
Unveiling the Differences in Signaling and Regulatory Mechanisms between Dopamine D2 and D3 Receptors and Their Impact on Behavioral Sensitization
Abstract
:1. Introduction
2. Desensitization and Intracellular Trafficking
2.1. Desensitization
2.2. Endocytosis
2.3. Roles of Small G Proteins in the Signaling and Endocytosis of D2R and D3R
2.3.1. RalA
2.3.2. ARF6
2.4. Roles of Regulators of G Protein Signaling (RGS) in the Signaling of D2R and D3R
3. Biased Signaling
4. Dimerization of Dopamine D2 and D3 Receptors
5. Posttranslational Modifications
5.1. Glycosylation
5.2. Phosphorylation
5.3. Ubiquitination
5.4. Palmitoylation
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, T.; Seeman, P.; Rajput, A.; Farley, I.J.; Hornykiewicz, O. Receptor basis for dopaminergic supersensitivity in Parkinson’s disease. Nature 1978, 273, 59–61. [Google Scholar] [CrossRef]
- Seeman, P. Dopamine receptors and the dopamine hypothesis of schizophrenia. Synapse 1987, 1, 133–152. [Google Scholar] [CrossRef] [PubMed]
- Cunnah, D.; Besser, M. Management of prolactinomas. Clin. Endocrinol. 1991, 34, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H.; Makman, M.H. Stimulation by dopamine of adenylate cyclase in retinal homogenates and of adeno-sine-3′:5′-cyclic monophosphate formation in intact retina. Proc. Natl. Acad. Sci. USA 1972, 69, 539–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Camilli, P.; Macconi, D.; Spada, A. Dopamine inhibits adenylate cyclase in human prolactin-secreting pituitary adenomas. Nature 1979, 278, 252–254. [Google Scholar] [CrossRef]
- Kebabian, J.W.; Calne, D.B. Multiple receptors for dopamine. Nature 1979, 277, 93–96. [Google Scholar] [CrossRef]
- Dearry, A.; Gingrich, J.A.; Falardeau, P.; Bates, M.D.; Caron, M.G. Molecular cloning and expression of the gene for a human D1 dopamine receptor. Nature 1990, 347, 72–76. [Google Scholar] [CrossRef]
- Sunahara, R.K.; Niznik, H.B.; Weiner, D.M.; Stormann, T.M.; Brann, M.R.; Kennedy, J.L.; Gelernter, J.E.; Rozmahel, R.; Yang, Y.; Israel, Y.; et al. Human dopamine D1 receptor encoded by an intronless gene on chromosome 5. Nature 1990, 347, 80–83. [Google Scholar] [CrossRef]
- Sunahara, R.K.; Guan, H.-C.; O’Dowd, B.F.; Seeman, P.; Laurier, L.G.; Ng, G.; George, S.R.; Torchia, J.; Van Tol, H.H.M.; Niznik, H.B. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D1. Nature 1991, 350, 614–619. [Google Scholar] [CrossRef]
- Bunzow, J.R.; Van Tol, H.H.M.; Grandy, D.K.; Albert, P.; Salon, J.; Christie, M.; Machida, C.A.; Neve, K.A.; Civelli, O. Cloning and expression of a rat D2 dopamine receptor cDNA. Nature 1988, 336, 783–787. [Google Scholar] [CrossRef]
- Sokoloff, P.; Giros, B.; Martres, M.P.; Bouthenet, M.L.; Schwartz, J.C. Molecular cloning and characterization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 1990, 347, 146–151. [Google Scholar] [CrossRef]
- Van Tol, H.H.M.; Bunzow, J.R.; Guan, H.-C.; Sunahara, R.K.; Seeman, P.; Niznik, H.B.; Civelli, O. Cloning of the gene for a human dopamine D4 receptor with high affinity for the antipsychotic clozapine. Nature 1991, 350, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Giros, B.; Martres, M.P.; Sokoloff, P.; Schwartz, J.C. Gene cloning of human dopaminergic D3 receptor and identification of its chromosome. C R Acad. Sci. III 1990, 311, 501–508. [Google Scholar] [PubMed]
- Chio, C.L.; Lajiness, E.M.; Huff, R.M. Activation of heterologously expressed D3 dopamine receptors: Comparison with D2 dopamine receptors. Mol. Pharmacol. 1994, 45, 51–60. [Google Scholar] [PubMed]
- Freedman, S.B.; Patel, S.; Marwood, R.; Emms, F.; Seabrook, G.R.; Knowles, M.R.; McAllister, G. Expression and pharmaco-logical characterization of the human D3 dopamine receptor. J. Pharmacol. Exp. Ther. 1994, 268, 417–426. [Google Scholar]
- Robinson, S.W.; Caron, M.G. Selective Inhibition of Adenylyl Cyclase Type V by the Dopamine D3Receptor. Mol. Pharmacol. 1997, 52, 508–514. [Google Scholar] [CrossRef]
- Ganz, M.B.; Pachter, A.J.; Barber, D.L. Multiple receptors coupled to adenylate cyclase regulate Na-H exchange independent of cAMP. J. Biol. Chem. 1990, 265, 8989–8992. [Google Scholar] [CrossRef]
- Neve, K.A.; Kozlowski, M.R.; Rosser, M.P. Dopamine D2 receptor stimulation of Na+/H+ exchange assessed by quantifica-tion of extracellular acidification. J. Biol. Chem. 1992, 267, 25748–25753. [Google Scholar] [CrossRef]
- Pilon, C.; Levesque, D.; Dimitriadou, V.; Griffon, N.; Martres, M.P.; Schwartz, J.C.; Sokoloff, P. Functional coupling of the hu-man dopamine D3 receptor in a transfected NG 108-15 neuroblastoma-glioma hybrid cell line. Eur. J. Pharmacol. 1994, 268, 129–139. [Google Scholar] [CrossRef]
- Welsh, G.I.; Hall, D.A.; Warnes, A.; Strange, P.G.; Proud, C.G. Activation of microtubule-associated protein kinase (Erk) and p70 S6 kinase by D2 dopamine receptors. J. Neurochem. 2002, 70, 2139–2146. [Google Scholar] [CrossRef] [Green Version]
- Cussac, D.; Newman-Tancredi, A.; Pasteau, V.; Millan, M.J. Human Dopamine D3Receptors Mediate Mitogen-Activated Protein Kinase Activation via a Phosphatidylinositol 3-Kinase and an Atypical Protein Kinase C-Dependent Mechanism. Mol. Pharmacol. 1999, 56, 1025–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, L.; Todd, R.D.; O’Malley, K.L. Dopamine D2 and D3 receptors inhibit dopamine release. Experiment 1994, 270, 475–479. [Google Scholar]
- Werner, P.I.A.; Hussy, N.; Buell, G.; Jones, K.A.; North, R.A. D2, D3, and D4 dopamine receptors couple to G protein-regulated potassium channels in Xenopus oocytes. Mol. Pharmacol. 1996, 49, 656–661. [Google Scholar] [PubMed]
- Kuzhikandathil, E.V.; Yu, W.; Oxford, G.S. Human dopamine D3 and D2L receptors couple to inward rectifier potassium channels in mammalian cell lines. Mol. Cell. Neurosci. 1998, 12, 390–402. [Google Scholar] [CrossRef]
- Bouthenet, M.-L.; Souil, E.; Martres, M.-P.; Sokoloff, P.; Giros, B.; Schwartz, J.-C. Localization of dopamine D3 receptor mRNA in the rat brain using in situ hybridization histochemistry: Comparison with dopamine D2 receptor mRNA. Brain Res. 1991, 564, 203–219. [Google Scholar] [CrossRef]
- Robinson, S.W.; Jarvie, K.R.; Caron, M.G. High affinity agonist binding to the dopamine D3 receptor: Chimeric receptors delineate a role for intracellular domains. Mol. Pharmacol. 1994, 46, 352–356. [Google Scholar]
- Joyce, J.N. Dopamine D3 receptor as a therapeutic target for antipsychotic and antiparkinsonian drugs. Pharmacol. Ther. 2001, 90, 231–259. [Google Scholar] [CrossRef]
- Joyce, J.N.; Millan, M.J. Dopamine D3 receptor antagonists as therapeutic agents. Drug Discov. Today 2005, 10, 917–925. [Google Scholar] [CrossRef]
- Newman, A.H.; Grundt, P.; Nader, M.A. Dopamine D3 Receptor Partial Agonists and Antagonists as Potential Drug Abuse Therapeutic Agents. J. Med. Chem. 2005, 48, 3663–3679. [Google Scholar] [CrossRef]
- Kim, K.M.; Valenzano, K.J.; Robinson, S.R.; Yao, W.D.; Barak, L.S.; Caron, M.G. Differential regulation of the dopamine D2 and D3 receptors by G protein-coupled receptor kinases and beta-arrestins. J. Biol. Chem. 2001, 276, 37409–37414. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Le, H.T.; Zhang, X.; Zheng, M.; Choi, B.-G.; Kim, K.-M. Palmitoylation on the carboxyl terminus tail is required for the selective regulation of dopamine D2 versus D3 receptors. Biochim. Biophys. Acta (BBA) Biomembr. 2016, 1858, 2152–2162. [Google Scholar] [CrossRef] [PubMed]
- Min, X.; Sun, N.; Wang, S.; Zhang, X.; Kim, K.M. Sequestration of Gbetagamma by deubiquitinated arrestins into the nucleus as a novel desensitization mechanism of G protein-coupled receptors. Cell Commun. Signal. 2023, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Boeckler, F.; Gmeiner, P. The structural evolution of dopamine D3 receptor ligands: Structure–activity relationships and selected neuropharmacological aspects. Pharmacol. Ther. 2006, 112, 281–333. [Google Scholar] [CrossRef] [PubMed]
- Leggio, G.M.; Bucolo, C.; Platania, C.B.M.; Salomone, S.; Drago, F. Current drug treatments targeting dopamine D3 receptor. Pharmacol. Ther. 2016, 165, 164–177. [Google Scholar] [CrossRef]
- McCorvy, J.D.; Butler, K.V.; Kelly, B.; Rechsteiner, K.; Karpiak, J.; Betz, R.M.; Kormos, B.L.; Shoichet, B.K.; Dror, R.O.; Jin, J.; et al. Structure-inspired design of beta-arrestin-biased ligands for aminergic GPCRs. Nat. Chem. Biol. 2018, 14, 126–134. [Google Scholar] [CrossRef]
- Kiss, B.; Laszlovszky, I.; Kramos, B.; Visegrady, A.; Bobok, A.; Levay, G.; Lendvai, B.; Roman, V. Neuronal Dopamine D3 Re-ceptors: Translational Implications for Preclinical Research and CNS Disorders. Biomolecules 2021, 11, 104. [Google Scholar] [CrossRef]
- Sokoloff, P.; Le Foll, B. A Historical Perspective on the Dopamine D3 Receptor. Curr. Top Behav. Neurosci. 2022, 1–28. [Google Scholar] [CrossRef]
- Gainetdinov, R.R.; Premont, R.T.; Bohn, L.M.; Lefkowitz, R.J.; Caron, M.G. Desensitization of g Protein–Coupled Receptors and Neuronal Functions. Annu. Rev. Neurosci. 2004, 27, 107–144. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.T.; Ingram, S.L.; Henderson, G.; Chavkin, C.; von Zastrow, M.; Schulz, S.; Koch, T.; Evans, C.J.; Christie, M.J. Regula-tion of mu-opioid receptors: Desensitization, phosphorylation, internalization, and tolerance. Pharmacol. Rev. 2013, 65, 223–254. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Kim, K.-M. Mechanistic diversity involved in the desensitization of G protein-coupled receptors. Arch. Pharmacal Res. 2021, 44, 342–353. [Google Scholar] [CrossRef]
- Nambi, P.; Peters, J.R.; Sibley, D.R.; Lefkowitz, R.J. Desensitization of the turkey erythrocyte beta-adrenergic receptor in a cell-free system. Evidence that multiple protein kinases can phosphorylate and desensitize the receptor. J. Biol. Chem. 1985, 260, 2165–2171. [Google Scholar] [CrossRef] [PubMed]
- Benovic, J.L.; Pike, L.J.; Cerione, R.A.; Staniszewski, C.; Yoshimasa, T.; Codina, J.; Caron, M.G.; Lefkowitz, R.J. Phosphoryla-tion of the mammalian beta-adrenergic receptor by cyclic AMP-dependent protein kinase. Regulation of the rate of receptor phos-phorylation and dephosphorylation by agonist occupancy and effects on coupling of the receptor to the stimulatory guanine nucleo-tide regulatory protein. J. Biol. Chem. 1985, 260, 7094–7101. [Google Scholar] [PubMed]
- Cho, E.-Y.; Cho, D.-I.; Park, J.H.; Kurose, H.; Caron, M.G.; Kim, K.-M. Roles of Protein Kinase C and Actin-Binding Protein 280 in the Regulation of Intracellular Trafficking of Dopamine D3 Receptor. Mol. Endocrinol. 2007, 21, 2242–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrens, D.L.; Altenbach, C.; Yang, K.; Hubbell, W.L.; Khorana, H.G. Requirement of Rigid-Body Motion of Transmembrane Helices for Light Activation of Rhodopsin. Science 1996, 274, 768–770. [Google Scholar] [CrossRef] [PubMed]
- Scheerer, P.; Park, J.H.; Hildebrand, P.W.; Kim, Y.J.; Krauß, N.; Choe, H.-W.; Hofmann, K.P.; Ernst, O.P. Crystal structure of opsin in its G-protein-interacting conformation. Nature 2008, 455, 497–502. [Google Scholar] [CrossRef]
- Rasmussen, S.G.F.; DeVree, B.T.; Zou, Y.; Kruse, A.C.; Chung, K.Y.; Kobilka, T.S.; Thian, F.S.; Chae, P.S.; Pardon, E.; Calinski, D.; et al. Crystal structure of the β2 adrenergic receptor–Gs protein complex. Nature 2011, 477, 549–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepek, M.; Beyrière, F.; Hofmann, K.P.; Elgeti, M.; Kazmin, R.; Rose, A.; Bartl, F.J.; von Stetten, D.; Heck, M.; Sommer, M.E.; et al. Crystal structure of a common GPCR-binding interface for G protein and arrestin. Nat. Commun. 2014, 5, 4801. [Google Scholar] [CrossRef] [Green Version]
- Kumari, P.; Srivastava, A.; Banerjee, R.; Ghosh, E.; Gupta, P.; Ranjan, R.; Chen, X.; Gupta, B.; Gupta, C.; Jaiman, D.; et al. Functional competence of a partially engaged GPCR-beta-arrestin complex. Nat. Commun. 2016, 7, 13416. [Google Scholar] [CrossRef] [Green Version]
- Hausdorff, W.P.; Caron, M.G.; Lefkowitz, R.J. Turning off the signal: Desensitization of beta-adrenergic receptor function. FASEB J. 1990, 4, 2881–2889. [Google Scholar] [CrossRef]
- Westrich, L.; Kuzhikandathil, E.V. The tolerance property of human D3 dopamine receptor is determined by specific ami-no acid residues in the second cytoplasmic loop. Biochim. Biophys. Acta 2007, 1773, 1747–1758. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.; Zheng, M.; Min, C.; Ma, L.; Kurose, H.; Park, J.H.; Kim, K.M. Agonist-induced endocytosis and receptor phosphoryla-tion mediate resensitization of dopamine D(2) receptors. Mol. Endocrinol. 2010, 24, 574–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, C.; Zheng, M.; Zhang, X.; Caron, M.G.; Kim, K.M. Novel roles for beta-arrestins in the regulation of pharmacological sequestration to predict agonist-induced desensitization of dopamine D3 receptors. Br. J. Pharmacol. 2013, 170, 1112–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Namkung, Y.; Dipace, C.; Javitch, J.; Sibley, D.R. G Protein-coupled Receptor Kinase-mediated Phosphorylation Regulates Post-endocytic Trafficking of the D2 Dopamine Receptor. J. Biol. Chem. 2009, 284, 15038–15051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, C.; Zhang, X.; Zheng, M.; Sun, N.; Acharya, S.; Zhang, X.; Kim, K.-M. Molecular Signature That Determines the Acute Tolerance of G Protein-Coupled Receptors. Biomol. Ther. 2017, 25, 239–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Zhang, X.; Min, X.; Sun, N.; Kim, K.-M. Cytoplasmic recruitment of Mdm2 as a common characteristic of G pro-tein-coupled receptors that undergo desensitization. Biochem. Biophys. Res. Commun. 2020, 530, 181–188. [Google Scholar] [CrossRef]
- Namkung, Y.; Sibley, D.R. Protein Kinase C Mediates Phosphorylation, Desensitization, and Trafficking of the D2 Dopamine Receptor. J. Biol. Chem. 2004, 279, 49533–49541. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.I.; Min, C.; Jung, K.S.; Cheong, S.Y.; Zheng, M.; Cheong, S.J.; Oak, M.H.; Cheong, J.H.; Lee, B.K.; Kim, K.-M. The N-terminal region of the dopamine D2 receptor, a rhodopsin-like GPCR, regulates correct integration into the plasma membrane and endocytic routes. Br. J. Pharmacol. 2012, 166, 659–675. [Google Scholar] [CrossRef] [Green Version]
- Min, C.; Zheng, M.; Zhang, X.; Guo, S.; Kwon, K.J.; Shin, C.Y.; Kim, H.S.; Cheon, S.H.; Kim, K.M. N-linked Glycosylation on the N-terminus of the dopamine D2 and D3 receptors determines receptor association with specific microdomains in the plasma membrane. Biochim. Biophys. Acta 2015, 1853, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Sun, N.; Zheng, M.; Kim, K.-M. Clathrin-mediated endocytosis is responsible for the lysosomal degradation of dopamine D3 receptor. Biochem. Biophys. Res. Commun. 2016, 476, 245–251. [Google Scholar] [CrossRef]
- Zhang, X.; Kim, K.-M. Multifactorial Regulation of G Protein-Coupled Receptor Endocytosis. Biomol. Ther. 2017, 25, 26–43. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.S.; Downey, W.E.; Colapietro, A.M., 3rd; Barak, L.S.; Menard, L.; Caron, M.G. Role of beta-arrestin in mediating agonist-promoted G protein-coupled receptor internalization. Science 1996, 271, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, S.S.; Caron, M.G. G protein-coupled receptor adaptation mechanisms. Semin. Cell Dev. Biol. 1998, 9, 119–127. [Google Scholar] [CrossRef]
- Hinshaw, J.E.; Schmid, S.L. Dynamin self-assembles into rings suggesting a mechanism for coated vesicle budding. Nature 1995, 374, 190–192. [Google Scholar] [CrossRef] [PubMed]
- Laporte, S.A.; Oakley, R.H.; Holt, J.A.; Barak, L.S.; Caron, M.G. The interaction of beta-arrestin with the AP-2 adaptor is re-quired for the clustering of beta 2-adrenergic receptor into clathrin-coated pits. J. Biol. Chem. 2000, 275, 23120–23126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oakley, R.H.; Laporte, S.A.; Holt, J.A.; Caron, M.G.; Barak, L.S. Differential affinities of visual arrestin, beta arrestin1, and beta arrestin2 for G protein-coupled receptors delineate two major classes of receptors. J. Biol. Chem. 2000, 275, 17201–17210. [Google Scholar] [CrossRef] [Green Version]
- Oakley, R.H.; Laporte, S.A.; Holt, J.A.; Barak, L.S.; Caron, M.G. Molecular determinants underlying the formation of stable intracellular G protein-coupled receptor-beta-arrestin complexes after receptor endocytosis. J. Biol. Chem. 2001, 276, 19452–19460. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.I.; Zheng, M.; Min, C.; Kwon, K.J.; Shin, C.Y.; Choi, H.K.; Kim, K.M. ARF6 and GASP-1 are post-endocytic sorting pro-teins selectively involved in the intracellular trafficking of dopamine D(2) receptors mediated by GRK and PKC in transfected cells. Br. J. Pharmacol. 2013, 168, 1355–1374. [Google Scholar] [CrossRef] [Green Version]
- Mostafapour, S.; Kobilka, B.K.; von Zastrow, M. Pharmacological sequestration of a chimeric beta 3/beta 2 adrenergic re-ceptor occurs without a corresponding amount of receptor internalization. Recept. Signal Transduct. 1996, 6, 151–163. [Google Scholar]
- Burgoyne, R.D. Small GTP-binding proteins. Trends Biochem. Sci. 1989, 14, 394–396. [Google Scholar] [CrossRef]
- Takai, Y.; Sasaki, T.; Matozaki, T. Small GTP-binding proteins. Physiol. Rev. 2001, 81, 153–208. [Google Scholar] [CrossRef]
- Segev, N. GTPases in intracellular trafficking: An overview. Semin. Cell Dev. Biol. 2011, 22, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenmark, H.; Olkkonen, V.M. The Rab GTPase family. Genome Biol. 2001, 2, REVIEWS3007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, T.; Ferguson, C.; Wainwright, B.; Parton, R.; Wicking, C. Rab23, a Negative Regulator of Hedgehog Signaling, Localizes to the Plasma Membrane and the Endocytic Pathway. Traffic 2003, 4, 869–884. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.; Zhang, X.; Guo, S.; Zhang, X.; Min, C.; Cheon, S.H.; Oak, M.-H.; Kim, Y.R.; Kim, K.-M. Agonist-induced changes in RalA activities allows the prediction of the endocytosis of G protein-coupled receptors. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2016, 1863, 77–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Zhang, X.; Sun, N.; Min, C.; Zhang, X.; Kim, K.M. RalA employs GRK2 and beta-arrestins for the filamin A-mediated regulation of trafficking and signaling of dopamine D and D receptor. Biochim. Biophys. Acta 2016, 1863, 2072–2083. [Google Scholar] [CrossRef]
- Iwata, K.; Ito, K.; Fukuzaki, A.; Inaki, K.; Haga, T. Dynamin and rab5 regulate GRK2-dependent internalization of dopamine D2 receptors. JBIC J. Biol. Inorg. Chem. 1999, 263, 596–602. [Google Scholar] [CrossRef] [Green Version]
- Lin, R.; Karpa, K.; Kabbani, N.; Goldman-Rakic, P.; Levenson, R. Dopamine D2 and D3 receptors are linked to the actin cy-toskeleton via interaction with filamin A. Proc. Natl. Acad. Sci. USA 2001, 98, 5258–5263. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Bermak, J.C.; Wang, Z.W.; Zhou, Q.Y. Modulation of Dopamine D2Receptor Signaling by Actin-Binding Protein (ABP-280). Mol. Pharmacol. 2000, 57, 446–452. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, C.; Weingarten, P.; Bunzow, J.R.; Grandy, D.K.; Zhou, Q.Y. Association of dopamine D(3) receptors with ac-tin-binding protein 280 (ABP-280). Biochem. Pharmacol. 2002, 63, 859–863. [Google Scholar] [CrossRef]
- Kim, K.M.; Gainetdinov, R.R.; Laporte, S.A.; Caron, M.G.; Barak, L.S. G protein-coupled receptor kinase regulates dopamine D3 receptor signaling by modulating the stability of a receptor-filamin-beta-arrestin complex. A case of autoreceptor regulation. J. Biol. Chem. 2005, 280, 12774–12780. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishna, H.; Donaldson, J.G. ADP-Ribosylation Factor 6 Regulates a Novel Plasma Membrane Recycling Pathway. J. Cell Biol. 1997, 139, 49–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagnac, G.; de Forges, H.; Smythe, E.; Gueudry, C.; Romao, M.; Salamero, J.; Chavrier, P. Decoupling of Activation and Effector Binding Underlies ARF6 Priming of Fast Endocytic Recycling. Curr. Biol. 2011, 21, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claing, A.; Chen, W.; Miller, W.E.; Vitale, N.; Moss, J.; Premont, R.T.; Lefkowitz, R.J. beta-Arrestin-mediated ADP-ribosylation factor 6 activation and beta 2-adrenergic receptor endocytosis. J. Biol. Chem. 2001, 276, 42509–42513. [Google Scholar] [CrossRef] [Green Version]
- Gilman, A.G. G proteins: Transducers of receptor-generated signals. Annu. Rev. Biochem. 1987, 56, 615–649. [Google Scholar] [CrossRef] [PubMed]
- Angleson, J.K.; Wensel, T.G. A GTPase-accelerating factor for transducin, distinct from its effector cGMP phosphodiester-ase, in rod outer segment membranes. Neuron 1993, 11, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Arshavsky, V.Y.; Pugh, E.N. Lifetime Regulation of G Protein–Effector Complex: Emerging Importance of RGS Proteins. Neuron 1998, 20, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Ross, E.M.; Wilkie, T.M. GTPase-Activating Proteins for Heterotrimeric G Proteins: Regulators of G Protein Signaling (RGS) and RGS-Like Proteins. Annu. Rev. Biochem. 2000, 69, 795–827. [Google Scholar] [CrossRef] [Green Version]
- Taymans, J.-M.; Kia, H.K.; Claes, R.; Cruz, C.; Leysen, J.; Langlois, X. Dopamine receptor-mediated regulation of RGS2 and RGS4 mRNA differentially depends on ascending dopamine projections and time. Eur. J. Neurosci. 2004, 19, 2249–2260. [Google Scholar] [CrossRef]
- Taymans, J.-M.; Leysen, J.E.; Langlois, X. Striatal gene expression of RGS2 and RGS4 is specifically mediated by dopamine D1 and D2 receptors: Clues for RGS2 and RGS4 functions. J. Neurochem. 2003, 84, 1118–1127. [Google Scholar] [CrossRef]
- Rorabaugh, B.R.; Rose, M.J.; Stoops, T.S.; Stevens, A.A.; Seeley, S.L.; D’Souza, M.S. Regulators of G-protein signaling 2 and 4 differentially regulate cocaine-induced rewarding effects. Physiol. Behav. 2018, 195, 9–19. [Google Scholar] [CrossRef]
- Rahman, Z.; Schwarz, J.; Gold, S.J.; Zachariou, V.; Wein, M.N.; Choi, K.-H.; Kovoor, A.; Chen, C.-K.; DiLeone, R.; Schwarz, S.C.; et al. RGS9 Modulates Dopamine Signaling in the Basal Ganglia. Neuron 2003, 38, 941–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Vera, T.M.; Hernandez, S.; Earls, L.R.; Medkova, M.; Sundgren-Andersson, A.K.; Surmeier, D.J.; Hamm, H.E. RGS9-2 modulates D 2 dopamine receptor-mediated Ca 2+ channel inhibition in rat striatal cholinergic interneurons. Proc. Natl. Acad. Sci. USA 2004, 101, 16339–16344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovoor, A.; Seyffarth, P.; Ebert, J.; Barghshoon, S.; Chen, C.K.; Schwarz, S.; Axelrod, J.D.; Cheyette, B.N.; Simon, M.I.; Lester, H.A.; et al. D2 dopamine receptors colocalize regulator of G-protein signaling 9-2 (RGS9-2) via the RGS9 DEP domain, and RGS9 knock-out mice develop dyskinesias associated with dopamine pathways. J. Neurosci. 2005, 25, 2157–2165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, A.; Katz, A.; Simon, M. A fifth member of the mammalian G-protein beta-subunit family. Expression in brain and activation of the beta 2 isotype of phospholipase C. J. Biol. Chem. 1994, 269, 22150–22156. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Hamm, H.E. DEP Domains: More Than Just Membrane Anchors. Dev. Cell 2006, 11, 436–438. [Google Scholar] [CrossRef] [Green Version]
- Cheever, M.L.; Snyder, J.T.; Gershburg, S.; Siderovski, D.P.; Harden, T.K.; Sondek, J. Crystal structure of the multifunctional Gbeta5-RGS9 complex. Nat. Struct. Mol. Biol. 2008, 15, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Masuho, I.; Wakasugi-Masuho, H.; Posokhova, E.N.; Patton, J.R.; Martemyanov, K.A. Type 5 G protein beta subunit (Gbeta5) controls the interaction of regulator of G protein signaling 9 (RGS9) with membrane anchors. J. Biol. Chem. 2011, 286, 21806–21813. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Cheong, S.Y.; Min, C.; Jin, M.; Cho, D.I.; Kim, K.M. Beta-arrestin2 plays permissive roles in the inhibitory activities of RGS9-2 on G protein-coupled receptors by maintaining RGS9-2 in the open conformation. Mol. Cell Biol. 2011, 31, 4887–4901. [Google Scholar] [CrossRef] [Green Version]
- Benovic, J.L.; Kuhn, H.; Weyand, I.; Codina, J.; Caron, M.G.; Lefkowitz, R.J. Functional desensitization of the isolated be-ta-adrenergic receptor by the beta-adrenergic receptor kinase: Potential role of an analog of the retinal protein arrestin (48-kDa pro-tein). Proc. Natl. Acad. Sci. USA 1987, 84, 8879–8882. [Google Scholar] [CrossRef] [Green Version]
- Lohse, M.J.; Benovic, J.L.; Codina, J.; Caron, M.G.; Lefkowitz, R.J. Beta-Arrestin: A protein that regulates beta-adrenergic re-ceptor function. Science 1990, 248, 1547–1550. [Google Scholar] [CrossRef]
- Wei, H.; Ahn, S.; Shenoy, S.K.; Karnik, S.S.; Hunyady, L.; Luttrell, L.M.; Lefkowitz, R.J. Independent beta-arrestin 2 and G pro-tein-mediated pathways for angiotensin II activation of extracellular signal-regulated kinases 1 and 2. Proc. Natl. Acad. Sci. USA 2003, 100, 10782–10787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, J.D.; Clarke, W.P.; von Zastrow, M.; Nichols, D.E.; Kobilka, B.; Weinstein, H.; Javitch, J.A.; Roth, B.L.; Christopoulos, A.; Sexton, P.M.; et al. Functional Selectivity and Classical Concepts of Quantitative Pharmacology. Experiment 2006, 320, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Violin, J.D.; DeWire, S.M.; Yamashita, D.; Rominger, D.H.; Nguyen, L.; Schiller, K.; Whalen, E.J.; Gowen, M.; Lark, M.W. Selec-tively engaging beta-arrestins at the angiotensin II type 1 receptor reduces blood pressure and increases cardiac performance. J. Pharmacol. Exp. Ther. 2010, 335, 572–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, E.; Ahn, S.; Shukla, A.K.; Lefkowitz, R.J. Molecular mechanism of beta-arrestin-biased agonism at sev-en-transmembrane receptors. Annu. Rev. Pharmacol. Toxicol. 2012, 52, 179–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevich, V.V.; Gurevich, E.V. Biased GPCR signaling: Possible mechanisms and inherent limitations. Pharmacol. Ther. 2020, 211, 107540. [Google Scholar] [CrossRef] [PubMed]
- Soergel, D.G.; Subach, R.A.; Burnham, N.; Lark, M.W.; James, I.E.; Sadler, B.M.; Skobieranda, F.; Violin, J.D.; Webster, L.R. Bi-ased agonism of the mu-opioid receptor by TRV130 increases analgesia and reduces on-target adverse effects versus morphine: A randomized, double-blind, placebo-controlled, crossover study in healthy volunteers. Pain 2014, 155, 1829–1835. [Google Scholar] [CrossRef]
- Violin, J.D.; Crombie, A.L.; Soergel, D.G.; Lark, M.W. Biased ligands at G-protein-coupled receptors: Promise and progress. Trends Pharmacol. Sci. 2014, 35, 308–316. [Google Scholar] [CrossRef]
- Sanchez-Soto, M.; Verma, R.K.; Willette, K.A.B.; Gonye, E.C.; Moore, A.M.; Moritz, A.E.; Boateng, C.A.; Yano, H.; Free, R.B.; Shi, L.; et al. A structural basis for how ligand binding site changes can allosterically regulate GPCR signaling and engender func-tional selectivity. Sci. Signal. 2020, 13, eaaw5885. [Google Scholar] [CrossRef]
- Donthamsetti, P.; Gallo, E.F.; Buck, D.C.; Stahl, E.L.; Zhu, Y.; Lane, J.R.; Bohn, L.M.; Neve, K.A.; Kellendonk, C.; Javitch, J.A. Ar-restin recruitment to dopamine D2 receptor mediates locomotion but not incentive motivation. Mol. Psychiatry 2020, 25, 2086–2100. [Google Scholar] [CrossRef]
- Lan, H.; Liu, Y.; Bell, M.I.; Gurevich, V.V.; Neve, K.A. A dopamine D2 receptor mutant capable of G protein-mediated signaling but deficient in arrestin binding. Mol. Pharmacol. 2009, 75, 113–123. [Google Scholar] [CrossRef]
- Peterson, S.M.; Pack, T.F.; Wilkins, A.D.; Urs, N.M.; Urban, D.J.; Bass, C.E.; Lichtarge, O.; Caron, M.G. Elucidation of G-protein and beta-arrestin functional selectivity at the dopamine D2 receptor. Proc. Natl. Acad. Sci. USA 2015, 112, 7097–7102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballesteros, J.A.; Weinstein, H. Integrated methods for the construction of three-dimensional models and computational probing of structure-function relations in G protein-coupled receptors. Methods Neurosci. 1995, 25, 366–428. [Google Scholar]
- Kim, K.M.; Caron, M.G. Complementary roles of the DRY motif and C-terminus tail of GPCRS for G protein coupling and beta-arrestin interaction. Biochem. Biophys. Res. Commun. 2008, 366, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Cho, E.-Y.; Min, C.; Park, J.H.; Kim, K.-M. Characterization of functional roles of DRY motif in the 2nd intracellular loop of dopamine D2 and D3 receptors. Arch. Pharmacal Res. 2008, 31, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Roth, C.B.; Hanson, M.A.; Stevens, R.C. Stabilization of the human beta2-adrenergic receptor TM4-TM3-TM5 helix interface by mutagenesis of Glu122(3.41), a critical residue in GPCR structure. J. Mol. Biol. 2008, 376, 1305–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtarge, O.; Bourne, H.R.; Cohen, F.E. An Evolutionary Trace Method Defines Binding Surfaces Common to Protein Families. J. Mol. Biol. 1996, 257, 342–358. [Google Scholar] [CrossRef] [Green Version]
- Peterson, S.M.; Pack, T.F.; Caron, M.G. Receptor, Ligand and Transducer Contributions to Dopamine D2 Receptor Functional Selectivity. PLoS ONE 2015, 10, e0141637. [Google Scholar] [CrossRef]
- Kroeze, W.K.; Sassano, M.F.; Huang, X.-P.; Lansu, K.; McCorvy, J.D.; Giguère, P.M.; Sciaky, N.; Roth, B.L. PRESTO-Tango as an open-source resource for interrogation of the druggable human GPCRome. Nat. Struct. Mol. Biol. 2015, 22, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Wang, X.; Tocker, A.M.; Huang, P.; Reith, M.E.; Liu-Chen, L.Y.; Smith, A.B.; Kortagere, S., 3rd. Functional Characterization of a Novel Series of Biased Signaling Dopamine D3 Receptor Agonists. ACS Chem. Neurosci. 2017, 8, 486–500. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Bearoff, F.; Kortagere, S. G-protein biased signaling agonists of Dopamine D3 receptor promote distinct activation patterns of ERK1/2. Pharmacol. Res. Off. J. Ital. Pharmacol. Soc. 2022, 179, 106223. [Google Scholar] [CrossRef]
- Quan, W.; Kim, J.H.; Albert, P.R.; Choi, H.; Kim, K.M. Roles of G protein and beta-arrestin in dopamine D2 receptor-mediated ERK activation. Biochem. Biophys. Res. Commun. 2008, 377, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; McCorvy, J.D.; Fischer, M.G.; Butler, K.V.; Shen, Y.; Roth, B.L.; Jin, J. Discovery of G Protein-Biased D2 Dopamine Receptor Partial Agonists. J. Med. Chem. 2016, 59, 10601–10618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonifazi, A.; Yano, H.; Guerrero, A.M.; Kumar, V.; Hoffman, A.F.; Lupica, C.R.; Shi, L.; Newman, A.H. Novel and Potent Do-pamine D(2) Receptor Go-Protein Biased Agonists. ACS Pharmacol. Transl. Sci. 2019, 2, 52–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, E.Y.; Liu, W.; Zhao, Q.; Katritch, V.; Han, G.; Hanson, W.M.; Shi, A.; Newman, L.A.; Javitch, H.J.; Cherezov, A.V.; et al. Structure of the human dopamine D3 receptor in complex with a D2/D3 selective antagonist. Science 2010, 330, 1091–1095. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Che, T.; Levit, A.; Shoichet, B.K.; Wacker, D.; Roth, B.L. Structure of the D2 dopamine receptor bound to the atypical antipsychotic drug risperidone. Nature 2018, 555, 269–273. [Google Scholar] [CrossRef]
- Yin, J.; Chen, K.-Y.M.; Clark, M.J.; Hijazi, M.; Kumari, P.; Bai, X.-C.; Sunahara, R.K.; Barth, P.; Rosenbaum, D.M. Structure of a D2 dopamine receptor–G-protein complex in a lipid membrane. Nature 2020, 584, 125–129. [Google Scholar] [CrossRef]
- Xu, P.; Huang, S.; Mao, C.; Krumm, B.E.; Zhou, X.E.; Tan, Y.; Huang, X.P.; Liu, Y.; Shen, D.D.; Jiang, Y.; et al. Structures of the human dopamine D3 receptor-G(i) complexes. Mol. Cell. 2021, 81, 1147–1159.e4. [Google Scholar] [CrossRef]
- Platania, C.B.M.; Salomone, S.; Leggio, G.M.; Drago, F.; Bucolo, C. Homology Modeling of Dopamine D2 and D3 Receptors: Molecular Dynamics Refinement and Docking Evaluation. PLoS ONE 2012, 7, e44316. [Google Scholar] [CrossRef]
- Lane, J.R.; Abramyan, A.M.; Adhikari, P.; Keen, A.C.; Lee, K.H.; Sanchez, J.; Verma, R.K.; Lim, H.D.; Yano, H.; Javitch, J.A.; et al. Distinct inactive conformations of the dopamine D2 and D3 receptors correspond to different extents of inverse agonism. Elife 2020, 9, e52189. [Google Scholar] [CrossRef]
- Lee, S.P.; So, C.H.; Rashid, A.J.; Varghese, G.; Cheng, R.; Lança, A.J.; O’Dowd, B.F.; George, S.R. Dopamine D1 and D2 Receptor Co-activation Generates a Novel Phospholipase C-mediated Calcium Signal. J. Biol. Chem. 2004, 279, 35671–35678. [Google Scholar] [CrossRef] [Green Version]
- Rashid, A.J.; So, C.H.; Kong, M.M.; Furtak, T.; El-Ghundi, M.; Cheng, R.; O’Dowd, B.F.; George, S.R. D1-D2 dopamine receptor heterooligomers with unique pharmacology are coupled to rapid activation of Gq/11 in the striatum. Proc. Natl. Acad. Sci. USA 2007, 104, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasbi, A.; Fan, T.; Alijaniaram, M.; Nguyen, T.; Perreault, M.L.; O’Dowd, B.F.; George, S.R. Calcium signaling cascade links dopamine D1–D2 receptor heteromer to striatal BDNF production and neuronal growth. Proc. Natl. Acad. Sci. USA 2009, 106, 21377–21382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dowd, B.F.; Ji, X.; Nguyen, T.; George, S.R. Two amino acids in each of D1 and D2 dopamine receptor cytoplasmic regions are involved in D1–D2 heteromer formation. Biochem. Biophys. Res. Commun. 2012, 417, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Łukasiewicz, S.; Faron-Górecka, A.; Dobrucki, J.; Polit, A.; Dziedzicka-Wasylewska, M. Studies on the role of the receptor protein motifs possibly involved in electrostatic interactions on the dopamine D1 and D2 receptor oligomerization. FEBS J. 2008, 276, 760–775. [Google Scholar] [CrossRef] [PubMed]
- Łukasiewicz, S.; Polit, A.; Kędracka-Krok, S.; Wędzony, K.; Maćkowiak, M.; Dziedzicka-Wasylewska, M. Hetero-dimerization of serotonin 5-HT2A and dopamine D2 receptors. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2010, 1803, 1347–1358. [Google Scholar] [CrossRef] [Green Version]
- Ciruela, F.; Burgueno, J.; Casado, V.; Canals, M.; Marcellino, D.; Goldberg, S.R.; Bader, M.; Fuxe, K.; Agnati, L.F.; Lluis, C.; et al. Combining mass spectrometry and pull-down techniques for the study of receptor heteromerization. Direct epitope-epitope electrostatic interactions between adenosine A2A and dopamine D2 receptors. Anal. Chem. 2004, 76, 5354–5363. [Google Scholar] [CrossRef] [PubMed]
- Ridray, S.; Griffon, N.; Mignon, V.; Souil, E.; Carboni, S.; Diaz, J.; Schwartz, J.-C.; Sokoloff, P. Coexpression of dopamine D1and D3receptors in islands of Calleja and shell of nucleus accumbens of the rat: Opposite and synergistic functional interactions. Eur. J. Neurosci. 1998, 10, 1676–1686. [Google Scholar] [CrossRef]
- Schwartz, J.C.; Diaz, J.; Bordet, R.; Griffon, N.; Perachon, S.; Pilon, C.; Ridray, S.; Sokoloff, P. Functional implications of multi-ple dopamine receptor subtypes: The D1/D3 receptor coexistence. Brain Res. Brain Res. Rev. 1998, 26, 236–242. [Google Scholar] [CrossRef]
- Cho, D.-I.; Quan, W.; Oak, M.-H.; Choi, H.J.; Lee, K.Y.; Kim, K.-M. Functional interaction between dopamine receptor subtypes for the regulation of c-fos expression. Biochem. Biophys. Res. Commun. 2007, 357, 1113–1118. [Google Scholar] [CrossRef]
- Marcellino, D.; Ferre, S.; Casado, V.; Cortes, A.; Le Foll, B.; Mazzola, C.; Drago, F.; Saur, O.; Stark, H.; Soriano, A.; et al. Identification of dopamine D1-D3 receptor heteromers. Indications for a role of synergistic D1-D3 receptor interactions in the striatum. J. Biol. Chem. 2008, 283, 26016–26025. [Google Scholar] [CrossRef] [Green Version]
- Fiorentini, C.; Busi, C.; Gorruso, E.; Gotti, C.; Spano, P.; Missale, C. Reciprocal Regulation of Dopamine D1 and D3 Receptor Function and Trafficking by Heterodimerization. Mol. Pharmacol. 2008, 74, 59–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guitart, X.; Moreno, E.; Rea, W.; Sánchez-Soto, M.; Cai, N.-S.; Quiroz, C.; Kumar, V.; Bourque, L.; Cortés, A.; Canela, E.I.; et al. Biased G Protein-Independent Signaling of Dopamine D1-D3 Receptor Heteromers in the Nucleus Accumbens. Mol. Neurobiol. 2019, 56, 6756–6769. [Google Scholar] [CrossRef] [PubMed]
- Bontempi, L.; Savoia, P.; Bono, F.; Fiorentini, C.; Missale, C. Dopamine D3 and acetylcholine nicotinic receptor heteromeriza-tion in midbrain dopamine neurons: Relevance for neuroplasticity. Eur. Neuropsychopharmacol. 2017, 27, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, B.; Diaz, J.; Sokoloff, P. Increased dopamine D3 receptor expression accompanying behavioral sensitization to nicotine in rats. Synapse 2003, 47, 176–183. [Google Scholar] [CrossRef]
- Smith, L.N.; Bachus, S.E.; McDonald, C.G.; Smith, R.F. Role of the D3 dopamine receptor in nicotine sensitization. Behav. Brain Res. 2015, 289, 92–104. [Google Scholar] [CrossRef]
- Le Foll, B.; Goldberg, S.R.; Sokoloff, P. Dopamine D3receptor ligands for the treatment of tobacco dependence. Expert Opin. Investig. Drugs 2006, 16, 45–57. [Google Scholar] [CrossRef]
- Acharya, S.; Kundu, D.; Choi, H.J.; Kim, K.M. Metabotropic signaling cascade involved in alpha4beta2 nicotinic acetylcho-line receptor-mediated PKCbetaII activation. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118721. [Google Scholar] [CrossRef]
- Ramazi, S.; Zahiri, J. Posttranslational modifications in proteins: Resources, tools and prediction methods. Database 2021, 2021, baab012. [Google Scholar] [CrossRef]
- Krishna, R.G.; Wold, F. Post-translational modification of proteins. Adv. Enzymol. Relat. Areas Mol. Biol. 1993, 67, 265–298. [Google Scholar]
- Patwardhan, A.; Cheng, N.; Trejo, J. Post-Translational Modifications of G Protein–Coupled Receptors Control Cellular Signaling Dynamics in Space and Time. Pharmacol. Rev. 2020, 73, 120–151. [Google Scholar] [CrossRef]
- Duarte, M.L.; Devi, L.A. Post-translational Modifications of Opioid Receptors. Trends Neurosci. 2020, 43, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Haselbeck, A.; Tanner, W. Dolichyl phosphate-mediated mannosyl transfer through liposomal membranes. Proc. Natl. Acad. Sci. USA 1982, 79, 1520–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bause, E. Structural requirements of N-glycosylation of proteins. Studies with proline peptides as conformational probes. Biochem. J. 1983, 209, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Cantagrel, V.; Lefeber, D.J. From glycosylation disorders to dolichol biosynthesis defects: A new class of metabolic diseases. J. Inherit. Metab. Dis. 2011, 34, 859–867. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Miller, L.J.; Dong, M. Role of N-linked glycosylation in biosynthesis, trafficking, and function of the human glu-cagon-like peptide 1 receptor. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E62–E68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackman, J.J.; Goth, C.K.; Halim, A.; Vakhrushev, S.Y.; Clausen, H.; Petaja-Repo, U.E. Site-specific O-glycosylation of N-terminal serine residues by polypeptide GalNAc-transferase 2 modulates human delta-opioid receptor turnover at the plasma membrane. Cell. Signal. 2018, 42, 184–193. [Google Scholar] [CrossRef]
- Wang, T.; Nakagawa, S.; Miyake, T.; Setsu, G.; Kunisue, S.; Goto, K.; Hirasawa, A.; Okamura, H.; Yamaguchi, Y.; Doi, M. Identi-fication and functional characterisation of N-linked glycosylation of the orphan G protein-coupled receptor Gpr176. Sci. Rep. 2020, 10, 4429. [Google Scholar] [CrossRef] [Green Version]
- Free, R.B.; Hazelwood, L.A.; Cabrera, D.M.; Spalding, H.N.; Namkung, Y.; Rankin, M.L.; Sibley, D.R. D1 and D2 Dopamine Receptor Expression Is Regulated by Direct Interaction with the Chaperone Protein Calnexin. J. Biol. Chem. 2007, 282, 21285–21300. [Google Scholar] [CrossRef] [Green Version]
- Wilden, U.; Kuhn, H. Light-dependent phosphorylation of rhodopsin: Number of phosphorylation sites. Biochemistry 1982, 21, 3014–3022. [Google Scholar] [CrossRef]
- Stadel, J.M.; Nambi, P.; Shorr, R.G.; Sawyer, D.F.; Caron, M.G.; Lefkowitz, R.J. Catecholamine-induced desensitization of tur-key erythrocyte adenylate cyclase is associated with phosphorylation of the beta-adrenergic receptor. Proc. Natl. Acad. Sci. USA 1983, 80, 3173–3177. [Google Scholar] [CrossRef] [Green Version]
- Benovic, J.L.; Strasser, R.H.; Caron, M.G.; Lefkowitz, R.J. Beta-adrenergic receptor kinase: Identification of a novel protein kinase that phosphorylates the agonist-occupied form of the receptor. Proc. Natl. Acad. Sci. USA 1986, 83, 2797–2801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawrence, R.T.; Searle, B.C.; Llovet, A.; Villén, J. Plug-and-play analysis of the human phosphoproteome by targeted high-resolution mass spectrometry. Nat. Methods 2016, 13, 431–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Sharma, K.; Zangrandi, L.; Chen, C.; Humphrey, S.J.; Chiu, Y.-T.; Spetea, M.; Liu-Chen, L.-Y.; Schwarzer, C.; Mann, M. In vivo brain GPCR signaling elucidated by phosphoproteomics. Science 2018, 360, eaao4927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinna, L.A.; Ruzzene, M. How do protein kinases recognize their substrates? Biochim. Biophys. Acta (BBA) Mol. Cell Res. 1996, 1314, 191–225. [Google Scholar] [CrossRef] [Green Version]
- Sibley, D.R.; Peters, J.R.; Nambi, P.; Caron, M.G.; Lefkowitz, R.J. Desensitization of turkey erythrocyte adenylate cyclase. Be-ta-adrenergic receptor phosphorylation is correlated with attenuation of adenylate cyclase activity. J. Biol. Chem. 1984, 259, 9742–9749. [Google Scholar]
- Shichi, H.; Somers, R. Light-dependent phosphorylation of rhodopsin. Purification and properties of rhodopsin kinase. J. Biol. Chem. 1978, 253, 7040–7046. [Google Scholar] [CrossRef]
- Palczewski, K.; Buczyłko, J.; Kaplan, M.; Polans, A.; Crabb, J. Mechanism of rhodopsin kinase activation. J. Biol. Chem. 1991, 266, 12949–12955. [Google Scholar] [CrossRef]
- Samama, P.; Cotecchia, S.; Costa, T.; Lefkowitz, R. A mutation-induced activated state of the beta 2-adrenergic receptor. Extending the ternary complex model. J. Biol. Chem. 1993, 268, 4625–4636. [Google Scholar] [CrossRef]
- Benovic, J.; Onorato, J.; Lohse, M.; Dohlman, H.; Staniszewski, C.; Caron, M.; Lefkowitz, R. Synthetic peptides of the hamster beta 2-adrenoceptor as substrates and inhibitors of the beta-adrenoceptor kinase. Br. J. Clin. Pharmacol. 1990, 30, 3S–12S. [Google Scholar] [CrossRef]
- Chen, C.Y.; Dion, S.B.; Kim, C.M.; Benovic, J.L. Beta-adrenergic receptor kinase. Agonist-dependent receptor binding pro-motes kinase activation. J. Biol. Chem. 1993, 268, 7825–7831. [Google Scholar] [CrossRef]
- Komolov, K.E.; Benovic, J.L. G protein-coupled receptor kinases: Past, present and future. Cell. Signal. 2018, 41, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.G.; Fowles, C.; Sharma, R.; Akhtar, M. Mechanistic studies on rhodopsin kinase. Eur. J. Biochem. 1992, 208, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Pickart, C.M. Mechanisms underlying ubiquitination. Annu. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef] [PubMed]
- Ardley, H.C.; Robinson, P.A. E3 ubiquitin ligases. Essays Biochem. 2005, 41, 15–30. [Google Scholar] [CrossRef] [PubMed]
- Suresh, B.; Lee, J.; Kim, H.; Ramakrishna, S. Regulation of pluripotency and differentiation by deubiquitinating enzymes. Cell Death Differ. 2016, 23, 1257–1264. [Google Scholar] [CrossRef] [Green Version]
- Dittmar, G.; Winklhofer, K.F. Linear Ubiquitin Chains: Cellular Functions and Strategies for Detection and Quantification. Front. Chem. 2020, 7, 915. [Google Scholar] [CrossRef]
- Kennedy, J.E.; Marchese, A. Regulation of GPCR Trafficking by Ubiquitin. Prog. Mol. Biol. Transl. Sci. 2015, 132, 15–38. [Google Scholar] [CrossRef] [Green Version]
- Imai, Y.; Soda, M.; Inoue, H.; Hattori, N.; Mizuno, Y.; Takahashi, R. An Unfolded Putative Transmembrane Polypeptide, which Can Lead to Endoplasmic Reticulum Stress, Is a Substrate of Parkin. Cell 2001, 105, 891–902. [Google Scholar] [CrossRef] [Green Version]
- Marchese, A.; Benovic, J.L. Agonist-promoted Ubiquitination of the G Protein-coupled Receptor CXCR4 Mediates Lysosomal Sorting. J. Biol. Chem. 2001, 276, 45509–45512. [Google Scholar] [CrossRef] [Green Version]
- Aicart-Ramos, C.; Valero, R.A.; Rodriguez-Crespo, I. Protein palmitoylation and subcellular trafficking. Biochim. Biophys. Acta (BBA) Biomembr. 2011, 1808, 2981–2994. [Google Scholar] [CrossRef] [Green Version]
- Resh, M.D. Membrane Targeting of Lipid Modified Signal Transduction Proteins. Sub Cell. Biochem. 2004, 37, 217–232. [Google Scholar] [CrossRef]
- Fukata, Y.; Bredt, D.S.; Fukata, M. Protein Palmitoylation by DHHC Protein Family. In The Dynamic Synapse: Molecular Methods in Ionotropic Receptor Biology; Kittler, J.T., Moss, S.J., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2006. [Google Scholar]
- Dunphy, J.T.; Schroeder, H.; Leventis, R.; Greentree, W.K.; Knudsen, J.K.; Silvius, J.R.; Linder, M.E. Differential effects of acyl-CoA binding protein on enzymatic and non-enzymatic thioacylation of protein and peptide substrates. Biochim. Biophys. Acta 2000, 1485, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Koster, K.P.; Yoshii, A. Depalmitoylation by Palmitoyl-Protein Thioesterase 1 in Neuronal Health and Degeneration. Front. Synaptic Neurosci. 2019, 11, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, A.; Yang, H.-C.; Friedman, P.; Johnson, C.; Dennis, E. A specific human lysophospholipase: cDNA cloning, tissue distribution and kinetic characterization. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 1999, 1437, 157–169. [Google Scholar] [CrossRef]
- Rocks, O.; Peyker, A.; Kahms, M.; Verveer, P.J.; Koerner, C.; Lumbierres, M.; Kuhlmann, J.; Waldmann, H.; Wittinghofer, A.; Bastiaens, P.I.H. An Acylation Cycle Regulates Localization and Activity of Palmitoylated Ras Isoforms. Science 2005, 307, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Verkruyse, L.A.; Hofmann, S.L. Lysosomal Targeting of Palmitoyl-protein Thioesterase. J. Biol. Chem. 1996, 271, 15831–15836. [Google Scholar] [CrossRef] [Green Version]
- Qanbar, R.; Bouvier, M. Role of palmitoylation/depalmitoylation reactions in G-protein-coupled receptor function. Pharmacol. Ther. 2002, 97, 1–33. [Google Scholar] [CrossRef]
- Chini, B.; Parenti, M. G-protein-coupled receptors, cholesterol and palmitoylation: Facts about fats. J. Mol. Endocrinol. 2009, 42, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Ng, G.Y.K.; O’Dowd, B.F.; Caron, M.; Dennis, M.; Brann, M.R.; George, S.R. Phosphorylation and Palmitoylation of the Human D2L Dopamine Receptor in Sf9 Cells. J. Neurochem. 2002, 63, 1589–1595. [Google Scholar] [CrossRef]
- Grunewald, S.; Haase, W.; Reilander, H.; Michel, H. Glycosylation, palmitoylation, and localization of the human D2S receptor in baculovirus-infected insect cells. Biochemistry 1996, 35, 15149–15161. [Google Scholar] [CrossRef]
- Ebersole, B.; Petko, J.; Woll, M.; Murakami, S.; Sokolina, K.; Wong, V.; Stagljar, I.; Lüscher, B.; Levenson, R. Effect of C-Terminal S-Palmitoylation on D2 Dopamine Receptor Trafficking and Stability. PLoS ONE 2015, 10, e0140661. [Google Scholar] [CrossRef]
- Wolf, M.; Dahlin, S.; Hu, X.-T.; Xue, C.-J.; White, K. Effects of lesions of prefrontal cortex, amygdala, or fornix on behavioral sensitization to amphetamine: Comparison with N-methyl-d-aspartate antagonists. Neuroscience 1995, 69, 417–439. [Google Scholar] [CrossRef] [PubMed]
- Cador, M.; Bjijou, Y.; Cailhol, S.; Stinus, L. d-Amphetamine-induced behavioral sensitization: Implication of a glutamatergic medial prefrontal cortex–ventral tegmental area innervation. Neuroscience 1999, 94, 705–721. [Google Scholar] [CrossRef] [PubMed]
- Richtand, N.M. Behavioral Sensitization, Alternative Splicing, and D3 Dopamine Receptor-Mediated Inhibitory Function. Neuropsychopharmacology 2006, 31, 2368–2375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordet, R.; Ridray, S.; Carboni, S.; Diaz, J.; Sokoloff, P.; Schwartz, J.-C. Induction of dopamine D 3 receptor expression as a mechanism of behavioral sensitization to levodopa. Proc. Natl. Acad. Sci. USA 1997, 94, 3363–3367. [Google Scholar] [CrossRef] [Green Version]
- Damsma, G.; Bottema, T.; Westerink, B.H.; Tepper, P.G.; Dijkstra, D.; Pugsley, T.A.; MacKenzie, R.G.; Heffner, T.G.; Wikstrom, H. Pharmacological aspects of R-(+)-7-OH-DPAT, a putative dopamine D3 receptor ligand. Eur. J. Pharmacol. 1993, 249, R9–R10. [Google Scholar] [CrossRef]
- Meller, E.; Bohmaker, K.; Goldstein, M.; Basham, D.A. Evidence that striatal synthesis-inhibiting autoreceptors are dopa-mine D3 receptors. Eur. J. Pharmacol. 1993, 249, R5–R6. [Google Scholar] [CrossRef]
- Koeltzow, T.E.; Xu, M.; Cooper, D.C.; Hu, X.-T.; Tonegawa, S.; Wolf, M.E.; White, F.J. Alterations in Dopamine Release but Not Dopamine Autoreceptor Function in Dopamine D3Receptor Mutant Mice. J. Neurosci. 1998, 18, 2231–2238. [Google Scholar] [CrossRef] [Green Version]
- Kreiss, D.S.; Bergstrom, D.A.; Gonzalez, A.M.; Huang, K.-X.; Sibley, D.R.; Walters, J.R. Dopamine receptor agonist potencies for inhibition of cell firing correlate with dopamine D3 receptor binding affinities. Eur. J. Pharmacol. 1995, 277, 209–214. [Google Scholar] [CrossRef]
- Piercey, M.F.; Hoffmann, W.E.; Smith, M.W.; Hyslop, D.K. Inhibition of dopamine neuron firing by pramipexole, a dopamine D3 receptor-preferring agonist: Comparison to other dopamine receptor agonists. Eur. J. Pharmacol. 1996, 312, 35–44. [Google Scholar] [CrossRef]
- Daly, S.A.; Waddington, J.L. Behavioural effects of the putative D-3 dopamine receptor agonist 7-OH-DPAT in relation to other “D-2-like” agonists. Neuropharmacology 1993, 32, 509–510. [Google Scholar] [CrossRef] [PubMed]
- Boer, P.; Enrico, P.; Wright, J.; Wise, L.D.; Timmerman, W.; Moor, E.; Dijkstra, D.; Wikstrom, H.V.; Westerink, B.H. Charac-terization of the effect of dopamine D3 receptor stimulation on locomotion and striatal dopamine levels. Brain Res. 1997, 758, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Waters, N.; Haadsma-Svensson, S.; Svensson, K.; Sonesson, C.; Carlsson, A. Differential effects of dopamine D2 and D3 receptor antagonists in regard to dopamine release, in vivo receptor displacement and behaviour. J. Neural Transm. 1994, 98, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Sotnikova, T.D.; Gainetdinov, R.R.; Grekhova, T.V.; Rayevsky, K.S. Effects of intrastriatal infusion of D2 and D3 dopamine receptor preferring antagonists on dopamine release in rat dorsal striatum (in vivo microdialysis study). Pharmacol. Res. 2001, 43, 283–290. [Google Scholar] [CrossRef]
- Li, T.; Hou, Y.; Yan, C.X.; Chen, T.; Zhao, Y.; Li, S.B. Dopamine D3 receptor knock-out mice display deficits in locomotor sen-sitization after chronic morphine administration. Neurosci. Lett. 2010, 485, 256–260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
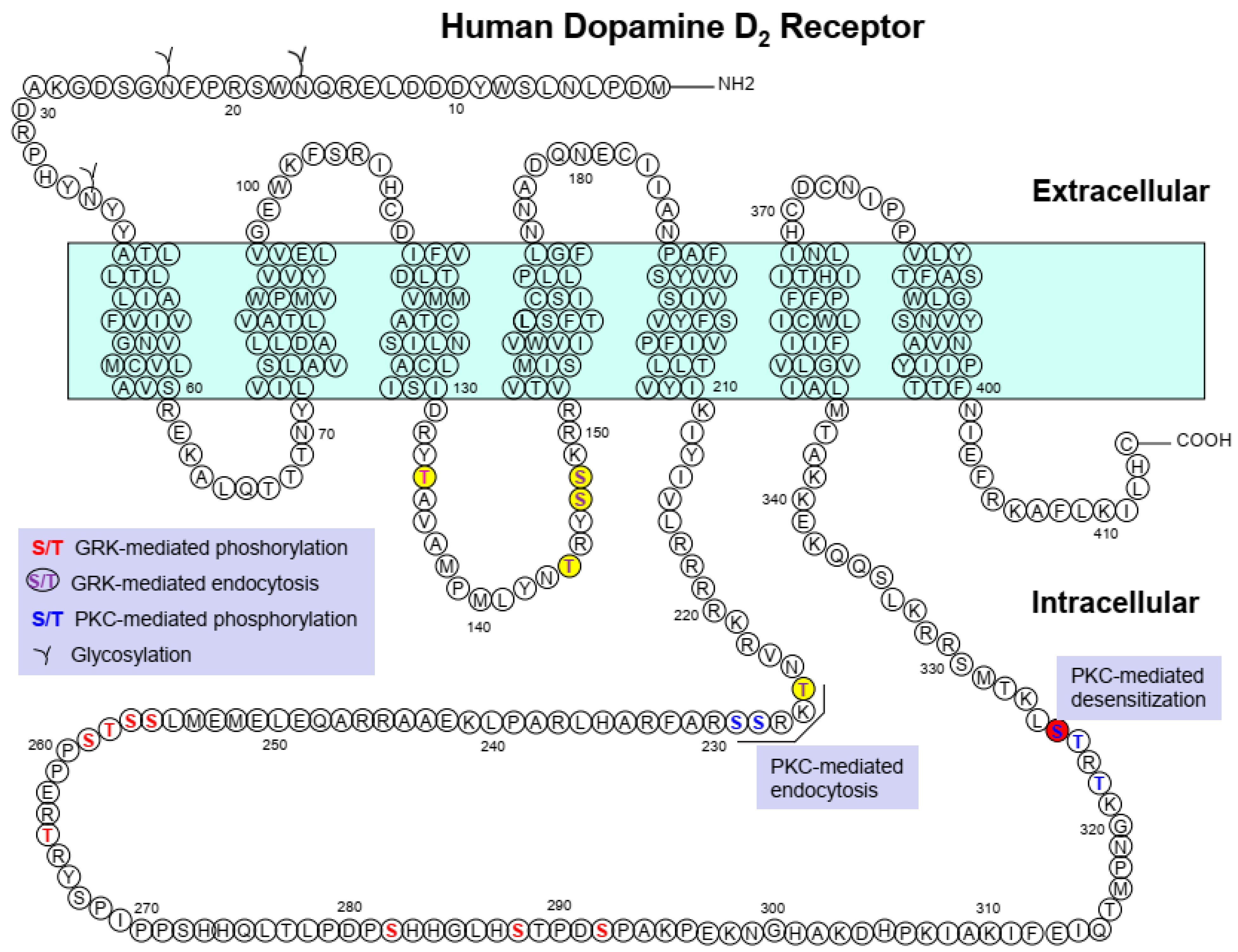
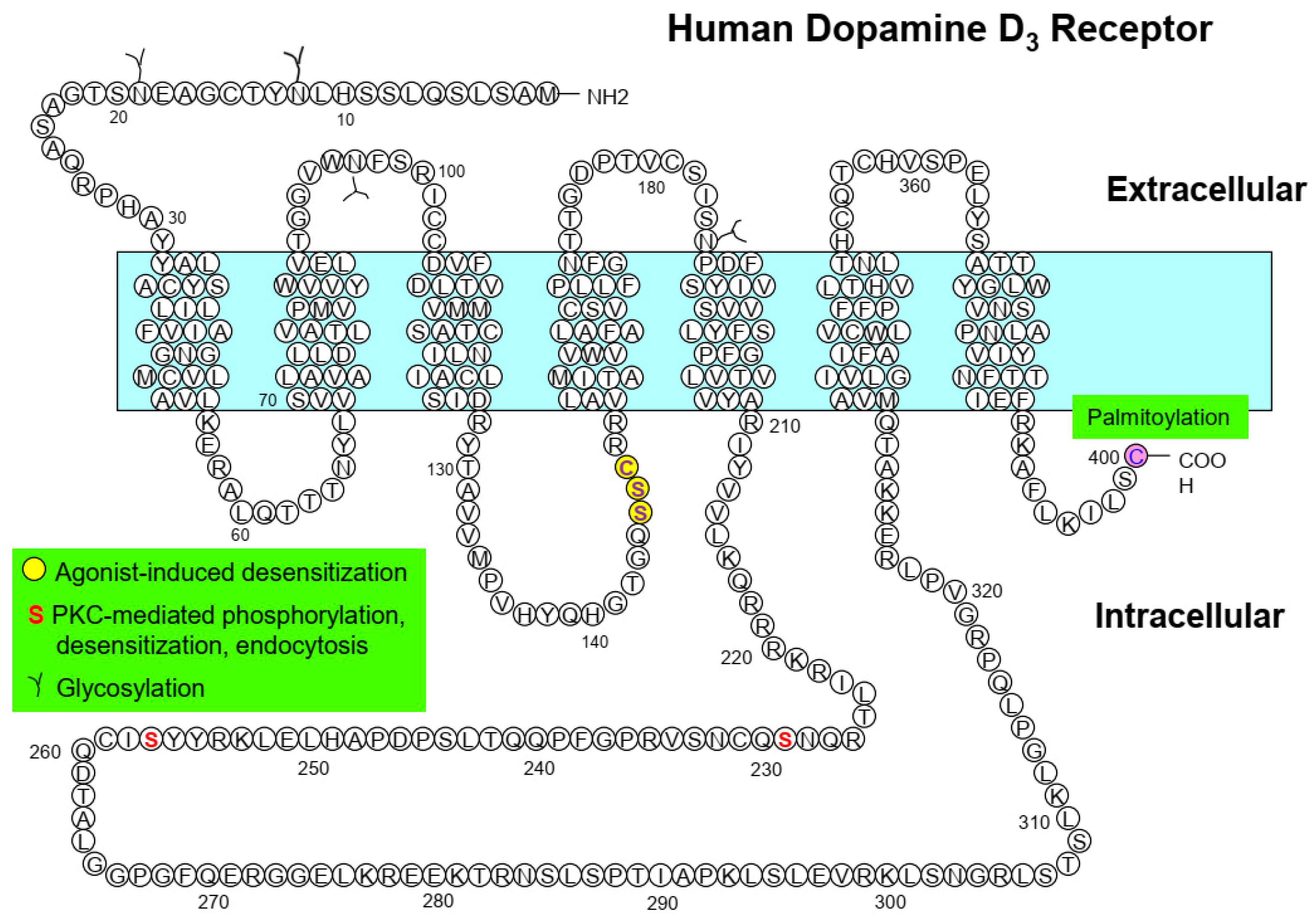
| PTM | Receptor | Sites (Region) | Functional Roles |
|---|---|---|---|
| Glycosylation | D2R | N5, N17, N23 (N-terminus) | Caveolar endocytosis [57,58] |
| D3R | N12, N19 (N-terminus) | Clathrin-mediated endocytosis [58] | |
| N97 (ECL1), N173 (ECL2) | Receptor endocytosis [58] | ||
| Phosphorylation | D2R | S256, S257, T258, S259, T264, S282, S288, S292 | Agonist-induced phosphorylation, receptor recycling [51,53] |
| S228, S229, T322, T324, S325 | PKC-mediated phosphorylation, receptor endocytosis; desensitization (S325) [56] | ||
| D3R | S229, S257 | PKC-mediated phosphorylation, endocytosis, desensitization [43] | |
| Ubiquitination | D2R | None | |
| D3R | ? | PMA-induced lysosomal downregulation [59] | |
| Palmitoylation | D2R | None | |
| D3R | C400 | Cell-surface expression, PKC-mediated endocytosis, agonist affinity, desensitization [31] |
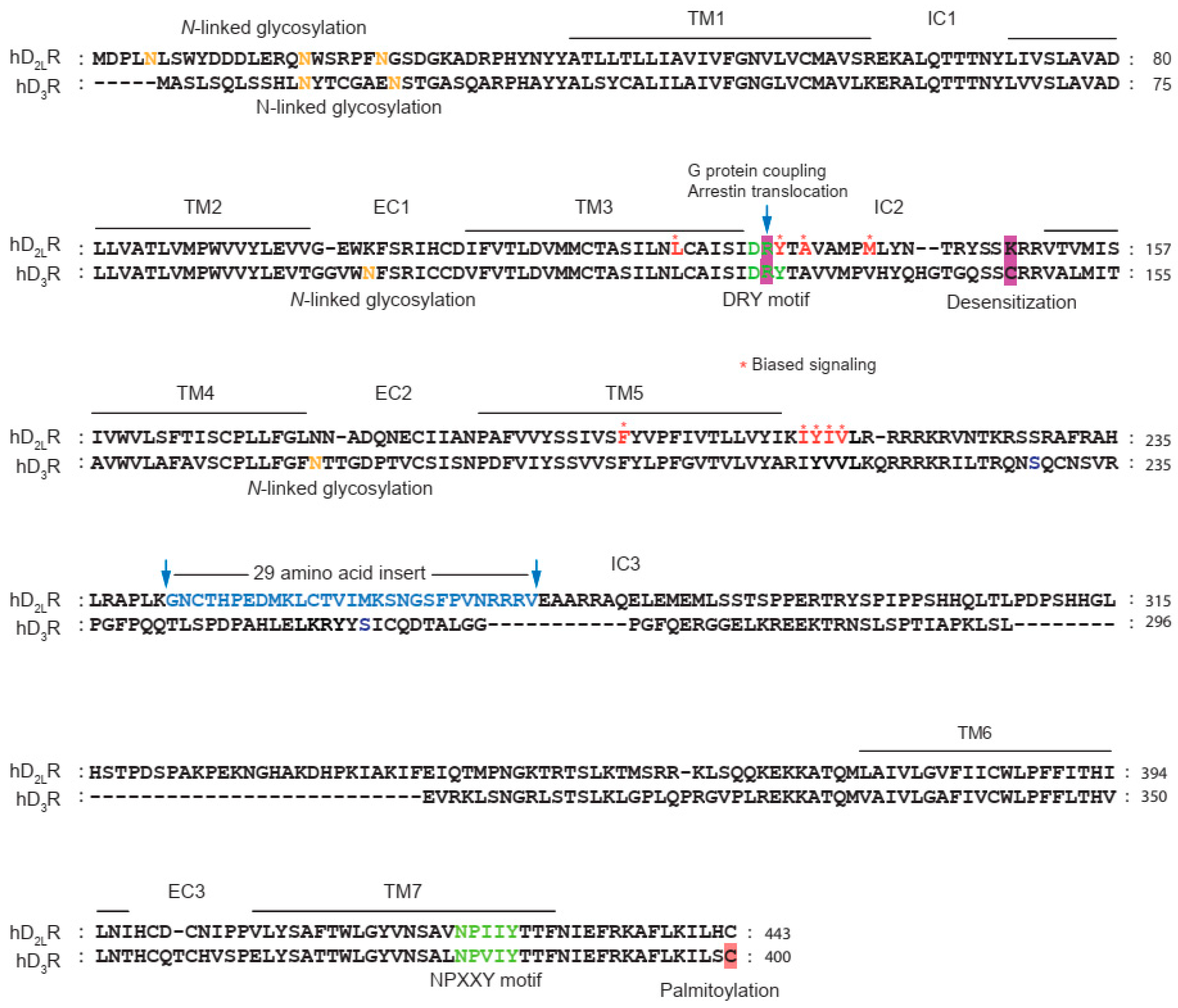
| Sites | Specificity (G Protein vs. Arrestin) |
|---|---|
| F198A (F5.38) | G protein > Arrestin [108] |
| R132L/L123W (R3.50/L3.41) | G protein > Arrestin [109] |
| 212IYIV215 → AAAA | G protein > Arrestin [110] |
| L125N/Y133L | G protein > Arrestin [111] |
| A135R/M140D | G protein < Arrestin [111] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, K.-M. Unveiling the Differences in Signaling and Regulatory Mechanisms between Dopamine D2 and D3 Receptors and Their Impact on Behavioral Sensitization. Int. J. Mol. Sci. 2023, 24, 6742. https://doi.org/10.3390/ijms24076742
Kim K-M. Unveiling the Differences in Signaling and Regulatory Mechanisms between Dopamine D2 and D3 Receptors and Their Impact on Behavioral Sensitization. International Journal of Molecular Sciences. 2023; 24(7):6742. https://doi.org/10.3390/ijms24076742
Chicago/Turabian StyleKim, Kyeong-Man. 2023. "Unveiling the Differences in Signaling and Regulatory Mechanisms between Dopamine D2 and D3 Receptors and Their Impact on Behavioral Sensitization" International Journal of Molecular Sciences 24, no. 7: 6742. https://doi.org/10.3390/ijms24076742