
Structural Insights into the Binding of Red Fluorescent Protein mCherry-Specific Nanobodies
 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Protein Expression, Purification, and Characterization
2.2. Crystal Structures of a Complex of mCherry and Nanobodies
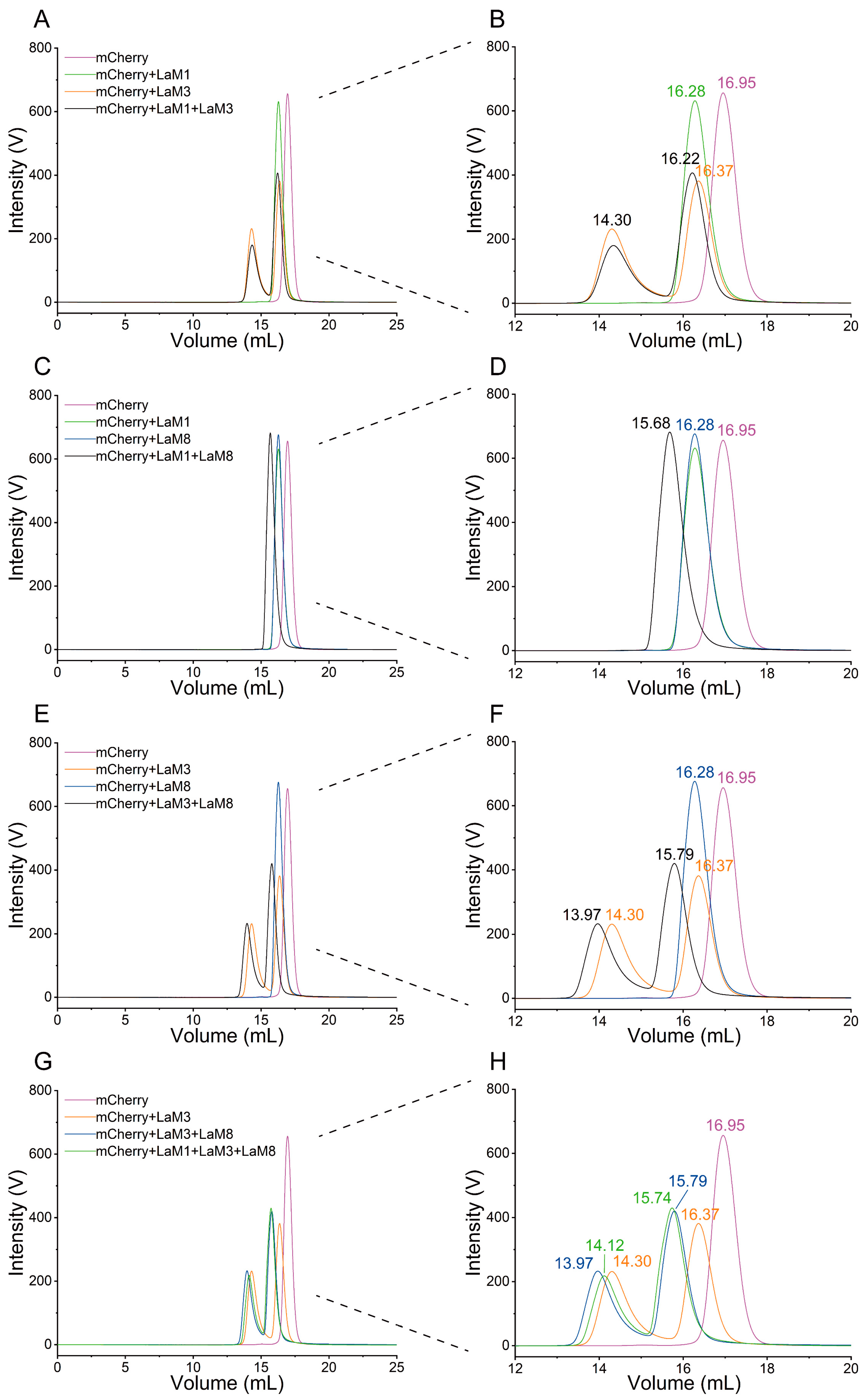
2.3. Epitope Mapping of LaM Nanobodies Binding to mCherry by FSEC
2.4. Binding Kinetics and Thermal Dynamics between mCherry and Different LaM Nanobodies
2.5. Comparison of the Emission Fluorescence Spectrum of mCherry Binding with Different LaMs
2.6. Design and Verification of Multivalent Nanobodies Binding with mCherry
3. Discussion
3.1. Comparison of the Binding Mode of LaM Nanobodies to mCherry
3.2. Structural Insights of the Binding between DsRed-LaM3 and DsRed-LaM4
3.3. Possible Applications of Multimeric Tandem Nanobodies
4. Materials and Methods
4.1. Protein Expression
4.2. Protein Purification
4.3. Crystallization and Structural Determination of mCherry-LaM1, mCherry-LaM3, and mCherry-LaM8
4.4. Verification of Related Protein or Protein Complex by Mass Spectrometry
4.5. Fluorescence Detected Size Exclusion Chromatography (FSEC)
4.6. Affinity and Kinetic Parameter Determination by BLI (OctetRed96)
4.7. Isothermal Titration Calorimetry (ITC)
4.8. Fluorescence Spectroscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shimomura, O. Discovery of Green Fluorescent Protein (GFP) (Nobel Lecture). Angew. Chem. Int. Ed. 2009, 48, 5590–5602. [Google Scholar] [CrossRef]
- Tan, J.X.; Finkel, T. A phosphoinositide signalling pathway mediates rapid lysosomal repair. Nature 2022, 609, 815–821. [Google Scholar] [CrossRef]
- Csibra, E.; Stan, G.-B. Absolute protein quantification using fluorescence measurements with FPCountR. Nat. Commun. 2022, 13, 6600. [Google Scholar] [CrossRef]
- Lin, C.S.; Shih, J.R.; Hsu, C.H.; Wen, Z.H.; Chakraborty, C. Expression of Green Fluorescence Protein (GFP) in Zebrafish Muscle through Injection: A Gene Therapy Model. Nat. Preced. 2008. [Google Scholar] [CrossRef]
- Shakhova, E.S.; Markina, N.M.; Mitiouchkina, T.; Bugaeva, E.N.; Karataeva, T.A.; Palkina, K.A.; Fakhranurova, L.I.; Yampolsky, I.V.; Sarkisyan, K.S.; Mishin, A.S. Systematic Comparison of Plant Promoters in Nicotiana spp. Expression Systems. Int. J. Mol. Sci. 2022, 23, 15441. [Google Scholar] [CrossRef] [PubMed]
- Pollok, B.A.; Heim, R. Using GFP in FRET-based applications. Trends Cell Biol. 1999, 9, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Słabicki, M.; Yoon, H.; Koeppel, J.; Nitsch, L.; Roy Burman, S.S.; Di Genua, C.; Donovan, K.A.; Sperling, A.S.; Hunkeler, M.; Tsai, J.M.; et al. Small-molecule-induced polymerization triggers degradation of BCL6. Nature 2020, 588, 164–168. [Google Scholar] [CrossRef]
- Oliinyk, O.S.; Baloban, M.; Clark, C.L.; Carey, E.; Pletnev, S.; Nimmerjahn, A.; Verkhusha, V.V. Single-domain near-infrared protein provides a scaffold for antigen-dependent fluorescent nanobodies. Nat. Methods 2022, 19, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Liang, Q.N.; Du, S.Q.; Hu, X.J.; Ding, Y. The crystal structure of red fluorescent protein TagRFP-T reveals the mechanism of its superior photostability. Biochem. Biophys. Res. Commun. 2016, 477, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Leben, R.; Lindquist, R.L.; Hauser, A.E.; Niesner, R.; Rakhymzhan, A. Two-Photon Excitation Spectra of Various Fluorescent Proteins within a Broad Excitation Range. Int. J. Mol. Sci. 2022, 23, 13407. [Google Scholar] [CrossRef]
- Hall, C.; von Grabowiecki, Y.; Pearce, S.P.; Dive, C.; Bagley, S.; Muller, P.A.J. iRFP (near-infrared fluorescent protein) imaging of subcutaneous and deep tissue tumours in mice highlights differences between imaging platforms. Cancer Cell Int. 2021, 21, 247. [Google Scholar] [CrossRef]
- Zipfel, W.R.; Williams, R.M.; Webb, W.W. Nonlinear magic: Multiphoton microscopy in the biosciences. Nat. Biotechnol. 2003, 21, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Subach, O.M.; Vlaskina, A.V.; Agapova, Y.K.; Piatkevich, K.D.; Patrushev, M.V.; Samygina, V.R.; Subach, F.V. LSSmScarlet2 and LSSmScarlet3, Chemically Stable Genetically Encoded Red Fluorescent Proteins with a Large Stokes’ Shift. Int. J. Mol. Sci. 2022, 23, 11051. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, M.R.; Chen, T.Y.; Wu, T.C.; Liu, S.W.; Hsu, C.H.; Liou, G.G.; Kao, Y.Y.; Dong, G.C.; Chu, P.H.; et al. Cardiac fibrosis in mouse expressing DsRed tetramers involves chronic autophagy and proteasome degradation insufficiency. Oncotarget 2016, 7, 54274–54289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaner, N.C.; Campbell, R.E.; Steinbach, P.A.; Giepmans, B.N.; Palmer, A.E.; Tsien, R.Y. Improved monomeric red, orange and yellow fluorescent proteins derived from Discosoma sp. red fluorescent protein. Nat. Biotechnol. 2004, 22, 1567–1572. [Google Scholar] [CrossRef]
- Shui, B.; Ozer, A.; Zipfel, W.; Sahu, N.; Singh, A.; Lis, J.T.; Shi, H.; Kotlikoff, M.I. RNA aptamers that functionally interact with green fluorescent protein and its derivatives. Nucleic Acids Res. 2012, 40, e39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chevrel, A.; Urvoas, A.; de la Sierra-Gallay, I.L.; Aumont-Nicaise, M.; Moutel, S.; Desmadril, M.; Perez, F.; Gautreau, A.; van Tilbeurgh, H.; Minard, P.; et al. Specific GFP-binding artificial proteins (αRep): A new tool for in vitro to live cell applications. Biosci. Rep. 2015, 35, e00223. [Google Scholar] [CrossRef] [PubMed]
- Brauchle, M.; Hansen, S.; Caussinus, E.; Lenard, A.; Ochoa-Espinosa, A.; Scholz, O.; Sprecher, S.G.; Plückthun, A.; Affolter, M. Protein interference applications in cellular and developmental biology using DARPins that recognize GFP and mCherry. Biol. Open 2014, 3, 1252–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubala, M.H.; Kovtun, O.; Alexandrov, K.; Collins, B.M. Structural and thermodynamic analysis of the GFP:GFP-nanobody complex. Protein Sci. A Publ. Protein Soc. 2010, 19, 2389–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourelis, J.; Marchal, C.; Posbeyikian, A.; Harant, A.; Kamoun, S. NLR immune receptor–nanobody fusions confer plant disease resistance. Science 2023, 379, 934–939. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, Y.; Ding, Y.; Hattori, M. Structure-based engineering of anti-GFP nanobody tandems as ultra-high-affinity reagents for purification. Sci. Rep. 2020, 10, 6239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirchhofer, A.; Helma, J.; Schmidthals, K.; Frauer, C.; Cui, S.; Karcher, A.; Pellis, M.; Muyldermans, S.; Casas-Delucchi, C.S.; Cardoso, M.C.; et al. Modulation of protein properties in living cells using nanobodies. Nat. Struct. Mol. Biol. 2010, 17, 133–138. [Google Scholar] [CrossRef]
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Wang, Z.; Cheng, S.; Zhang, Y.; Jiang, H.; Liu, R.; Ding, Y. Structural insights into two distinct nanobodies recognizing the same epitope of green fluorescent protein. Biochem. Biophys. Res. Commun. 2021, 565, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Pleiner, T.; Bates, M.; Trakhanov, S.; Lee, C.-T.; Schliep, J.E.; Chug, H.; Böhning, M.; Stark, H.; Urlaub, H.; Görlich, D. Nanobodies: Site-specific labeling for super-resolution imaging, rapid epitope-mapping and native protein complex isolation. eLife 2015, 4, e11349. [Google Scholar] [CrossRef]
- Muyldermans, S. Applications of Nanobodies. Annu. Rev. Anim. Biosci. 2021, 9, 401–421. [Google Scholar] [CrossRef]
- Meng, J.; Xu, C.; Lafon, P.-A.; Roux, S.; Mathieu, M.; Zhou, R.; Scholler, P.; Blanc, E.; Becker, J.A.J.; Le Merrer, J.; et al. Nanobody-based sensors reveal a high proportion of mGlu heterodimers in the brain. Nat. Chem. Biol. 2022, 18, 894–903. [Google Scholar] [CrossRef]
- Nachit, M.; Montemagno, C.; Clerc, R.; Ahmadi, M.; Briand, F.; Bacot, S.; Devoogdt, N.; Serdjebi, C.; Ghezzi, C.; Sulpice, T.; et al. Molecular imaging of liver inflammation using an anti-VCAM-1 nanobody. Nat. Commun. 2023, 14, 1062. [Google Scholar] [CrossRef]
- Xu, J.; Xu, K.; Jung, S.; Conte, A.; Lieberman, J.; Muecksch, F.; Lorenzi, J.C.C.; Park, S.; Schmidt, F.; Wang, Z.; et al. Nanobodies from camelid mice and llamas neutralize SARS-CoV-2 variants. Nature 2021, 595, 278–282. [Google Scholar] [CrossRef]
- Nagata, K.; Utsumi, D.; Asaka, M.N.; Maeda, R.; Shirakawa, K.; Kazuma, Y.; Nomura, R.; Horisawa, Y.; Yanagida, Y.; Kawai, Y.; et al. Intratracheal trimerized nanobody cocktail administration suppresses weight loss and prolongs survival of SARS-CoV-2 infected mice. Commun. Med. 2022, 2, 152. [Google Scholar] [CrossRef]
- Liu, M.; Li, L.; Jin, D.; Liu, Y. Nanobody-A versatile tool for cancer diagnosis and therapeutics. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnology 2021, 13, e1697. [Google Scholar] [CrossRef] [PubMed]
- Bao, C.; Gao, Q.; Li, L.L.; Han, L.; Zhang, B.; Ding, Y.; Song, Z.; Zhang, R.; Zhang, J.; Wu, X.H. The Application of Nanobody in CAR-T Therapy. Biomolecules 2021, 11, 238. [Google Scholar] [CrossRef]
- Helma, J.; Cardoso, M.C.; Muyldermans, S.; Leonhardt, H. Nanobodies and recombinant binders in cell biology. J. Cell Biol. 2015, 209, 633–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, M.J.; Papasergi-Scott, M.M.; He, F.; Seven, A.B.; Meyerowitz, J.G.; Panova, O.; Peroto, M.C.; Che, T.; Skiniotis, G. Structure determination of inactive-state GPCRs with a universal nanobody. Nat. Struct. Mol. Biol. 2022, 29, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Butler, Y.R.; Liu, Y.; Kumbhar, R.; Zhao, P.; Gadhave, K.; Wang, N.; Li, Y.; Mao, X.; Wang, W. α-Synuclein fibril-specific nanobody reduces prion-like α-synuclein spreading in mice. Nat. Commun. 2022, 13, 4060. [Google Scholar] [CrossRef] [PubMed]
- Fridy, P.C.; Li, Y.; Keegan, S.; Thompson, M.K.; Nudelman, I.; Scheid, J.F.; Oeffinger, M.; Nussenzweig, M.C.; Fenyo, D.; Chait, B.T.; et al. A robust pipeline for rapid production of versatile nanobody repertoires. Nat Methods 2014, 11, 1253–1260. [Google Scholar] [CrossRef] [Green Version]
- Cong, A.T.Q.; Witter, T.L.; Schellenberg, M.J. High-efficiency recombinant protein purification using mCherry and YFP nanobody affinity matrices. Protein Sci. A Publ. Protein Soc. 2022, 31, e4383. [Google Scholar] [CrossRef]
- Wang, Z.; Li, L.; Hu, R.; Zhong, P.; Zhang, Y.; Cheng, S.; Jiang, H.; Liu, R.; Ding, Y. Structural insights into the binding of nanobodies LaM2 and LaM4 to the red fluorescent protein mCherry. Protein Sci. A Publ. Protein Soc. 2021, 30, 2298–2309. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Williams, K.L.; Appel, R.D.; Hochstrasser, D.F. Protein identification and analysis tools in the ExPASy server. Methods Mol. Biol. 1999, 112, 531–552. [Google Scholar] [CrossRef]
- Pletnev, S.; Subach, F.V.; Dauter, Z.; Wlodawer, A.; Verkhusha, V.V. Understanding Blue-to-Red Conversion in Monomeric Fluorescent Timers and Hydrolytic Degradation of Their Chromophores. J. Am. Chem. Soc. 2010, 132, 2243–2253. [Google Scholar] [CrossRef] [Green Version]
- Hattori, M.; Hibbs, R.E.; Gouaux, E. A Fluorescence-Detection Size-Exclusion Chromatography-Based Thermostability Assay for Membrane Protein Precrystallization Screening. Structure 2012, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Jin, F.; Funato, Y.; Xu, Z.; Zhu, W.; Wang, J.; Sun, M.; Zhao, Y.; Yu, Y.; Miki, H.; et al. Structural basis for the Mg2+ recognition and regulation of the CorC Mg2+ transporter. Sci. Adv. 2021, 7, eabe6140. [Google Scholar] [CrossRef]
- Wang, Q.-S.; Zhang, K.-H.; Cui, Y.; Wang, Z.-J.; Pan, Q.-Y.; Liu, K.; Sun, B.; Zhou, H.; Li, M.-J.; Xu, Q.; et al. Upgrade of macromolecular crystallography beamline BL17U1 at SSRF. Nucl. Sci. Tech. 2018, 29, 68. [Google Scholar] [CrossRef]
- Minor, W.; Cymborowski, M.; Otwinowski, Z.; Chruszcz, M. HKL-3000: The integration of data reduction and structure solution—From diffraction images to an initial model in minutes. Acta Cryst. D Biol. Cryst. 2006, 62, 859–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.-W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. Sect. D 2019, 75, 861–877. [Google Scholar] [CrossRef] [Green Version]
- The CCP4 suite: Programs for protein crystallography. Acta Cryst. D Biol. Cryst. 1994, 50, 760–763. [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Cryst. D Biol. Cryst. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 1.8; Schrodinger, LLC: New York, NY, USA, 2015.
- OriginPro, Version 2021; OriginLab Corporation: Northampton, MA, USA, 2021.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| mCherry-LaM1 | mCherry-LaM3 | mCherry-Lam8 | |
|---|---|---|---|
| Wavelength | 0.979 | 0.979 | 0.979 |
| Resolution range | 31.24−2.05 (2.123−2.05) | 66.8−3.29 (3.408−3.29) | 50.36−1.31 (1.357−1.31) |
| Space group | P 1 | C 1 2 1 | P 21 21 21 |
| Unit cell | 68.07 74.83 94.64 111.33 100.68 104.42 | 134.867 104.831 68.5085 90 102.828 90 | 63.0748 75.3103 83.6246 90 90 90 |
| Total reflections | 349,349 (33,627) | 93,208 (7800) | 1,055,598 (41,572) |
| Unique reflections | 97,578 (9741) | 14,164 (1388) | 95,041 (8553) |
| Multiplicity | 3.6 (3.5) | 6.6 (5.6) | 11.1 (4.9) |
| Completeness (%) | 96.90 (96.43) | 99.43 (99.64) | 98.81 (90.40) |
| Mean I/sigma(I) | 8.97 (1.90) | 4.67 (1.59) | 18.48 (1.71) |
| Wilson B-factor | 41.13 | 65.43 | 14.25 |
| R-merge | 0.08522 (1.024) | 0.4263 (1.557) | 0.06234 (0.7762) |
| R-meas | 0.1008 (1.218) | 0.4681 (1.733) | 0.06521 (0.8674) |
| R-pim | 0.05308 (0.6503) | 0.19 (0.7503) | 0.01877 (0.3708) |
| CC1/2 | 0.996 (0.565) | 0.835 (0.472) | 0.999 (0.645) |
| CC* | 0.999 (0.85) | 0.954 (0.801) | 1 (0.886) |
| Reflections used in refinement | 97,558 (9740) | 14,108 (1383) | 95,021 (8548) |
| Reflections used for R-free | 4722 (448) | 682 (90) | 2000 (180) |
| R-work | 0.2043 (0.3369) | 0.2638 (0.3527) | 0.1780 (0.2628) |
| R-free | 0.2350 (0.3622) | 0.3123 (0.4489) | 0.1896 (0.2445) |
| CC (work) | 0.961 (0.806) | 0.897 (0.810) | 0.967 (0.803) |
| CC (free) | 0.946 (0.716) | 0.811 (0.638) | 0.971 (0.763) |
| Number of non-hydrogen atoms | 11,069 | 5076 | 3231 |
| macromolecules | 10,428 | 5030 | 2621 |
| ligands | 114 | 46 | 23 |
| solvent | 527 | 0 | 587 |
| Protein residues | 1353 | 671 | 338 |
| RMS (bonds) | 0.002 | 0.002 | 0.006 |
| RMS (angles) | 0.53 | 0.54 | 0.9 |
| Ramachandran favored (%) | 98.42 | 95.27 | 98.8 |
| Ramachandran allowed (%) | 1.51 | 3.97 | 1.2 |
| Ramachandran outliers (%) | 0.08 | 0.76 | 0 |
| Rotamer outliers (%) | 0.95 | 0.41 | 0 |
| Clashscore | 4.34 | 5.03 | 3.67 |
| Average B-factor | 48.56 | 74.35 | 19.15 |
| macromolecules | 48.44 | 74.38 | 16.2 |
| ligands | 53.5 | 71.52 | 19.52 |
| solvent | 49.87 | 32.29 |
| Antibody | Antigen | KD (M) | kon (1/Ms) | kdis (1/s) | R2 |
|---|---|---|---|---|---|
| LaM1 | mCherry | 1.29 × 10−8 ±2.80 × 10−10 | 9.13 × 104 ±1.76 × 103 | 1.17 × 10−3 ±1.19 × 10−5 | 0.9426 |
| LaM3 | mCherry | 3.06 × 10−8 ±2.94 × 10−10 | 1.06 × 105 ±9.72 × 102 | 3.24 × 10−3 ±9.63 × 10−6 | 0.9896 |
| LaM8 | mCherry | 3.68 × 10−8 ±1.50 × 10−9 | 2.52 × 105 ±9.88 × 103 | 9.26 × 10−3 ±1.03 × 10−4 | 0.9354 |
| LaM3 | DsRed | 7.13 × 10−6 ±1.31 × 10−7 | 62.2 ±1.02 | 4.43 × 10−4 ±3.56 × 10−6 | 0.9838 |
| Group | Temp (°C) | N (Sites) | KD (M) | ∆H (kJ/mol) | ∆G (kJ/mol) | −T∆S (kJ/mol) | ΔS (J/[mol × K]) |
|---|---|---|---|---|---|---|---|
| LaM1 to mCherry | 25 | 1.000 ±1.50 × 10−2 | 2.89 × 10−7 ±5.18 × 10−8 | −120 ±3.79 | −37.4 | 82.9 | −278.19 |
| LaM3 to mCherry | 25.1 | 0.913 ±9.50 × 10−3 | 1.37 × 10−8 ±5.21 × 10−9 | −57.5 ±0.657 | −44.9 | 12.5 | −41.93 |
| LaM8 to mCherry | 25.2 | 0.842 ±4.80 × 10−2 | 1.99 × 10−7 ±1.34 × 10−7 | −46.8 ±6.32 | −38.3 | 8.51 | −28.54 |
| LaM8 to mCherry with LaM1 | 25.1 | 0.854 ±5.40 × 10−3 | 5.40 × 10−8 ±7.68 × 10−9 | −43 ±0.569 | −41.5 | 1.51 | −5.07 |
| LaM8 to mCherry with LaM3 | 25.1 | 0.799 ±3.00 × 10−2 | 2.36 × 10−7 ±1.03 × 10−7 | −41.8 ±2.85 | −37.9 | 3.95 | −13.25 |
| Antibody. | Antigen | KD (M) | KD2 (M) | kon (1/Ms) | kon2 (1/Ms) | kdis1 (1/s) | kdis2 (1/s) | R2 |
|---|---|---|---|---|---|---|---|---|
| LaM1-LaM8 | mCherry | 1.87 × 10−9 | <1.0 × 10−12 | 2.96 × 105 | 4.67 × 104 | 5.52 × 10−4 | <1.0 × 10−7 | 0.986 |
| error | <1.0 × 10−12 | 4.52 × 10−10 | 2.10 × 103 | 1.69 × 103 | 4.63 × 10−6 | − | ||
| LaM8-LaM4 | mCherry | 1.38 × 10−9 | 1.29 × 10−9 | 1.73 × 105 | 5.06 × 104 | 2.39 × 10−4 | 6.51 × 10−5 | 0.990 |
| error | <1.0 × 10−12 | 1.01 × 10−10 | 1.12 × 103 | 1.93 × 103 | 1.89 × 10−6 | 4.47 × 10−6 | ||
| LaM8-LaM4 | DsRed | 2.64 × 10−9 | 4.12 × 10−9 | 8.81 × 104 | 1.99 × 104 | 2.32 × 10−4 | 8.20 × 10−5 | 0.991 |
| error | <1.0 × 10−12 | 3.12 × 10−10 | 5.05 × 102 | 9.92 × 102 | 1.71 × 10−6 | 4.69 × 10−6 |
| Nanobody | Temp (°C) | N (Sites) | KD (M) | ∆H (kJ/mol) | ∆G (kJ/mol) | −T∆S (kJ/mol) | ΔS (J/[mol × K]) |
|---|---|---|---|---|---|---|---|
| LaM1-LaM8 | 20.1 | 0.671 ±1.20 × 10−2 | 4.27 × 10−8 ±1.82 × 10−8 | −144 ±5.00 | −41.4 | 103 | −351.42 |
| LaM8-LaM4 | 20.2 | 0.750 ±3.70 × 10−2 | 3.10 × 10−7 ±1.70 × 10−7 | −60.1 ±6.09 | −36.6 | 23.5 | −80.15 |
| Protein 1 | Nat | Nres | Surface (Å) | Protein 2 | Nat | Nres | Surface (Å) | Interface Area (Å2) | ΔG (kcal/mol) | ΔG (p-Value) | NHB | NSB | NDS | CSS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| LaM1 (8IM1) | 64 | 18 | 6299 | mCherry | 75 | 20 | 9663 | 627.0 | −1.2 | 0.494 | 13 | 3 | 0 | 0.000 |
| LaM2 (6IR2) | 71 | 17 | 6365 | mCherry | 90 | 22 | 9926 | 679.6 | −5.1 | 0.160 | 9 | 1 | 0 | 0.000 |
| LaM3 (8ILX) | 60 | 16 | 6108 | mCherry | 67 | 20 | 9257 | 597.2 | −8.6 | 0.235 | 6 | 0 | 0 | 0.100 |
| LaM4 (6IR1) | 76 | 23 | 6127 | mCherry | 72 | 21 | 10,088 | 757.8 | −7.0 | 0.172 | 6 | 0 | 0 | 0.100 |
| LaM6 (7SAL) | 59 | 15 | 6268 | mCherry | 77 | 21 | 10,081 | 611.0 | −5.2 | 0.257 | 4 | 1 | 0 | 0.000 |
| LaM8 (8IM0) | 66 | 20 | 6295 | mCherry | 75 | 23 | 9879 | 688.8 | 1.5 | 0.734 | 11 | 4 | 0 | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Ma, Z.; Wang, Z.; Zhong, P.; Li, R.; Jiang, H.; Zong, X.; Zhong, C.; Liu, X.; Liu, P.; et al. Structural Insights into the Binding of Red Fluorescent Protein mCherry-Specific Nanobodies. Int. J. Mol. Sci. 2023, 24, 6952. https://doi.org/10.3390/ijms24086952
Liang H, Ma Z, Wang Z, Zhong P, Li R, Jiang H, Zong X, Zhong C, Liu X, Liu P, et al. Structural Insights into the Binding of Red Fluorescent Protein mCherry-Specific Nanobodies. International Journal of Molecular Sciences. 2023; 24(8):6952. https://doi.org/10.3390/ijms24086952
Chicago/Turabian StyleLiang, Hui, Zhiqiang Ma, Ziying Wang, Peiyu Zhong, Ran Li, He Jiang, Xin Zong, Chao Zhong, Xihuan Liu, Peng Liu, and et al. 2023. "Structural Insights into the Binding of Red Fluorescent Protein mCherry-Specific Nanobodies" International Journal of Molecular Sciences 24, no. 8: 6952. https://doi.org/10.3390/ijms24086952