
Inter-Alpha Inhibitor Proteins Modify the Microvasculature after Exposure to Hypoxia–Ischemia and Hypoxia in Neonatal Rats

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
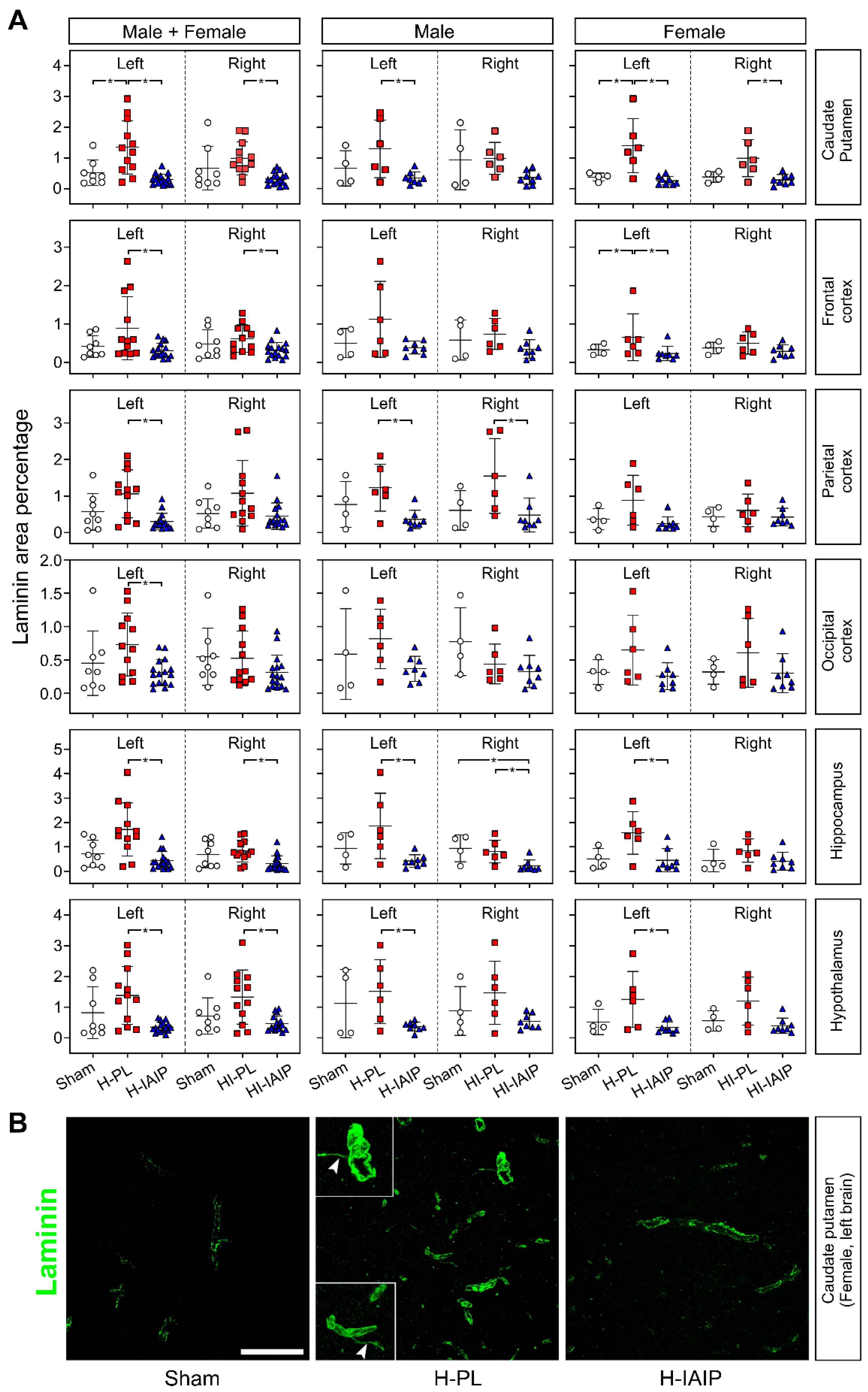
2.1. Effects of Treatment with hIAIPs on the Percent of Laminin-Stained Area in Brain Regions after Exposure of Neonatal Rats to Hypoxia–Ischemia and Systemic Hypoxia
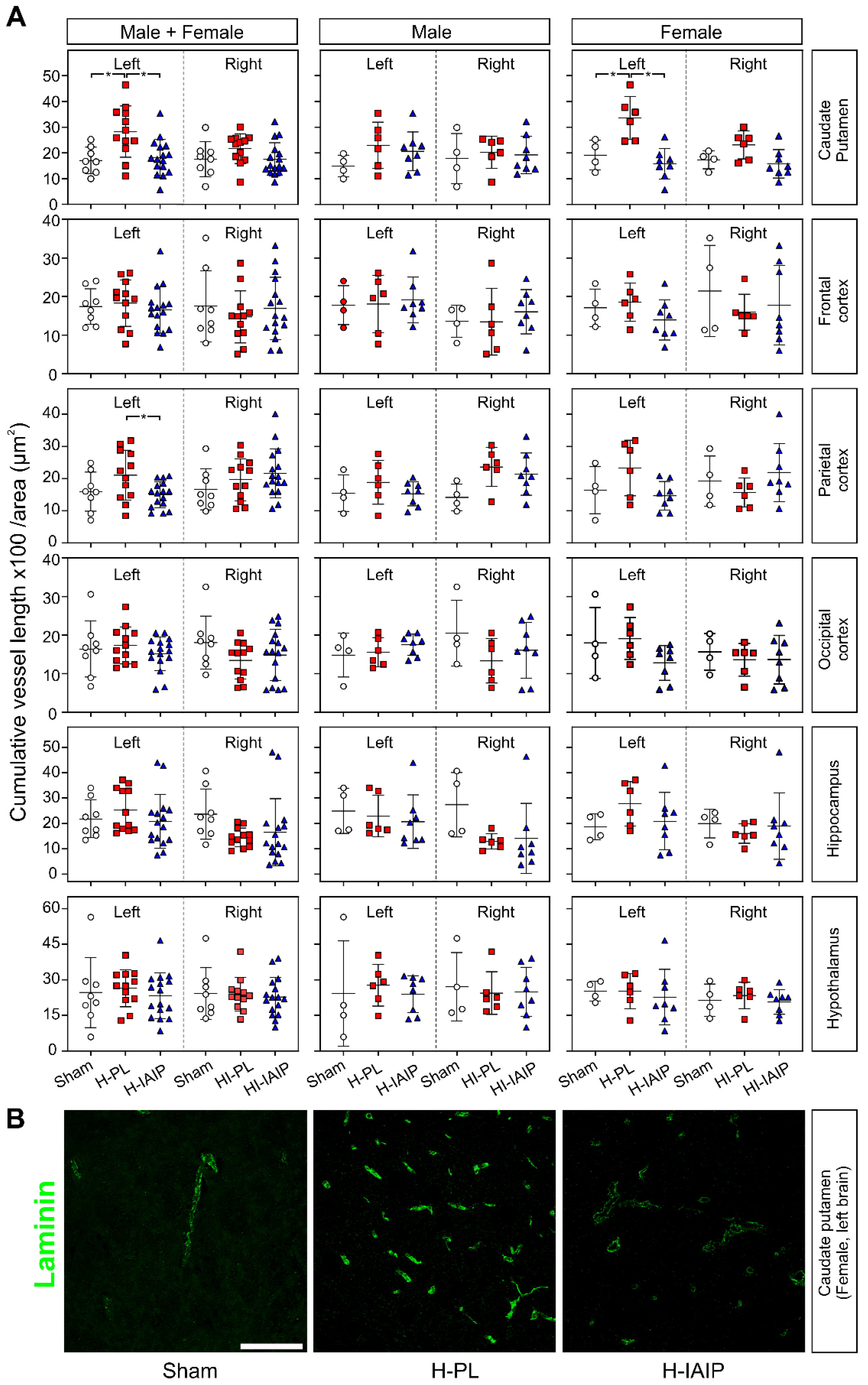
2.2. Treatment with hIAIPs Reduces Cumulative Blood Vessel Length in the Brains of Neonatal Rats after Exposure to Systemic Hypoxia
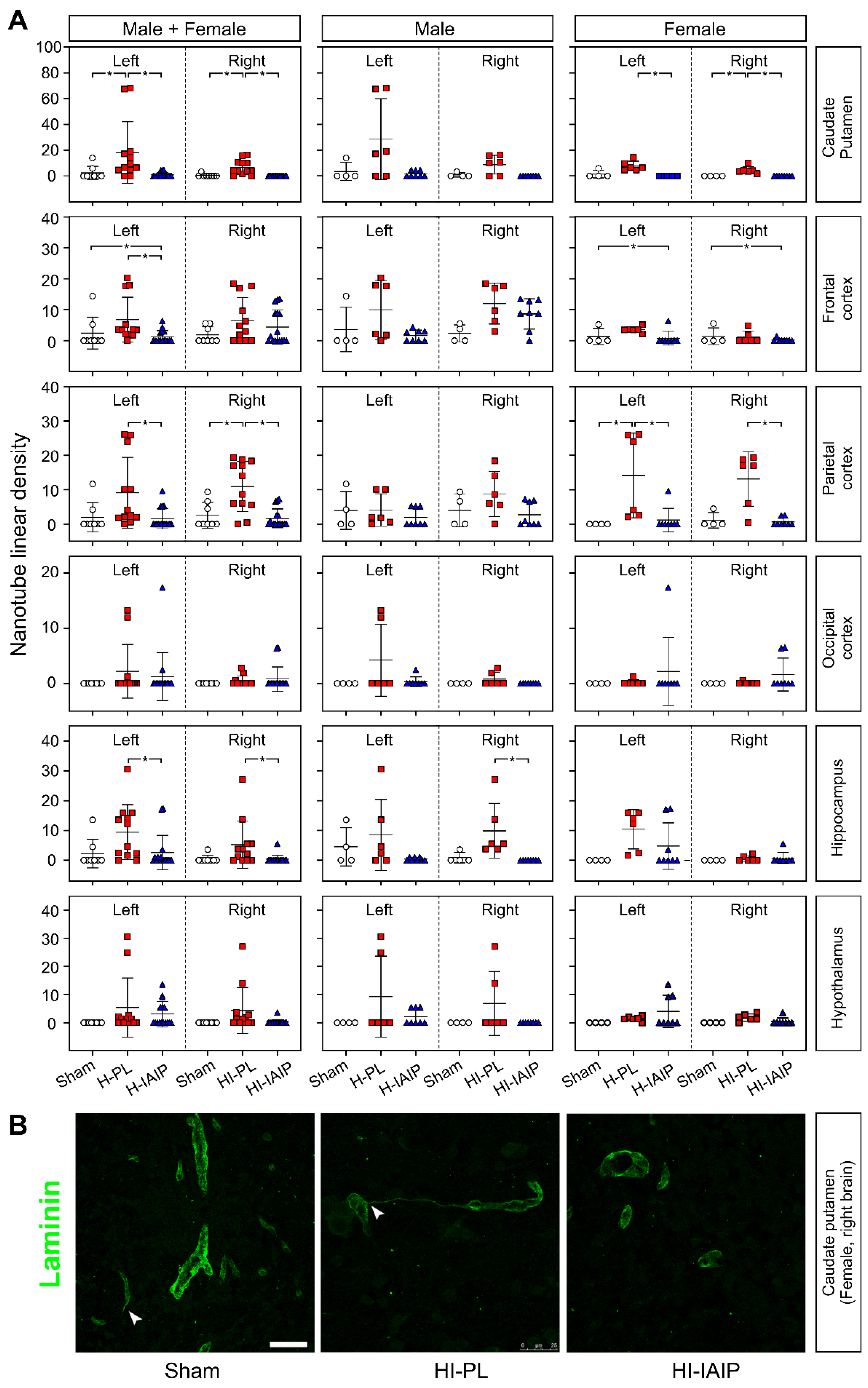
2.3. Treatment with hIAIPs Affects the Linear Density of Tunneling Nanotubes in More Brain Regions of the Female Than Male Neonatal Rats after Exposure to Systemic Hypoxia and Hypoxia–Ischemia
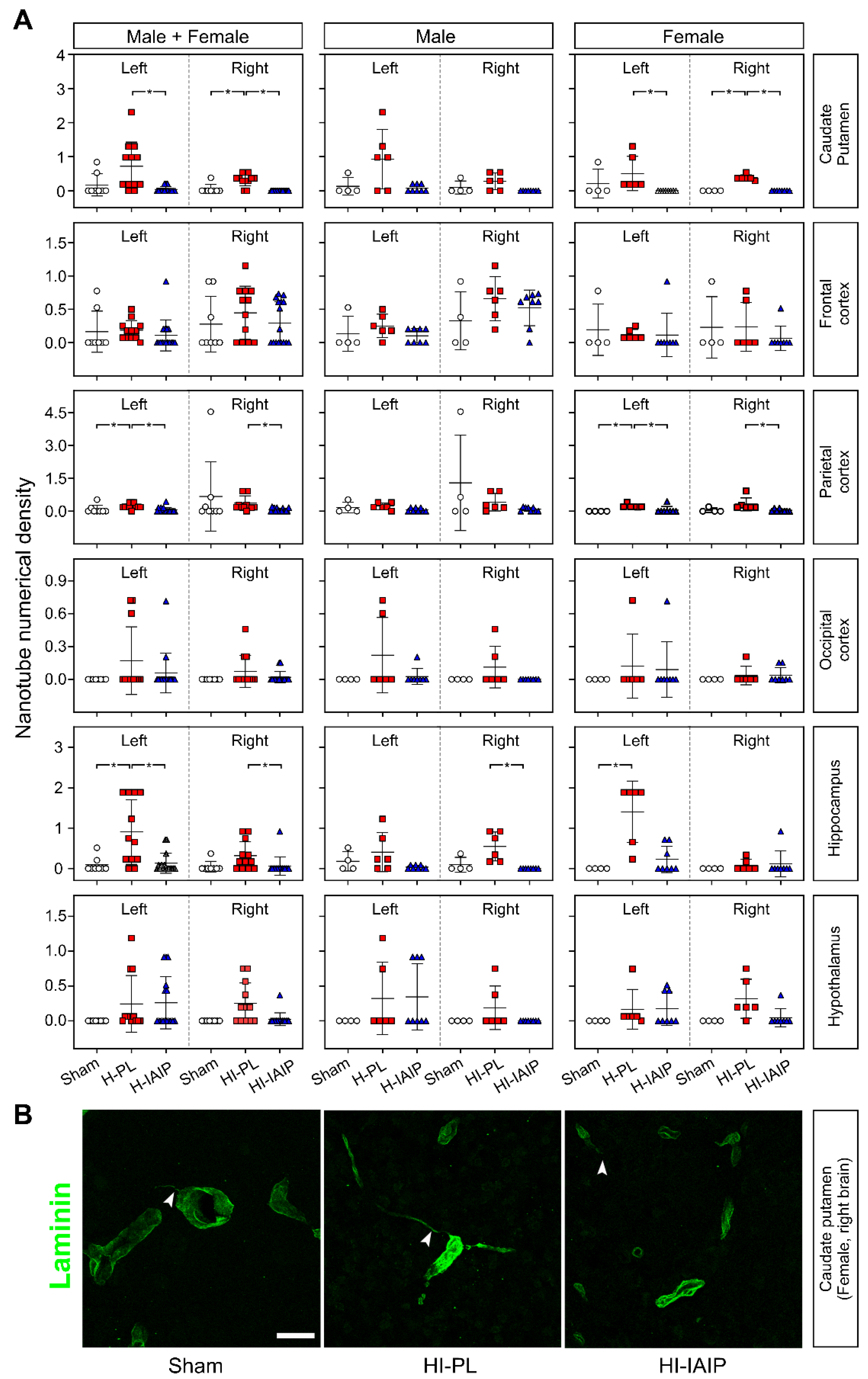
2.4. Treatment with hIAIPs Affects Tunneling Nanotube Numeric Density in More Brain Regions of the Female Compared with the Male Neonatal Rats after Exposure to Systemic Hypoxia and Hypoxia–Ischemia
3. Discussion
4. Materials and Methods
4.1. Production and Purification of Human Inter-Alpha Inhibitor Proteins (hIAIPs)
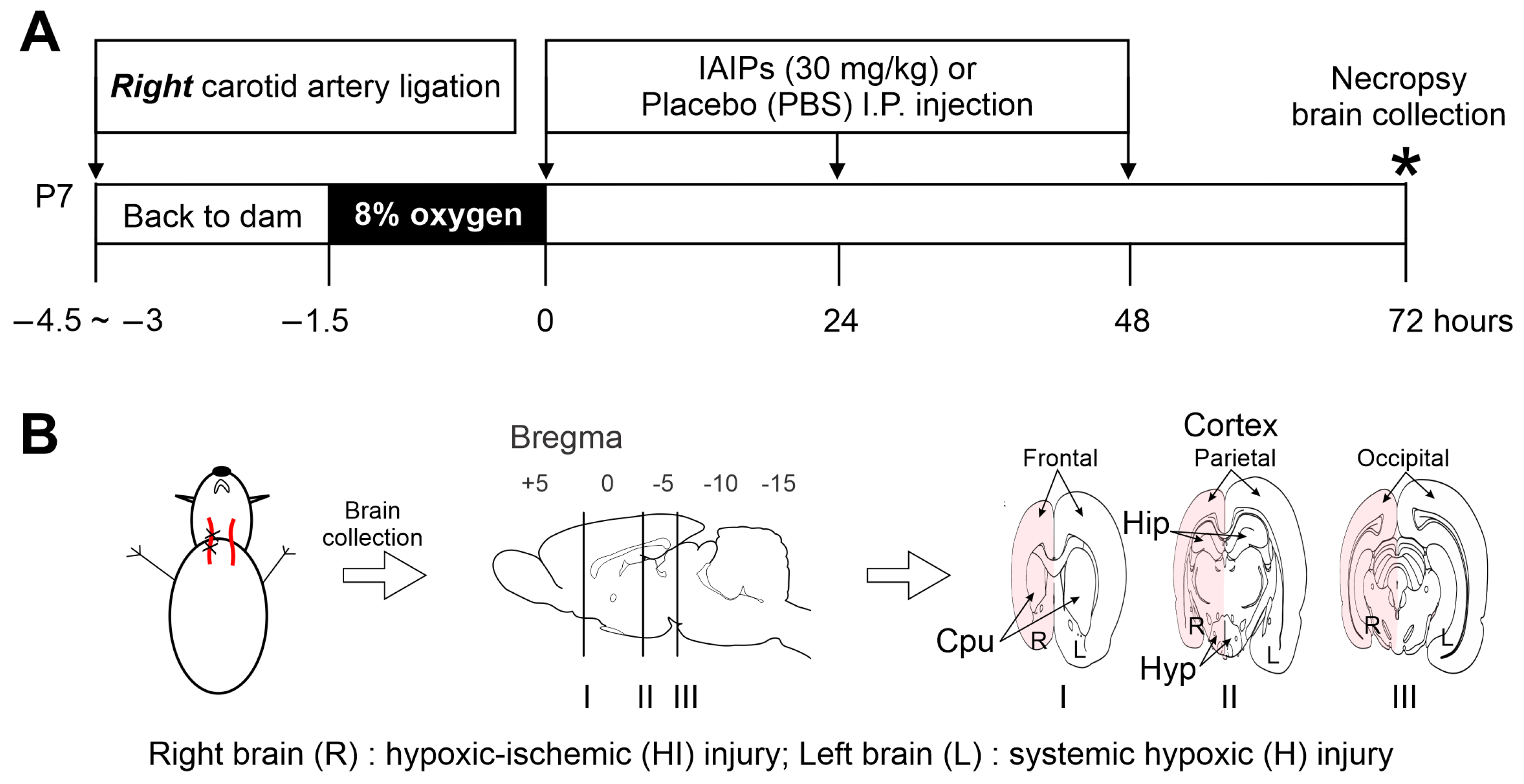
4.2. Experimental Animals and Hypoxic–Ischemic (HI) Procedures
4.3. Histology and Immunofluorescence
4.4. Morphometry and Quantitative Analysis
4.5. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barrett, R.D.; Bennet, L.; Davidson, J.; Dean, J.M.; George, S.; Emerald, B.S.; Gunn, A.J. Destruction and reconstruction: Hypoxia and the developing brain. Birth Defects Res. C Embryo Today 2007, 81, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Fatemi, A.; Wilson, M.A.; Johnston, M.V. Hypoxic-ischemic encephalopathy in the term infant. Clin. Perinatol. 2009, 36, 835–858, vii. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scafidi, J.; Fagel, D.M.; Ment, L.R.; Vaccarino, F.M. Modeling premature brain injury and recovery. Int. J. Dev. Neurosci. 2009, 27, 863–871. [Google Scholar] [CrossRef] [Green Version]
- Baburamani, A.A.; Ek, C.J.; Walker, D.W.; Castillo-Melendez, M. Vulnerability of the developing brain to hypoxic-ischemic damage: Contribution of the cerebral vasculature to injury and repair? Front. Physiol. 2012, 3, 424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centers for Disease, C. Prevention, Economic costs associated with mental retardation, cerebral palsy, hearing loss, and vision impairment--United States, 2003. MMWR Morb. Mortal. Wkly. Rep. 2004, 53, 57–59. [Google Scholar]
- Klahr, A.C.; Nadeau, C.A.; Colbourne, F. Temperature Control in Rodent Neuroprotection Studies: Methods and Challenges. Ther. Hypothermia Temp. Manag. 2017, 7, 42–49. [Google Scholar] [CrossRef]
- Cotten, C.M.; Shankaran, S. Hypothermia for hypoxic-ischemic encephalopathy. Expert Rev. Obstet. Gynecol. 2010, 5, 227–239. [Google Scholar] [CrossRef] [Green Version]
- Osredkar, D.; Thoresen, M.; Maes, E.; Flatebo, T.; Elstad, M.; Sabir, H. Hypothermia is not neuroprotective after infection-sensitized neonatal hypoxic-ischemic brain injury. Resuscitation 2014, 85, 567–572. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Gunn, A.J.; Wyatt, J.S. Hypothermia for neonates with hypoxic-ischemic encephalopathy. N. Engl. J. Med. 2006, 354, 1643–1645. [Google Scholar] [PubMed]
- Gunn, A.J.; Gunn, T.R.; de Haan, H.H.; Williams, C.E.; Gluckman, P.D. Dramatic neuronal rescue with prolonged selective head cooling after ischemia in fetal lambs. J. Clin. Investig. 1997, 99, 248–256. [Google Scholar] [CrossRef]
- Higgins, R.D.; Raju, T.N.; Perlman, J.; Azzopardi, D.V.; Blackmon, L.R.; Clark, R.H.; Edwards, A.D.; Ferriero, D.M.; Gluckman, P.D.; Gunn, A.J.; et al. Hypothermia and perinatal asphyxia: Executive summary of the National Institute of Child Health and Human Development workshop. J. Pediatr. 2006, 148, 170–175. [Google Scholar] [CrossRef]
- Shankaran, S.; Laptook, A.R.; Ehrenkranz, R.A.; Tyson, J.E.; McDonald, S.A.; Donovan, E.F.; Fanaroff, A.A.; Poole, W.K.; Wright, L.L.; Higgins, R.D.; et al. Whole-body hypothermia for neonates with hypoxic-ischemic encephalopathy. N. Engl. J. Med. 2005, 353, 1574–1584. [Google Scholar] [CrossRef]
- Shankaran, S.; Pappas, A.; McDonald, S.A.; Vohr, B.R.; Hintz, S.R.; Yolton, K.; Gustafson, K.E.; Leach, T.M.; Green, C.; Bara, R.; et al. Childhood outcomes after hypothermia for neonatal encephalopathy. N. Engl. J. Med. 2012, 366, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Disdier, C.; Stonestreet, B.S. Hypoxic-ischemic-related cerebrovascular changes and potential therapeutic strategies in the neonatal brain. J. Neurosci. Res. 2020, 98, 1468–1484. [Google Scholar] [CrossRef] [PubMed]
- Dixon, B.J.; Reis, C.; Ho, W.M.; Tang, J.; Zhang, J.H. Neuroprotective Strategies after Neonatal Hypoxic Ischemic Encephalopathy. Int. J. Mol. Sci. 2015, 16, 22368–22401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virgintino, D.; Girolamo, F.; Rizzi, M.; Ahmedli, N.; Sadowska, G.B.; Stopa, E.G.; Zhang, J.; Stonestreet, B.S. Ischemia/Reperfusion-induced neovascularization in the cerebral cortex of the ovine fetus. J. Neuropathol. Exp. Neurol. 2014, 73, 495–506. [Google Scholar] [CrossRef] [Green Version]
- Zaitoun, I.S.; Cikla, U.; Zafer, D.; Udho, E.; Almomani, R.; Suscha, A.; Cengiz, P.; Sorenson, C.M.; Sheibani, N. Attenuation of Retinal Vascular Development in Neonatal Mice Subjected to Hypoxic-Ischemic Encephalopathy. Sci. Rep. 2018, 8, 9166. [Google Scholar] [CrossRef] [Green Version]
- Loveridge-Easther, C.; Good, W.V. Ocular neovascularization following hypoxic ischemic encephalopathy. J. AAPOS 2022, 26, 216–218. [Google Scholar] [CrossRef]
- Threlkeld, S.W.; Gaudet, C.M.; La Rue, M.E.; Dugas, E.; Hill, C.A.; Lim, Y.P.; Stonestreet, B.S. Effects of inter-alpha inhibitor proteins on neonatal brain injury: Age, task and treatment dependent neurobehavioral outcomes. Exp. Neurol. 2014, 261, 424–433. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Nakada, S.; Donahue, J.E.; Chen, R.H.; Tucker, R.; Qiu, J.; Lim, Y.P.; Stopa, E.G.; Stonestreet, B.S. Neuroprotective effects of inter-alpha inhibitor proteins after hypoxic-ischemic brain injury in neonatal rats. Exp. Neurol. 2019, 317, 244–259. [Google Scholar] [CrossRef]
- McCullough, L.D.; Roy-O’Reilly, M.; Lai, Y.-J.; Patrizz, A.; Xu, Y.; Lee, J.; Holmes, A.; Kraushaar, D.C.; Chauhan, A.; Sansing, L.H.; et al. Exogenous inter-α inhibitor proteins prevent cell death and improve ischemic stroke outcomes in mice. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Barrios-Anderson, A.; Chen, X.; Nakada, S.; Chen, R.; Lim, Y.P.; Stonestreet, B.S. Inter-alpha Inhibitor Proteins Modulate Neuroinflammatory Biomarkers After Hypoxia-Ischemia in Neonatal Rats. J. Neuropathol. Exp. Neurol. 2019, 78, 742–755. [Google Scholar] [CrossRef]
- Logsdon, A.F.; Erickson, M.A.; Chen, X.; Qiu, J.; Lim, Y.P.; Stonestreet, B.S.; Banks, W.A. Inter-alpha inhibitor proteins attenuate lipopolysaccharide-induced blood-brain barrier disruption and downregulate circulating interleukin 6 in mice. J. Cereb. Blood Flow Metab. 2020, 40, 1090–1102. [Google Scholar] [CrossRef]
- Schuffels, S.; Nakada, S.; Wu, Y.; Lim, Y.P.; Chen, X.; Stonestreet, B.S. Effects of inter-alpha inhibitor proteins on brain injury after exposure of neonatal rats to severe hypoxia-ischemia. Exp. Neurol. 2020, 334, 113442. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, J.; Wu, Y.; Tucker, R.; Baird, G.L.; Domonoske, R.; Barrios-Anderson, A.; Lim, Y.P.; Bath, K.; Walsh, E.G.; et al. Inter-alpha Inhibitor Proteins Ameliorate Brain Injury and Improve Behavioral Outcomes in a Sex-Dependent Manner After Exposure to Neonatal Hypoxia Ischemia in Newborn and Young Adult Rats. Neurotherapeutics 2022, 19, 528–549. [Google Scholar] [CrossRef]
- Gaudet, C.M.; Lim, Y.P.; Stonestreet, B.S.; Threlkeld, S.W. Effects of age, experience and inter-alpha inhibitor proteins on working memory and neuronal plasticity after neonatal hypoxia-ischemia. Behav. Brain Res. 2016, 302, 88–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Threlkeld, S.W.; Lim, Y.P.; La Rue, M.; Gaudet, C.; Stonestreet, B.S. Immuno-modulator inter-alpha inhibitor proteins ameliorate complex auditory processing deficits in rats with neonatal hypoxic-ischemic brain injury. Brain Behav. Immun. 2017, 64, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Cowan, W.M. The development of the brain. Sci. Am. 1979, 241, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Dobbing, J.; Sands, J. Comparative aspects of the brain growth spurt. Early Hum. Dev. 1979, 3, 79–83. [Google Scholar] [CrossRef]
- Semple, B.D.; Blomgren, K.; Gimlin, K.; Ferriero, D.M.; Noble-Haeusslein, L.J. Brain development in rodents and humans: Identifying benchmarks of maturation and vulnerability to injury across species. Prog. Neurobiol. 2013, 106–107, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, L.M.; Nguyen, K.; Chen, X.; Santoso, A.; Tucker, R.; Lim, Y.P.; Stonestreet, B.S. Effects of Three Different Doses of Inter-Alpha Inhibitor Proteins on Severe Hypoxia-Ischemia-Related Brain Injury in Neonatal Rats. Int. J. Mol. Sci. 2022, 23, 13473. [Google Scholar] [CrossRef]
- Chen, X.; Song, D.; Nakada, S.; Qiu, J.; Iwamoto, K.; Chen, R.H.; Lim, Y.P.; Jusko, W.J.; Stonestreet, B.S. Pharmacokinetics of Inter-Alpha Inhibitor Proteins and Effects on Hemostasis after Hypoxic-Ischemic Brain Injury in Neonatal Rats. Curr. Pharm. Des. 2020, 26, 3997–4006. [Google Scholar] [CrossRef]
- Alarcon-Martinez, L.; Villafranca-Baughman, D.; Quintero, H.; Kacerovsky, J.B.; Dotigny, F.; Murai, K.K.; Prat, A.; Drapeau, P.; Di Polo, A. Interpericyte tunnelling nanotubes regulate neurovascular coupling. Nature 2020, 585, 91–95. [Google Scholar] [CrossRef]
- Errede, M.; Mangieri, D.; Longo, G.; Girolamo, F.; de Trizio, I.; Vimercati, A.; Serio, G.; Frei, K.; Perris, R.; Virgintino, D. Tunneling nanotubes evoke pericyte/endothelial communication during normal and tumoral angiogenesis. Fluids Barriers CNS 2018, 15, 28. [Google Scholar] [CrossRef] [Green Version]
- Hallmann, R.; Hannocks, M.J.; Song, J.; Zhang, X.; Di Russo, J.; Luik, A.L.; Burmeister, M.; Gerwien, H.; Sorokin, L. The role of basement membrane laminins in vascular function. Int. J. Biochem. Cell Biol. 2020, 127, 105823. [Google Scholar] [CrossRef]
- Girolamo, F.; de Trizio, I.; Errede, M.; Longo, G.; d’Amati, A.; Virgintino, D. Neural crest cell-derived pericytes act as pro-angiogenic cells in human neocortex development and gliomas. Fluids Barriers CNS 2021, 18, 14. [Google Scholar] [CrossRef]
- Alarcon-Martinez, L.; Shiga, Y.; Villafranca-Baughman, D.; Belforte, N.; Quintero, H.; Dotigny, F.; Cueva Vargas, J.L.; Di Polo, A. Pericyte dysfunction and loss of interpericyte tunneling nanotubes promote neurovascular deficits in glaucoma. Proc. Natl. Acad. Sci. USA 2022, 119, e2110329119. [Google Scholar] [CrossRef]
- Timpl, R.; Rohde, H.; Robey, P.G.; Rennard, S.I.; Foidart, J.M.; Martin, G.R. Laminin--a glycoprotein from basement membranes. J. Biol. Chem. 1979, 254, 9933–9937. [Google Scholar] [CrossRef] [PubMed]
- Risau, W.; Lemmon, V. Changes in the vascular extracellular matrix during embryonic vasculogenesis and angiogenesis. Dev. Biol. 1988, 125, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Milner, R.; Campbell, I.L. Developmental regulation of beta1 integrins during angiogenesis in the central nervous system. Mol. Cell Neurosci. 2002, 20, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Natah, S.S.; Srinivasan, S.; Pittman, Q.; Zhao, Z.; Dunn, J.F. Effects of acute hypoxia and hyperthermia on the permeability of the blood-brain barrier in adult rats. J. Appl. Physiol. (1985) 2009, 107, 1348–1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Guan, J.; Jiang, Z.; Yang, Y.; Liu, J.; Hua, W.; Mao, Y.; Li, C.; Lu, W.; Qian, J.; et al. Brain-targeted drug delivery by manipulating protein corona functions. Nat. Commun. 2019, 10, 3561. [Google Scholar] [CrossRef] [Green Version]
- Bertossi, M.; Virgintino, D.; Errede, M.; Roncali, L. Immunohistochemical and ultrastructural characterization of cortical plate microvasculature in the human fetus telencephalon. Microvasc. Res. 1999, 58, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Eriksdotter-Nilsson, M.; Bjorklund, H.; Olson, L. Laminin immunohistochemistry: A simple method to visualize and quantitate vascular structures in the mammalian brain. J. Neurosci. Methods 1986, 17, 275–286. [Google Scholar] [CrossRef]
- Hatayama, K.; Riddick, S.; Awa, F.; Chen, X.; Virgintino, D.; Stonestreet, B.S. Time Course of Changes in the Neurovascular Unit after Hypoxic-Ischemic Injury in Neonatal Rats. Int. J. Mol. Sci. 2022, 23, 4180. [Google Scholar] [CrossRef]
- Girolamo, F.; Errede, M.; Longo, G.; Annese, T.; Alias, C.; Ferrara, G.; Morando, S.; Trojano, M.; Kerlero de Rosbo, N.; Uccelli, A.; et al. Defining the role of NG2-expressing cells in experimental models of multiple sclerosis. A biofunctional analysis of the neurovascular unit in wild type and NG2 null mice. PLoS ONE 2019, 14, e0213508. [Google Scholar] [CrossRef] [Green Version]
- Virgintino, D.; Maiorano, E.; Errede, M.; Vimercati, A.; Greco, P.; Selvaggi, L.; Roncali, L.; Bertossi, M. Astroglia-microvessel relationship in the developing human telencephalon. Int. J. Dev. Biol. 1998, 42, 1165–1168. [Google Scholar]
- Zapata-Acevedo, J.F.; Garcia-Perez, V.; Cabezas-Perez, R.; Losada-Barragan, M.; Vargas-Sanchez, K.; Gonzalez-Reyes, R.E. Laminin as a Biomarker of Blood-Brain Barrier Disruption under Neuroinflammation: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 6788. [Google Scholar] [CrossRef]
- Gautam, J.; Xu, L.; Nirwane, A.; Nguyen, B.; Yao, Y. Loss of mural cell-derived laminin aggravates hemorrhagic brain injury. J. Neuroinflam. 2020, 17, 103. [Google Scholar] [CrossRef] [Green Version]
- Gautam, J.; Zhang, X.; Yao, Y. The role of pericytic laminin in blood brain barrier integrity maintenance. Sci. Rep. 2016, 6, 36450. [Google Scholar] [CrossRef] [Green Version]
- Stober, V.P.; Lim, Y.P.; Opal, S.; Zhuo, L.; Kimata, K.; Garantziotis, S. Inter-alpha-inhibitor Ameliorates Endothelial Inflammation in Sepsis. Lung 2019, 197, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Baranova, N.S.; Foulcer, S.J.; Briggs, D.C.; Tilakaratna, V.; Enghild, J.J.; Milner, C.M.; Day, A.J.; Richter, R.P. Inter-alpha-inhibitor impairs TSG-6-induced hyaluronan cross-linking. J. Biol. Chem. 2013, 288, 29642–29653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huth, S.; Huth, L.; Marquardt, Y.; Fietkau, K.; Dahl, E.; Esser, P.R.; Martin, S.F.; Heise, R.; Merk, H.F.; Baron, J.M. Inter-alpha-Trypsin Inhibitor Heavy Chain 5 (ITIH5) Is a Natural Stabilizer of Hyaluronan That Modulates Biological Processes in the Skin. Skin Pharmacol. Physiol. 2020, 33, 198–206. [Google Scholar] [CrossRef]
- Rugg, M.S.; Willis, A.C.; Mukhopadhyay, D.; Hascall, V.C.; Fries, E.; Fulop, C.; Milner, C.M.; Day, A.J. Characterization of complexes formed between TSG-6 and inter-alpha-inhibitor that act as intermediates in the covalent transfer of heavy chains onto hyaluronan. J. Biol. Chem. 2005, 280, 25674–25686. [Google Scholar] [CrossRef] [Green Version]
- He, H.; Li, W.; Tseng, D.Y.; Zhang, S.; Chen, S.Y.; Day, A.J.; Tseng, S.C. Biochemical characterization and function of complexes formed by hyaluronan and the heavy chains of inter-alpha-inhibitor (HC*HA) purified from extracts of human amniotic membrane. J. Biol. Chem. 2009, 284, 20136–20146. [Google Scholar] [CrossRef] [Green Version]
- Shay, E.; He, H.; Sakurai, S.; Tseng, S.C. Inhibition of angiogenesis by HC.HA, a complex of hyaluronan and the heavy chain of inter-alpha-inhibitor, purified from human amniotic membrane. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2669–2678. [Google Scholar] [CrossRef]
- Slevin, M.; Krupinski, J.; Gaffney, J.; Matou, S.; West, D.; Delisser, H.; Savani, R.C.; Kumar, S. Hyaluronan-mediated angiogenesis in vascular disease: Uncovering RHAMM and CD44 receptor signaling pathways. Matrix. Biol. 2007, 26, 58–68. [Google Scholar] [CrossRef]
- Al Ahmad, A.; Gassmann, M.; Ogunshola, O.O. Maintaining blood-brain barrier integrity: Pericytes perform better than astrocytes during prolonged oxygen deprivation. J. Cell Physiol. 2009, 218, 612–622. [Google Scholar] [CrossRef]
- Volpe, J.J. Intraventricular hemorrhage and brain injury in the premature infant. Neuropathology and pathogenesis. Clin. Perinatol. 1989, 16, 361–386. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Mahmood, A.; Chopp, M. Angiogenesis, neurogenesis and brain recovery of function following injury. Curr. Opin. Investig. Drugs 2010, 11, 298–308. [Google Scholar]
- Ogunshola, O.O.; Stewart, W.B.; Mihalcik, V.; Solli, T.; Madri, J.A.; Ment, L.R. Neuronal VEGF expression correlates with angiogenesis in postnatal developing rat brain. Brain Res. Dev. Brain Res. 2000, 119, 139–153. [Google Scholar] [CrossRef] [PubMed]
- Mu, D.; Jiang, X.; Sheldon, R.A.; Fox, C.K.; Hamrick, S.E.; Vexler, Z.S.; Ferriero, D.M. Regulation of hypoxia-inducible factor 1alpha and induction of vascular endothelial growth factor in a rat neonatal stroke model. Neurobiol. Dis. 2003, 14, 524–534. [Google Scholar] [CrossRef] [PubMed]
- Rehncrona, S.; Rosen, I.; Siesjo, B.K. Brain lactic acidosis and ischemic cell damage: 1. Biochemistry and neurophysiology. J. Cereb. Blood Flow Metab. 1981, 1, 297–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baburamani, A.A.; Castillo-Melendez, M.; Walker, D.W. VEGF expression and microvascular responses to severe transient hypoxia in the fetal sheep brain. Pediatr. Res. 2013, 73, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.C.; Chang, Y.C.; Lin, Y.C.; Sze, C.I.; Huang, C.C.; Ho, C.J. Cerebral microvascular damage occurs early after hypoxia-ischemia via nNOS activation in the neonatal brain. J. Cereb. Blood Flow Metab. 2014, 34, 668–676. [Google Scholar] [CrossRef] [Green Version]
- Jain, A.; Kratimenos, P.; Koutroulis, I.; Jain, A.; Buddhavarapu, A.; Ara, J. Effect of Intranasally Delivered rh-VEGF165 on Angiogenesis Following Cerebral Hypoxia-Ischemia in the Cerebral Cortex of Newborn Piglets. Int. J. Mol. Sci. 2017, 18, 2356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dupont, M.; Souriant, S.; Lugo-Villarino, G.; Maridonneau-Parini, I.; Verollet, C. Tunneling Nanotubes: Intimate Communication between Myeloid Cells. Front. Immunol. 2018, 9, 43. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, M.; Abounit, S.; Marzo, L.; Danckaert, A.; Chamoun, Z.; Roux, P.; Zurzolo, C. Transfer of polyglutamine aggregates in neuronal cells occurs in tunneling nanotubes. J. Cell Sci. 2013, 126 Pt 16, 3678–3685. [Google Scholar] [CrossRef] [Green Version]
- Abounit, S.; Zurzolo, C. Wiring through tunneling nanotubes--from electrical signals to organelle transfer. J. Cell Sci. 2012, 125 Pt 5, 1089–1098. [Google Scholar] [CrossRef] [Green Version]
- Tardivel, M.; Begard, S.; Bousset, L.; Dujardin, S.; Coens, A.; Melki, R.; Buee, L.; Colin, M. Tunneling nanotube (TNT)-mediated neuron-to neuron transfer of pathological Tau protein assemblies. Acta Neuropathol. Commun. 2016, 4, 117. [Google Scholar] [CrossRef] [Green Version]
- Vargas, J.Y.; Loria, F.; Wu, Y.J.; Cordova, G.; Nonaka, T.; Bellow, S.; Syan, S.; Hasegawa, M.; van Woerden, G.M.; Trollet, C.; et al. The Wnt/Ca(2+) pathway is involved in interneuronal communication mediated by tunneling nanotubes. EMBO J. 2019, 38, e101230. [Google Scholar] [CrossRef]
- Ariazi, J.; Benowitz, A.; De Biasi, V.; Den Boer, M.L.; Cherqui, S.; Cui, H.; Douillet, N.; Eugenin, E.A.; Favre, D.; Goodman, S.; et al. Tunneling Nanotubes and Gap Junctions-Their Role in Long-Range Intercellular Communication during Development, Health, and Disease Conditions. Front. Mol. Neurosci. 2017, 10, 333. [Google Scholar] [CrossRef] [PubMed]
- Marzo, L.; Gousset, K.; Zurzolo, C. Multifaceted roles of tunneling nanotubes in intercellular communication. Front. Physiol. 2012, 3, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eugenin, E.A.; Gaskill, P.J.; Berman, J.W. Tunneling nanotubes (TNT): A potential mechanism for intercellular HIV trafficking. Commun. Integr. Biol. 2009, 2, 243–244. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, M.H.; Lehmann, J.M.; Erfle, V. Nef-induced CCL2 Expression Contributes to HIV/SIV Brain Invasion and Neuronal Dysfunction. Front. Immunol. 2019, 10, 2447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y. Tunneling-nanotube. Commun. Integr. Biol. 2011, 4, 324–325. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.; Rocha-Ferreira, E.; Fleiss, B.; Nijboer, C.H.; Gressens, P.; Mallard, C.; Hagberg, H. Neuroprotection offered by mesenchymal stem cells in perinatal brain injury: Role of mitochondria, inflammation, and reactive oxygen species. J. Neurochem. 2021, 158, 59–73. [Google Scholar] [CrossRef]
- Pisani, F.; Castagnola, V.; Simone, L.; Loiacono, F.; Svelto, M.; Benfenati, F. Role of pericytes in blood-brain barrier preservation during ischemia through tunneling nanotubes. Cell Death Dis. 2022, 13, 582. [Google Scholar] [CrossRef]
- Dion-Albert, L.; Cadoret, A.; Doney, E.; Kaufmann, F.N.; Dudek, K.A.; Daigle, B.; Parise, L.F.; Cathomas, F.; Samba, N.; Hudson, N.; et al. Vascular and blood-brain barrier-related changes underlie stress responses and resilience in female mice and depression in human tissue. Nat. Commun. 2022, 13, 164. [Google Scholar] [CrossRef]
- Elahy, M.; Jackaman, C.; Mamo, J.C.; Lam, V.; Dhaliwal, S.S.; Giles, C.; Nelson, D.; Takechi, R. Blood-brain barrier dysfunction developed during normal aging is associated with inflammation and loss of tight junctions but not with leukocyte recruitment. Immun. Ageing 2015, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Frias-Anaya, E.; Gromnicova, R.; Kraev, I.; Rogachevsky, V.; Male, D.K.; Crea, F.; Hawkes, C.A.; Romero, I.A. Age-related ultrastructural neurovascular changes in the female mouse cortex and hippocampus. Neurobiol. Aging 2021, 101, 273–284. [Google Scholar] [CrossRef] [PubMed]
- Villapol, S.; Faivre, V.; Joshi, P.; Moretti, R.; Besson, V.C.; Charriaut-Marlangue, C. Early Sex Differences in the Immune-Inflammatory Responses to Neonatal Ischemic Stroke. Int. J. Mol. Sci. 2019, 20, 3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Threlkeld, S.W.; Cummings, E.E.; Juan, I.; Makeyev, O.; Besio, W.G.; Gaitanis, J.; Banks, W.A.; Sadowska, G.B.; Stonestreet, B.S. Ischemia-reperfusion impairs blood-brain barrier function and alters tight junction protein expression in the ovine fetus. Neuroscience 2012, 226, 89–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koehn, L.M.; Chen, X.; Logsdon, A.F.; Lim, Y.P.; Stonestreet, B.S. Novel Neuroprotective Agents to Treat Neonatal Hypoxic-Ischemic Encephalopathy: Inter-Alpha Inhibitor Proteins. Int. J. Mol. Sci. 2020, 21, 9193. [Google Scholar] [CrossRef]
- Singh, K.; Zhang, L.X.; Bendelja, K.; Heath, R.; Murphy, S.; Sharma, S.; Padbury, J.F.; Lim, Y.P. Inter-alpha inhibitor protein administration improves survival from neonatal sepsis in mice. Pediatr. Res. 2010, 68, 242–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opal, S.M.; Lim, Y.P.; Siryaporn, E.; Moldawer, L.L.; Pribble, J.P.; Palardy, J.E.; Souza, S. Longitudinal studies of inter-alpha inhibitor proteins in severely septic patients: A potential clinical marker and mediator of severe sepsis. Crit. Care Med. 2007, 35, 387–392. [Google Scholar] [CrossRef]
- Sadowska, G.B.; Chen, X.; Zhang, J.; Lim, Y.P.; Cummings, E.E.; Makeyev, O.; Besio, W.G.; Gaitanis, J.; Padbury, J.F.; Banks, W.A.; et al. Interleukin-1beta transfer across the blood-brain barrier in the ovine fetus. J. Cereb. Blood Flow Metab. 2015, 35, 1388–1395. [Google Scholar] [CrossRef]
- Chen, X.; Sadowska, G.B.; Zhang, J.; Kim, J.E.; Cummings, E.E.; Bodge, C.A.; Lim, Y.P.; Makeyev, O.; Besio, W.G.; Gaitanis, J.; et al. Neutralizing anti-interleukin-1beta antibodies modulate fetal blood-brain barrier function after ischemia. Neurobiol. Dis. 2015, 73, 118–129. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Sadowska, G.B.; Chen, X.; Park, S.Y.; Kim, J.E.; Bodge, C.A.; Cummings, E.; Lim, Y.P.; Makeyev, O.; Besio, W.G.; et al. Anti-IL-6 neutralizing antibody modulates blood-brain barrier function in the ovine fetus. FASEB J. 2015, 29, 1739–1753. [Google Scholar] [CrossRef] [Green Version]
- Al-Ahmad, A.J.; Patel, R.; Palecek, S.P.; Shusta, E.V. Hyaluronan impairs the barrier integrity of brain microvascular endothelial cells through a CD44-dependent pathway. J. Cereb. Blood Flow Metab. 2019, 39, 1759–1775. [Google Scholar] [CrossRef]
- Mallard, E.C.; Williams, C.E.; Gunn, A.J.; Gunning, M.I.; Gluckman, P.D. Frequent episodes of brief ischemia sensitize the fetal sheep brain to neuronal loss and induce striatal injury. Pediatr. Res. 1993, 33, 61–65. [Google Scholar] [CrossRef] [Green Version]
- Pathak, A.P.; Schmainda, K.M.; Ward, B.D.; Linderman, J.R.; Rebro, K.J.; Greene, A.S. MR-derived cerebral blood volume maps: Issues regarding histological validation and assessment of tumor angiogenesis. Magn. Reson. Med. 2001, 46, 735–747. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.; Lee, E.; Plummer, C.; Gil, S.; Popel, A.S.; Pathak, A.P. Vasculature-specific MRI reveals differential anti-angiogenic effects of a biomimetic peptide in an orthotopic breast cancer model. Angiogenesis 2015, 18, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Cebulla, J.; Ward, B.D.; Rhie, K.; Zhang, J.; Pathak, A.P. Assessing breast cancer angiogenesis in vivo: Which susceptibility contrast MRI biomarkers are relevant? Magn. Reson. Med. 2013, 70, 1106–1116. [Google Scholar] [CrossRef]
- Choi, J.P.; Foley, M.; Zhou, Z.; Wong, W.Y.; Gokoolparsadh, N.; Arthur, J.S.; Li, D.Y.; Zheng, X. Micro-CT Imaging Reveals Mekk3 Heterozygosity Prevents Cerebral Cavernous Malformations in Ccm2-Deficient Mice. PLoS ONE 2016, 11, e0160833. [Google Scholar] [CrossRef] [Green Version]
- Menezes, K.; Nascimento, M.A.; Goncalves, J.P.; Cruz, A.S.; Lopes, D.V.; Curzio, B.; Bonamino, M.; de Menezes, J.R.; Borojevic, R.; Rossi, M.I.; et al. Human mesenchymal cells from adipose tissue deposit laminin and promote regeneration of injured spinal cord in rats. PLoS ONE 2014, 9, e96020. [Google Scholar] [CrossRef]
- Malaeb, S.; Dammann, O. Fetal inflammatory response and brain injury in the preterm newborn. J. Child Neurol. 2009, 24, 1119–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segovia, K.N.; McClure, M.; Moravec, M.; Luo, N.L.; Wan, Y.; Gong, X.; Riddle, A.; Craig, A.; Struve, J.; Sherman, L.S.; et al. Arrested oligodendrocyte lineage maturation in chronic perinatal white matter injury. Ann. Neurol. 2008, 63, 520–530. [Google Scholar] [CrossRef] [Green Version]
- Lim, D.K.; Wylie, R.G.; Langer, R.; Kohane, D.S. Selective binding of C-6 OH sulfated hyaluronic acid to the angiogenic isoform of VEGF(165). Biomaterials 2016, 77, 130–138. [Google Scholar] [CrossRef] [Green Version]
- Gonul, E.; Duz, B.; Kahraman, S.; Kayali, H.; Kubar, A.; Timurkaynak, E. Early pericyte response to brain hypoxia in cats: An ultrastructural study. Microvasc. Res. 2002, 64, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Kaczkurkin, A.N.; Raznahan, A.; Satterthwaite, T.D. Sex differences in the developing brain: Insights from multimodal neuroimaging. Neuropsychopharmacology 2019, 44, 71–85. [Google Scholar] [CrossRef]
- Lim, Y.P. ProThera Biologics, Inc.: A novel immunomodulator and biomarker for life-threatening diseases. RI Med. J. 2013, 96, 16–18. [Google Scholar]
- Opal, S.M.; Lim, Y.P.; Cristofaro, P.; Artenstein, A.W.; Kessimian, N.; Delsesto, D.; Parejo, N.; Palardy, J.E.; Siryaporn, E. Inter-alpha inhibitor proteins: A novel therapeutic strategy for experimental anthrax infection. Shock 2011, 35, 42–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, Y.P.; Josic, D.; Callanan, H.; Brown, J.; Hixson, D.C. Affinity purification and enzymatic cleavage of inter-alpha inhibitor proteins using antibody and elastase immobilized on CIM monolithic disks. J. Chromatogr. A 2005, 1065, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Rice, J.E., III; Vannucci, R.C.; Brierley, J.B. The influence of immaturity on hypoxic-ischemic brain damage in the rat. Ann. Neurol. 1981, 9, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Vannucci, S.J.; Hagberg, H. Hypoxia-ischemia in the immature brain. J. Exp. Biol. 2004, 207 Pt 18, 3149–3154. [Google Scholar] [CrossRef] [Green Version]
- Towfighi, J.; Mauger, D.; Vannucci, R.C.; Vannucci, S.J. Influence of age on the cerebral lesions in an immature rat model of cerebral hypoxia-ischemia: A light microscopic study. Brain Res. Dev. Brain Res. 1997, 100, 149–160. [Google Scholar] [CrossRef]
- Thoresen, M.; Hobbs, C.E.; Wood, T.; Chakkarapani, E.; Dingley, J. Cooling combined with immediate or delayed xenon inhalation provides equivalent long-term neuroprotection after neonatal hypoxia-ischemia. J. Cereb. Blood Flow Metab. 2009, 29, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Thoresen, M.; Bagenholm, R.; Loberg, E.M.; Apriccna, F. The stress of being restrained reduces brain damage after a hypoxic-ischaemic insult in the 7-day-old rat. Neuroreport 1996, 7, 481–484. [Google Scholar] [CrossRef] [PubMed]
- Dingley, J.; Tooley, J.; Porter, H.; Thoresen, M. Xenon provides short-term neuroprotection in neonatal rats when administered after hypoxia-ischemia. Stroke 2006, 37, 501–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoresen, M.; Bagenholm, R.; Loberg, E.M.; Apricena, F.; Kjellmer, I. Posthypoxic cooling of neonatal rats provides protection against brain injury. Arch. Dis. Child.-Fetal Neonatal Ed. 1996, 74, F3–F9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Compact; Elsevier Science: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Chen, X.; Zhang, J.; Kim, B.; Jaitpal, S.; Meng, S.S.; Adjepong, K.; Imamura, S.; Wake, H.; Nishibori, M.; Stopa, E.G.; et al. High-mobility group box-1 translocation and release after hypoxic ischemic brain injury in neonatal rats. Exp. Neurol. 2019, 311, 1–14. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analysis | Brain Regions | HI-IAIP Vs. Placebo | |||
|---|---|---|---|---|---|
| Male + Female | Male | Female | |||
| Laminin area percentage | Caudate Putamen | Left | ↓ | ↓ | ↓ |
| Right | ↓ | ↔ | ↓ | ||
| Frontal cortex | Left | ↓ | ↔ | ↓ | |
| Right | ↓ | ↔ | |||
| Parietal cortex | Left | ↓ | ↓ | ↔ | |
| Right | ↔ | ↓ | ↔ | ||
| Occipital cortex | Left | ↓ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Hippocampus | Left | ↓ | ↓ | ↓ | |
| Right | ↓ | ↓ | ↔ | ||
| Hypothalamus | Left | ↓ | ↓ | ↓ | |
| Right | ↓ | ↔ | ↔ | ||
| Cumulative vessel length ×100/area (µm2) | Caudate Putamen | Left | ↓ | ↔ | ↓ |
| Right | ↔ | ↔ | ↔ | ||
| Frontal cortex | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Parietal cortex | Left | ↓ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Occipital cortex | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Hippocampus | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Hypothalamus | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Nanotube linear density | Caudate Putamen | Left | ↓ | ↔ | ↓ |
| Right | ↓ | ↔ | ↓ | ||
| Frontal cortex | Left | ↓ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Parietal cortex | Left | ↓ | ↔ | ↓ | |
| Right | ↓ | ↔ | ↓ | ||
| Occipital cortex | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Hippocampus | Left | ↓ | ↔ | ↔ | |
| Right | ↓ | ↓ | ↔ | ||
| Hypothalamus | Left | ↔ | ↔ | ↔ | |
| Right | v | ↔ | ↔ | ||
| Nanotube Numerical density | Caudate Putamen | Left | ↓ | ↔ | ↓ |
| Right | ↓ | ↔ | ↓ | ||
| Frontal cortex | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Parietal cortex | Left | ↓ | ↔ | ↓ | |
| Right | ↓ | ↔ | ↓ | ||
| Occipital cortex | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
| Hippocampus | Left | ↓ | ↔ | ↔ | |
| Right | ↓ | ↓ | ↔ | ||
| Hypothalamus | Left | ↔ | ↔ | ↔ | |
| Right | ↔ | ↔ | ↔ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Girolamo, F.; Lim, Y.-P.; Virgintino, D.; Stonestreet, B.S.; Chen, X.F. Inter-Alpha Inhibitor Proteins Modify the Microvasculature after Exposure to Hypoxia–Ischemia and Hypoxia in Neonatal Rats. Int. J. Mol. Sci. 2023, 24, 6743. https://doi.org/10.3390/ijms24076743
Girolamo F, Lim Y-P, Virgintino D, Stonestreet BS, Chen XF. Inter-Alpha Inhibitor Proteins Modify the Microvasculature after Exposure to Hypoxia–Ischemia and Hypoxia in Neonatal Rats. International Journal of Molecular Sciences. 2023; 24(7):6743. https://doi.org/10.3390/ijms24076743
Chicago/Turabian StyleGirolamo, Francesco, Yow-Pin Lim, Daniela Virgintino, Barbara S. Stonestreet, and Xiaodi F. Chen. 2023. "Inter-Alpha Inhibitor Proteins Modify the Microvasculature after Exposure to Hypoxia–Ischemia and Hypoxia in Neonatal Rats" International Journal of Molecular Sciences 24, no. 7: 6743. https://doi.org/10.3390/ijms24076743