
Identification and Functional Analysis of Tartrate-Resistant Acid Phosphatase Type 5b (TRAP5b) in Oreochromis niloticus
 ,
,
Abstract
:1. Introduction
2. Results
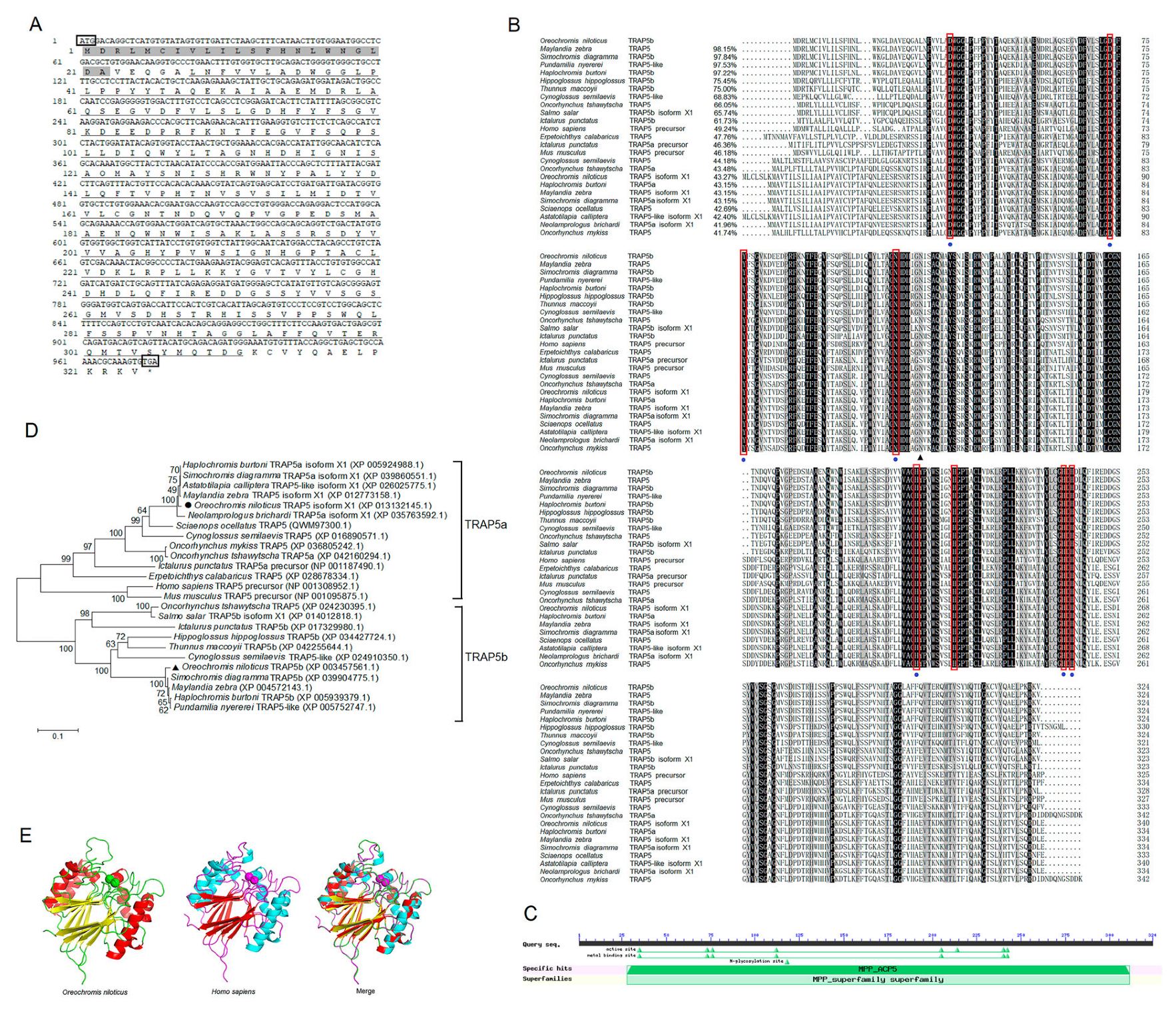
2.1. Molecular Characterization of OnTRAP5b
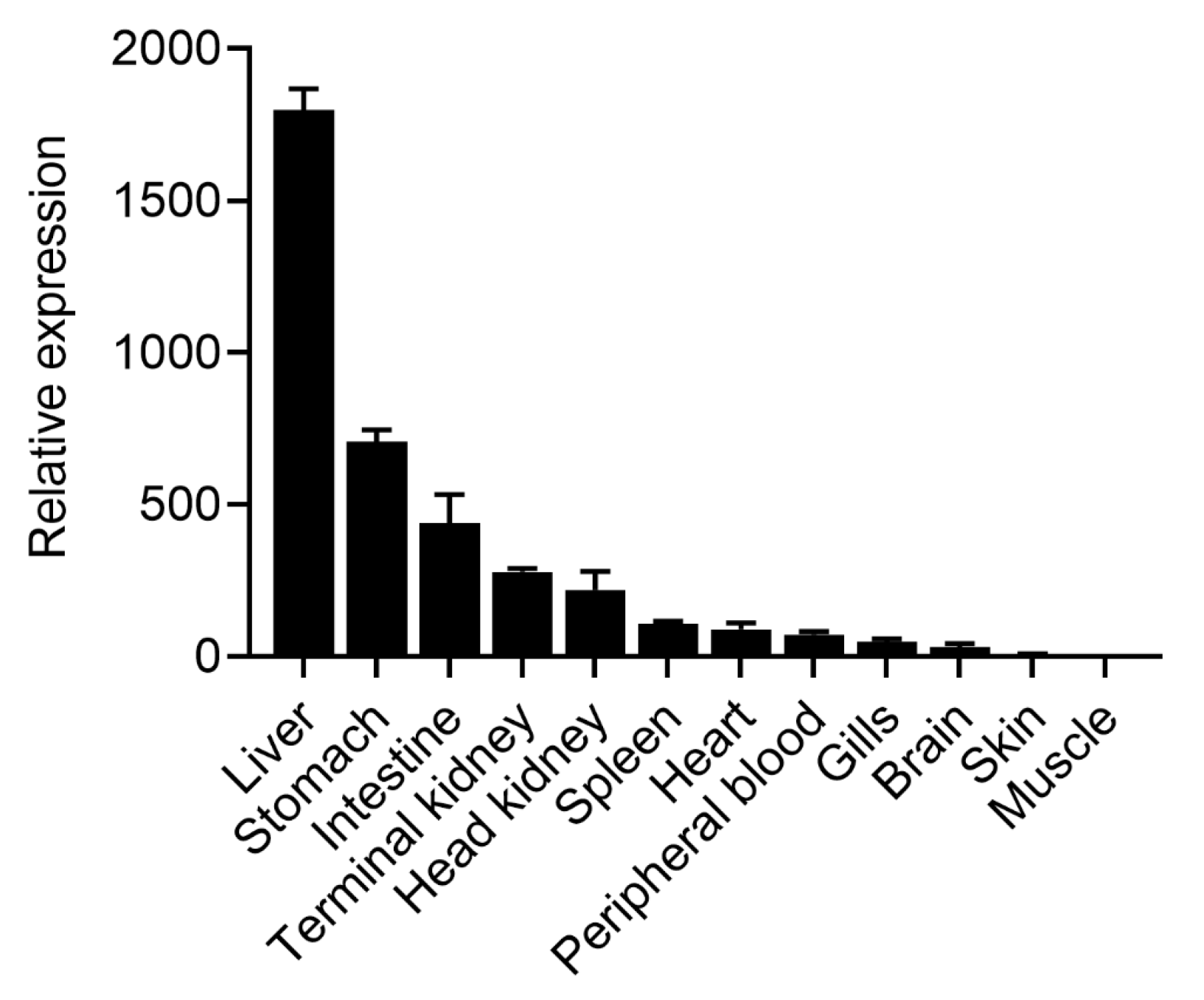
2.2. Tissue Distribution Analysis of OnTRAP5b
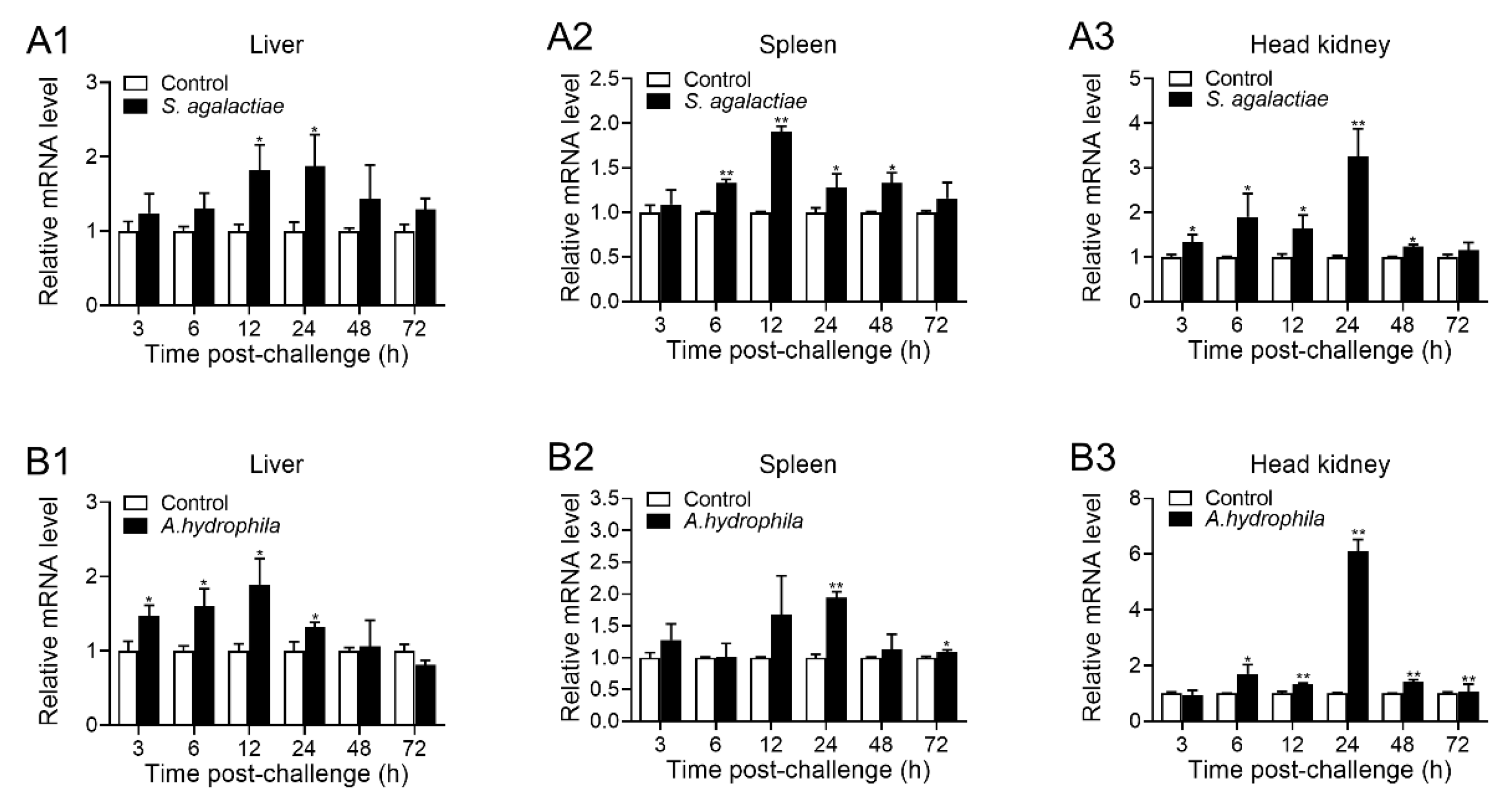
2.3. Expression Pattern of OnTRAP5b upon S. agalactiae and A. hydrophila Challenged In Vivo
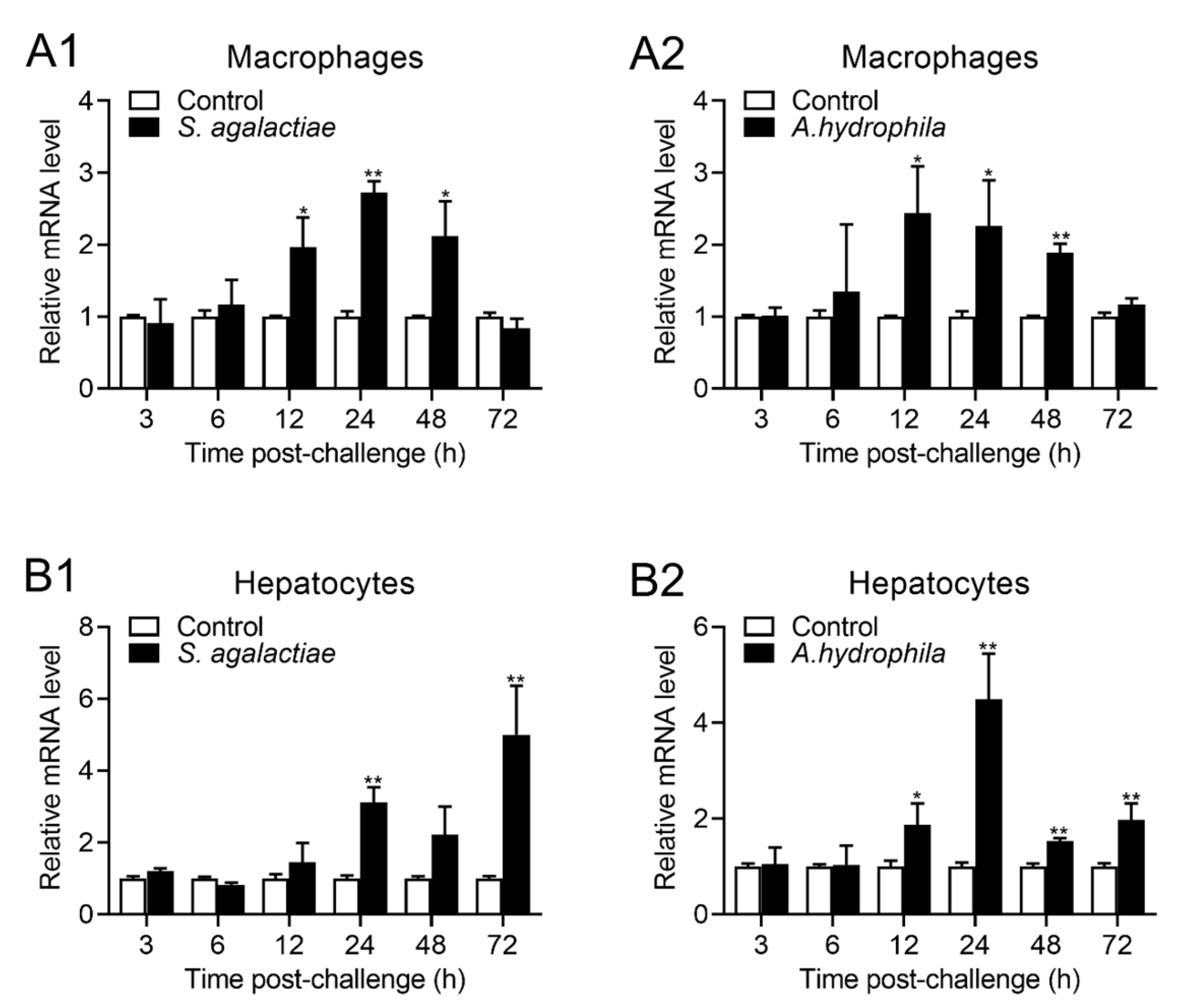
2.4. Expression of OnTRAP5b after Stimulation In Vitro
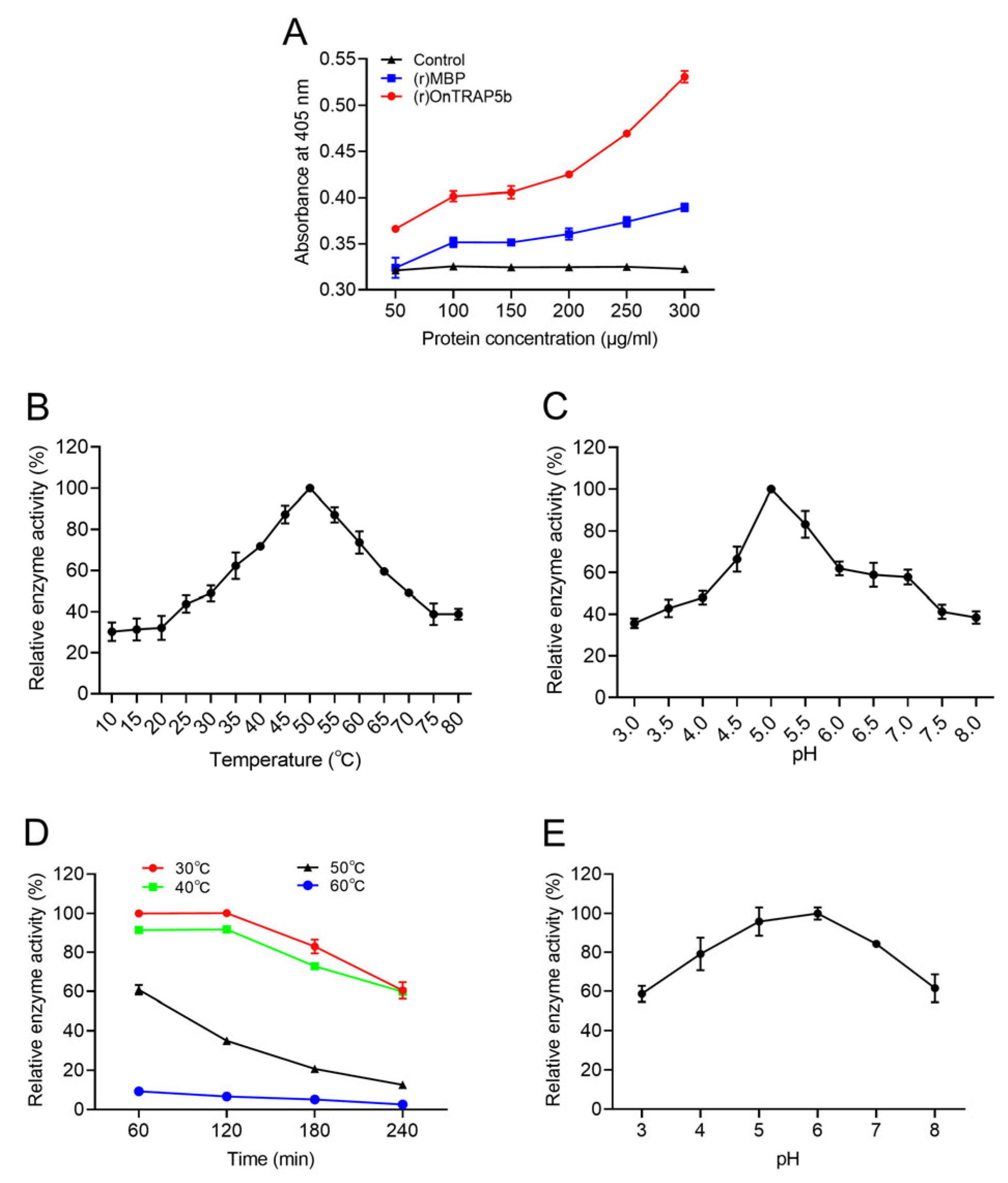
2.5. Purification and Phosphatase Activity of Recombinant (r)OnTRAP5b
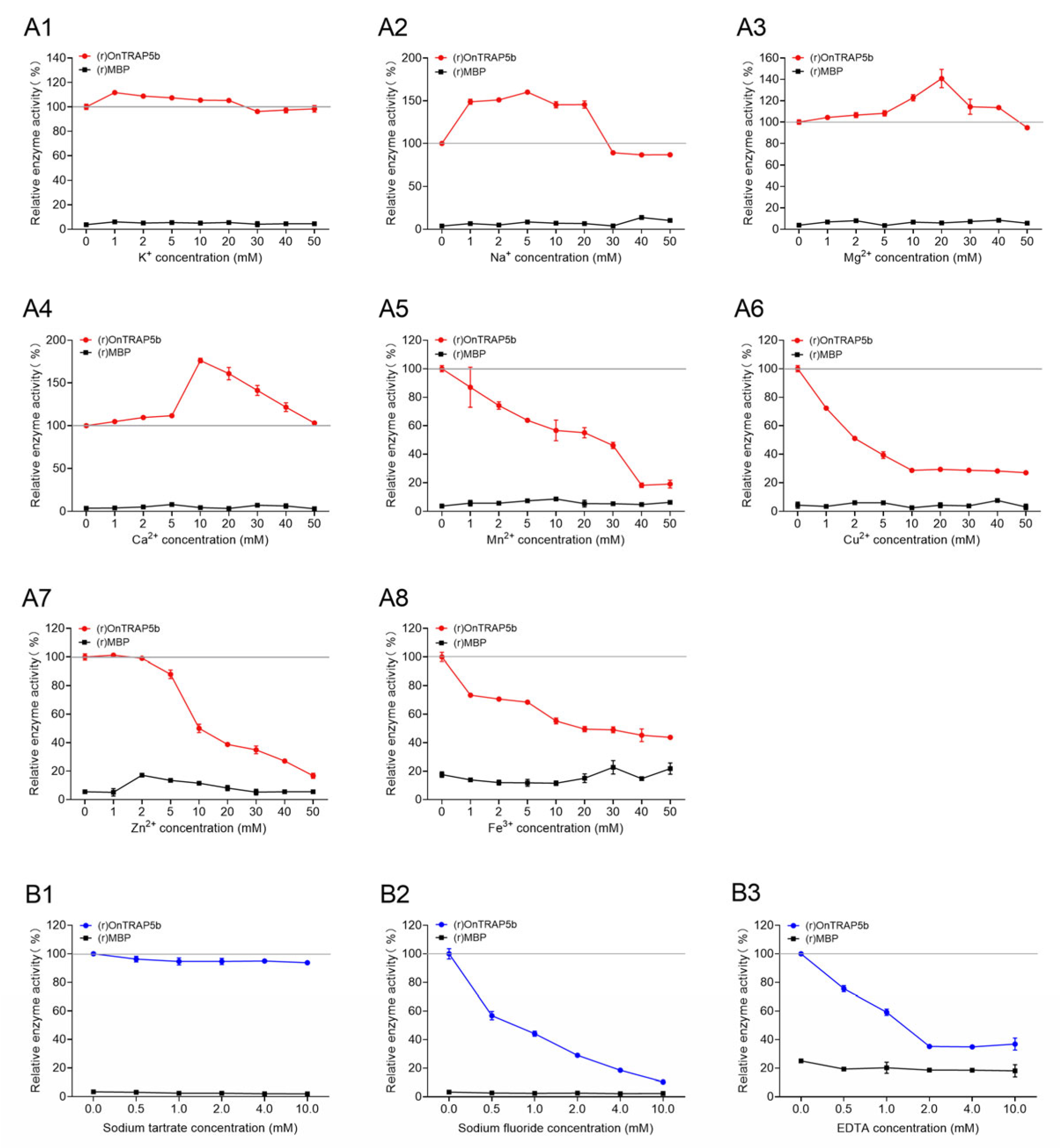
2.6. Effect of Metal Ions and Inhibitors on (r)OnTRAP5b Phosphatase Activity
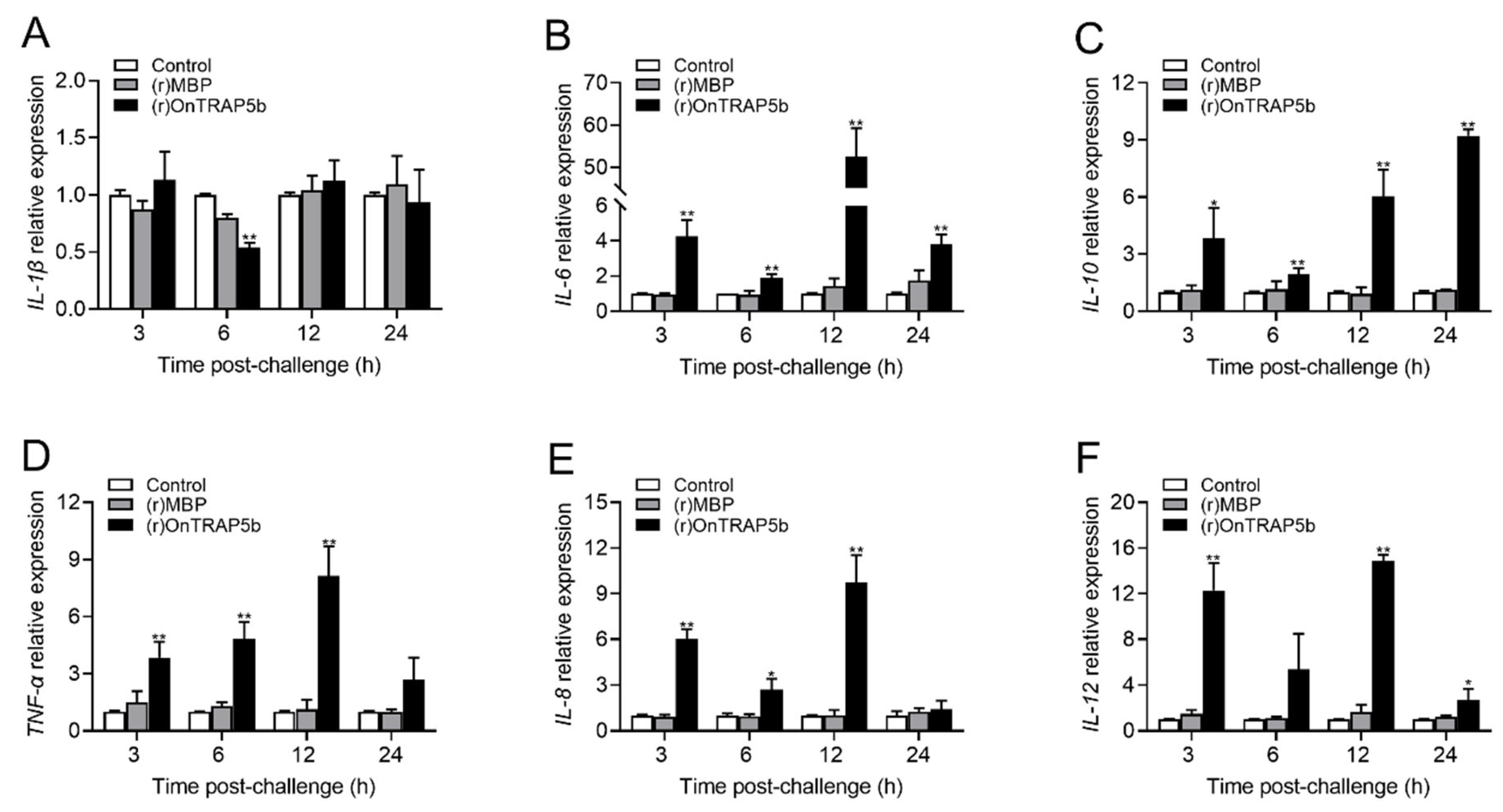
2.7. Effects of (r)OnTRAP5b on mRNA Expression of Inflammatory-Related Cytokines of Tilapia Head Kidney Macrophages
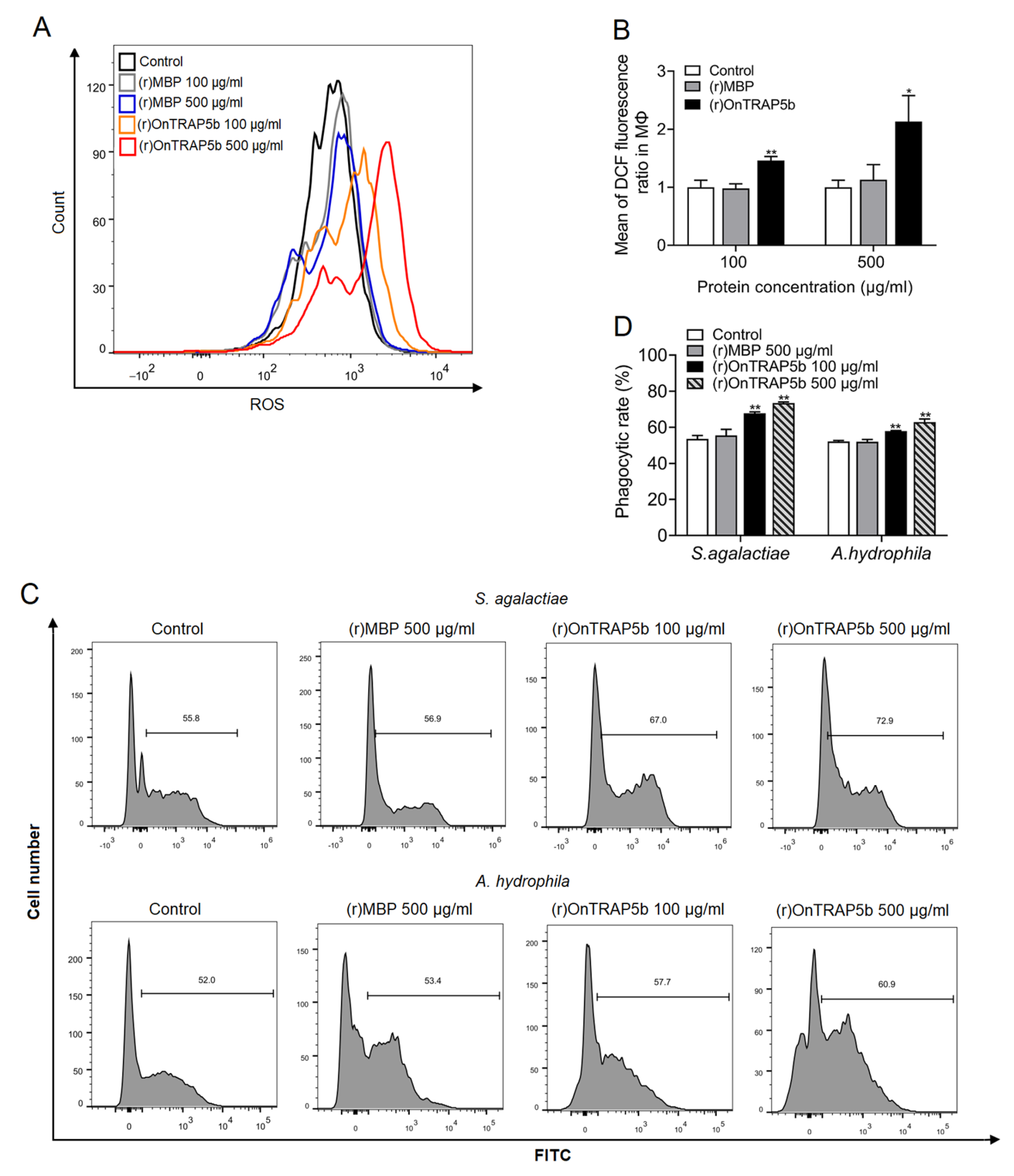
2.8. Effects of (r)OnTRAP5b on the Production of ROS and Phagocytic Activity in MΦ
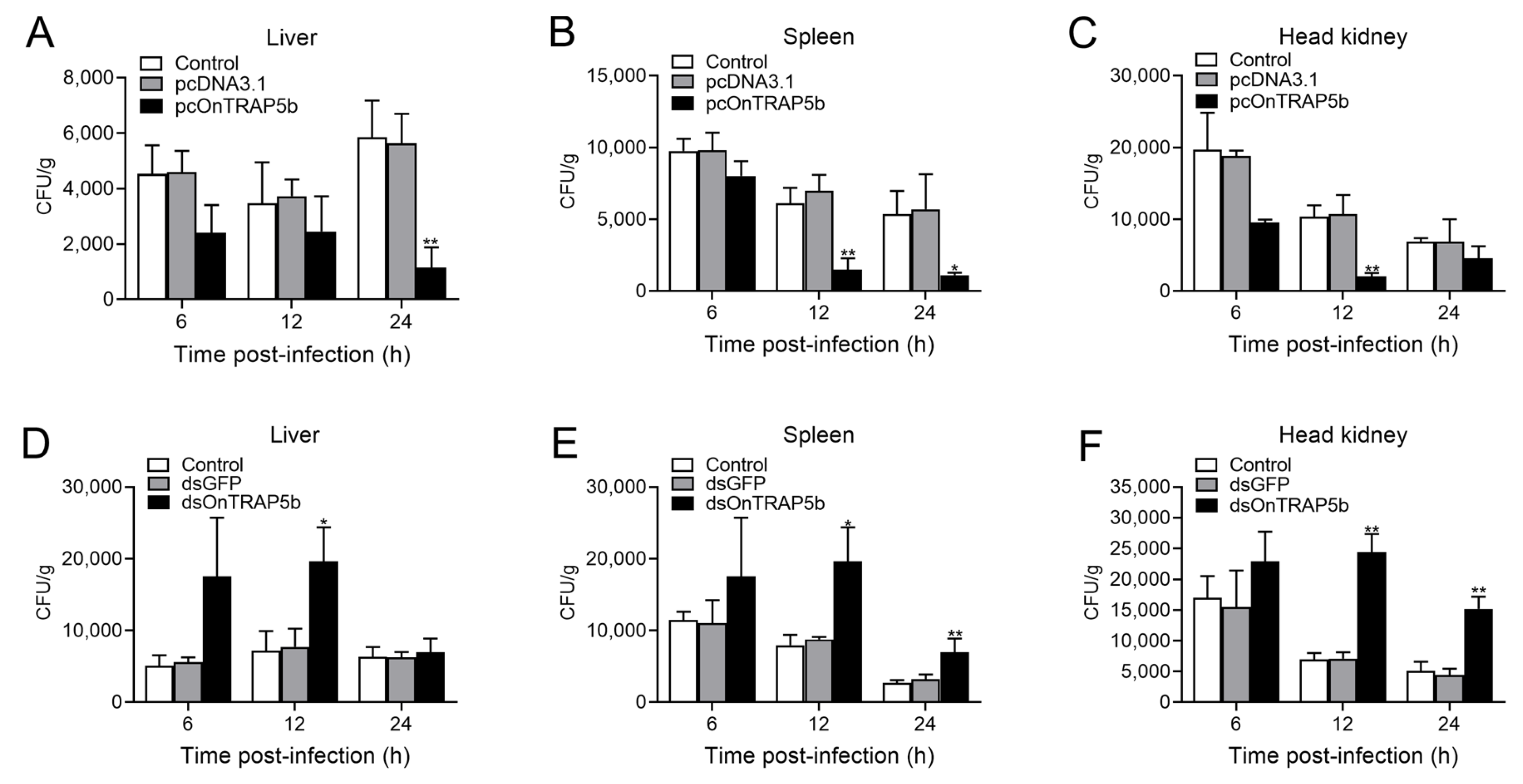
2.9. Effect of OnTRAP5b Overexpression and Knockdown on Bacteria Load In Vivo
3. Discussion
4. Materials and Methods
4.1. Fish
4.2. Total RNA Isolation, cDNA Synthesis, and Real-Time Quantitative PCR
4.3. Cloning and Sequence Analyses of OnTRAP5b
4.4. OnTRAP5b Expression in Different Tissues under Normal Physiological Conditions
4.5. Expression Pattern of OnTRAP5b following S. agalactiae and A. hydrophila Stimulation In Vivo
4.6. Isolation of Head Kidney MΦ and Hepatocytes
4.7. Plasmid Construction
4.8. Expression and Purification of Recombinant Protein
4.9. Phosphatase Activity of Recombinant Protein (r)OnTRAP5b
4.10. Effect of Different Metal Ions, Phosphatase Specific Inhibitors or EDTA on Phosphatase Activity of (r)OnTRAP5b
4.11. Effect of (r)OnTRAP5b on Immune-Related Genes
4.12. Measurement of Reactive Oxygen Species and Phagocytosis in MΦ
4.13. Effects of Overexpression and Knockdown of OnTRAP5b on S. agalactiae Infection
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| BHI | Brain heart infusion |
| CFU | Colony-Forming Units |
| DCFH-DA | 2,7-dichlorofluorescin diacetate |
| DCs | Dendritic cells |
| EDTA | Ethylene Diamine Tetraacetic Acid |
| FBS | Fetal bovine serum |
| FITC | Fluorescein isothiocyanate |
| h p.i | Hour post infection |
| IL-1β | Interleukin IL-1β |
| IL-6 | Interleukin IL-6 |
| IL-8 | Interleukin IL-8 |
| IL-10 | Interleukin IL-10 |
| IL-12 | Interleukin IL-12 |
| IPTG | Isopropyl-β-D-thiogalactopyranoside |
| LB | Luria-Bertani |
| LPS | Lipopolysaccharide |
| MPP | Metallophosphatase |
| MΦ | Macrophages |
| NCBI | National Center for Biotechnology Information |
| O.D. | Optical density |
| OnTRAP5b | Oreochromis niloticus TRAP5b |
| ORF | Open reading frame |
| P/S | Penicillin G and Streptomycin |
| PBS | Phosphate buffer saline |
| PCR | Polymerase Chain Reaction |
| pNPP | p-nitrophenyl phosphate |
| qRT-PCR | Quantitative real-time PCR |
| RNAi | RNA interference |
| (r)OnTRAP5b | Recombinant protein OnTRAP5b |
| ROS | Reactive oxygen species |
| RPMI-1640 | Roswell Park Memorial Institute 1640 medium |
| SD | Standard deviation |
| SDS-PAGE | SDS-polyacrylamide gel electrophoresis |
| siRNA | Synthesize small interfering RNA |
| TNF-α | Tumor necrosis factor-α |
| TRAP5 | Tartrate-resistant acid phosphatase type 5 |
References
- Jemtland, R.; Lee, K.; Segre, G.V. Heterogeneity among Cells That Express Osteoclast-Associated Genes in Developing Bone. Endocrinology 1998, 139, 340–349. [Google Scholar] [CrossRef]
- Kaida, R.; Hayashi, T.; Kaneko, T.S. Purple Acid Phosphatase in the Walls of Tobacco Cells. Phytochemistry 2008, 69, 2546–2551. [Google Scholar] [CrossRef]
- Veljanovski, V.; Vanderbeld, B.; Knowles, V.L.; Snedden, W.A.; Plaxton, W.C. Biochemical and Molecular Characterization of AtPAP26, a Vacuolar Purple Acid Phosphatase up-Regulated in Phosphate-Deprived Arabidopsis Suspension Cells and Seedlings. Plant Physiol. 2006, 142, 1282–1293. [Google Scholar] [CrossRef] [Green Version]
- Siddiqua, A.; Rehmat, M.; Saeed, A.; Amin, S.; Naz, R.; Sherazi, M.; Khan, G.M.; Saeed, A. Acid Phosphatases from the Liver of Labeo Rohita: Purification and Characterization. Biol. Pharm. Bull. 2008, 31, 802–808. [Google Scholar] [CrossRef] [Green Version]
- Vasileva-Tonkova, E.S.; Galabova, D.N.; Balasheva, M.A.; Sotirova, A.V. Purification and Partial Characterization of Acid Phosphatase from Candida Lipolytica. J. Gen. Microbiol. 1993, 139, 479–483. [Google Scholar] [CrossRef] [Green Version]
- Passariello, C.; Forleo, C.; Micheli, V.; Schippa, S.; Leone, R.; Mangani, S.; Thaller, M.C.; Rossolini, G.M. Biochemical Characterization of the Class B Acid Phosphatase (AphA) of Escherichia Coli MG1655. Biochim. Biophys. Acta 2006, 1764, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Yeung, S.-L.; Cheng, C.; Lui, T.K.O.; Tsang, J.S.H.; Chan, W.-T.; Lim, B.L. Purple Acid Phosphatase-like Sequences in Prokaryotic Genomes and the Characterization of an Atypical Purple Alkaline Phosphatase from Burkholderia Cenocepacia J2315. Gene 2009, 440, 1–8. [Google Scholar] [CrossRef]
- Minkin, C. Bone Acid Phosphatase: Tartrate-Resistant Acid Phosphatase as a Marker of Osteoclast Function. Calcif. Tissue Int. 1982, 34, 285–290. [Google Scholar] [CrossRef]
- Hayman, A.R.; Bune, A.J.; Bradley, J.R.; Rashbass, J.; Cox, T.M. Osteoclastic Tartrate-Resistant Acid Phosphatase (Acp 5): Its Localization to Dendritic Cells and Diverse Murine Tissues. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2000, 48, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Hayman, A.R.; Macary, P.; Lehner, P.J.; Cox, T.M. Tartrate-Resistant Acid Phosphatase (Acp 5): Identification in Diverse Human Tissues and Dendritic Cells. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2001, 49, 675–684. [Google Scholar] [CrossRef] [Green Version]
- Angel, N.Z.; Walsh, N.; Forwood, M.R.; Ostrowski, M.C.; Cassady, A.I.; Hume, D.A. Transgenic Mice Overexpressing Tartrate-Resistant Acid Phosphatase Exhibit an Increased Rate of Bone Turnover. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2000, 15, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Sträter, N.; Jasper, B.; Scholte, M.; Krebs, B.; Duff, A.P.; Langley, D.B.; Han, R.; Averill, B.A.; Freeman, H.C.; Guss, J.M. Crystal Structures of Recombinant Human Purple Acid Phosphatase with and without an Inhibitory Conformation of the Repression Loop. J. Mol. Biol. 2005, 351, 233–246. [Google Scholar] [CrossRef]
- Davis, J.C.; Lin, S.S.; Averill, B.A. Kinetics and Optical Spectroscopic Studies on the Purple Acid Phosphatase from Beef Spleen. Biochemistry 1981, 20, 4062–4067. [Google Scholar] [CrossRef] [PubMed]
- Ketcham, C.M.; Baumbach, G.A.; Bazer, F.W.; Roberts, R.M. The Type 5, Acid Phosphatase from Spleen of Humans with Hairy Cell Leukemia. Purification, Properties, Immunological Characterization, and Comparison with Porcine Uteroferrin. J. Biol. Chem. 1985, 260, 5768–5776. [Google Scholar] [CrossRef]
- Robinson, D.B.; Glew, R.H. A Tartrate-Resistant Acid Phosphatase from Gaucher Spleen. Purification and Properties. J. Biol. Chem. 1980, 255, 5864–5870. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.H.; Nuttleman, P.R.; Ketcham, C.M.; Roberts, R.M. Purification and Characterization of Human Bone Tartrate-Resistant Acid Phosphatase. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1989, 4, 47–55. [Google Scholar] [CrossRef]
- Efstratiadis, T.; Moss, D.W. Tartrate-Resistant Acid Phosphatase of Human Lung: Apparent Identity with Osteoclastic Acid Phosphatase. Enzyme 1985, 33, 34–40. [Google Scholar] [CrossRef] [PubMed]
- Ketcham, C.M.; Roberts, R.M.; Simmen, R.C.; Nick, H.S. Molecular Cloning of the Type 5, Iron-Containing, Tartrate-Resistant Acid Phosphatase from Human Placenta. J. Biol. Chem. 1989, 264, 557–563. [Google Scholar] [CrossRef]
- Doi, K.; Antanaitis, B.C.; Aisen, P. The Binuclear Iron Centers of Uteroferrin and the Purple Acid Phosphatases. In Bioinorganic Chemistry; Springer: Berlin/Heidelberg, Germany, 1988; pp. 1–26. [Google Scholar]
- Schenk, G.; Guddat, L.W.; Ge, Y.; Carrington, L.E.; Hume, D.A.; Hamilton, S.; de Jersey, J. Identification of Mammalian-like Purple Acid Phosphatases in a Wide Range of Plants. Gene 2000, 250, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.C.; Averill, B.A. Evidence for a Spin-Coupled Binuclear Iron Unit at the Active Site of the Purple Acid Phosphatase from Beef Spleen. Proc. Natl. Acad. Sci. USA 1982, 79, 4623–4627. [Google Scholar] [CrossRef] [Green Version]
- Halleen, J.M.; Räisänen, S.R.; Alatalo, S.L.; Väänänen, H.K. Potential Function for the ROS-Generating Activity of TRACP. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2003, 18, 1908–1911. [Google Scholar] [CrossRef]
- Andersson, G.; Ek-Rylander, B.; Hollberg, K.; Ljusberg-Sjölander, J.; Lång, P.; Norgård, M.; Wang, Y.; Zhang, S.-J. TRACP as an Osteopontin Phosphatase. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2003, 18, 1912–1915. [Google Scholar] [CrossRef]
- Räisänen, S.R.; Alatalo, S.L.; Ylipahkala, H.; Halleen, J.M.; Cassady, A.I.; Hume, D.A.; Väänänen, H.K. Macrophages Overexpressing Tartrate-Resistant Acid Phosphatase Show Altered Profile of Free Radical Production and Enhanced Capacity of Bacterial Killing. Biochem. Biophys. Res. Commun. 2005, 331, 120–126. [Google Scholar] [CrossRef]
- Bune, A.J.; Hayman, A.R.; Evans, M.J.; Cox, T.M. Mice Lacking Tartrate-Resistant Acid Phosphatase (Acp 5) Have Disordered Macrophage Inflammatory Responses and Reduced Clearance of the Pathogen, Staphylococcus Aureus. Immunology 2001, 102, 103–113. [Google Scholar] [CrossRef]
- Suter, A.; Everts, V.; Boyde, A.; Jones, S.J.; Lüllmann-Rauch, R.; Hartmann, D.; Hayman, A.R.; Cox, T.M.; Evans, M.J.; Meister, T.; et al. Overlapping Functions of Lysosomal Acid Phosphatase (LAP) and Tartrate-Resistant Acid Phosphatase (Acp5) Revealed by Doubly Deficient Mice. Dev. Camb. Engl. 2001, 128, 4899–4910. [Google Scholar] [CrossRef]
- Lord, D.K.; Cross, N.C.; Bevilacqua, M.A.; Rider, S.H.; Gorman, P.A.; Groves, A.V.; Moss, D.W.; Sheer, D.; Cox, T.M. Type 5 Acid Phosphatase. Sequence, Expression and Chromosomal Localization of a Differentiation-Associated Protein of the Human Macrophage. Eur. J. Biochem. 1990, 189, 287–293. [Google Scholar] [CrossRef]
- Ek-Rylander, B.; Bill, P.; Norgård, M.; Nilsson, S.; Andersson, G. Cloning, Sequence, and Developmental Expression of a Type 5, Tartrate-Resistant, Acid Phosphatase of Rat Bone. J. Biol. Chem. 1991, 266, 24684–24689. [Google Scholar] [CrossRef]
- Ling, P.; Roberts, R.M. Uteroferrin and Intracellular Tartrate-Resistant Acid Phosphatases Are the Products of the Same Gene. J. Biol. Chem. 1993, 268, 6896–6902. [Google Scholar] [CrossRef]
- Wang, Y.; Norgård, M.; Andersson, G. N-Glycosylation Influences the Latency and Catalytic Properties of Mammalian Purple Acid Phosphatase. Arch. Biochem. Biophys. 2005, 435, 147–156. [Google Scholar] [CrossRef]
- Palacios, M.C.; Haros, M.; Rosell, C.M.; Sanz, Y. Characterization of an Acid Phosphatase from Lactobacillus Pentosus: Regulation and Biochemical Properties. J. Appl. Microbiol. 2005, 98, 229–237. [Google Scholar] [CrossRef]
- Jing, G.; Li, L.; Li, Y.; Xie, L.; Zhang, R. Purification and Partial Characterization of Two Acid Phosphatase Forms from Pearl Oyster (Pinctada Fucata). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 229–235. [Google Scholar] [CrossRef]
- Dos-Santos, A.L.A.; Dick, C.F.; Lopes, L.R.; Rocco-Machado, N.; Muzi-Filho, H.; Freitas-Mesquita, A.L.; Paes-Vieira, L.; Vieyra, A.; Meyer-Fernandes, J.R. Tartrate-Resistant Phosphatase Type 5 in Trypanosoma Cruzi Is Important for Resistance to Oxidative Stress Promoted by Hydrogen Peroxide. Exp. Parasitol. 2019, 205, 107748. [Google Scholar] [CrossRef] [PubMed]
- Tagad, C.K.; Sabharwal, S.G. Purification and Characterization of Acid Phosphatase from Macrotyloma Uiflorum Seeds. J. Food Sci. Technol. 2018, 55, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Lam, W.K.; Eastlund, D.T.; Li, C.Y.; Yam, L.T. Biochemical Properties of Tartrate-Resistant Acid Phosphatase in Serum of Adults and Children. Clin. Chem. 1978, 24, 1105–1108. [Google Scholar] [CrossRef] [PubMed]
- Tham, S.-J.; Chang, C.-D.; Huang, H.-J.; Lee, Y.-F.; Huang, T.-S.; Chang, C.-C. Biochemical Characterization of an Acid Phosphatase from Thermus Thermophilus. Biosci. Biotechnol. Biochem. 2010, 74, 727–735. [Google Scholar] [CrossRef]
- Lei, Y.; Cao, Z.; Hou, Y.; Du, X.; Zhang, H.; Du, H.; Zhou, Y.; Sun, Y. Molecular Characterization and Functional Analysis of Tartrate-Resistant Acid Phosphatase (ACP5) Gene in Red Drum (Sciaenops ocellatus). Fish Shellfish Immunol. 2023, 132, 108495. [Google Scholar] [CrossRef]
- Yang, L.H.; Xiao, B.; Wang, X.J.; Hou, Y.L.; Wang, J.; Sun, J. Characterization of acid phosphatase from Ctenopharyngodon idellus and effects of metal ions on the enzyme activity. J. Fish. Sci. China 2010, 17, 969–976. (In Chinese) [Google Scholar]
- Halleen, J.; Hentunen, T.A.; Hellman, J.; Väänänen, H.K. Tartrate-Resistant Acid Phosphatase from Human Bone: Purification and Development of an Immunoassay. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1996, 11, 1444–1452. [Google Scholar] [CrossRef]
- Janckila, A.J.; Latham, M.D.; Lam, K.W.; Chow, K.C.; Li, C.Y.; Yam, L.T. Heterogeneity of Hairy Cell Tartrate-Resistant Acid Phosphatase. Clin. Biochem. 1992, 25, 437–443. [Google Scholar] [CrossRef]
- Igarashi, Y.; Lee, M.Y.; Matsuzaki, S. Heparin Column Analysis of Serum Type 5 Tartrate-Resistant Acid Phosphatase Isoforms. J. Chromatogr. B. Biomed. Sci. App. 2001, 757, 269–276. [Google Scholar] [CrossRef]
- Oddie, G.W.; Schenk, G.; Angel, N.Z.; Walsh, N.; Guddat, L.W.; de Jersey, J.; Cassady, A.I.; Hamilton, S.E.; Hume, D.A. Structure, Function, and Regulation of Tartrate-Resistant Acid Phosphatase. Bone 2000, 27, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halleen, J.M.; Räisänen, S.; Salo, J.J.; Reddy, S.V.; Roodman, G.D.; Hentunen, T.A.; Lehenkari, P.P.; Kaija, H.; Vihko, P.; Väänänen, H.K. Intracellular Fragmentation of Bone Resorption Products by Reactive Oxygen Species Generated by Osteoclastic Tartrate-Resistant Acid Phosphatase. J. Biol. Chem. 1999, 274, 22907–22910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Mu, L.; Yin, X.; Wu, H.; Han, K.; Guo, Z.; Ye, J. MAp34 Regulates the Non-Specific Cell Immunity of Monocytes/Macrophages and Inhibits the Lectin Pathway of Complement Activation in a Teleost Fish. Front. Immunol. 2020, 11, 1706. [Google Scholar] [CrossRef] [PubMed]
- Munro, E.S.; Gahlawat, S.K.; Acosta, F.; Ellis, A.E. In Infectious Pancreatic Necrosis Virus Carrier Atlantic Salmon, Salmo Salar L., Post-Smolts, Almost All Kidney Macrophages Ex Vivo Contain a Low Level of Non-Replicating Virus. J. Fish Dis. 2006, 29, 43–48. [Google Scholar] [CrossRef]
- Yin, X.; Chen, N.; Mu, L.; Bai, H.; Wu, H.; Qi, W.; Huang, Y.; Jian, J.; Wang, A.; Ye, J. Identification and Characterization of Hepcidin from Nile Tilapia (Oreochromis niloticus) in Response to Bacterial Infection and Iron Overload. Aquaculture 2022, 546, 737317. [Google Scholar] [CrossRef]
- Fagerlund, K.M.; Ylipahkala, H.; Tiitinen, S.L.; Janckila, A.J.; Hamilton, S.; Mäentausta, O.; Väänänen, H.K.; Halleen, J.M. Effects of Proteolysis and Reduction on Phosphatase and ROS-Generating Activity of Human Tartrate-Resistant Acid Phosphatase. Arch. Biochem. Biophys. 2006, 449, 1–7. [Google Scholar] [CrossRef]
- Zhang, J.; Chen, L.; Sun, L. SmLMWPTP, a Teleost Low Molecular Weight Protein Tyrosine Phosphatase, Inhibits the Immune Response of Peripheral Blood Leukocytes in a Manner That Depends on the Conserved P-Loop. Dev. Comp. Immunol. 2013, 40, 103–111. [Google Scholar] [CrossRef]
- Zhou, M.; Tang, C.; Xu, J.; Yu, H. Genome Mining and Characterization of a New Mandelate Racemase. Sheng Wu Gong Cheng Xue Bao 2018, 34, 897–905. (In Chinese) [Google Scholar] [CrossRef]
- He, J.; Liu, X.; Li, Y.; Wang, Z.; Shi, H.; Kan, Y.; Yao, L.; Tang, C. High Level Expression of Nicotinamide Nucleoside Kinase from Saccharomyces cerevisiae and its Purification and Immobilization by One-Step Method. Front. Bioeng. Biotechnol. 2023, 11, 1134152. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′ → 3′) | Usage |
|---|---|---|
| OnTRAP5b-F1 | ATGGACAGGCTCATGTGTATAGTGT | Cloning of OnTRAP5b ORF sequence |
| OnTRAP5b-R1 | CACTTTGCGTTTTGGCAGC | |
| OnTRAP5b-F2 | CCGGATATCATGGTGGAACAAGGTGCCCT | Amplification of sequence of mature OnTRAP5b gene |
| OnTRAP5b-R2 | CCGGGATCCCACTTTGCGTTTTGGCAGC | |
| OnTRAP5b-F3 | CCGGGATCCGCCACCATGGACAGGCTCATGTGTATAGTGT | Overexpression of OnTRAP5b |
| OnTRAP5b-R3 | CCGGATATCCACTTTGCGTTTTGGCAGC | |
| dsOnTRAP5b-P1 | GGATCCTAATACGACTCACTATAGGTGTCTTCTCTCAGCCAT | Knockdown of OnTRAP5b |
| dsOnTRAP5b-P2 | AAATGGCTGAGAGAAGACACCTATAGTGAGTCGTATTAGGATCC | |
| dsOnTRAP5b-P3 | GGATCCTAATACGACTCACTATAATGGCTGAGAGAAGACACC | |
| dsOnTRAP5b-P4 | AAGGTGTCTTCTCTCAGCCATTATAGTGAGTCGTATTAGGATCC | |
| dsGFP-P1 | GGATCCTAATACGACTCACTATAGCCACAACGTCTATATCAT | Synthetic GFP siRNA |
| dsGFP-P2 | AAATGATATAGACGTTGTGGCTATAGTGAGTCGTATTAGGATCC | |
| dsGFP-P3 | GGATCCTAATACGACTCACTATAATGATATAGACGTTGTGGC | |
| dsGFP-P4 | AAGCCACAACGTCTATATCATTATAGTGAGTCGTATTAGGATCC | |
| qOnTRAP5b-F1 | ACTTCTATTTTAGCGGCGTCAA | qRT-PCR of OnTRAP5b gene |
| qOnTRAP5b-R1 | GCACCACCGTATCAATCATCA | |
| qIL-1β-F | CGTGCCAACAGTGAGAAAGCG | qRT-PCR analysis |
| qIL-1β-R | CAGGAGGGACGGAAGGGAT | |
| qIL-6-F | ACAGAGGAGGCGGAGATG | |
| qIL-6-R | GCAGTGCTTCGGGATAGAG | |
| qIL-10-F | TGGAGGGCTTCCCCGTCAG | |
| qIL-10-R | CTGTCGGCAGAACCGTGTCC | |
| qTNFα-F | GCTGAGGCTCCTGGACAAAA | |
| qTNFα-R | TCTGCCATTCCACTGAGGTCTT | |
| qIL-8-F | GATAAGCAACAGAATCATTGTCAGC | |
| qIL-8-R | CCTCGCAGTGGGAGTTGG | |
| qIL-12-F | GTGGTCTTGGTAAAGGCCGA | |
| qIL-12-R | AGTTTTGCCGGTCTCACGAT | |
| qβ-actin-F | CGAGAGGGAAATCGTGCGTGACA | |
| qβ-actin-R | AGGAAGGAAGGCTGGAAGAGGGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lei, Y.; Fu, S.; Yang, Y.; Chen, J.; Li, B.; Guo, Z.; Ye, J. Identification and Functional Analysis of Tartrate-Resistant Acid Phosphatase Type 5b (TRAP5b) in Oreochromis niloticus. Int. J. Mol. Sci. 2023, 24, 7179. https://doi.org/10.3390/ijms24087179
Lei Y, Fu S, Yang Y, Chen J, Li B, Guo Z, Ye J. Identification and Functional Analysis of Tartrate-Resistant Acid Phosphatase Type 5b (TRAP5b) in Oreochromis niloticus. International Journal of Molecular Sciences. 2023; 24(8):7179. https://doi.org/10.3390/ijms24087179
Chicago/Turabian StyleLei, Yang, Shengli Fu, Yanjian Yang, Jianlin Chen, Bingxi Li, Zheng Guo, and Jianmin Ye. 2023. "Identification and Functional Analysis of Tartrate-Resistant Acid Phosphatase Type 5b (TRAP5b) in Oreochromis niloticus" International Journal of Molecular Sciences 24, no. 8: 7179. https://doi.org/10.3390/ijms24087179