
Regulatory Effects of Three-Dimensional Cultured Lipopolysaccharide-Pretreated Periodontal Ligament Stem Cell-Derived Secretome on Macrophages
 ,
,
Abstract
:1. Introduction
2. Results
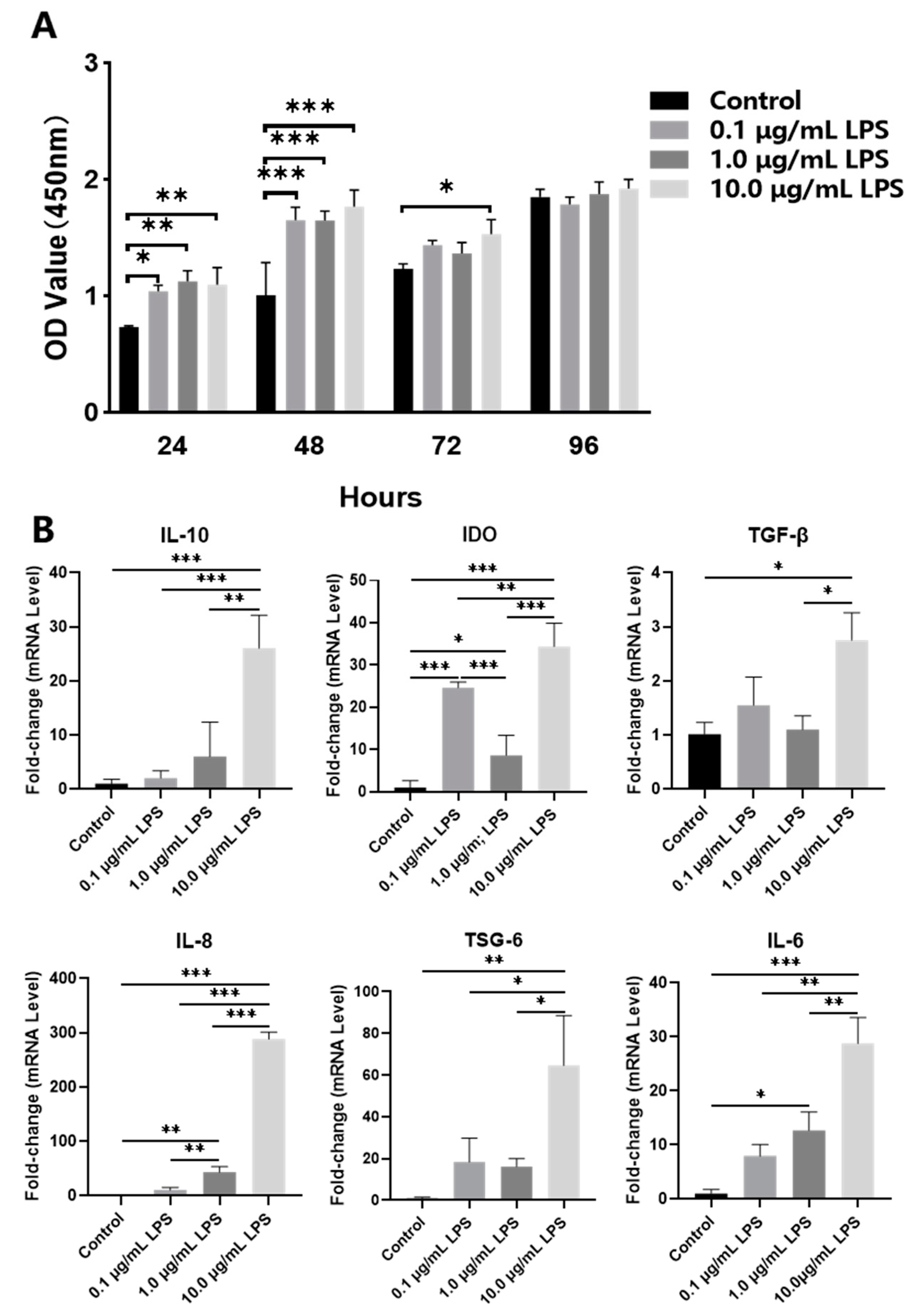
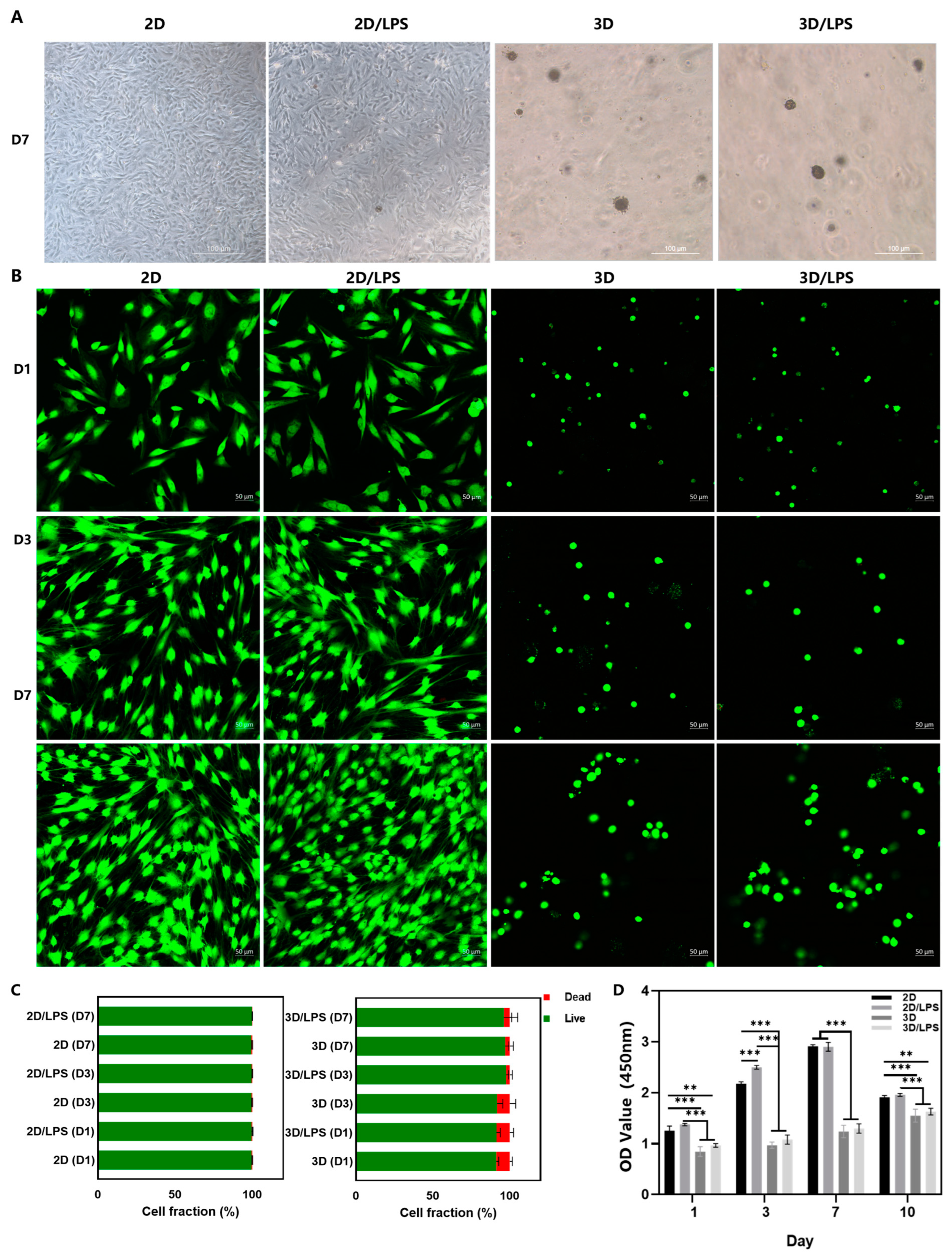
2.1. Effects of Lipopolysaccharide (LPS) Pretreatment on Periodontal Ligament Stem Cells (PDLSCs)
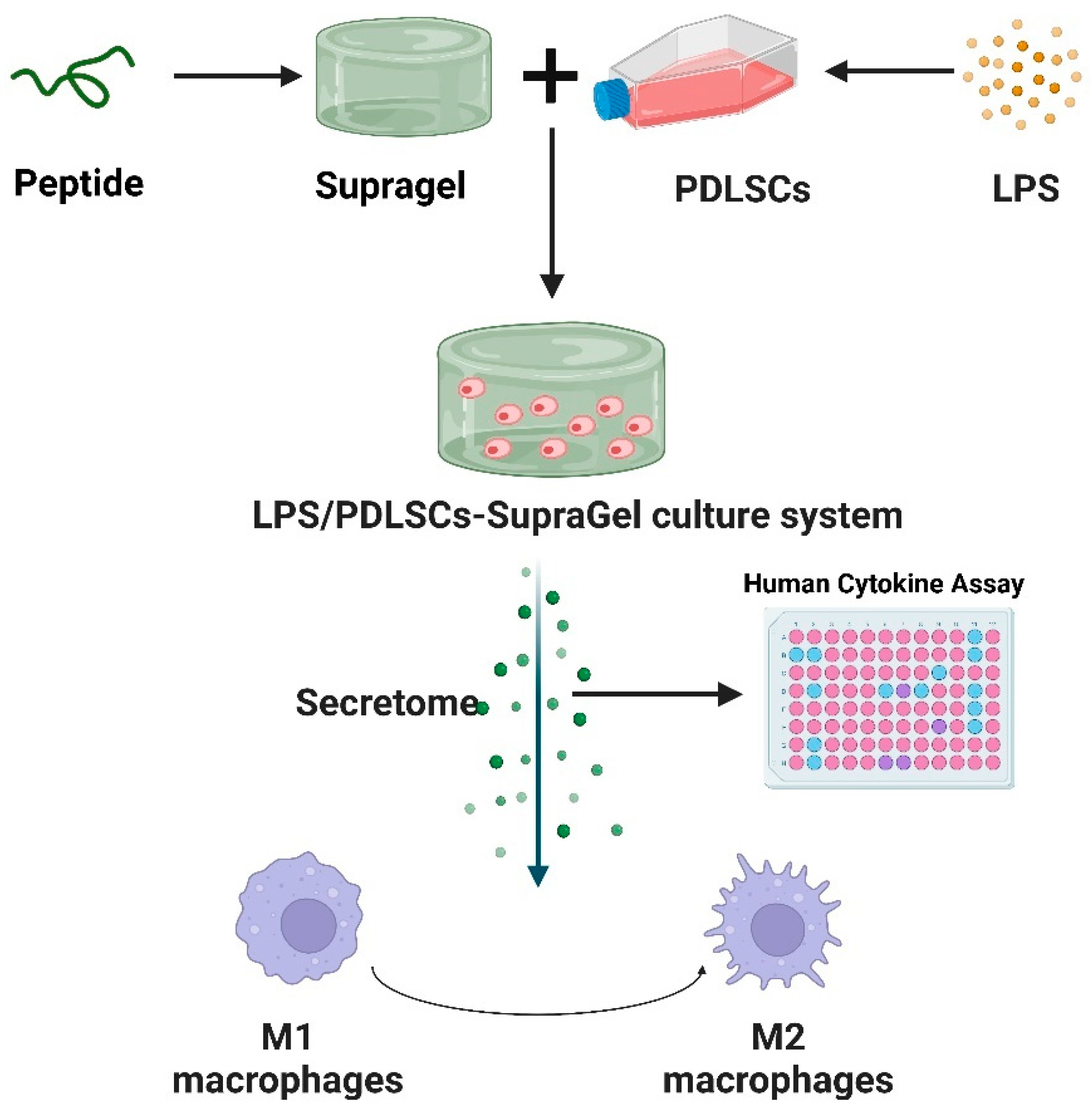
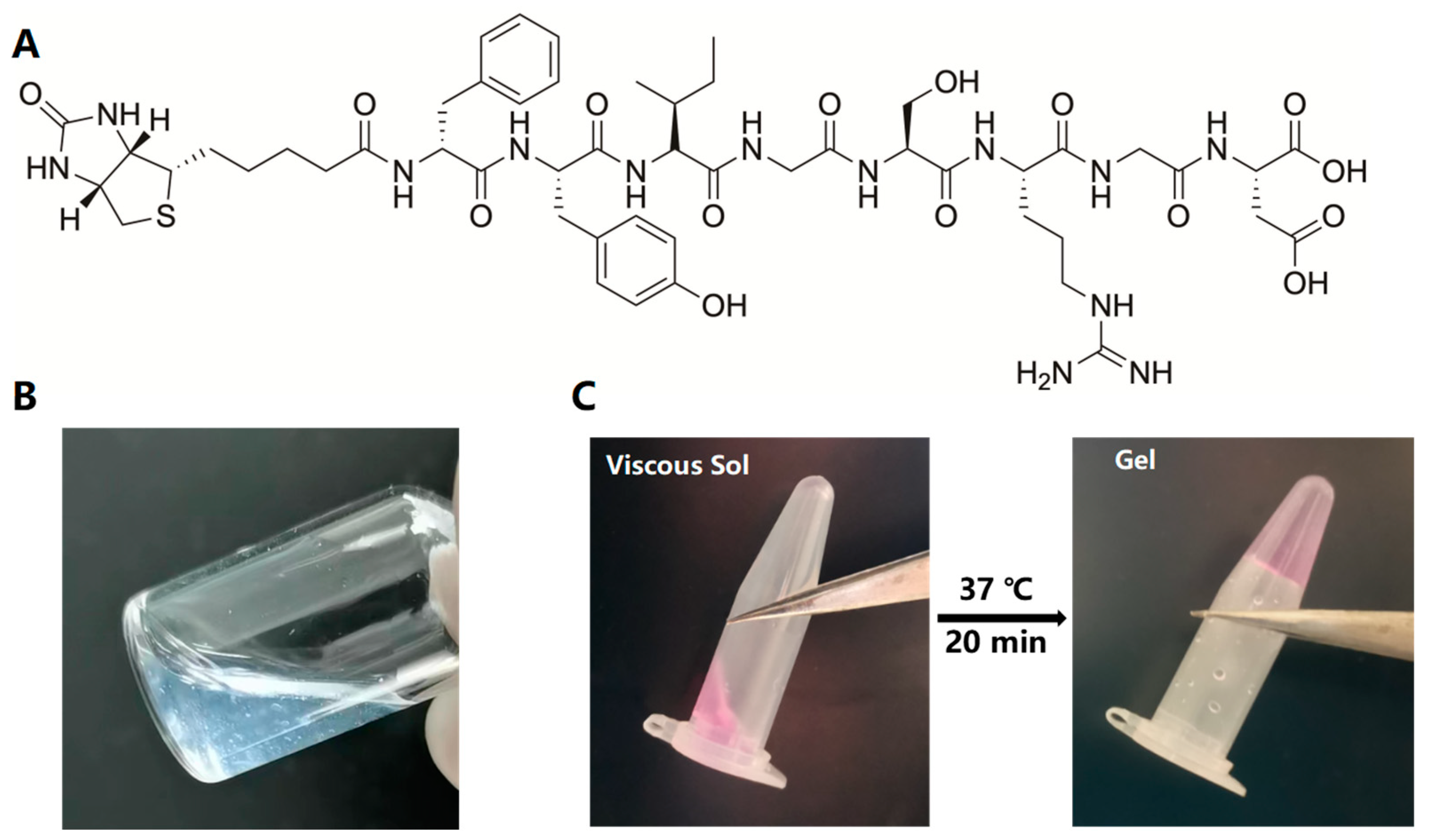
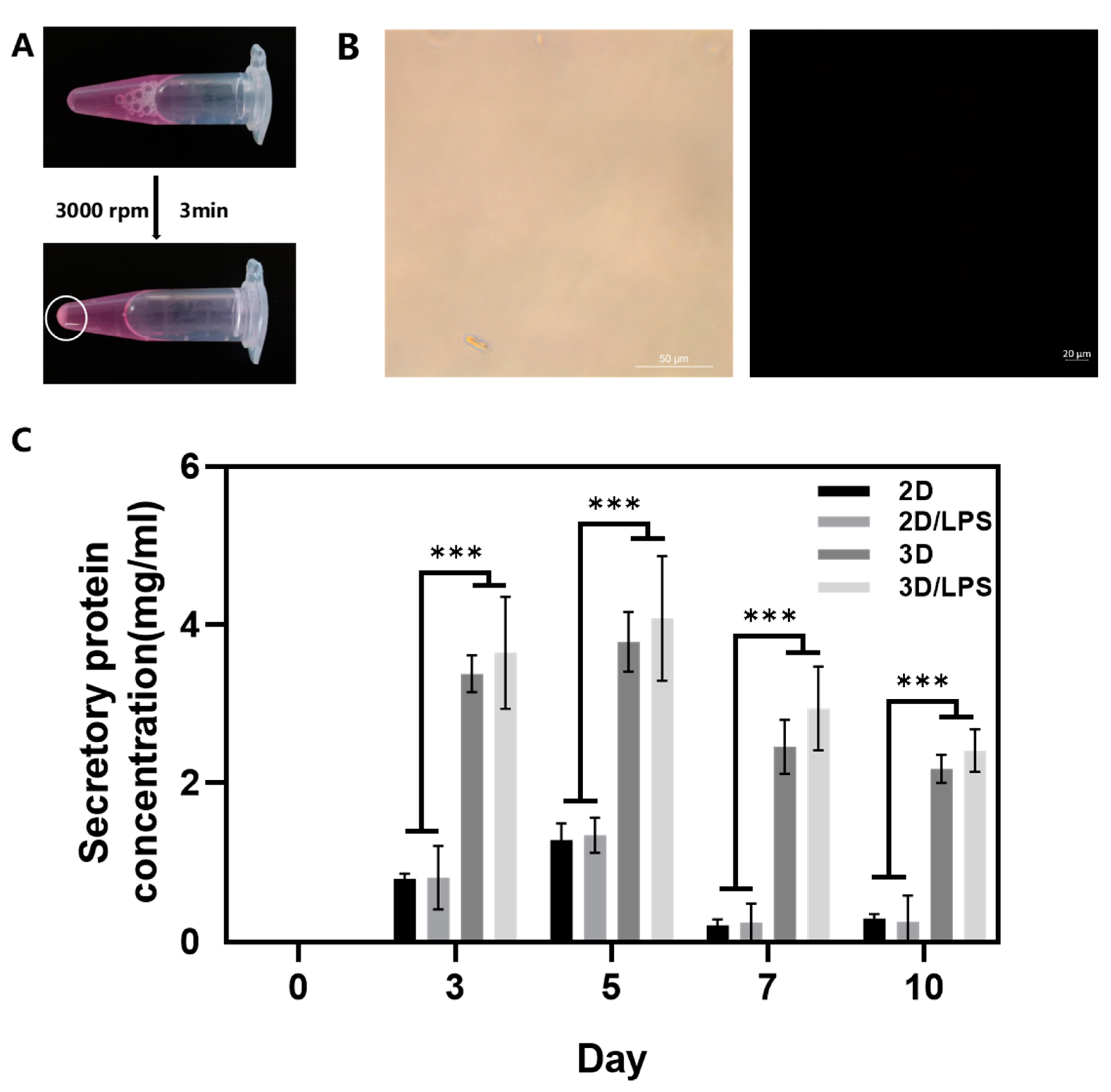
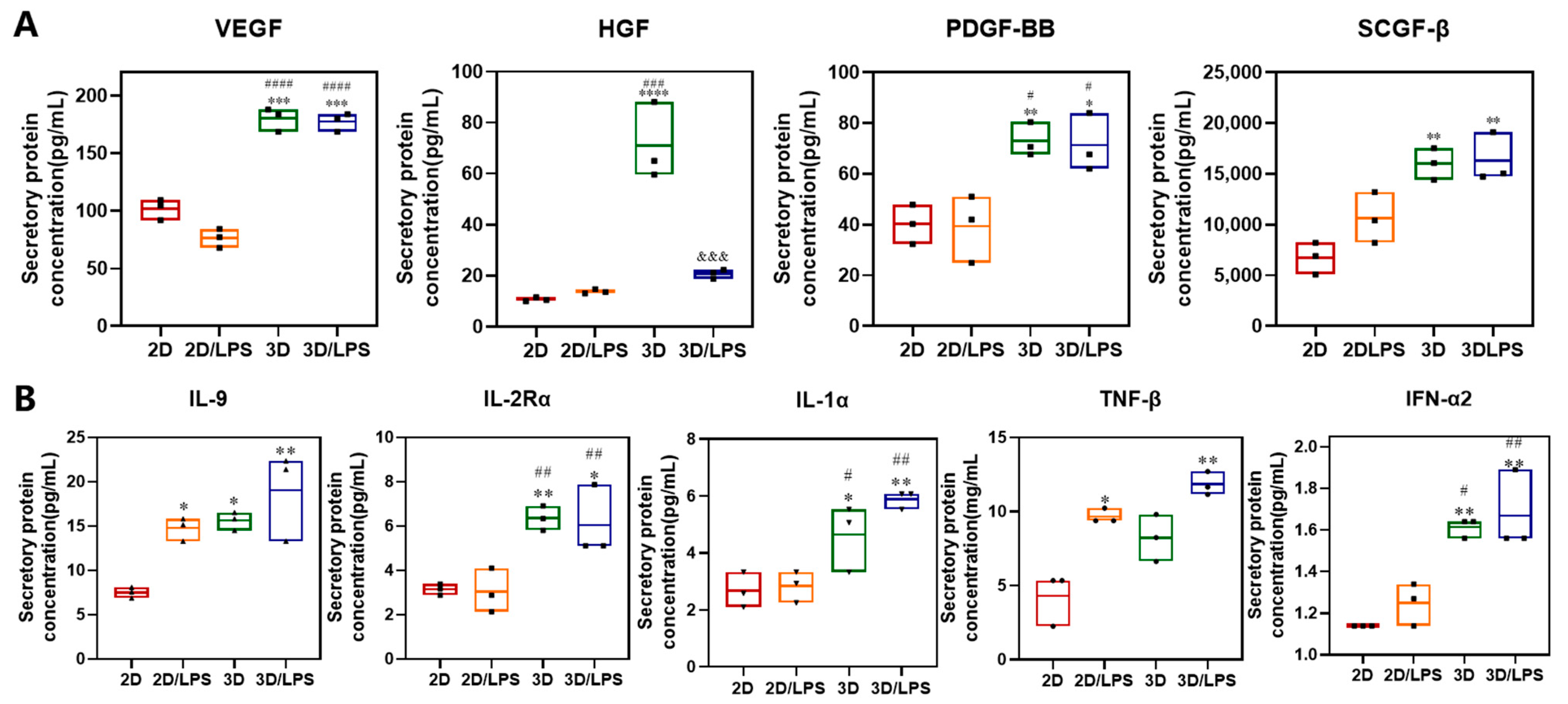
2.2. Construction of PDLSC–SupraGel Culture System
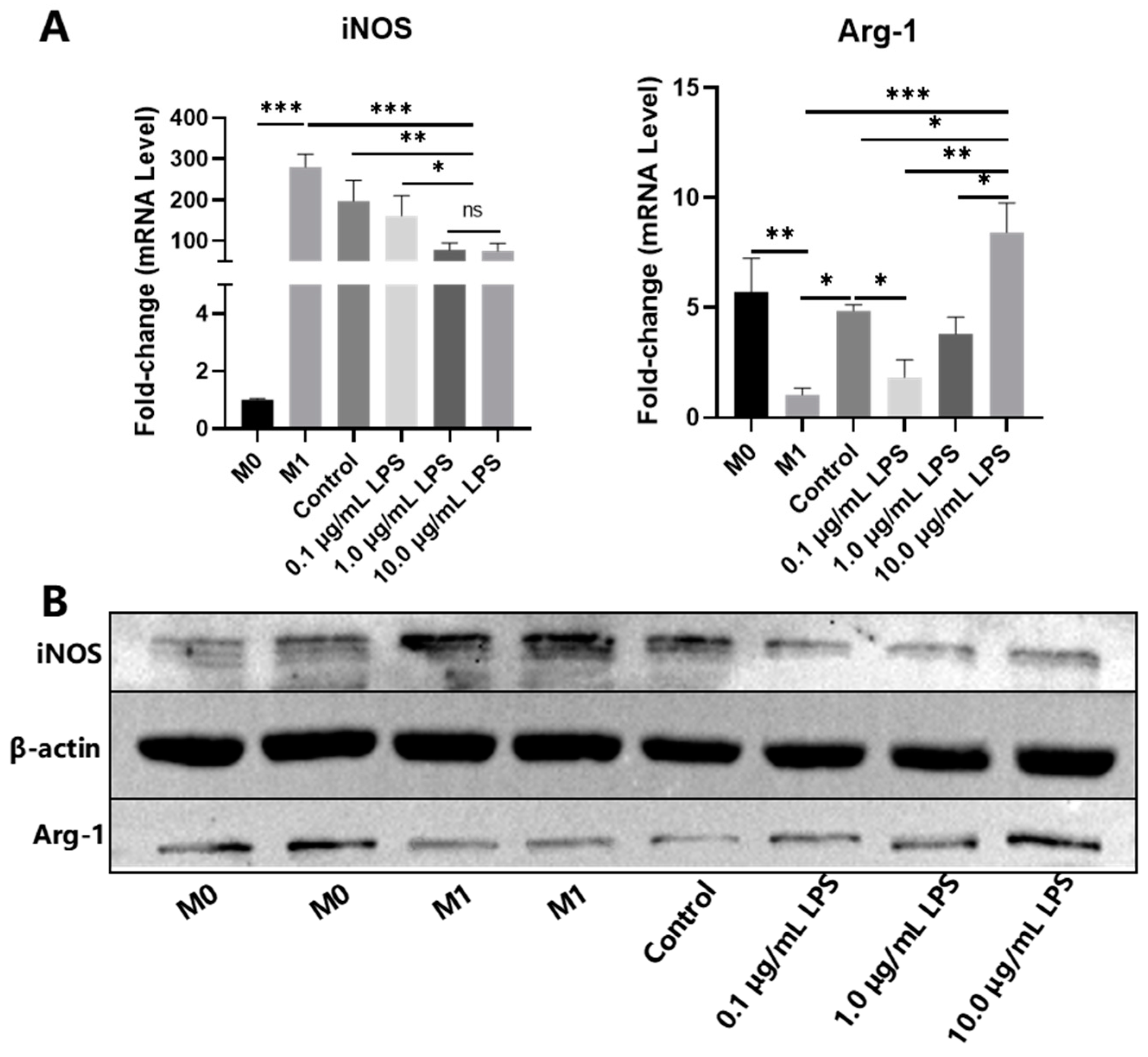
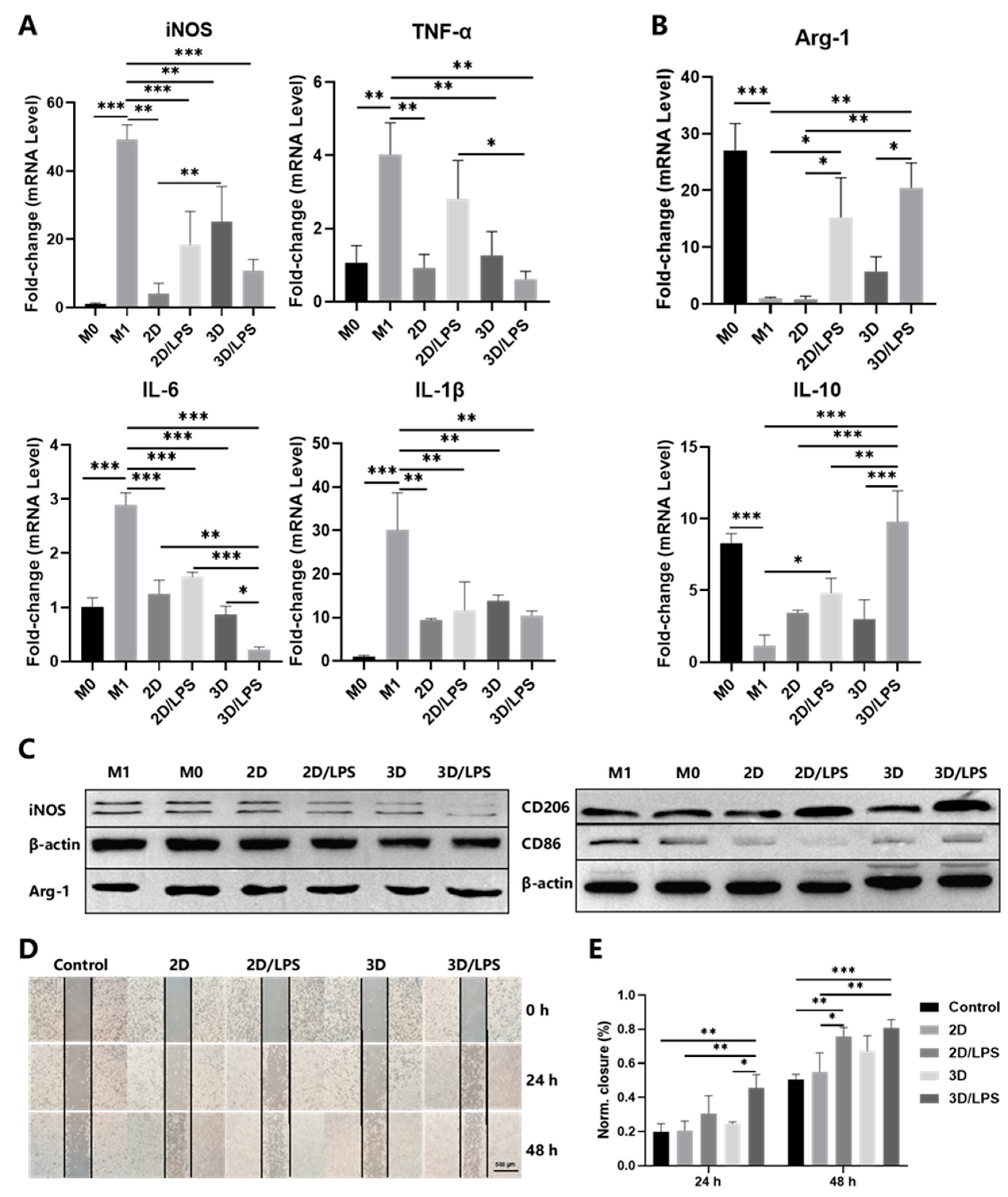
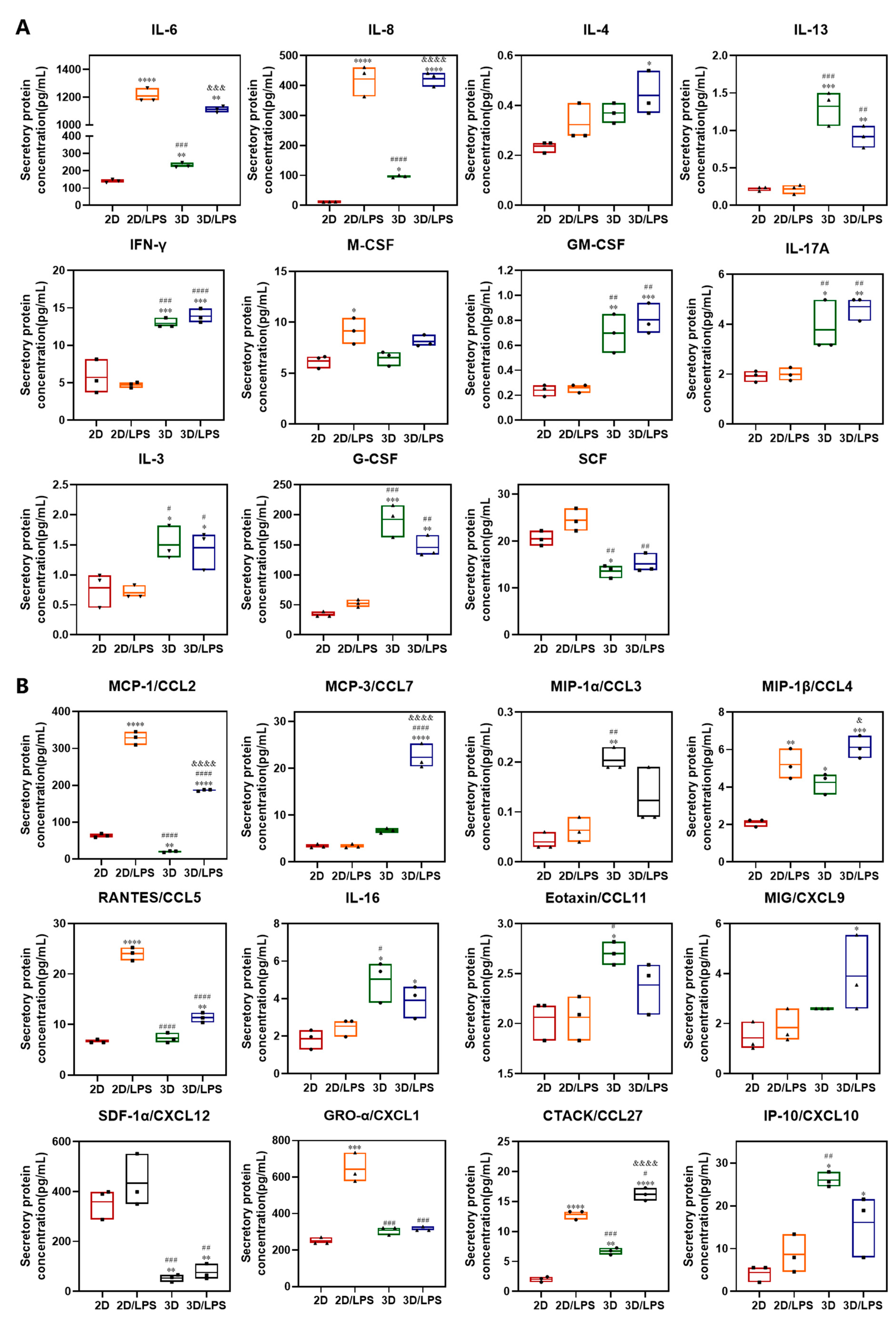
2.3. Effects of Secretome on Macrophage Polarization and Migration
3. Discussion
4. Materials and Methods
4.1. Proliferation Assay of Periodontal Ligament Stem Cells (PDLSCs)
4.2. Macrophage Polarization
4.3. LPS Pretreatment of PDLSCs and Conditioned Medium (CM) Culture of M1 Macrophages
4.4. Construction of a PDLSC–SupraGel Culture System
4.5. PDLSC Viability in SupraGel
4.6. Collection of Secretome
4.7. Secretome Culture of M1 Macrophages
4.8. Wound Healing
4.9. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.10. Western Blot
4.11. Multiplex Cytokine Assay of the Secretome
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, J.; Horng, T. Lipid metabolism in regulation of macrophage functions. Trends Cell Biol. 2020, 30, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef] [PubMed]
- Kwon, T.; Lamster, I.B.; Levin, L. Current concepts in the management of periodontitis. Int. Dent. J. 2021, 71, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Pang, X.; Li, Z.; Chen, Z.; Wang, Y. Immunomodulation in the treatment of periodontitis: Progress and perspectives. Front. Immunol. 2021, 12, 781378. [Google Scholar] [CrossRef] [PubMed]
- Sima, C.; Viniegra, A.; Glogauer, M. Macrophage immunomodulation in chronic osteolytic diseases-the case of periodontitis. J. Leukoc. Biol. 2019, 105, 473–487. [Google Scholar] [CrossRef]
- Sun, X.; Gao, J.; Meng, X.; Lu, X.; Zhang, L.; Chen, R. Polarized macrophages in periodontitis: Characteristics, function, and molecular signaling. Front. Immunol. 2021, 12, 763334. [Google Scholar] [CrossRef] [PubMed]
- Sarsenova, M.; Kim, Y.; Raziyeva, K.; Kazybay, B.; Ogay, V.; Saparov, A. Recent advances to enhance the immunomodulatory potential of mesenchymal stem cells. Front. Immunol. 2022, 13, 1010399. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Xu, C.; Cheng, W.; Zhao, Y.; Sui, L.; Zhao, Y. Pretreated mesenchymal stem cells and their secretome: Enhanced immunotherapeutic strategies. Int. J. Mol. Sci. 2023, 24, 1277. [Google Scholar] [CrossRef]
- Yang, Y.; Lee, E.H.; Yang, Z. Hypoxia-conditioned mesenchymal stem cells in tissue regeneration application. Tissue Eng. Part B Rev. 2022, 28, 966–977. [Google Scholar] [CrossRef]
- Mitchell, R.; Mellows, B.; Sheard, J.; Antonioli, M.; Kretz, O.; Chambers, D.; Zeuner, M.-T.; Tomkins, J.E.; Denecke, B.; Musante, L.; et al. Secretome of adipose-derived mesenchymal stem cells promotes skeletal muscle regeneration through synergistic action of extracellular vesicle cargo and soluble proteins. Stem Cell Res. Ther. 2019, 10, 116. [Google Scholar] [CrossRef]
- Eleuteri, S.; Fierabracci, A. Insights into the secretome of mesenchymal stem cells and its potential applications. Int. J. Mol. Sci. 2019, 20, 4597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.; Veenendaal, T.; Wiendels, M.; Ruiz-Zapata, A.M.; Van Laar, J.; Kyranas, R.; Enting, H.; Van Cranenbroek, B.; Koenen, H.J.P.M.; Mihaila, S.M.; et al. Synthetic extracellular matrices as a toolbox to tune stem cell secretome. ACS Appl. Mater. Interfaces 2020, 12, 56723–56730. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, M.E.; Rao, V.V.; Borelli, A.N.; Anseth, K.S. Engineering the MSC secretome: A hydrogel focused approach. Adv. Healthc. Mater. 2021, 10, e2001948. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Discher, D.E.; Péault, B.M.; Phinney, D.G.; Hare, J.M.; Caplan, A.I. Mesenchymal stem cell perspective: Cell biology to clinical progress. NPJ Regen. Med. 2019, 4, 22. [Google Scholar] [CrossRef] [Green Version]
- Asgari Taei, A.; Khodabakhsh, P.; Nasoohi, S.; Farahmandfar, M.; Dargahi, L. Paracrine effects of mesenchymal stem cells in ischemic stroke: Opportunities and challenges. Mol. Neurobiol. 2022, 59, 6281–6306. [Google Scholar] [CrossRef]
- Vizoso, F.J.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Wang, Y.; Li, Q.; Liu, K.; Hou, J.; Shao, C.; Wang, Y. Immunoregulatory mechanisms of mesenchymal stem and stromal cells in inflammatory diseases. Nat. Rev. Nephrol. 2018, 14, 493–507. [Google Scholar] [CrossRef]
- Weiss, A.R.R.; Dahlke, M.H. Immunomodulation by mesenchymal stem cells (MSCs): Mechanisms of action of living, apoptotic, and dead MSCs. Front. Immunol. 2019, 10, 1191. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, R.F.; Sá-Correia, I.; Valvano, M.A. Lipopolysaccharide modification in Gram-negative bacteria during chronic infection. FEMS Microbiol. Rev. 2016, 40, 480–493. [Google Scholar] [CrossRef] [Green Version]
- Ti, D.; Hao, H.; Tong, C.; Liu, J.; Dong, L.; Zheng, J.; Zhao, Y.; Liu, H.; Fu, X.; Han, W. LPS-preconditioned mesenchymal stromal cells modify macrophage polarization for resolution of chronic inflammation via exosome-shuttled let-7b. J. Transl. Med. 2015, 13, 308. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Zhang, F.; Chai, R.; Zhou, W.; Hu, M.; Liu, B.; Chen, X.; Liu, M.; Xu, Q.; Liu, N.; et al. Exosomes derived from pro-inflammatory bone marrow-derived mesenchymal stem cells reduce inflammation and myocardial injury via mediating macrophage polarization. J. Cell. Mol. Med. 2019, 23, 7617–7631. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Liu, Q.; Liu, L.; Huo, F.; Guo, S.; Tian, W. Lipopolysaccharide-preconditioned dental follicle stem cells derived small extracellular vesicles treating periodontitis via reactive oxygen species/mitogen-activated protein kinase signaling-mediated antioxidant effect. Int. J. Nanomed. 2022, 17, 799–819. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.Y.; Liu, Y.; Lu, Y.; Qin, Y.-R.; Di, G.-H.; Lei, Y.-H.; Liu, H.-X.; Li, Y.-Q.; Wu, C.; Hu, X.-W.; et al. Short-term memory of danger signals or environmental stimuli in mesenchymal stem cells: Implications for therapeutic potential. Cell. Mol. Immunol. 2016, 13, 369–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosh, T.J.; Ylöstalo, J.H.; Mohammadipoor, A.; Bazhanov, N.; Coble, K.; Claypool, K.; Lee, R.H.; Choi, H.; Prockop, D.J. Aggregation of human mesenchymal stromal cells (MSCs) into 3D spheroids enhances their antiinflammatory properties. Proc. Natl. Acad. Sci. USA 2010, 107, 13724–13729. [Google Scholar] [CrossRef] [Green Version]
- Ylöstalo, J.H.; Bartosh, T.J.; Coble, K.; Prockop, D.J. Human mesenchymal stem/stromal cells cultured as spheroids are self-activated to produce prostaglandin E2 that directs stimulated macrophages into an anti-inflammatory phenotype. Stem Cells 2012, 30, 2283–2296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartosh, T.J.; Ylöstalo, J.H.; Bazhanov, N.; Kuhlman, J.; Prockop, D.J. Dynamic compaction of human mesenchymal stem/precursor cells into spheres self-activates caspase-dependent IL1 signaling to enhance secretion of modulators of inflammation and immunity (PGE2, TSG6, and STC1). Stem Cells 2013, 31, 2443–2456. [Google Scholar] [CrossRef] [Green Version]
- Zimmermann, J.A.; Mcdevitt, T.C. Pre-conditioning mesenchymal stromal cell spheroids for immunomodulatory paracrine factor secretion. Cytotherapy 2014, 16, 331–345. [Google Scholar] [CrossRef]
- Bicer, M.; Cottrell, G.S.; Widera, D. Impact of 3D cell culture on bone regeneration potential of mesenchymal stromal cells. Stem Cell Res. Ther. 2021, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Ai, S.; Li, H.; Zheng, H.; Liu, J.; Gao, J.; Liu, J.; Chen, Q.; Yang, Z. A SupraGel for efficient production of cell spheroids. Sci. China Mater. 2022, 65, 1655–1661. [Google Scholar] [CrossRef]
- Miceli, V.; Bulati, M.; Iannolo, G.; Zito, G.; Gallo, A.; Conaldi, P.G. Therapeutic properties of mesenchymal stromal/stem cells: The need of cell priming for cell-free therapies in regenerative medicine. Int. J. Mol. Sci. 2021, 22, 763. [Google Scholar] [CrossRef]
- Gorgun, C.; Ceresa, D.; Lesage, R.; Villa, F.; Reverberi, D.; Balbi, C.; Santamaria, S.; Cortese, K.; Malatesta, P.; Geris, L.; et al. Dissecting the effects of preconditioning with inflammatory cytokines and hypoxia on the angiogenic potential of mesenchymal stromal cell (MSC)-derived soluble proteins and extracellular vesicles (EVs). Biomaterials 2021, 269, 120633. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Deng, M.; Hao, M.; Tang, J. Periodontal ligament stem cells in the periodontitis niche: Inseparable interactions and mechanisms. J. Leukoc. Biol. 2021, 110, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Waterman, R.S.; Tomchuck, S.L.; Henkle, S.L.; Betancourt, A.M. A new mesenchymal stem cell (MSC) paradigm: Polarization into a pro-inflammatory MSC1 or an Immunosuppressive MSC2 phenotype. PLoS ONE 2010, 5, e10088. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Li, J.; Sun, L.; Sun, Y.; Zhao, F.; Liu, P.; Peng, X.; Xuan, X.; Li, Y.; Wang, P.; et al. Exosomes derived from LPS-stimulated human thymic mesenchymal stromal cells enhance inflammation via thrombospondin-1. Biosci. Rep. 2021, 41, BSR20203573. [Google Scholar] [CrossRef] [PubMed]
- Kurte, M.; Vega-Letter, A.M.; Luz-Crawford, P.; Djouad, F.; Noël, D.; Khoury, M.; Carrión, F. Time-dependent LPS exposure commands MSC immunoplasticity through TLR4 activation leading to opposite therapeutic outcome in EAE. Stem Cell Res. Ther. 2020, 11, 416. [Google Scholar] [CrossRef]
- François, M.; Romieu-Mourez, R.; Li, M.; Galipeau, J. Human MSC suppression correlates with cytokine induction of indoleamine 2,3-dioxygenase and bystander M2 macrophage differentiation. Mol. Ther. 2012, 20, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Huang, F.; Wang, W.; Xie, Y.; Bin Wang, B. Interleukin-10 genetically modified clinical-grade mesenchymal stromal cells markedly reinforced functional recovery after spinal cord injury via directing alternative activation of macrophages. Cell. Mol. Biol. Lett. 2022, 27, 27. [Google Scholar] [CrossRef]
- Wang, Y.; Han, B.; Wang, Y.; Wang, C.; Zhang, H.; Xue, J.; Wang, X.; Niu, T.; Niu, Z.; Chen, Y. Mesenchymal stem cell-secreted extracellular vesicles carrying TGF-β1 up-regulate miR-132 and promote mouse M2 macrophage polarization. J. Cell. Mol. Med. 2020, 24, 12750–12764. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.-Y.; Song, T.; Zhang, L.; Eirin, A.; Conley, S.; Tang, H.; Saadiq, I.; Jordan, K.; Lerman, A.; et al. Mesenchymal stem cells protect renal tubular cells via TSG-6 regulating macrophage function and phenotype switching. Am. J. Physiol. Renal. Physiol. 2021, 320, F454–F463. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wu, F.; Zhou, Y.; Bao, Z.; Li, H.; Zheng, P.; Zhao, S. Gastric cancer-derived mesenchymal stromal cells trigger M2 macrophage polarization that promotes metastasis and EMT in gastric cancer. Cell Death Dis. 2019, 10, 918. [Google Scholar] [CrossRef] [Green Version]
- Rao, V.V.; Vu, M.K.; Ma, H.; Killaars, A.R.; Anseth, K.S. Rescuing mesenchymal stem cell regenerative properties on hydrogel substrates post serial expansion. Bioeng. Transl. Med. 2019, 4, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, Q.; Xu, N.; Xu, D.; Dong, M.; Shi, X.; Wang, Y.; Hao, Z.; Zhu, S.; Zhao, D.; Jin, H.; et al. Comparison of senescence-related changes between three- and two-dimensional cultured adipose-derived mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 226. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Shi, T.; Xu, A.; Zhang, L. 3D spheroid culture enhances survival and therapeutic capacities of MSCs injected into ischemic kidney. J. Cell. Mol. Med. 2016, 20, 1203–1213. [Google Scholar] [CrossRef] [Green Version]
- Miranda, J.P.; Camões, S.P.; Gaspar, M.M.; Rodrigues, J.; Carvalheiro, M.; Bárcia, R.N.; da Cruz, P.E.; Cruz, H.; Simões, S.; Santos, J.M. The secretome derived from 3D-cultured umbilical cord tissue MSCs counteracts manifestations typifying rheumatoid arthritis. Front. Immunol. 2019, 10, 18. [Google Scholar] [CrossRef]
- Cesarz, Z.; Tamama, K. Spheroid culture of mesenchymal stem cells. Stem Cells Int. 2016, 2016, 9176357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drzeniek, N.M.; Mazzocchi, A.; Schlickeiser, S.; Forsythe, S.D.; Moll, G.; Geißler, S.; Reinke, P.; Gossen, M.; Gorantla, V.S.; Volk, H.-D.; et al. Bio-instructive hydrogel expands the paracrine potency of mesenchymal stem cells. Biofabrication 2021, 13, 045002. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.; Feng, W.; Cao, X.; Ren, K.; Quan, M.; Chen, A.; Xu, C.; Qiu, Y.; Cao, J.; Li, X.; et al. Genistein inhibits stemness of SKOV3 cells induced by macrophages co-cultured with ovarian cancer stem-like cells through IL-8/STAT3 axis. J. Exp. Clin. Cancer Res. 2019, 38, 19. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Ma, T.; Lin, Y.; Lu, X.; Zhang, C.; Chen, S.; Jian, Z. IL-6/STAT3 pathway intermediates M1/M2 macrophage polarization during the development of hepatocellular carcinoma. J. Cell Biochem. 2018, 119, 9419–9432. [Google Scholar] [CrossRef]
- Liu, C.; Xu, Y.; Lu, Y.; Du, P.; Li, X.; Wang, C.; Guo, P.; Diao, L.; Lu, G. Mesenchymal stromal cells pretreated with proinflammatory cytokines enhance skin wound healing via IL-6-dependent M2 polarization. Stem Cell Res. Ther. 2022, 13, 414. [Google Scholar] [CrossRef]
- Xie, Z.; Hao, H.; Tong, C.; Cheng, Y.; Liu, J.; Pang, Y.; Si, Y.; Guo, Y.; Zang, L.; Mu, Y.; et al. Human umbilical cord-derived mesenchymal stem cells elicit macrophages into an anti-inflammatory phenotype to alleviate insulin resistance in type 2 diabetic rats. Stem Cells 2016, 34, 627–639. [Google Scholar] [CrossRef]
- Li, D.; Li, J.; Liu, H.; Zhai, L.; Hu, W.; Xia, N.; Tang, T.; Jiao, J.; Lv, B.; Nie, S.; et al. Pathogenic Tconvs promote inflammatory macrophage polarization through GM-CSF and exacerbate abdominal aortic aneurysm formation. FASEB J. 2022, 36, e22172. [Google Scholar] [CrossRef] [PubMed]
- Orecchioni, M.; Ghosheh, Y.; Pramod, A.B.; Ley, K. Macrophage polarization: Different gene signatures in M1(LPS+) vs. classically and M2(LPS-) vs. alternatively activated macrophages. Front. Immunol. 2019, 10, 1084. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.E.; Ahn, S.H.; Marks, R.M.; Monsanto, S.P.; Fazleabas, A.T.; Koti, M.; Tayade, C. IL-17A modulates peritoneal macrophage recruitment and M2 Polarization in endometriosis. Front. Immunol. 2020, 11, 108. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, K.; Seo, N.; Torii, M.; Ma, N.; Muraoka, D.; Tawara, I.; Masuya, M.; Tanaka, K.; Takei, Y.; Shiku, H.; et al. Interleukin-17 induces an atypical M2-like macrophage subpopulation that regulates intestinal inflammation. PLoS ONE 2014, 9, e108494. [Google Scholar] [CrossRef]
- Zhang, Q.; Atsuta, I.; Liu, S.; Chen, C.; Shi, S.; Shi, S.; Le, A.D. IL-17-mediated M1/M2 macrophage alteration contributes to pathogenesis of bisphosphonate-related osteonecrosis of the jaws. Clin. Cancer Res. 2013, 19, 3176–3188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, T.A.; Zhao, C.; Pavicic, P.G.J.; Datta, S. Myeloid colony-stimulating factors as regulators of macrophage polarization. Front. Immunol. 2014, 5, 554. [Google Scholar] [CrossRef] [Green Version]
- Mehta, H.M.; Malandra, M.; Corey, S.J. G-CSF and GM-CSF in neutropenia. J. Immunol. 2015, 195, 1341–1349. [Google Scholar] [CrossRef] [Green Version]
- Dhaiban, S.; Al-Ani, M.; Elemam, N.M.; Maghazachi, A.A. Targeting chemokines and chemokine receptors in multiple sclerosis and experimental autoimmune encephalomyelitis. J. Inflamm. Res. 2020, 13, 619–633. [Google Scholar] [CrossRef]
- Huang, Y.; Du, K.L.; Guo, P.Y.; Zhao, R.M.; Wang, B.; Zhao, X.L.; Zhang, C.Q. IL-16 regulates macrophage polarization as a target gene of mir-145-3p. Mol. Immunol. 2019, 107, 1–9. [Google Scholar] [CrossRef]
- Li, C.; Xu, M.; Coyne, J.; Wang, W.B.; Davila, M.L.; Wang, Y.; Xiong, N. Psoriasis-associated impairment of CCL27/CCR10-derived regulation leads to IL-17A/IL-22-producing skin T-cell overactivation. J. Allergy Clin. Immunol. 2021, 147, 759–763.e9. [Google Scholar] [CrossRef]
- Li, X.; Bu, W.; Meng, L.; Liu, X.; Wang, S.; Jiang, L.; Ren, M.; Fan, Y.; Sun, H. CXCL12/CXCR4 pathway orchestrates CSC-like properties by CAF recruited tumor associated macrophage in OSCC. Exp. Cell Res. 2019, 378, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Vimalraj, S. A concise review of VEGF, PDGF, FGF, Notch, angiopoietin, and HGF signalling in tumor angiogenesis with a focus on alternative approaches and future directions. Int. J. Biol. Macromol. 2022, 221, 1428–1438. [Google Scholar] [CrossRef]
- Desole, C.; Gallo, S.; Vitacolonna, A.; Montarolo, F.; Bertolotto, A.; Vivien, D.; Comoglio, P.; Crepaldi, T. HGF and MET: From brain development to neurological disorders. Front. Cell Dev. Biol. 2021, 9, 683609. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Liu, G.; Liu, X.; Zhou, Y.; Sun, Q.; Zhen, G.; Wang, X.; Hu, Y.; Gao, P.; Demehri, S.; et al. Angiogenesis stimulated by elevated PDGF-BB in subchondral bone contributes to osteoarthritis development. JCI Insight 2020, 5, e135446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.M.; Feng, F.E.; Wang, Q.M.; Zhu, X.L.; Fu, H.X.; Xu, L.P.; Liu, K.Y.; Huang, X.J.; Zhang, X.H. Platelet-derived growth factor-bb protects mesenchymal stem cells (MSCs) derived from immune thrombocytopenia patients against apoptosis and senescence and maintains MSC-mediated immunosuppression. Stem Cells Transl. Med. 2016, 5, 1631–1643. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, C.; Xie, L.; Wang, X.; Zhang, L.; Chen, C.; Jiang, B. Traditional two-dimensional mesenchymal stem cells (MSCs) are better than spheroid MSCs on promoting retinal ganglion cells survival and axon regeneration. Exp. Eye Res. 2019, 185, 107699. [Google Scholar] [CrossRef]
- Li, J.; Chen, S.; Xiao, X.; Zhao, Y.; Ding, W.; Li, X.C. IL-9 and Th9 cells in health and diseases—From tolerance to immunopathology. Cytokine Growth Factor Rev. 2017, 37, 47–55. [Google Scholar] [CrossRef]
- Permanyer, M.; Bošnjak, B.; Glage, S.; Friedrichsen, M.; Floess, S.; Huehn, J.; Patzer, G.E.; Odak, I.; Eckert, N.; Zargari, R.; et al. Efficient IL-2R signaling differentially affects the stability, function, and composition of the regulatory T-cell pool. Cell Mol. Immunol. 2021, 18, 398–414. [Google Scholar] [CrossRef]
- Cao, Z.; Xie, Y.; Yu, L.; Li, Y.; Wang, Y. Hepatocyte growth factor (HGF) and stem cell factor (SCF) maintained the stemness of human bone marrow mesenchymal stem cells (hBMSCs) during long-term expansion by preserving mitochondrial function via the PI3K/AKT, ERK1/2, and STAT3 signaling pathways. Stem Cell Res. Ther. 2020, 11, 329. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | |
|---|---|---|
| Homo GAPDH | Forward | CCGCATCTTCTTTTGCGTCG |
| Reverse | GGACTCCACGACGTACTCAG | |
| Homo IL-6 | Forward | ATGAACTCCTTCTCCACAAGCGC |
| Reverse | GAAGAGCCCTCAGGCTGGACTG | |
| Homo IL-8 | Forward | ACACTGCGCCAACACAGAAATTA |
| Reverse | TTTGCTTGAAGTTTCACTGGCATC | |
| Homo IL-10 | Forward | GATCTCCGAGATGCCTTCAG |
| Reverse | ATCGATGACAGCGCCGTAGC | |
| Homo IDO | Forward | GCCCTTCAAGTGTTTCACCAA |
| Reverse | CCAGCCAGACAAATATATGCGA | |
| Homo TSG-6 | Forward | TGTCTGTGCTGCTGGATGGAT |
| Reverse | TGTGGGTTGTAGCAATAGGCAT | |
| Homo TGF-β | Forward | CTAATGGTGGAAACCCACAACG |
| Reverse | TATCGCCAGGAATTGTTGCTG | |
| Mus GAPDH | Forward | CACGACATACTCAGCACCAG |
| Reverse | TCCAGTATGACTCTACCCAC | |
| Mus Arg-1 | Forward | ATGTCCCTAATGACAGCTCCT |
| Reverse | GCTTCCAACTGCCAGACTGT | |
| Mus iNOS | Forward | CCCTATTTCACCTGCAACAG |
| Reverse | GCTTGTCCAGGGATTCTGG | |
| Mus IL-β | Forward | TGCCACCTTTTGACAGTGATG |
| Reverse | TGATGTGCTGCTGCGAGATT | |
| Mus TNF-α | Forward | TAGCCCACGTCGTAGCAAAC |
| Reverse | GCAGCCTTGTCCCTTGAAGA | |
| Mus IL-6 | Forward | TGATGGATGCTACCAAACTGGA |
| Reverse | TGTGACTCCAGCTTATCTCTTGG | |
| Mus IL-10 | Forward | GCTGGACAACATACTGCTAACCG |
| Reverse | CACAGGGGAGAAATCGATGACAG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, Y.; Ai, S.; Shen, Y.; Cheng, W.; Xu, C.; Sui, L.; Zhao, Y. Regulatory Effects of Three-Dimensional Cultured Lipopolysaccharide-Pretreated Periodontal Ligament Stem Cell-Derived Secretome on Macrophages. Int. J. Mol. Sci. 2023, 24, 6981. https://doi.org/10.3390/ijms24086981
Su Y, Ai S, Shen Y, Cheng W, Xu C, Sui L, Zhao Y. Regulatory Effects of Three-Dimensional Cultured Lipopolysaccharide-Pretreated Periodontal Ligament Stem Cell-Derived Secretome on Macrophages. International Journal of Molecular Sciences. 2023; 24(8):6981. https://doi.org/10.3390/ijms24086981
Chicago/Turabian StyleSu, Yuran, Sifan Ai, Youqing Shen, Wen Cheng, Chenyu Xu, Lei Sui, and Yanhong Zhao. 2023. "Regulatory Effects of Three-Dimensional Cultured Lipopolysaccharide-Pretreated Periodontal Ligament Stem Cell-Derived Secretome on Macrophages" International Journal of Molecular Sciences 24, no. 8: 6981. https://doi.org/10.3390/ijms24086981