
Spleen Transcriptome Profiling Reveals Divergent Immune Responses to LPS and Poly (I:C) Challenge in the Yellow Drum (Nibea albiflora)

Abstract
:1. Introduction
2. Results
2.1. Data Filtering and Transcriptome Assembly
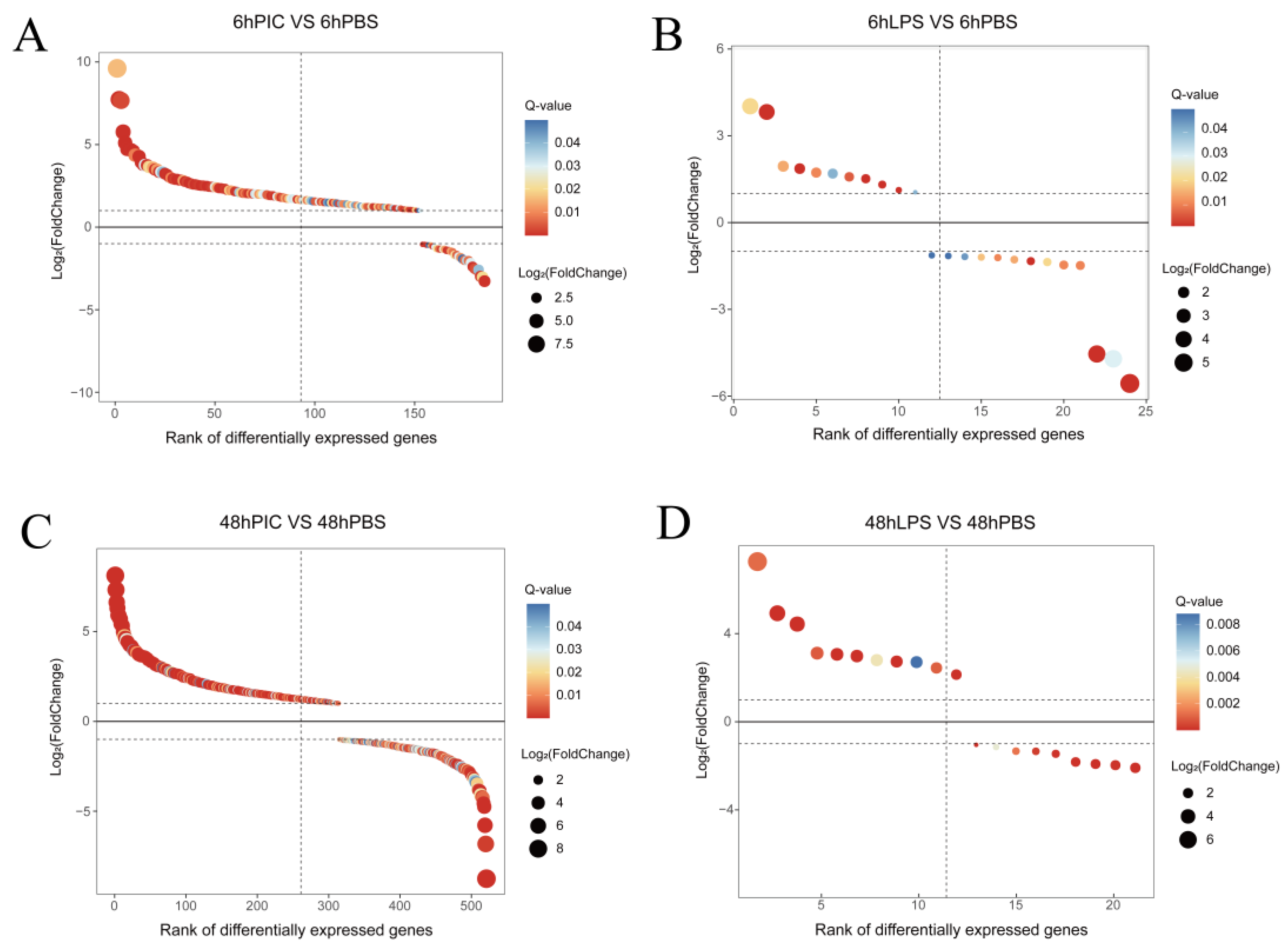
2.2. Gene Expression Quantification and Differentially Expressed Genes (DEGs) Screening
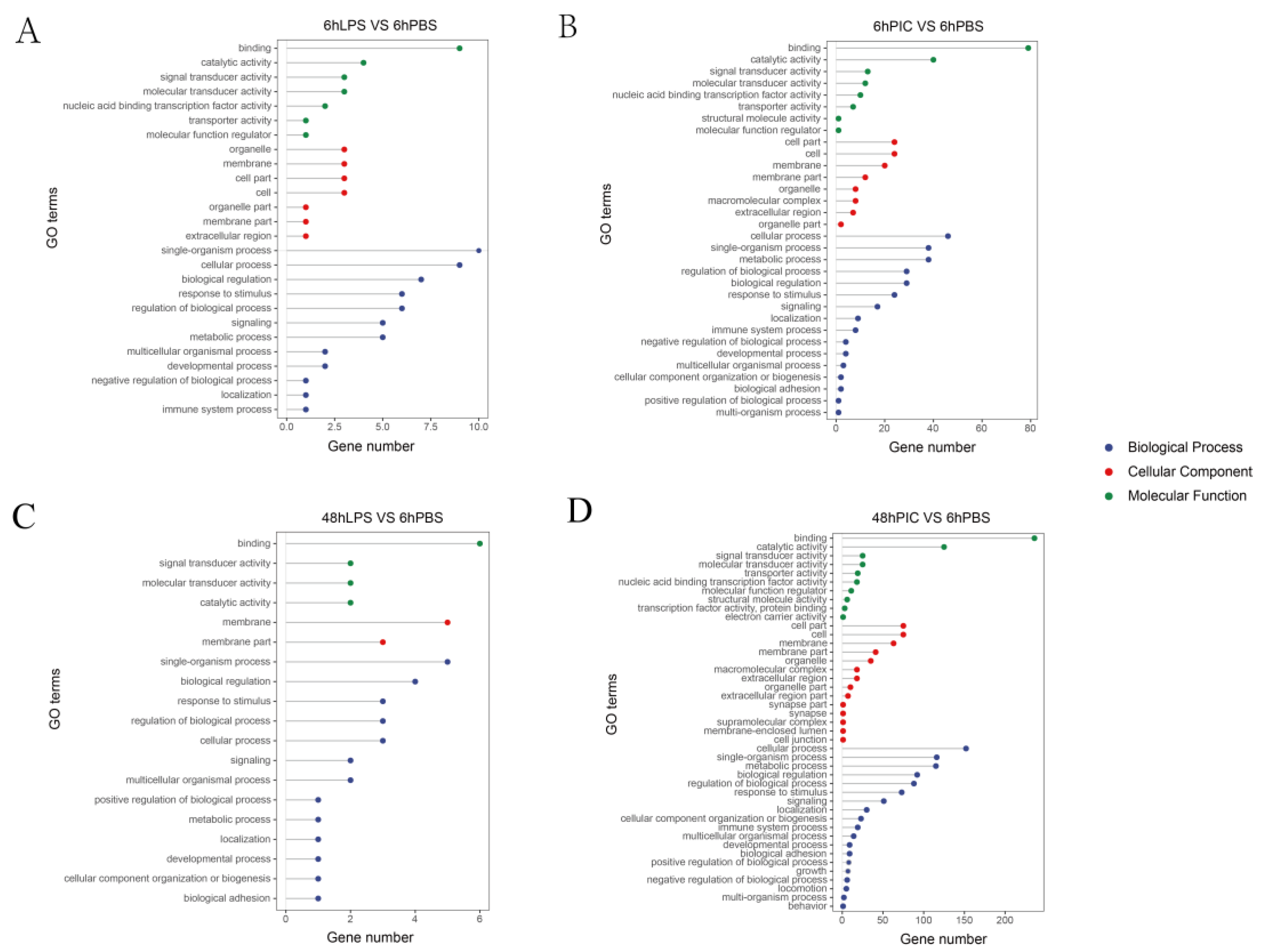
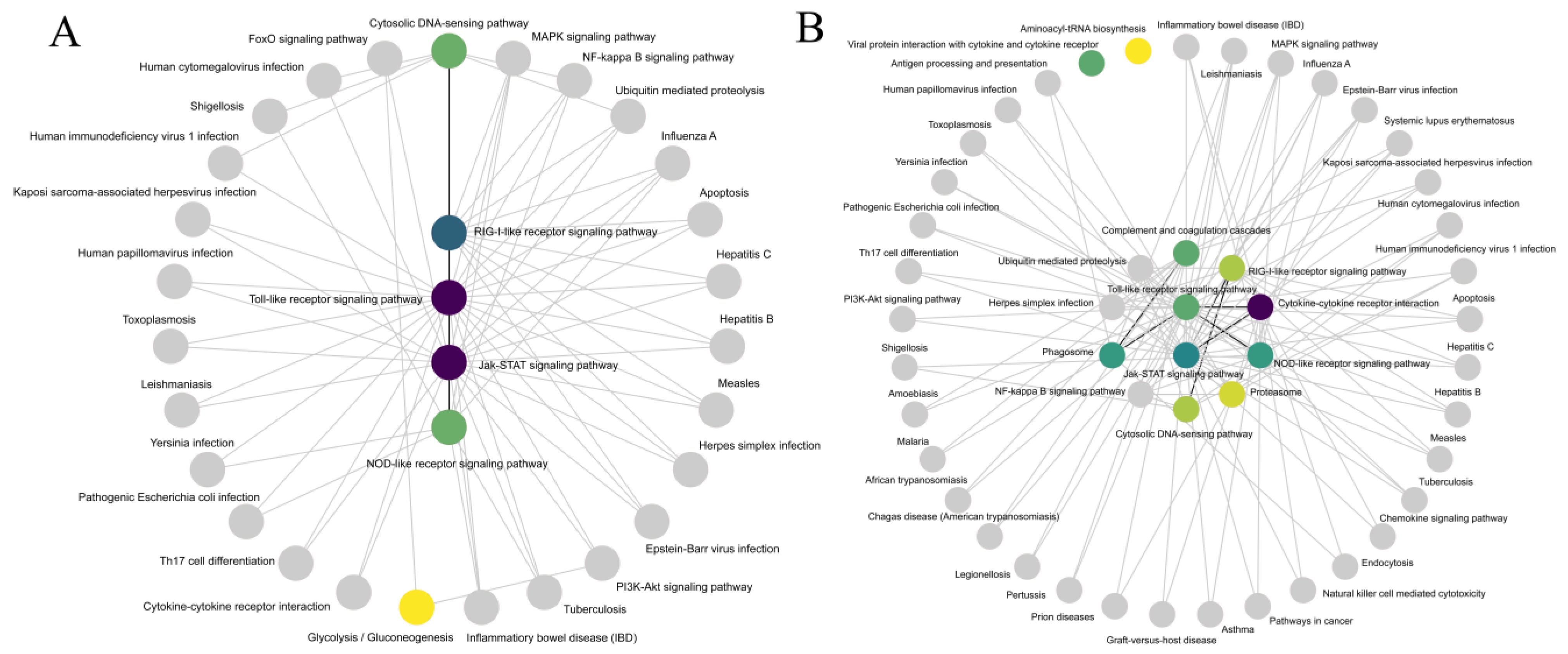
2.3. Gene Ontology (GO) Annotation and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of DEGs
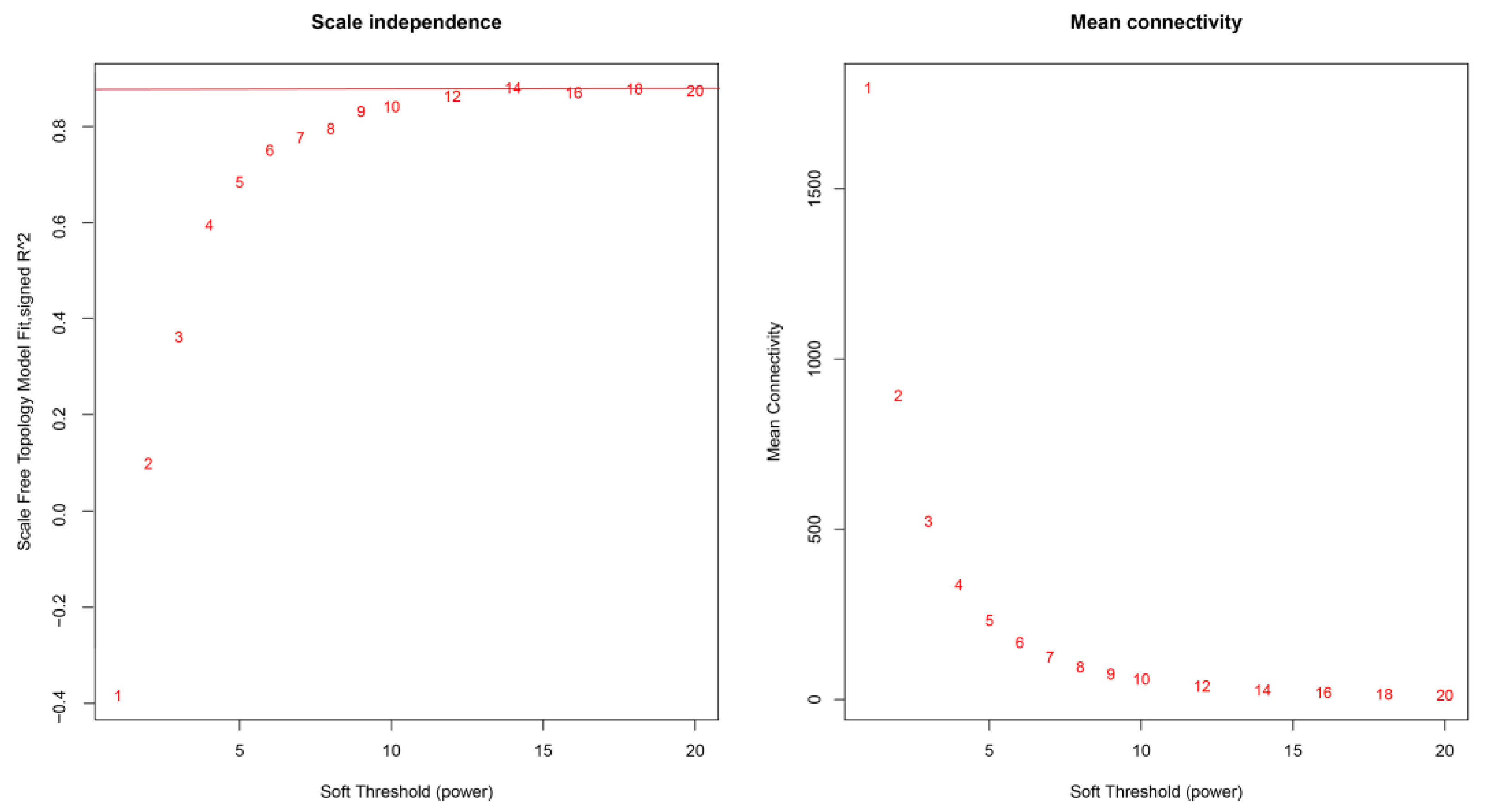
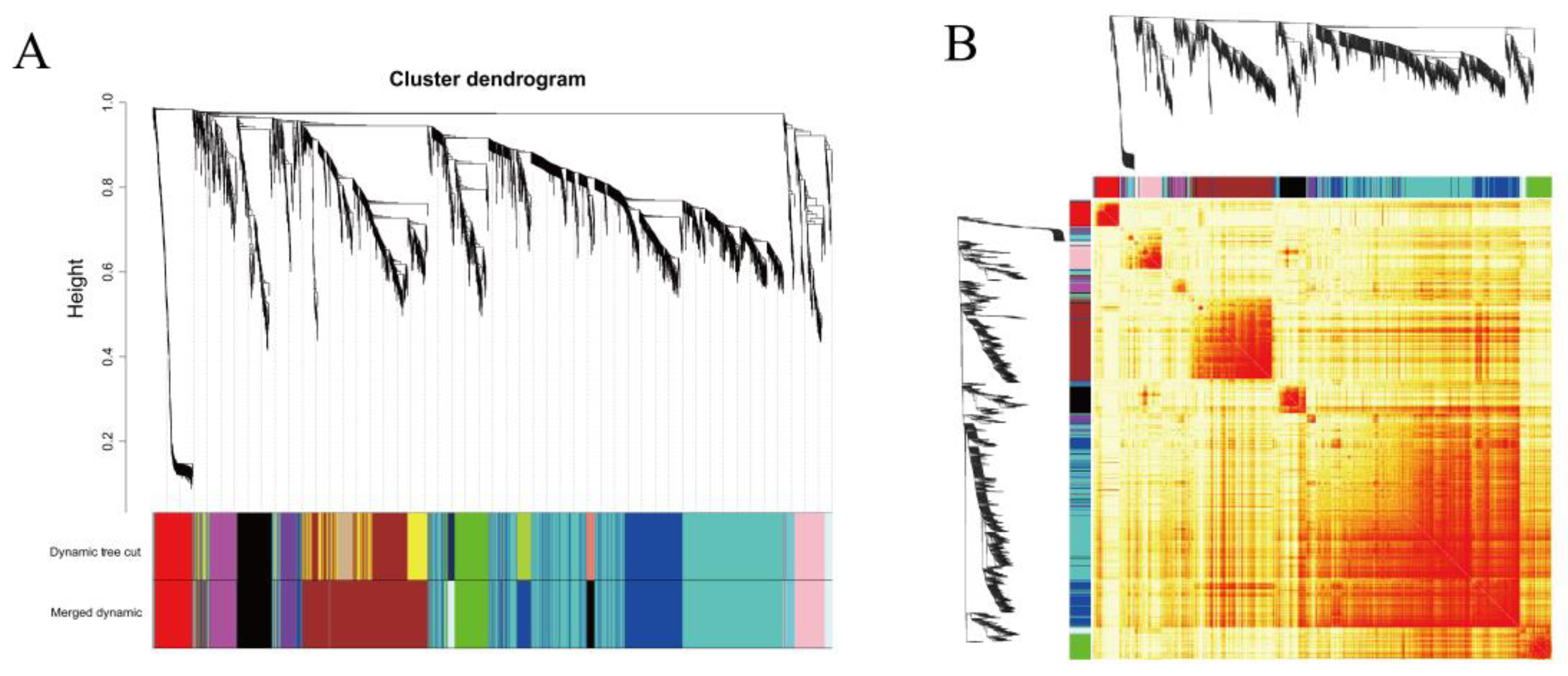
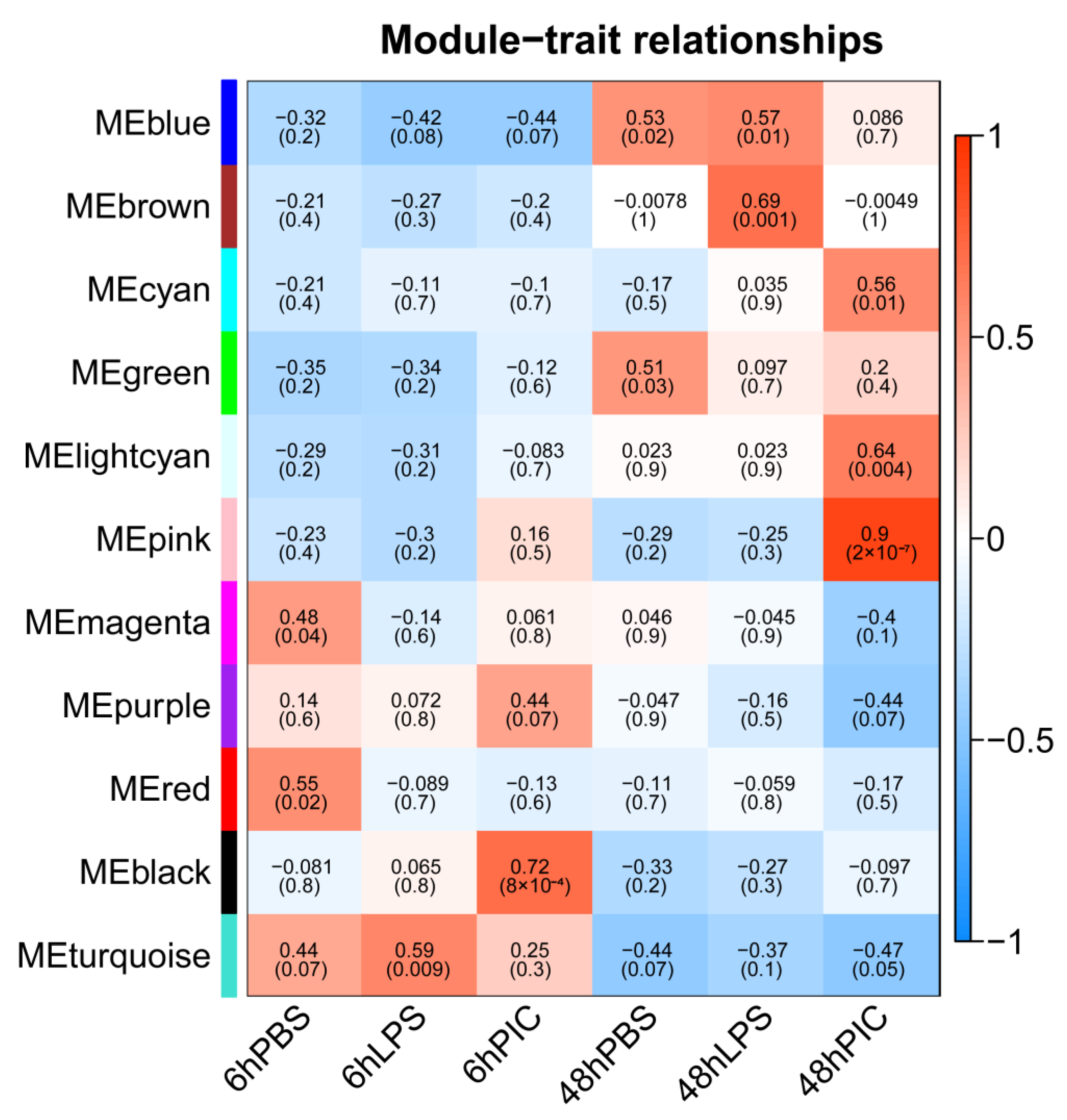
2.4. Co-Expression Network Construction and Key Gene Screening Using WGCNA
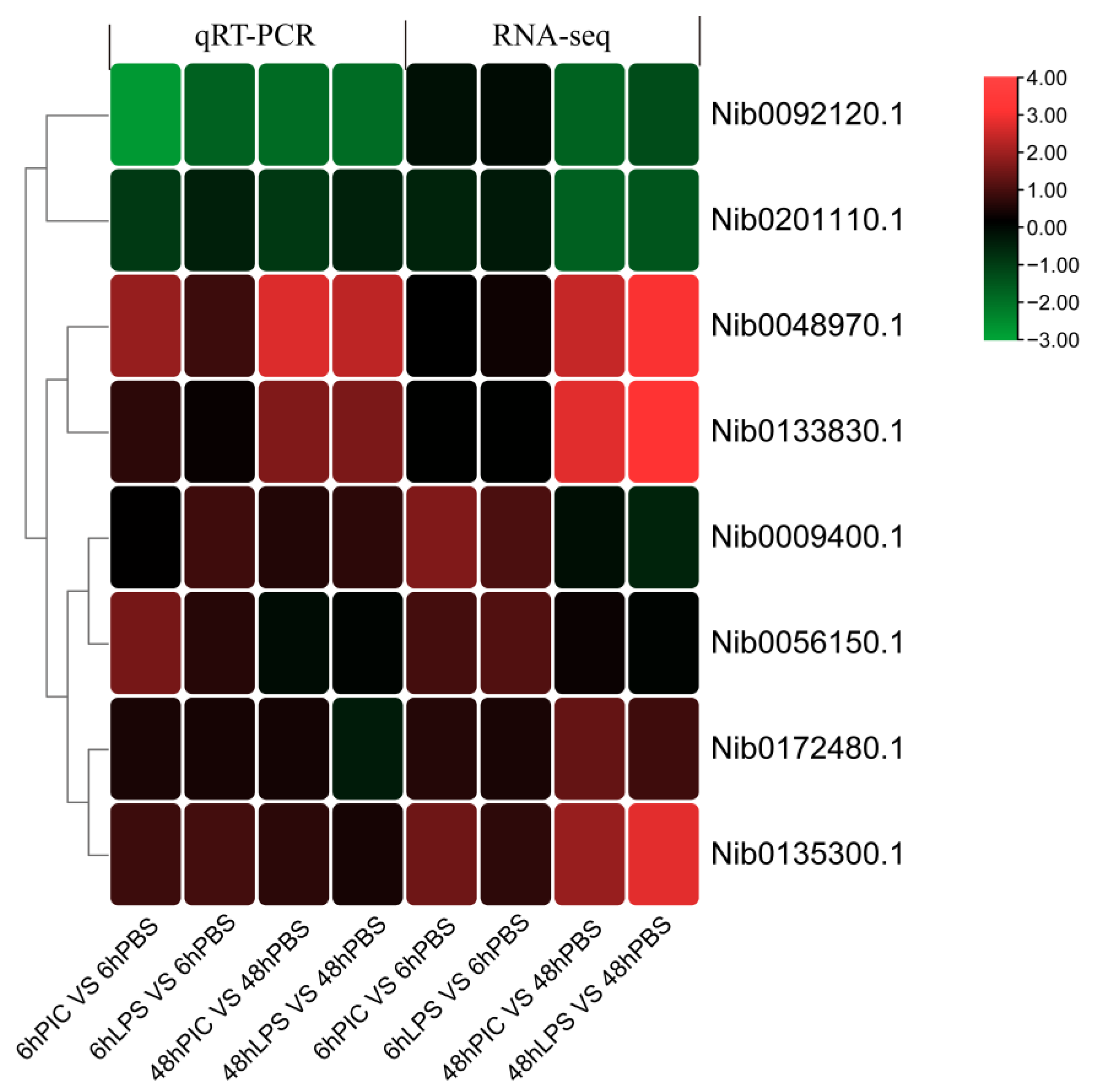
2.5. Validation of Transcriptomic Data via qRT-PCR
3. Discussion
3.1. Immunoregulatory Pathways and Key Genes under the LPS Injection Stress
3.2. Immunoregulatory Pathways and Key Genes under the Poly (I:C) Injection Stress
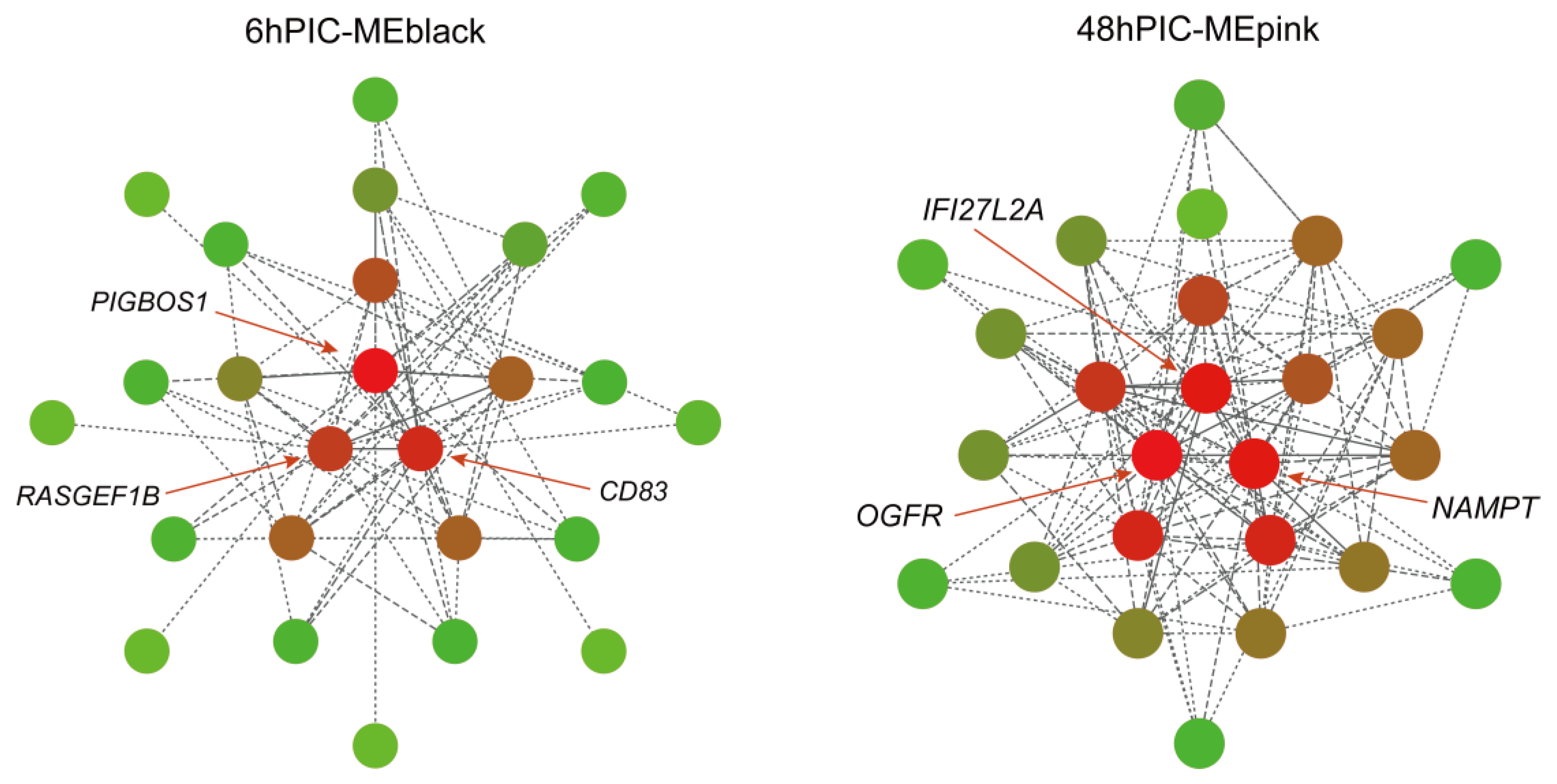
3.3. Key Hub Genes Related to Poly (I:C) Injection Stress Identified by WGCNA
4. Materials and Methods
4.1. Ethics Statement
4.2. Experimental Fishes and Injection Program
4.3. RNA Extraction and Illumina Sequencing
4.4. Data Filtering and Mapping
4.5. Differential Expression Genes (DEGs) Analysis and Functional Enrichment
4.6. Weighted Gene Co-Expression Network Analysis (WGCNA)
4.7. Quantitative Real-Time PCR Validation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Takita, T. Studies on the early life history of Nibea albiflora (Richardson) in Ariake sound. Bull. Fac. Fish. Nagasaki Univ. 1974, 38, 1–55. [Google Scholar]
- Liu, G.; Dong, L.; Gu, L.; Han, Z.; Zhang, W.; Fang, M.; Wang, Z. Evaluation of genomic selection for seven economic traits in yellow drum (Nibea albiflora). Mar. Biotechnol. 2019, 21, 806–812. [Google Scholar] [CrossRef]
- Ma, R.; Yu, Y.; Liu, X.; Lei, Y.; Zhou, S.; Xie, X.; Jin, S.; Qian, D.; Yin, F. Transcriptomic analysis of Nibea albiflora skin in response to infection by Cryptocaryon irritans. Fish Shellfish Immunol. 2020, 98, 819–831. [Google Scholar] [CrossRef]
- Xie, X.; Kong, J.; Huang, J.; Zhou, L.; Jiang, Y.; Miao, R.; Yin, F. Integration of metabolomic and transcriptomic analyses to characterize the influence of the gill metabolism of Nibea albiflora on the response to Cryptocaryon irritans infection. Vet. Parasitol. 2021, 298, 109533. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Y.; Maha, I.F.; Kong, J.; Xie, X.; Yin, F. Label-free quantitative proteomics analysis of skin of yellow drum (Nibea albiflora) reveals immune mechanism against Cryptocaryon irritans. Fish Shellfish Immunol. 2020, 101, 284–290. [Google Scholar] [CrossRef]
- Zhang, J.; Cui, Z.; Hu, G.; Jiang, X.; Wang, J.; Qiao, G.; Li, Q. Transcriptome analysis provides insights into the antiviral response in the spleen of gibel carp (Carassius auratus gibelio) after poly I: C treatment. Fish Shellfish Immunol. 2020, 102, 13–19. [Google Scholar] [CrossRef]
- Gao, W.; Zheng, W.; Sun, Y.; Xu, T. MicroRNA-489 negatively modulates RIG-I signaling pathway via targeting TRAF6 in miiuy croaker after poly(I:C) stimulation. Fish Shellfish Immun. 2021, 113, 61–68. [Google Scholar] [CrossRef]
- Zhou, Z.X.; Zhang, B.C.; Sun, L. Poly(I:C) induces antiviral immune responses in Japanese flounder (Paralichthys olivaceus) that require TLR3 and MDA5 and is negatively regulated by Myd88. PLoS ONE 2014, 9, e112918. [Google Scholar] [CrossRef]
- Liu, S.; Jiang, C.; Duan, C.; Hu, L.; Zhang, S. Expression of virus-responsive genes and their response to challenge with poly(I:C) at different stages of the annual fish Nothobranchius guentheri: Implications for an asymmetric decrease in immunity. Fish Shellfish Immunol. 2015, 46, 493–500. [Google Scholar] [CrossRef]
- Yu, Z.; Zhao, Y.Y.; Jiang, N.; Zhang, A.Z.; Li, M.Y. Bioflocs attenuates lipopolysaccharide-induced inflammation, immunosuppression and oxidative stress in Channa argus. Fish Shellfish Immunol. 2021, 114, 218–228. [Google Scholar] [CrossRef]
- Mottaz, H.; Schönenberger, R.; Fischer, S.; Eggen, R.; Schirmer, K.; Groh, K.J. Dose-dependent effects of morphine on lipopolysaccharide (LPS)-induced inflammation, and involvement of multixenobiotic resistance (MXR) transporters in LPS efflux in teleost fish. Environ. Pollut. 2017, 221, 105–115. [Google Scholar] [CrossRef]
- Nishizawa, T.; Takami, I.; Yoshimizu, M.; Oh, M.J. Required dose of fish nervous necrosis virus (NNV) for poly(I:C) immunization of sevenband grouper Epinephelus septemfasciatus. Aquaculture 2011, 311, 100–104. [Google Scholar] [CrossRef]
- Saravia, J.; Paschke, K.; Pontigo, J.P.; Nualart, D.; Navarro, J.M.; Vargas-Chacoff, L. Effects of temperature on the innate immune response on Antarctic and sub-Antarctic fish Harpagifer antarcticus and Harpagifer bispinis challenged with two immunostimulants, LPS and Poly I:C: In vivo and in vitro approach. Fish Shellfish Immunol. 2022, 130, 391–408. [Google Scholar] [CrossRef]
- Nsrelden, R.M.; Horiuchi, H.; Furusawa, S. Expression of ayu antimicrobial peptide genes after LPS stimulation. J. Vet. Med. Sci. 2017, 79, 1072–1080. [Google Scholar] [CrossRef]
- Qiu, H.T.; Fernandes, J.; Hong, W.S.; Wu, H.X.; Zhang, Y.T.; Huang, S.; Liu, D.T.; Yu, H.; Wang, Q.; You, X.X.; et al. Paralogues from the expanded TLR11 gene family in mudskipper (Boleophthalmus pectinirostris) are under positive selection and respond differently to LPS/poly(I:C) challenge. Front Immunol. 2019, 10, 343. [Google Scholar] [CrossRef]
- Ciesielska, A.; Matyjek, M.; Kwiatkowska, K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell. Mol. Life Sci. 2021, 78, 1233–1261. [Google Scholar] [CrossRef]
- Jeong, Y.M.; Jin, T.E.; Choi, J.H.; Lee, M.S.; Kim, H.T.; Hwang, K.S.; Park, D.S.; Oh, H.W.; Choi, J.K.; Korzh, V.; et al. Induction of clusterin expression by neuronal cell death in Zebrafish. J. Genet. Genomics. 2014, 41, 583–589. [Google Scholar] [CrossRef]
- Lewis, J.C.; Jones, N.L.; Hermanns, M.I.; Röhrig, O.; Klein, C.L.; Kirkpatrick, C.J. Tissue factor expression during coculture of endothelial cells and monocytes. Exp. Mol. Pathol. 1995, 62, 207–218. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.N.; Tang, Y.Y.; Zhou, M.J.; Luo, S.; Li, Y.T.; Wang, G.; Zhang, D.Z.; Yang, H.; Tang, B.P.; He, W.F. Differentially expressed genes involved in immune pathways from yellowhead catfish (Tachysurus fulvidraco) after poly (I:C) challenge. Int. J. Biol. Macromol. 2021, 183, 340–345. [Google Scholar] [CrossRef]
- Liu, Y.; Xin, Z.Z.; Zhang, D.Z.; Wang, Z.F.; Zhu, X.Y.; Tang, B.P.; Jiang, S.H.; Zhang, H.B.; Zhou, C.L.; Chai, X.Y.; et al. Transcriptome analysis of yellow catfish (Pelteobagrus fulvidraco) liver challenged with polyriboinosinic polyribocytidylic acid (poly I:C). Fish Shellfish Immunol. 2017, 68, 395–403. [Google Scholar] [CrossRef]
- Lulijwa, R.; Alfaro, A.C.; Merien, F.; Burdass, M.; Meyer, J.; Venter, L.; Young, T. Metabolic and immune responses of Chinook salmon (Oncorhynchus tshawytscha) smolts to a short-term poly (I:C) challenge. J. Fish Biol. 2020, 96, 731–746. [Google Scholar] [CrossRef]
- Magnadottir, B. Innate immunity offish (overview). Fish Shellfish Immunol. 2006, 20, 137–151. [Google Scholar] [CrossRef]
- Neville, L.F.; Mathiak, G.; Bagasra, O. The immunobiology of interferon-gamma inducible protein 10 kD (IP-10): A novel, pleiotropic member of the C-X-C chemokine superfamily. Cytokine Growth Factor Rev. 1997, 8, 207e219. [Google Scholar] [CrossRef]
- Yoshie, O.; Imai, T.; Nomiyama, H. Chemokines in immunity. Adv. Immunol. 2001, 78, 57–110. [Google Scholar] [PubMed]
- Secombes, C.J.; Wang, T.; Bird, S. The interleukins of fish. Dev. Comp. Immunol. 2011, 35, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, J.; Zhang, R.; Liu, L.; Ma, G.; Zhu, H. Interleukin-6 in Siberian sturgeon (Acipenser baeri): Molecular characterization and immune functional activity. Fish Shellfish Immunol. 2020, 102, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.; Zou, J.; Savan, R.; Kono, T.; Sakai, M.; Woo, J.; Secombes, C. Characterisation and expression analysis of an interleukin 6 homologue in the Japanese pufferfish, Fugu rubripes. Dev. Comp. Immunol. 2005, 29, 775–789. [Google Scholar] [CrossRef]
- Iliev, D.B.; Castellana, B.; Mackenzie, S.; Planas, J.V.; Goetz, F.W. Cloning and expression analysis of an IL-6 homolog in rainbow trout (Oncorhynchus mykiss). Mol. Immunol. 2007, 44, 1803–1807. [Google Scholar] [CrossRef]
- Chen, H.H.; Lin, H.T.; Foung, Y.F.; Han-You Lin, J. The bioactivity of teleost IL-6: IL-6 protein in orange-spotted grouper (Epinephelus coioides) induces Th2 cell differentiation pathway and antibody production. Dev. Comp. Immunol. 2012, 38, 285–294. [Google Scholar] [CrossRef]
- Varela, M.; Dios, S.; Novoa, B.; Figueras, A. Characterisation, expression and ontogeny of interleukin-6 and its receptors in zebrafish (Danio rerio). Dev. Comp. Immunol. 2012, 37, 97–106. [Google Scholar] [CrossRef]
- Li, X.P.; Zhang, J. Tongue sole (Cynoglossus semilaevis) interleukin 10 receptors are involved in the immune response against bacterial infection. Dev. Comp. Immunol. 2021, 115, 103885. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Li, F.; Zhao, C.; Yan, L.; Fan, S.; Zheng, S.; Xu, H.; Qiu, L. Molecular characterization and functional analysis of TRAF6 in the spotted sea bass (Lateolabrax maculatus). Fish Shellfish Immunol. 2020, 105, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Li, X.; Sun, X.; Zhou, Z.; Cai, X.; Liu, X.; Wang, J.; Xiao, W. Zebrafish prmt2 attenuates antiviral innate immunity by targeting TRAF6. J. Immunol. 2021, 207, 2570–2580. [Google Scholar] [CrossRef]
- Du, X.; Li, Y.; Li, D.; Lian, F.; Yang, S.; Wu, J.; Liu, H.; Bu, G.; Meng, F.; Cao, X.; et al. Transcriptome profiling of spleen provides insights into the antiviral mechanism in Schizothorax prenanti after poly (I:C) challenge. Fish Shellfish Immunol. 2017, 62, 13–23. [Google Scholar] [CrossRef]
- Li, Q.; Wu, M.; Cui, K.; Zhu, S.; Mai, K.; Ai, Q. Characterization of antiviral immune response induced by poly(I:C) in macrophages of farmed large yellow croaker (Larimichthys crocea). Fish Shellfish Immunol. 2020, 104, 663–672. [Google Scholar] [CrossRef]
- Clark, T.C.; Boudinot, P.; Collet, B. Evolution of the IRF family in Salmonids. Genes 2021, 12, 238. [Google Scholar] [CrossRef] [PubMed]
- Langevin, C.; Boudinot, P.; Collet, B. IFN signaling in inflammation and viral infections: New insights from fish models. Viruses 2019, 11, 302. [Google Scholar] [CrossRef]
- Yan, X.; Zhao, X.; Huo, R.; Xu, T. IRF3 and IRF8 regulate NF-κB signaling by targeting MyD88 in teleost fish. Front. Immunol. 2020, 11, 606. [Google Scholar] [CrossRef]
- Liu, H.; Yang, M.; Tang, X.; Liu, J.; Zheng, L.; Xu, D.; Chi, C.; Lv, Z. Molecular insights of a novel fish Toll-like receptor 9 homologue in Nibea albiflora to reveal its function as PRRs. Fish Shellfish Immunol. 2021, 118, 321–332. [Google Scholar] [CrossRef]
- Chu, Q.; Martinez, T.F.; Novak, S.W.; Donaldson, C.J.; Tan, D.; Vaughan, J.M.; Chang, T.; Diedrich, J.K.; Andrade, L.; Kim, A.; et al. Regulation of the ER stress response by a mitochondrial microprotein. Nat. Commun. 2019, 10, 4883. [Google Scholar] [CrossRef]
- Doñate, C.; Roher, N.; Balasch, J.C.; Ribas, L.; Goetz, F.W.; Planas, J.V.; Tort, L.; MacKenzie, S. CD83 expression in sea bream macrophages is a marker for the LPS-induced inflammatory response. Fish Shellfish Immunol. 2007, 23, 877–885. [Google Scholar] [CrossRef]
- Buonocore, F.; Randelli, E.; Tranfa, P.; Scapigliati, G.A. CD83-like molecule in sea bass (Dicentrarchus labrax): Molecular characterization and modulation by viral and bacterial infection. Fish Shellfish Immunol. 2012, 32, 1179–1184. [Google Scholar] [CrossRef]
- Andrade, W.A.; Silva, A.M.; Alves, V.S.; Salgado, A.P.; Melo, M.B.; Andrade, H.M.; Dall’Orto, F.V.; Garcia, S.A.; Silveira, T.N.; Gazzinelli, R.T. Early endosome localization and activity of RasGEF1b, a toll-like receptor-inducible Ras guanine-nucleotide exchange factor. Genes Immun. 2010, 11, 447–457. [Google Scholar] [CrossRef]
- Epting, D.; Vorwerk, S.; Hageman, A.; Meyer, D. Expression of rasgef1b in zebrafish. Gene Expr. Patterns. 2007, 7, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Papac-Milicevic, N.; Breuss, J.M.; Zaujec, J.; Ryban, L.; Plyushch, T.; Wagner, G.A.; Fenzl, S.; Dremsek, P.; Cabaravdic, M.; Steiner, M.; et al. The interferon stimulated gene 12 inactivates vasculoprotective functions of NR4A nuclear receptors. Circ. Res. 2012, 110, e50–e63. [Google Scholar] [CrossRef] [PubMed]
- Matejusová, I.; Felix, B.; Sorsa-Leslie, T.; Gilbey, J.; Noble, L.R.; Jones, C.S.; Cunningham, C.O. Gene expression profiles of some immune relevant genes from skin of susceptible and responding Atlantic salmon (Salmo salar L.) infected with Gyrodactylus salaris (Monogenea) revealed by suppressive subtractive hybridization. Int. J. Parasitol. 2006, 36, 1175–1183. [Google Scholar] [CrossRef]
- Tang, Y.; Liu, X.; Feng, C.; Zhou, Z.; Liu, S. Nicotinamide phosphoribosyl transferase (Nampt) of hybrid crucian carp protects intestinal barrier and enhances host immune defense against bacterial infection. Dev. Comp. Immunoly 2022, 128, 104314. [Google Scholar] [CrossRef]
- Xiang, L.X.; Peng, B.; Dong, W.R.; Yang, Z.F.; Shao, J.Z. Lipopolysaccharide induces apoptosis in Carassius auratus lymphocytes, a possible role in pathogenesis of bacterial infection in fish. Dev. Comp. Immunol. 2008, 32, 992–1001. [Google Scholar] [CrossRef]
- Nya, E.J.; Austin, B. Use of bacterial lipopolysaccharide (LPS) as an immunostimulant for the control of Aeromonas hydrophila infections in rainbow trout Oncorhynchus mykiss (Walbaum). J. Appl. Microbiol. 2010, 108, 686–694. [Google Scholar] [CrossRef]
- Hou, C.Y.; Lin, J.H.; Lin, S.J.; Kuo, W.C.; Lin, H.T. Down-regulation of CD53 expression in Epinephelus coioides under LPS, poly (I:C), and cytokine stimulation. Fish Shellfish Immunol. 2016, 51, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- El-Gebali, S.; Mistry, J.; Bateman, A.; Eddy, S.R.; Luciani, A.; Potter, S.C.; Qureshi, M.; Richardson, L.J.; Salazar, G.A.; Smart, A.; et al. The Pfam protein families database in 2019. Nucleic Acids Res. 2019, 47, D427–D432. [Google Scholar] [CrossRef]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; López, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | Nr Annotation | Primer (5′ to 3′) |
|---|---|---|---|
| Nib0172480.1 | TLR22 | Toll-like receptor 22 | F: TGAAGTGAAGGAGGACAATAC R: TGTAACTGATTCGGCAAGAT |
| Nib0092120.1 | TLR14 | Toll-like receptor 14 | F: CAGGTGTTGGATGTTAGCA R: CTGATTGTGGTGAGCGATA |
| Nib0048970.1 | CLU | Clusterin | F: TCAACACCTCCTCAATCCT R: TTCTCTTCTCCATCTGACTTC |
| Nib0133830.1 | WDFY1 | WD repeat and FYVE domain-containing protein 1 | F: TGTTGCTGTATGCTGGAAG R: GTGTTGAAGACTTGACTGATG |
| Nib0135300.1 | CD36 | Coagulation factor IIIb | F: AGAACCTCTGGACTCTGATA R: ACGGCACTGATGTTACTC |
| Nib0201110.1 | GPCR68 | G-protein coupled receptor 68 | F: GCAACTTCATCACAGGAATA R: CAACAGCAGCACAGAATC |
| Nib0009400.1 | NR4A1 | Nuclear receptor subfamily 4 group A member 1 | F: AAGTGTTGGCAATCTGGAT R: GAGGCTGTGAGTAAGTTGT |
| Nib0056150.1 | NR4A3 | Nuclear receptor subfamily 4 group A member 3 | F: CGACTACAGCCAGTTCAG R: GCAGAATCTATGAGCAGGTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Zhang, Y.; Gao, T.; Song, N. Spleen Transcriptome Profiling Reveals Divergent Immune Responses to LPS and Poly (I:C) Challenge in the Yellow Drum (Nibea albiflora). Int. J. Mol. Sci. 2023, 24, 7735. https://doi.org/10.3390/ijms24097735
Zhao X, Zhang Y, Gao T, Song N. Spleen Transcriptome Profiling Reveals Divergent Immune Responses to LPS and Poly (I:C) Challenge in the Yellow Drum (Nibea albiflora). International Journal of Molecular Sciences. 2023; 24(9):7735. https://doi.org/10.3390/ijms24097735
Chicago/Turabian StyleZhao, Xiang, Yuan Zhang, Tianxiang Gao, and Na Song. 2023. "Spleen Transcriptome Profiling Reveals Divergent Immune Responses to LPS and Poly (I:C) Challenge in the Yellow Drum (Nibea albiflora)" International Journal of Molecular Sciences 24, no. 9: 7735. https://doi.org/10.3390/ijms24097735