rudiger, A.; et al. A Distinct Innate Immune Signature Marks Progression from Mild to Severe COVID-19. Cell Rep. Med. 2021, 2, 100166. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention Factors That Affect Your Risk of Getting Very Sick from COVID-19 2022. Available online: https://www.cdc.gov/coronavirus/2019-ncov/your-health/risks-getting-very-sick.html (accessed on 11 May 2023).
- Caramelo, F.; Ferreira, N.; Oliveiros, B. Estimation of Risk Factors for COVID-19 Mortality—Preliminary Results. medRxiv Epidemiology 2020. [Google Scholar] [CrossRef]
- The Severe COVID-19 GWAS Group Genomewide Association Study of Severe COVID-19 with Respiratory Failure. N. Engl. J. Med. 2020, 383, 1522–1534. [CrossRef]
- Grifoni, A.; Sidney, J.; Vita, R.; Peters, B.; Crotty, S.; Weiskopf, D.; Sette, A. SARS-CoV-2 Human T Cell Epitopes: Adaptive Immune Response against COVID-19. Cell Host Microbe 2021, 29, 1076–1092. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Cassaniti, I.; Percivalle, E.; Bergami, F.; Piralla, A.; Comolli, G.; Bruno, R.; Vecchia, M.; Sambo, M.; Colaneri, M.; Zuccaro, V.; et al. SARS-CoV-2 Specific T-Cell Immunity in COVID-19 Convalescent Patients and Unexposed Controls Measured by Ex Vivo ELISpot Assay. Clin. Microbiol. Infect. 2021, 27, 1029–1034. [Google Scholar] [CrossRef] [PubMed]
- Rovida, F.; Cassaniti, I.; Percivalle, E.; Sarasini, A.; Paolucci, S.; Klersy, C.; Cutti, S.; Novelli, V.; Marena, C.; Luzzaro, F.; et al. Incidence of SARS-CoV-2 Infection in Health Care Workers from Northern Italy Based on Antibody Status: Immune Protection from Secondary Infection- A Retrospective Observational Case-Controlled Study. Int. J. Infect. Dis. 2021, 109, 199–202. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and Efficacy of the ChAdOx1 NCoV-19 Vaccine (AZD1222) against SARS-CoV-2: An Interim Analysis of Four Randomised Controlled Trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Peng, Y.; Mentzer, A.J.; Liu, G.; Yao, X.; Yin, Z.; Dong, D.; Dejnirattisai, W.; Rostron, T.; Supasa, P.; Liu, C.; et al. Broad and Strong Memory CD4+ and CD8+ T Cells Induced by SARS-CoV-2 in UK Convalescent Individuals Following COVID-19. Nat. Immunol. 2020, 21, 1336–1345. [Google Scholar] [CrossRef]
- Moss, P. The T Cell Immune Response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef]
- Telenti, A.; Arvin, A.; Corey, L.; Corti, D.; Diamond, M.S.; García-Sastre, A.; Garry, R.F.; Holmes, E.C.; Pang, P.S.; Virgin, H.W. After the Pandemic: Perspectives on the Future Trajectory of COVID-19. Nature 2021, 596, 495–504. [Google Scholar] [CrossRef]
- Callaway, E. Beyond Omicron: What’s next for COVID’s Viral Evolution. Nature 2021, 600, 204–207. [Google Scholar] [CrossRef]
- Schultze, J.L.; Aschenbrenner, A.C. COVID-19 and the Human Innate Immune System. Cell 2021, 184, 1671–1692. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.S.; Kanneganti, T.-D. Innate Immunity: The First Line of Defense against SARS-CoV-2. Nat. Immunol. 2022, 23, 165–176. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Huang, S.; Yin, L. The Cytokine Storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef] [PubMed]
- Weyand, C.M.; Goronzy, J.J. Aging of the Immune System. Mechanisms and Therapeutic Targets. Ann. Am. Thorac. Soc. 2016, 13 (Suppl. 5), S422–S428. [Google Scholar] [CrossRef]
- Nikolich-Zugich, J.; Goodrum, F.; Knox, K.; Smithey, M.J. Known Unknowns: How Might the Persistent Herpesvirome Shape Immunity and Aging? Curr. Opin. Immunol. 2017, 48, 23–30. [Google Scholar] [CrossRef]
- Lissner, M.M.; Thomas, B.J.; Wee, K.; Tong, A.-J.; Kollmann, T.R.; Smale, S.T. Age-Related Gene Expression Differences in Monocytes from Human Neonates, Young Adults, and Older Adults. PLoS ONE 2015, 10, e0132061. [Google Scholar] [CrossRef]
- Fasano, R.; Malerba, E.; Prete, M.; Solimando, A.G.; Buonavoglia, A.; Silvestris, N.; Leone, P.; Racanelli, V. Impact of Antigen Presentation Mechanisms on Immune Response in Autoimmune Hepatitis. Front. Immunol. 2021, 12, 814155. [Google Scholar] [CrossRef]
- Brunetta, E.; Folci, M.; Bottazzi, B.; De Santis, M.; Gritti, G.; Protti, A.; Mapelli, S.N.; Bonovas, S.; Piovani, D.; Leone, R.; et al. Macrophage Expression and Prognostic Significance of the Long Pentraxin PTX3 in COVID-19. Nat. Immunol. 2021, 22, 19–24. [Google Scholar] [CrossRef]
- WHO Rapid Evidence Appraisal for COVID-19 Therapies (REACT) Working Group; Sterne, J.A.C.; Murthy, S.; Diaz, J.V.; Slutsky, A.S.; Villar, J.; Angus, D.C.; Annane, D.; Azevedo, L.C.P.; Berwanger, O.; et al. Association between Administration of Systemic Corticosteroids and Mortality among Critically Ill Patients with COVID-19: A Meta-Analysis. JAMA 2020, 324, 1330–1341. [Google Scholar] [CrossRef]
- Kalimuddin, S.; Tham, C.Y.L.; Qui, M.; de Alwis, R.; Sim, J.X.Y.; Lim, J.M.E.; Tan, H.-C.; Syenina, A.; Zhang, S.L.; Le Bert, N.; et al. Early T Cell and Binding Antibody Responses Are Associated with COVID-19 RNA Vaccine Efficacy Onset. Med 2021, 2, 682–688.e4. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological Memory to SARS-CoV-2 Assessed for up to 8 Months after Infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Vizcarra, P.; Pérez-Elías, M.J.; Quereda, C.; Moreno, A.; Vivancos, M.J.; Dronda, F.; Casado, J.L.; COVID-19 ID Team. Description of COVID-19 in HIV-Infected Individuals: A Single-Centre, Prospective Cohort. Lancet HIV 2020, 7, e554–e564. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.-E.; Peluso, M.J.; Margus, C.; Matias Lopes, J.P.; He, C.; Gaisa, M.M.; Osorio, G.; Aberg, J.A.; Mullen, M.P. Clinical Outcomes and Immunologic Characteristics of Coronavirus Disease 2019 in People With Human Immunodeficiency Virus. J. Infect. Dis. 2021, 223, 403–408. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.-A. HIV and Risk of COVID-19 Death: A Population Cohort Study from the Western Cape Province, South Africa. MedRxiv Prepr. Serv. Health Sci. 2020. [Google Scholar] [CrossRef]
- Chenneville, T.; Gabbidon, K.; Hanson, P.; Holyfield, C. The Impact of COVID-19 on HIV Treatment and Research: A Call to Action. Int. J. Environ. Res. Public. Health 2020, 17, 4548. [Google Scholar] [CrossRef]
- Mantovani, A.; Netea, M.G. Trained Innate Immunity, Epigenetics, and COVID-19. N. Engl. J. Med. 2020, 383, 1078–1080. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Brown, E.S.; Cheeseman, H.M.; Flight, K.E.; Higham, S.L.; Lemm, N.-M.; Pierce, B.F.; Stirling, D.C.; Wang, Z.; Pollock, K.M. Vaccines for COVID-19. Clin. Exp. Immunol. 2020, 202, 162–192. [Google Scholar] [CrossRef]
- Cassaniti, I.; Bergami, F.; Percivalle, E.; Gabanti, E.; Sammartino, J.C.; Ferrari, A.; Adzasehoun, K.M.G.; Zavaglio, F.; Zelini, P.; Comolli, G.; et al. Humoral and Cell-Mediated Response against SARS-CoV-2 Variants Elicited by MRNA Vaccine BNT162b2 in Healthcare Workers: A Longitudinal Observational Study. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2022, 28, 301.e1–301.e8. [Google Scholar] [CrossRef]
- Mateus, J.; Dan, J.M.; Zhang, Z.; Rydyznski Moderbacher, C.; Lammers, M.; Goodwin, B.; Sette, A.; Crotty, S.; Weiskopf, D. Low-Dose MRNA-1273 COVID-19 Vaccine Generates Durable Memory Enhanced by Cross-Reactive T Cells. Science 2021, 374, eabj9853. [Google Scholar] [CrossRef]
- Self, W.H.; Tenforde, M.W.; Rhoads, J.P.; Gaglani, M.; Ginde, A.A.; Douin, D.J.; Olson, S.M.; Talbot, H.K.; Casey, J.D.; Mohr, N.M.; et al. Comparative Effectiveness of Moderna, Pfizer-BioNTech, and Janssen (Johnson & Johnson) Vaccines in Preventing COVID-19 Hospitalizations among Adults without Immunocompromising Conditions - United States, March-August 2021. MMWR Morb. Mortal. Wkly. Rep. 2021, 70, 1337–1343. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.; Shao, W.; Chen, X.; Zhang, B.; Wang, G.; Zhang, W. Real-World Effectiveness of COVID-19 Vaccines: A Literature Review and Meta-Analysis. Int. J. Infect. Dis. 2022, 114, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Mateus, J.; Coelho, C.H.; Dan, J.M.; Moderbacher, C.R.; Gálvez, R.I.; Cortes, F.H.; Grifoni, A.; Tarke, A.; Chang, J.; et al. Humoral and Cellular Immune Memory to Four COVID-19 Vaccines. Cell 2022, 185, 2434–2451.e17. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Methot, N.; Yu, E.D.; Zhang, Y.; Dan, J.M.; Goodwin, B.; Rubiro, P.; Sutherland, A.; Wang, E.; et al. Impact of SARS-CoV-2 Variants on the Total CD4+ and CD8+ T Cell Reactivity in Infected or Vaccinated Individuals. Cell Rep. Med. 2021, 2, 100355. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Sheng, Y.; Huang, C.; Jin, Y.; Xiong, N.; Jiang, K.; Lu, H.; Liu, J.; Yang, J.; Dong, Y.; et al. Clinical Characteristics, Outcomes, and Risk Factors for Mortality in Patients with Cancer and COVID-19 in Hubei, China: A Multicentre, Retrospective, Cohort Study. Lancet Oncol. 2020, 21, 904–913. [Google Scholar] [CrossRef]
- Tian, J.; Yuan, X.; Xiao, J.; Zhong, Q.; Yang, C.; Liu, B.; Cai, Y.; Lu, Z.; Wang, J.; Wang, Y.; et al. Clinical Characteristics and Risk Factors Associated with COVID-19 Disease Severity in Patients with Cancer in Wuhan, China: A Multicentre, Retrospective, Cohort Study. Lancet Oncol. 2020, 21, 893–903. [Google Scholar] [CrossRef]
- Zhang, L.; Zhu, F.; Xie, L.; Wang, C.; Wang, J.; Chen, R.; Jia, P.; Guan, H.Q.; Peng, L.; Chen, Y.; et al. Clinical Characteristics of COVID-19-Infected Cancer Patients: A Retrospective Case Study in Three Hospitals within Wuhan, China. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2020, 31, 894–901. [Google Scholar] [CrossRef]
- Vuagnat, P.; Frelaut, M.; Ramtohul, T.; Basse, C.; Diakite, S.; Noret, A.; Bellesoeur, A.; Servois, V.; Hequet, D.; Laas, E.; et al. COVID-19 in Breast Cancer Patients: A Cohort at the Institut Curie Hospitals in the Paris Area. Breast Cancer Res. BCR 2020, 22, 55. [Google Scholar] [CrossRef]
- Assaad, S.; Avrillon, V.; Fournier, M.-L.; Mastroianni, B.; Russias, B.; Swalduz, A.; Cassier, P.; Eberst, L.; Steineur, M.-P.; Kazes, M.; et al. High Mortality Rate in Cancer Patients with Symptoms of COVID-19 with or without Detectable SARS-CoV-2 on RT-PCR. Eur. J. Cancer Oxf. Engl. 1990 2020, 135, 251–259. [Google Scholar] [CrossRef]
- Kuderer, N.M.; Choueiri, T.K.; Shah, D.P.; Shyr, Y.; Rubinstein, S.M.; Rivera, D.R.; Shete, S.; Hsu, C.-Y.; Desai, A.; de Lima Lopes, G.; et al. Clinical Impact of COVID-19 on Patients with Cancer (CCC19): A Cohort Study. Lancet 2020, 395, 1907–1918. [Google Scholar] [CrossRef]
- Garassino, M.C.; Whisenant, J.G.; Huang, L.-C.; Trama, A.; Torri, V.; Agustoni, F.; Baena, J.; Banna, G.; Berardi, R.; Bettini, A.C.; et al. COVID-19 in Patients with Thoracic Malignancies (TERAVOLT): First Results of an International, Registry-Based, Cohort Study. Lancet Oncol. 2020, 21, 914–922. [Google Scholar] [CrossRef]
- Boulad, F.; Kamboj, M.; Bouvier, N.; Mauguen, A.; Kung, A.L. COVID-19 in Children with Cancer in New York City. JAMA Oncol. 2020, 6, 1459–1460. [Google Scholar] [CrossRef]
- Miyashita, H.; Mikami, T.; Chopra, N.; Yamada, T.; Chernyavsky, S.; Rizk, D.; Cruz, C. Do Patients with Cancer Have a Poorer Prognosis of COVID-19? An Experience in New York City. Ann. Oncol. 2020, 31, 1088–1089. [Google Scholar] [CrossRef] [PubMed]
- Lisco, G.; De Tullio, A.; Stragapede, A.; Solimando, A.G.; Albanese, F.; Capobianco, M.; Giagulli, V.A.; Guastamacchia, E.; De Pergola, G.; Vacca, A.; et al. COVID-19 and the Endocrine System: A Comprehensive Review on the Theme. J. Clin. Med. 2021, 10, 2920. [Google Scholar] [CrossRef]
- Solimando, A.G.; Crudele, L.; Leone, P.; Argentiero, A.; Guarascio, M.; Silvestris, N.; Vacca, A.; Racanelli, V. Immune Checkpoint Inhibitor-Related Myositis: From Biology to Bedside. Int. J. Mol. Sci. 2020, 21, 3054. [Google Scholar] [CrossRef] [PubMed]
- Lasagna, A.; Agustoni, F.; Percivalle, E.; Borgetto, S.; Paulet, A.; Comolli, G.; Sarasini, A.; Bergami, F.; Sammartino, J.C.; Ferrari, A.; et al. A Snapshot of the Immunogenicity, Efficacy and Safety of a Full Course of BNT162b2 Anti-SARS-CoV-2 Vaccine in Cancer Patients Treated with PD-1/PD-L1 Inhibitors: A Longitudinal Cohort Study. ESMO Open 2021, 6, 100272. [Google Scholar] [CrossRef] [PubMed]
- De Gasparo, R.; Pedotti, M.; Simonelli, L.; Nickl, P.; Muecksch, F.; Cassaniti, I.; Percivalle, E.; Lorenzi, J.C.C.; Mazzola, F.; Magrì, D.; et al. Bispecific IgG Neutralizes SARS-CoV-2 Variants and Prevents Escape in Mice. Nature 2021, 593, 424–428. [Google Scholar] [CrossRef]
- Marra, A.; Generali, D.; Zagami, P.; Cervoni, V.; Gandini, S.; Venturini, S.; Morganti, S.; Passerini, R.; Orecchia, R.; Curigliano, G. Seroconversion in Patients with Cancer and Oncology Health Care Workers Infected by SARS-CoV-2. Ann. Oncol. 2021, 32, 113–119. [Google Scholar] [CrossRef]
- Kamar, N.; Abravanel, F.; Marion, O.; Couat, C.; Izopet, J.; Del Bello, A. Three Doses of an MRNA COVID-19 Vaccine in Solid-Organ Transplant Recipients. N. Engl. J. Med. 2021, 385, 661–662. [Google Scholar] [CrossRef]
- Yasin, A.I.; Aydin, S.G.; Sümbül, B.; Koral, L.; Şimşek, M.; Geredeli, Ç.; Öztürk, A.; Perkin, P.; Demirtaş, D.; Erdemoglu, E.; et al. Efficacy and Safety Profile of COVID-19 Vaccine in Cancer Patients: A Prospective, Multicenter Cohort Study. Future Oncol. Lond. Engl. 2022, 18, 1235–1244. [Google Scholar] [CrossRef]
- Iacono, D.; Cerbone, L.; Palombi, L.; Cavalieri, E.; Sperduti, I.; Cocchiara, R.A.; Mariani, B.; Parisi, G.; Garufi, C. Serological Response to COVID-19 Vaccination in Patients with Cancer Older than 80 Years. J. Geriatr. Oncol. 2021, 12, 1253–1255. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Zhang, D.; Li, Z.; Zhang, K. Predictors of Poor Serologic Response to COVID-19 Vaccine in Patients with Cancer: A Systematic Review and Meta-Analysis. Eur. J. Cancer Oxf. Engl. 1990 2022, 172, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Shahid, Z.; Patrick, A.L.; Wallander, M.L.; Donahue, E.E.; Trufan, S.J.; Tan, A.R.; Hwang, J.J.; Burgess, E.F.; Ragon, B.; Ghosh, N.; et al. COVID-19 Vaccine Uptake Trends in SARS-CoV-2 Previously Infected Cancer Patients. Vaccine X 2023, 14, 100289. [Google Scholar] [CrossRef] [PubMed]
- Mai, A.S.; Lee, A.R.Y.B.; Tay, R.Y.K.; Shapiro, L.; Thakkar, A.; Halmos, B.; Grinshpun, A.; Herishanu, Y.; Benjamini, O.; Tadmor, T.; et al. Booster Doses of COVID-19 Vaccines for Patients with Haematological and Solid Cancer: A Systematic Review and Individual Patient Data Meta-Analysis. Eur. J. Cancer Oxf. Engl. 1990 2022, 172, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Javadinia, S.A.; Alizadeh, K.; Mojadadi, M.-S.; Nikbakht, F.; Dashti, F.; Joudi, M.; Harati, H.; Welsh, J.S.; Farahmand, S.A.; Attarian, F. COVID-19 Vaccination in Patients With Malignancy; A Systematic Review and Meta-Analysis of the Efficacy and Safety. Front. Endocrinol. 2022, 13, 860238. [Google Scholar] [CrossRef]
- Desantis, V.; Saltarella, I.; Lamanuzzi, A.; Melaccio, A.; Solimando, A.G.; Mariggiò, M.A.; Racanelli, V.; Paradiso, A.; Vacca, A.; Frassanito, M.A. MicroRNAs-Based Nano-Strategies as New Therapeutic Approach in Multiple Myeloma to Overcome Disease Progression and Drug Resistance. Int. J. Mol. Sci. 2020, 21, 3084. [Google Scholar] [CrossRef]
- Vacca, A.; Melaccio, A.; Sportelli, A.; Solimando, A.G.; Dammacco, F.; Ria, R. Subcutaneous Immunoglobulins in Patients with Multiple Myeloma and Secondary Hypogammaglobulinemia: A Randomized Trial. Clin. Immunol. Orlando Fla 2018, 191, 110–115. [Google Scholar] [CrossRef]
- Gavriatopoulou, M.; Terpos, E.; Malandrakis, P.; Ntanasis-Stathopoulos, I.; Briasoulis, A.; Gumeni, S.; Fotiou, D.; Papanagnou, E.-D.; Migkou, M.; Theodorakakou, F.; et al. Myeloma Patients with COVID-19 Have Superior Antibody Responses Compared to Patients Fully Vaccinated with the BNT162b2 Vaccine. Br. J. Haematol. 2022, 196, 356–359. [Google Scholar] [CrossRef]
- Ballman, M.; Swift, S.; Mullenix, C.; Mallory, Y.; Zhao, C.; Szabo, E.; Shelat, M.; Sansone, S.; Steinberg, S.M.; McAdams, M.J.; et al. Tolerability of Coronavirus Disease 2019 Vaccines, BNT162b2 and MRNA-1273, in Patients with Thymic Epithelial Tumors. JTO Clin. Res. Rep. 2021, 2, 100229. [Google Scholar] [CrossRef]
- Trapani, D.; Curigliano, G. COVID-19 Vaccines in Patients with Cancer. Lancet Oncol. 2021, 22, 738–739. [Google Scholar] [CrossRef]
- Shahini, E.; Pesce, F.; Argentiero, A.; Solimando, A.G. Can Vitamin D Status Influence Seroconversion to SARS-CoV2 Vaccines? Front. Immunol. 2022, 13, 1038316. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Torres, V.; De Mendoza, C.; Mellor-Pita, S.; Martínez-Urbistondo, M.; Durán-del Campo, P.; Tutor-Ureta, P.; Vázquez-Comendador, J.-M.; Calderón-Parra, J.; Múñez-Rubio, E.; Ramos-Martínez, A.; et al. Systemic Autoimmune Diseases in Patients Hospitalized with COVID-19 in Spain: A Nation-Wide Registry Study. Viruses 2022, 14, 1631. [Google Scholar] [CrossRef] [PubMed]
- Brito-Zerón, P.; Sisó-Almirall, A.; Flores-Chavez, A.; Retamozo, S.; Ramos-Casals, M. SARS-CoV-2 Infection in Patients with Systemic Autoimmune Diseases. Clin. Exp. Rheumatol. 2021, 39, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Gianfrancesco, M.; Yazdany, J.; Robinson, P.C. Epidemiology and Outcomes of Novel Coronavirus 2019 in Patients with Immune-Mediated Inflammatory Diseases. Curr. Opin. Rheumatol. 2020, 32, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Gianfrancesco, M.; Hyrich, K.L.; Al-Adely, S.; Carmona, L.; Danila, M.I.; Gossec, L.; Izadi, Z.; Jacobsohn, L.; Katz, P.; Lawson-Tovey, S.; et al. Characteristics Associated with Hospitalisation for COVID-19 in People with Rheumatic Disease: Data from the COVID-19 Global Rheumatology Alliance Physician-Reported Registry. Ann. Rheum. Dis. 2020, 79, 859–866. [Google Scholar] [CrossRef]
- Haberman, R.H.; Um, S.; Axelrad, J.E.; Blank, R.B.; Uddin, Z.; Catron, S.; Neimann, A.L.; Mulligan, M.J.; Herat, R.S.; Hong, S.J.; et al. Methotrexate and TNF Inhibitors Affect Long-Term Immunogenicity to COVID-19 Vaccination in Patients with Immune-Mediated Inflammatory Disease. Lancet Rheumatol. 2022, 4, e384–e387. [Google Scholar] [CrossRef]
- Quartuccio, L.; Valent, F.; Pasut, E.; Tascini, C.; De Vita, S. Prevalence of COVID-19 among Patients with Chronic Inflammatory Rheumatic Diseases Treated with Biologic Agents or Small Molecules: A Population-Based Study in the First Two Months of COVID-19 Outbreak in Italy. Joint Bone Spine 2020, 87, 439–443. [Google Scholar] [CrossRef]
- Kotas, M.E.; Medzhitov, R. Homeostasis, Inflammation, and Disease Susceptibility. Cell 2015, 160, 816–827. [Google Scholar] [CrossRef]
- Solimando, A.G.; Vacca, A.; Ribatti, D. Inborn Error of Immunity: A Journey Through Novel Genes and Clinical Presentation. In Encyclopedia of Infection and Immunity; Elsevier: Amsterdam, The Netherlands, 2022; pp. 798–818. ISBN 978-0-323-90303-5. [Google Scholar]
- Solimando, A.G.; Desantis, V.; Palumbo, C.; Marasco, C.; Pappagallo, F.; Montagnani, M.; Ingravallo, G.; Cicco, S.; Di Paola, R.; Tabares, P.; et al. STAT1 Overexpression Triggers Aplastic Anemia: A Pilot Study Unravelling Novel Pathogenetic Insights in Bone Marrow Failure. Clin. Exp. Med. 2023. [Google Scholar] [CrossRef]
- Murdaca, G.; Paladin, F.; Tonacci, A.; Borro, M.; Greco, M.; Gerosa, A.; Isola, S.; Allegra, A.; Gangemi, S. Involvement of Il-33 in the Pathogenesis and Prognosis of Major Respiratory Viral Infections: Future Perspectives for Personalized Therapy. Biomedicines 2022, 10, 715. [Google Scholar] [CrossRef]
- Murdaca, G.; Greco, M.; Tonacci, A.; Negrini, S.; Borro, M.; Puppo, F.; Gangemi, S. IL-33/IL-31 Axis in Immune-Mediated and Allergic Diseases. Int. J. Mol. Sci. 2019, 20, 5856. [Google Scholar] [CrossRef] [PubMed]
- Bossuyt, X.; Vulsteke, J.-B.; Van Elslande, J.; Boon, L.; Wuyts, G.; Willebrords, S.; Frans, G.; Geukens, N.; Carpentier, S.; Tejpar, S.; et al. Antinuclear Antibodies in Individuals with COVID-19 Reflect Underlying Disease: Identification of New Autoantibodies in Systemic Sclerosis (CDK9) and Malignancy (RNF20, RCC1, TRIP13). Autoimmun. Rev. 2023, 22, 103288. [Google Scholar] [CrossRef]
- Böröcz, K.; Kinyó, Á.; Simon, D.; Erdő-Bonyár, S.; Németh, P.; Berki, T. Complexity of the Immune Response Elicited by Different COVID-19 Vaccines, in the Light of Natural Autoantibodies and Immunomodulatory Therapies. Int. J. Mol. Sci. 2023, 24, 6439. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, Y.; Liu, M.; Shi, S.; Tian, J. Impacts of Immunosuppression and Immunodeficiency on COVID-19: A Systematic Review and Meta-Analysis. J. Infect. 2020, 81, e93–e95. [Google Scholar] [CrossRef] [PubMed]
- Quinti, I.; Lougaris, V.; Milito, C.; Cinetto, F.; Pecoraro, A.; Mezzaroma, I.; Mastroianni, C.M.; Turriziani, O.; Bondioni, M.P.; Filippini, M.; et al. A Possible Role for B Cells in COVID-19? Lesson from Patients with Agammaglobulinemia. J. Allergy Clin. Immunol. 2020, 146, 211–213.e4. [Google Scholar] [CrossRef]
- Soresina, A.; Moratto, D.; Chiarini, M.; Paolillo, C.; Baresi, G.; Focà, E.; Bezzi, M.; Baronio, B.; Giacomelli, M.; Badolato, R. Two X-Linked Agammaglobulinemia Patients Develop Pneumonia as COVID-19 Manifestation but Recover. Pediatr. Allergy Immunol. Off. Publ. Eur. Soc. Pediatr. Allergy Immunol. 2020, 31, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Roschewski, M.; Lionakis, M.S.; Sharman, J.P.; Roswarski, J.; Goy, A.; Monticelli, M.A.; Roshon, M.; Wrzesinski, S.H.; Desai, J.V.; Zarakas, M.A.; et al. Inhibition of Bruton Tyrosine Kinase in Patients with Severe COVID-19. Sci. Immunol. 2020, 5, eabd0110. [Google Scholar] [CrossRef]
- Jenner, A.L.; Aogo, R.A.; Alfonso, S.; Crowe, V.; Deng, X.; Smith, A.P.; Morel, P.A.; Davis, C.L.; Smith, A.M.; Craig, M. COVID-19 Virtual Patient Cohort Suggests Immune Mechanisms Driving Disease Outcomes. PLoS Pathog. 2021, 17, e1009753. [Google Scholar] [CrossRef]
- Meyts, I.; Bucciol, G.; Quinti, I.; Neven, B.; Fischer, A.; Seoane, E.; Lopez-Granados, E.; Gianelli, C.; Robles-Marhuenda, A.; Jeandel, P.-Y.; et al. Coronavirus Disease 2019 in Patients with Inborn Errors of Immunity: An International Study. J. Allergy Clin. Immunol. 2021, 147, 520–531. [Google Scholar] [CrossRef]
- Solimando, A.G.; Annese, T.; Tamma, R.; Ingravallo, G.; Maiorano, E.; Vacca, A.; Specchia, G.; Ribatti, D. New Insights into Diffuse Large B-Cell Lymphoma Pathobiology. Cancers 2020, 12, 1869. [Google Scholar] [CrossRef]
- Shadbad, M.A.; Safaei, S.; Brunetti, O.; Derakhshani, A.; Lotfinejad, P.; Mokhtarzadeh, A.; Hemmat, N.; Racanelli, V.; Solimando, A.G.; Argentiero, A.; et al. A Systematic Review on the Therapeutic Potentiality of PD-L1-Inhibiting MicroRNAs for Triple-Negative Breast Cancer: Toward Single-Cell Sequencing-Guided Biomimetic Delivery. Genes 2021, 12, 1206. [Google Scholar] [CrossRef] [PubMed]
- Susca, N.; Solimando, A.G.; Borrelli, P.; Marziliano, D.; Monitillo, F.; Raimondo, P.; Vestito, D.; Lopizzo, A.; Brindicci, G.; Abumayyaleh, M.; et al. Electrocardiographic Pathological Findings Caused by the SARS-CoV-2 Virus Infection: Evidence from a Retrospective Multicenter International Cohort Longitudinal Pilot Study of 548 Subjects. J. Cardiovasc. Dev. Dis. 2023, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Gray, P.E.; Bartlett, A.W.; Tangye, S.G. Severe COVID-19 Represents an Undiagnosed Primary Immunodeficiency in a High Proportion of Infected Individuals. Clin. Transl. Immunol. 2022, 11, e1365. [Google Scholar] [CrossRef]
- Cazzato, G.; Cascardi, E.; Colagrande, A.; Foti, C.; Stellacci, A.; Marrone, M.; Ingravallo, G.; Arezzo, F.; Loizzi, V.; Solimando, A.G.; et al. SARS-CoV-2 and Skin: New Insights and Perspectives. Biomolecules 2022, 12, 1212. [Google Scholar] [CrossRef]
- Solimando, A.G.; Marziliano, D.; Ribatti, D. SARS-CoV-2 and Endothelial Cells: Vascular Changes, Intussusceptive Microvascular Growth and Novel Therapeutic Windows. Biomedicines 2022, 10, 2242. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.-A.K.; Moran, E.; Goodman, A.; Baxendale, H.; Bermingham, W.; Buckland, M.; AbdulKhaliq, I.; Jarvis, H.; Hunter, M.; Karanam, S.; et al. Treatment of Chronic or Relapsing COVID-19 in Immunodeficiency. J. Allergy Clin. Immunol. 2022, 149, 557–561.e1. [Google Scholar] [CrossRef]
- Milito, C.; Lougaris, V.; Giardino, G.; Punziano, A.; Vultaggio, A.; Carrabba, M.; Cinetto, F.; Scarpa, R.; Delle Piane, R.M.; Baselli, L.; et al. Clinical Outcome, Incidence, and SARS-CoV-2 Infection-Fatality Rates in Italian Patients with Inborn Errors of Immunity. J. Allergy Clin. Immunol. Pract. 2021, 9, 2904–2906.e2. [Google Scholar] [CrossRef]
- Giardino, G.; Milito, C.; Lougaris, V.; Punziano, A.; Carrabba, M.; Cinetto, F.; Scarpa, R.; Dellepiane, R.M.; Ricci, S.; Rivalta, B.; et al. The Impact of SARS-CoV-2 Infection in Patients with Inborn Errors of Immunity: The Experience of the Italian Primary Immunodeficiencies Network (IPINet). J. Clin. Immunol. 2022, 42, 935–946. [Google Scholar] [CrossRef]
- Rosenberg, J.M.; Peters, J.M.; Hughes, T.; Lareau, C.A.; Ludwig, L.S.; Massoth, L.R.; Austin-Tse, C.; Rehm, H.L.; Bryson, B.; Chen, Y.-B.; et al. JAK Inhibition in a Patient with a STAT1 Gain-of-Function Variant Reveals STAT1 Dysregulation as a Common Feature of Aplastic Anemia. Med 2022, 3, 42–57.e5. [Google Scholar] [CrossRef]
- Fattizzo, B.; Kulasekararaj, A.G.; Hill, A.; Benson-Quarm, N.; Griffin, M.; Munir, T.; Arnold, L.; Riley, K.; Ireland, R.; De Lavallade, H.; et al. Clinical and Morphological Predictors of Outcome in Older Aplastic Anemia Patients Treated with Eltrombopag. Haematologica 2019, 104, e494–e496. [Google Scholar] [CrossRef]
- Argentiero, A.; Solimando, A.G.; Brunetti, O.; Calabrese, A.; Pantano, F.; Iuliani, M.; Santini, D.; Silvestris, N.; Vacca, A. Skeletal Metastases of Unknown Primary: Biological Landscape and Clinical Overview. Cancers 2019, 11, 1270. [Google Scholar] [CrossRef] [PubMed]
- Notarangelo, L.D. Primary Immunodeficiencies (PIDs) Presenting with Cytopenias. Hematol. Am. Soc. Hematol. Educ. Program 2009, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Ferrarini, A.; Vacca, A.; Solimando, A.G.; Tavio, M.; Acquaviva, R.; Rocchi, M.; Nitti, C.; Salvi, A.; Menditto, V.; Luchetti Gentiloni, M.M.; et al. Early Administration of Tofacitinib in COVID-19 Pneumonitis: An Open Randomised Controlled Trial. Eur. J. Clin. Invest. 2022, 53, e13898. [Google Scholar] [CrossRef] [PubMed]
- Shields, A.M.; Burns, S.O.; Savic, S.; Richter, A.G.; Anantharachagan, A.; Arumugakani, G.; Baker, K.; Bahal, S.; Bermingham, W.; Bhole, M.; et al. COVID-19 in Patients with Primary and Secondary Immunodeficiency: The United Kingdom Experience. J. Allergy Clin. Immunol. 2021, 147, 870–875.e1. [Google Scholar] [CrossRef] [PubMed]


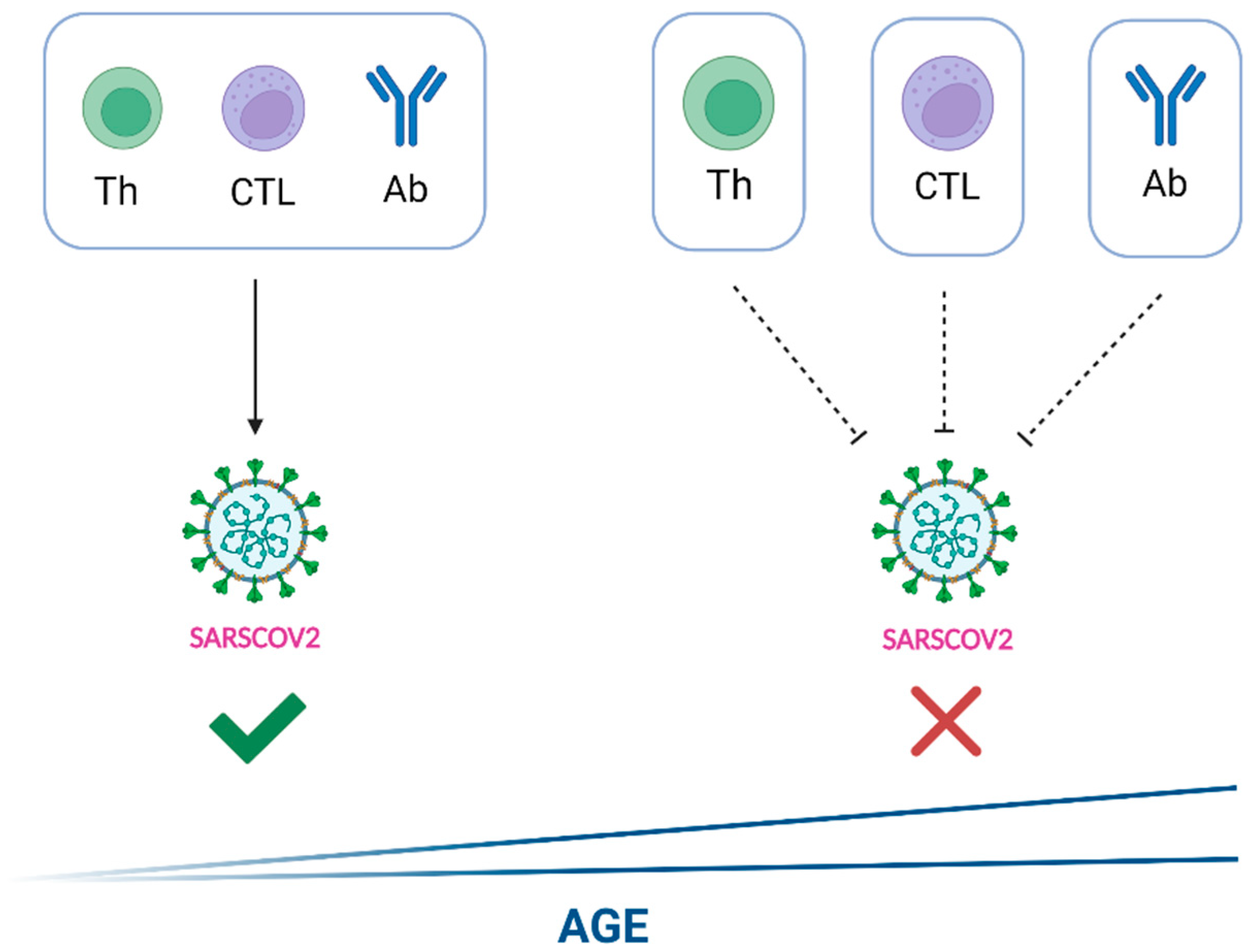{kind=link}
| Study | N, Neoplasms | Type | Risk Factors of Severe Outcome |
|---|---|---|---|
| China, Hubei [56] | 205, mixed | Retrospective | Hematological tumors, chemo <4 weeks, metastatic disease progression |
| China, Wuhan [57] | 232, mixed | Case-matched control | Advanced stage, ECOG, older patients |
| China, Wuhan [58] | 28, mixed | Retrospective | Antineoplastic tx <2 weeks, CT patchy consolidation |
| France, Paris [59] | 76, breast cancer | Prospective registry | Hypertension, older age |
| France, Lyon [60] | 302, mixed | Retrospective | Male gender, ECOG, PD cancer |
| CCC19 (USA, Canada, Spain) [61] | 928, mixed | Crowdsourcing | Age, male gender, NCDs, ECOG, PD cancer, smoking |
| TERAVOLT [62] | 200, thoracic cancer | Crowdsourcing | Smoking |
| US, NYC [63] | 20, childhood cancer | Retrospective | No increased risk of infection vs. non-cancer patients |
| US, NYC [64] | 334, mixed | Case-matched control | Older age |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solimando, A.G.; Bittrich, M.; Shahini, E.; Albanese, F.; Fritz, G.; Krebs, M. Determinants of COVID-19 Disease Severity–Lessons from Primary and Secondary Immune Disorders including Cancer. Int. J. Mol. Sci. 2023, 24, 8746. https://doi.org/10.3390/ijms24108746
Solimando AG, Bittrich M, Shahini E, Albanese F, Fritz G, Krebs M. Determinants of COVID-19 Disease Severity–Lessons from Primary and Secondary Immune Disorders including Cancer. International Journal of Molecular Sciences. 2023; 24(10):8746. https://doi.org/10.3390/ijms24108746
Chicago/Turabian StyleSolimando, Antonio G., Max Bittrich, Endrit Shahini, Federica Albanese, Georg Fritz, and Markus Krebs. 2023. "Determinants of COVID-19 Disease Severity–Lessons from Primary and Secondary Immune Disorders including Cancer" International Journal of Molecular Sciences 24, no. 10: 8746. https://doi.org/10.3390/ijms24108746