
H3K4me1 Modification Functions in Caste Differentiation in Honey Bees

1
Honeybee Research Institute, Jiangxi Agricultural University, Nanchang 330045, China
2
Jiangxi Province Key Laboratory of Honeybee Biology and Beekeeping, Nanchang 330045, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Int. J. Mol. Sci. 2023, 24(7), 6217; https://doi.org/10.3390/ijms24076217
Submission received: 4 February 2023
/
Revised: 13 March 2023
/
Accepted: 22 March 2023
/
Published: 25 March 2023
(This article belongs to the Special Issue Molecular Ecology, Physiology and Biochemistry of Insects 3.0)
Abstract
:Honey bees are important species for the study of epigenetics. Female honey bee larvae with the same genotype can develop into phenotypically distinct organisms (sterile workers and fertile queens) depending on conditions such as diet. Previous studies have shown that DNA methylation and histone modification can establish distinct gene expression patterns, leading to caste differentiation. It is unclear whether the histone methylation modification H3K4me1 can also impact caste differentiation. In this study, we analyzed genome-wide H3K4me1 modifications in both queen and worker larvae and found that H3K4me1 marks are more abundant in worker larvae than in queen larvae at both the second and fourth instars, and many genes associated with caste differentiation are differentially methylated. Notably, caste-specific H3K4me1 in promoter regions can direct worker development. Thus, our results suggest that H3K4me1 modification may act as an important regulatory factor in the establishment and maintenance of caste-specific transcriptional programs in honey bees; however, the potential influence of other epigenetic modifications cannot be excluded.
1. Introduction
Honey bees are eusocial insects and an important model organism for studies of caste development and caste differentiation in social insects. Their division of labor is mainly based on the differentiation of castes (queen and worker) [1]. After divergence, queens and workers have different morphological, physiological, behavioral, and longevity-related traits, despite sharing the same genome [2]. The mechanism underlying caste differentiation is not fully understood. However, there is evidence that differences in nutritional status between queens and workers modulate caste differentiation by altering DNA methylation patterns [3,4,5,6]. In addition, various signaling pathways, such as the Wnt signaling pathway [7], the target of rapamycin (TOR) nutrient sensing pathway [8,9], and the mitogen-activated protein kinase (MAPK) signaling pathway [10], are related to honey bee caste differentiation.
DNA and histone modifications are thought to primarily affect transcriptional events. The establishment, maintenance, and regulation of transcriptional programs during development depend on chromatin plasticity [11]. Recent evidence suggests that chromatin-based epigenetic mechanisms can influence nutrient-mediated caste differentiation in honey bees. RNAi knockdown of the DNA methyltransferase DNMT3 has a jelly-like effect on developmental trajectories, resulting in a significantly higher proportion of queens with fully developed ovaries [12]. Differences in DNA methylation also influence the alternative development of queens and workers [13]. H3K27ac has been shown to be a key chromatin modification, and the caste-specific region of intronic H3K27ac directs the worker caste [11]. However, for H3K4me3 and H3K36me3, there is no evidence that modification in specific regions can direct caste development [11]. Histone methylation is methylation that occurs on the N-terminal lysine (K) [14] or arginine [15] residues of H3 and H4 histones. Like histone acetylation, histone methylation contributes to almost all biological processes, from DNA repair, the cell cycle, stress responses, and transcription to development, differentiation, and aging [16,17,18,19,20]. It can also regulate the lifespan of model organisms such as rats [21], Caenorhabditis elegans [22], and Drosophila melanogaster [23], and even have a transgenerational effect on lifespan [24,25]. However, there is no evidence that histone methylation contributes to honey bee development.
Hundreds of genes involved in caste differentiation have been identified in honey bees [13,26]. Differences in chromatin levels may lead to differences in gene expression. Histone acetylation influences caste differentiation in honey bees [11]. However, the role of histone methylation in honey bee caste differentiation has not been determined. We performed the first genome-wide analysis of the distribution of H3K4me1 before (2nd instar) and after (4th instar) critical time points (3rd instar) [1,27] in honey bee caste differentiation. Compared with the 2nd instar, the numbers of differentially expressed genes (DEGs) and H3K4me1 markers were significantly increased at the 4th instar, and gene expression was negatively correlated with H3K4me1. Further analysis revealed that the chromatin patterns of queens and workers differed significantly at the 4th instar. H3K4me1 modification promotes larval development towards worker bees. These findings illustrate the important role of H3K4me1 in honey bee larval development and caste differentiation.
2. Results
2.1. H3K4me1 Modifications in the Honey Bee Are Enriched in Transcribed Regions
To investigate the chromatin structure of honey bees, we determined, for the first time, the genome-wide distribution of H3K4me1. We detected H3K4me1 enrichment around the transcriptional start sites (TSS) of genes in both queen larvae and worker larvae (Figure S1). These results are consistent with those for other species, including mammals, invertebrates, and plants [28,29,30]. Over 8 G of data were generated for all samples, with a mapping value and Q30 value of greater than 90% (Table S1). These results indicate that the sequencing results are reliable.
We further evaluated caste-specific patterns in the distribution of H3K4me1. In particular, we divided H3K4me1 into unique peaks (i.e., peaks detected in all three replicates in one sample, but not in any of the three replicates in the other sample) and differential peaks (p < 0.05 and fold change > 2). In 2Q (2nd instar queen) vs. 2W (2nd instar worker), we identified 36 unique H3K4me1 peaks in 2Q and 380 in 2W (Figure 1A). In 4Q vs. 4W, we identified 347 unique H3K4me1 peaks in 4Q and 1185 in 4W (Figure 1A). There were significantly more unique peaks for workers than queens at both the 2nd and 4th instars (Figure S2). There was no significant difference in the relative proportions of the positions of unique peaks between 2Q and 2W (Figure 1B; χ2 = 2.66, p = 0.27, chi-squared test). However, there was a difference in the distribution of unique peaks between 4Q and 4W (Figure 1C; χ2 = 80.18, p = 3.88 × 10−18, chi-squared test); 60% of the unique peaks in 4W were enriched in the promoter region, while 61% of the unique peaks in 4Q were enriched in intronic regions. The data for differential peaks between queens and workers were consistent with the results for unique peaks. In 2Q vs. 2W, we identified 18 differential H3K4me1 peaks in 2Q and 326 in 2W (Figure 1D). In 4Q (4th instar queen) vs. 4W (4th instar worker), we identified 432 differential H3K4me1 peaks in 4Q and 2233 in 4W (Figure 1D). There was no significant difference in the distribution of differential peaks between 2Q and 2W (Figure 1E; χ2 = 4.57, p = 0.10, chi-squared test). However, there was a difference in the distribution of differential peaks between 4Q and 4W (Figure 1F; χ2 = 45.32, p = 1.44 × 10−10, chi-squared test); 55% of unique peaks in workers were found in promoter regions, while 54% of unique peaks in queens were found in intronic regions. In order to study the functional significance of the caste-specific promoter H3K4me1, we performed a motif enrichment analysis using the MEME website (https://meme-suite.org/meme/tools/meme; accessed on 13 December 2022). The sequence of Drosophila melanogaster was used to annotate the honey bee motifs. We identified 2Q and 2W transcription factor binding sites that were significantly enriched, and the top three transcription factor binding sites were identical (Figure S3A). However, the top five enriched transcription factor binding sites of 4Q and 4W were not the same (Figure S3B).
2.2. Caste-Specific H3K4me1 Modification Patterns Correlate with Differential Gene Expression
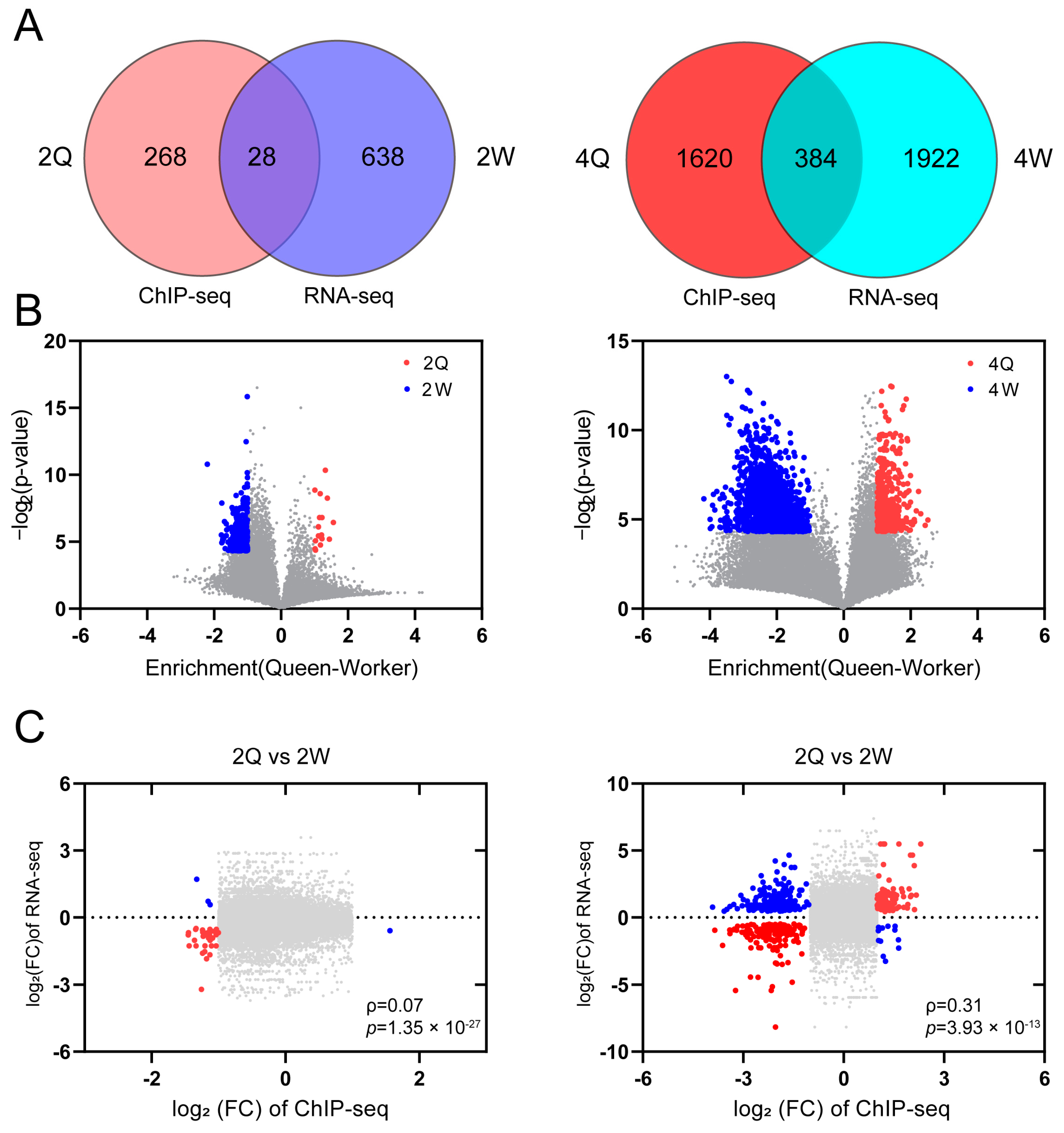
After identifying caste-specific differences in the distribution of H3K4me1 modifications, we next evaluated whether the caste-specific distributions were associated with gene expression patterns according to caste. Principal component analysis of our RNA-seq data revealed strong separation between the two castes (Figure S4). In 2Q vs. 2W, we identified 666 DEGs and 296 differential peak-associated genes (DPGs), of which 28 genes were commonly differentially expressed in RNA-seq and ChIP-seq (Figure 2A). In 4Q vs. 4W, 2306 DEGs and 2004 DPGs were identified, of which 384 genes were commonly differentially expressed in RNA-seq and ChIP-seq (Figure 2A). In 2Q vs. 2W, there were 18 up-regulated peaks and 326 down-regulated peaks (Figure 2B), and 224 up-regulated genes and 452 down-regulated genes (Figure S4). In 2Q vs. 2W, there were 432 up-regulated peaks and 2233 down-regulated peaks (Figure 2B), and 1355 up-regulated genes and 951 down-regulated genes (Figure S4). We found that the genes enriched for the H3K4me1 modification showed significant expression differences based on transcriptional data (Figure 2C; 2Q vs. 2W, ρ = 0.07, p = 1.35 × 10−27, Spearman test; 4Q vs. 4W, ρ = 0.31, p = 3.93 × 10−13, Spearman test). In addition, there were significant correlations between differential H3K4me1 peak signals and transcript levels (Figure S5A) and between DEGs and H3K4me1 peak signals (Figure S5B). Based on the enrichment with enhancer-associated histone H3K4me1 modifications, transcription factor binding sites, promoter sites, and changes in gene expression, worker-specific H3K4me1-enriched regions are markers of active enhancers and play an important role in caste differentiation.
To investigate the effect of H3K4me1-modified genes on caste differentiation, a KEGG analysis was performed. We detected enrichment for distinct developmental processes in the two castes at both the 2nd and 4th instars. Among the unique peak-associated genes of 2W and 4W, eight KEGG pathways were related to honey bee caste differentiation, while among the unique peak-associated genes of 2Q and 4Q, there were only three and five KEGG pathways related to honey bee caste differentiation, respectively (Figure 3A–D). This was consistent with our previous analysis [31], again showing that H3K4me1 modification favors the development of honey bee larvae into worker bees. More DPGs in 4th instar larvae than in 2nd instar larvae were involved in caste differentiation (Figure 3E,F).
2.3. Caste Features of Worker Bees May Be Induced by H3K4me1
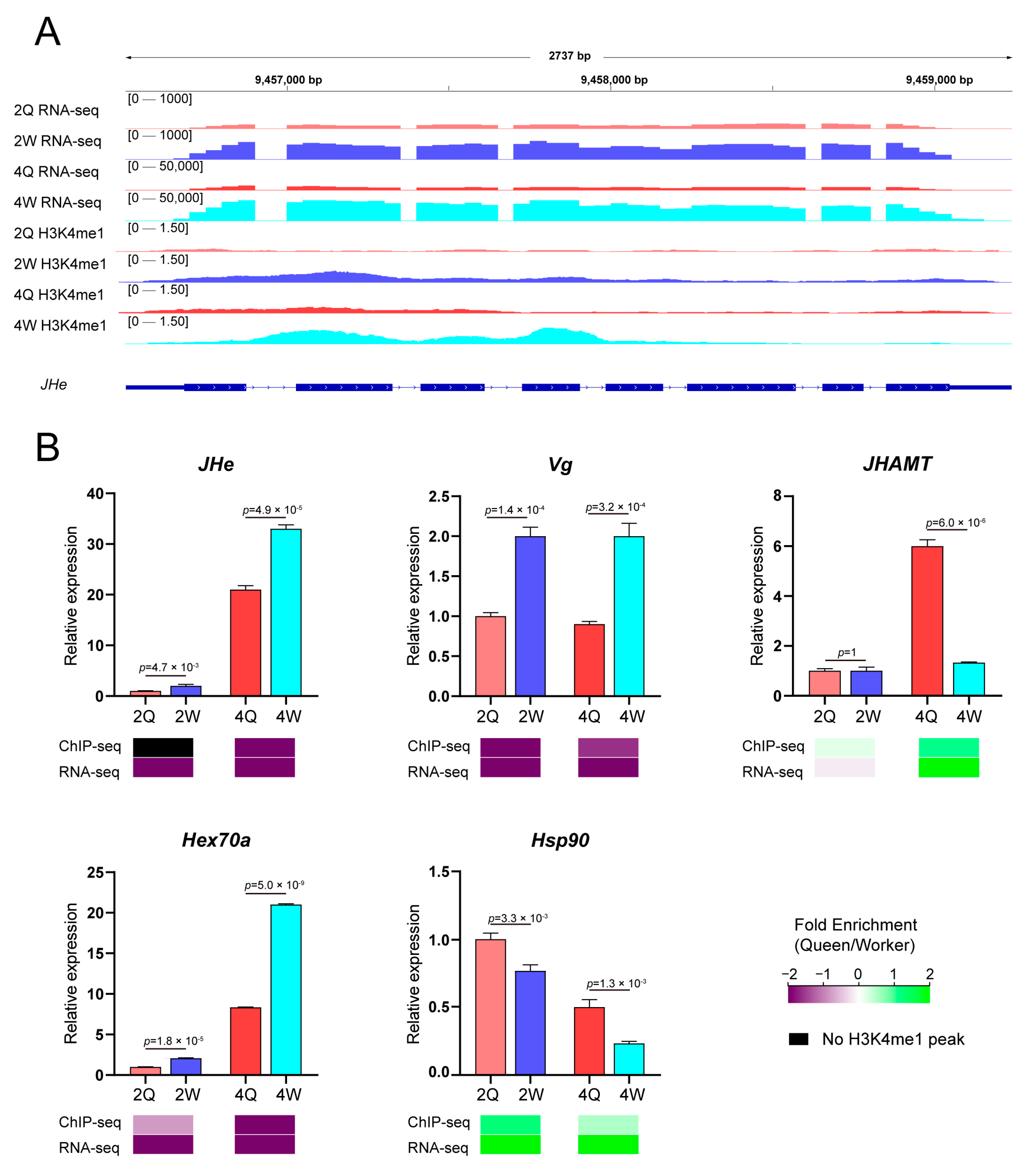
A representative gene involved in highly significant caste-specific changes in both H3K4me1 enrichment and gene expression analysis is shown in Figure 4A. Juvenile hormone esterase (JHe; LOC406066) is shown as an example of a physio-metabolic worker-specific gene [32], in which H3K4me1 enrichment differences are associated with the TSS. Similarly, the other two caste differentiation-related genes, P450-6a17 and IGF, showed significant differences in gene expression and H3K4me1 enrichment in both TSS and gene ontology regions (Figure S6).
In addition, we selected five genes with established roles in the caste differentiation of honey bees for verification, namely JHe [32], Vg (vitellogenin, LOC406088) [33], JHAMT (juvenile hormone acid O-methyltransferase, LOC724216) [34], Hex70a (hexamerin 70a, LOC726848) [35], and Hsp90 (heat shock protein 90, LOC408928) [36]. Our transcriptome results were consistent with those of previous studies [32,35,36,37], and trends in H3K4me1 enrichment were consistent with trends in transcriptome data, thus supporting the reliability of our results. Juvenile hormone (JH) is a master regulator of caste differentiation in honey bees [38]. Vitellogenin is an antagonist of JH. H3K4me1 enrichment in the Vg genes of workers was significantly higher than that in queens at the 2nd and 4th instars. These results further support the role of H3K4me1 modification in regulating transcription, and reveal the effect of H3K4me1 and transcript co-regulation on caste differentiation in honey bees.
3. Discussion
We used ChIP-seq to characterize genome-wide caste-specific chromatin patterns in honey bees and revealed the chromatin patterns related to reproductive division of labor in social insects. Combined with an RNA-seq analysis, the specific modification of H3K4me1 appeared after the critical time point for caste differentiation. There was no significant difference in H3K4me1 modification patterns between worker larvae and queen larvae until the irreversible stage of caste differentiation. A significant number of queen–worker chromatin differences are associated with caste-specific transcription. Importantly, a number of enhancers identified in caste-specific regions may be involved in honey bee caste differentiation.
Previous studies of Drosophila melanogaster [39,40], mice [41,42], and embryonic stem cells from humans and mice have demonstrated that changes in histone methylation affect development and regulate cell fate outcomes [43,44,45]. In honey bees, DNA methylation is predominantly present in gene body regions and the 5′ ends of genes [3,46,47]. Many previous studies have demonstrated the role of DNA methylation in caste differentiation. Different patterns of CpG methylation were detected between the queens and workers [3]. The results of the present study suggest that the developmental asymmetry between queen and worker larvae is associated with an asymmetry in H3K4me1 modification patterns. This is consistent with the distribution of DNA methylation in queen and worker bees [3]. Studies have shown that the degree of enrichment of H3K4me1 differs [48]. In addition, the differential histone methylation of transcription factors that regulate development is thought to alter cell fate decisions [44]. At the same time, the present study also showed that H3K4me1 modification can significantly regulate the transcript level of regulatory factors related to honey bee caste differentiation (such as JHe, Vg, Hex70a, and JHAMT). Signaling pathways such as FoxO and TOR are known to influence caste differentiation in honey bees [38,49], and the present study clearly shows that differential H3K4me1 levels also affect the transcription of genes in these signaling pathways. Notably, caste-specific H3K4me1-modified genes (such as Hex70a and Vg) are related to reproductive division of labor and nutrient metabolism [37,50,51]. These results suggest that H3K4me1 marks contribute to honey bee caste differentiation.
We found that H3K4me1 modification is closely involved in the modification of worker larvae, and is mainly involved in pathways related to honey bee caste differentiation. Caste-specific differences in H3K4me1 were mainly detected in the promoter and intronic regions. Queen-specific H3K4me1 was mainly located in intronic regions. In contrast, worker-specific H3K4me1 was mainly localized in promoter regions, close to transcription initiation sites. In addition, abundant caste-specific H3K4me1 in promoter regions was associated with high levels of caste-specific gene expression, suggesting that these regions play an important cis-regulatory role. Monomethylation of histone H3 at lysine 4 (H3K4me1) is a hallmark of activated enhancers in both vertebrates and invertebrates [28,52]. Enhancers are cis-regulatory DNA sequences and can increase the transcription of target genes. The opening of repressive enhancer–promoter loops leads to transcriptionally active enhancer–promoter regulation as a fundamental mechanism underlying differential transcriptional regulation [53,54]. The spatial organization of chromatin, including long-range enhancers adjacent to target promoters in cis, also modulates gene expression [55]. Therefore, we analyzed the caste-specific promoter region H3K4me1 for conserved transcription factor binding motifs. GATA2 and SPIB accounted for the most transcription factors in 4th instar queens and workers, respectively. Both GATA2 and SPIB can regulate development and cell differentiation [56,57,58,59]. Therefore, H3K4me1 modification may mediate caste-specific enhancer activation, thus directing larval development.
Taken together, we speculate that the worker-specific promoter H3K4me1 region has the hallmarks of an active enhancer. In addition, most worker genes enriched in the promoter H3K4me1 region are also transcription factors, suggesting that enhancers are associated with upstream and downstream genes during the development of worker castes. Queen-specific regions may also be caste-specific enhancers; however, further identification is needed.
4. Materials and Methods
4.1. Insects
Honey bees (Apis mellifera) were obtained from Jiangxi Agricultural University in 2020. Queens were restricted for 6 h (8 am to 2 pm) to a plastic frame designed by Pan et al. [60] to lay eggs in worker cells. The queen laid eggs on a removable plastic base, which was transferred to a plastic queen cell without touching the egg itself. Half of the eggs were transferred to queen cells at 2 pm on the second day after laying and before hatching, while the other half remained in the worker cells. All eggs (in both queen and worker cells) were cared for by workers. Eggs hatched on the third day after laying. To collect the queen and worker larvae at instars 2 and 4, larvae were sampled from both queen and worker cells in each of three colonies at 4 pm on days 5 and 7 after laying. Larvae were picked with sterilized tweezers and rinsed in ddH2O three times. Filter paper was used to drain the water from the larvae, and larvae were placed immediately in liquid nitrogen. Whole larvae were used for sequencing, as in previous studies, since they were very small (2nd instar worker and queen larvae: 2 mg/larva; 4th instar worker larvae: 12 mg/larva; 4th instar queen larvae: 25 mg/larva).
4.2. ChIP-seq Assay and Analysis
Chromatin immunoprecipitation was performed as described by Wojciechowski et al. [11], with slight modifications. Approximately 800 mg of larvae (320 larvae per sample in 2Q and 2W; 32 larvae per sample in 4Q and 4W) were cross-linked for 10 min in 1% ChIP-seq–grade formaldehyde. The H3K4me1 antibody (ab195391; Abcam, Cambridge, UK) was used for immunoprecipitation. The H3K4me1 library was sequenced (50 bp single-end reads or 150 bp paired-end reads) on an Illumina HiSeq3000 sequencer.
The genome assembly Amel_HAv3.1 (GCF_003254395.2) was downloaded from NCBI and indexed using Bowtie 2 (v2.3.3). ChIP-seq samples were mapped to this indexed genome using Bowtie 2 with the default parameters. Detailed mapping statistics for each sample are available in Tables S2 and S3.
4.3. RNA-seq Analysis
Hierarchical Indexing for Spliced Alignment of Transcripts (HISAT; version 2.1.5) was used to align the RNA-seq reads to the reference genome (Amel_HAv3.1). Expression levels were reported as FPKM values to normalize the length of the annotated transcripts and the total number of reads aligned to the transcriptome. Differential expression was analyzed using the DESeq2 R package (1.30.0). p-values were corrected for multiple comparisons with a false discovery rate (value < 0.05). Genes with p-adj ≤ 0.05 were defined as DEGs. Detailed statistics are available in Tables S4 and S5.
4.4. Verification of Gene Expression Differences by qRT-PCR
The 2nd and 4th instar larvae of queens and workers were sampled. Three samples were collected for each type of larvae, with each sample coming from a different colony. Therefore, a total of 12 larval samples across three different colonies were evaluated.
qRT-PCR was performed according to previously described methods [61]. Briefly, total RNAs were extracted using the TransZol Up Plus RNA Kit (TransGen Biotech, Beijing, China), and then transcribed into cDNA using a PrimeScript RT Reagent Kit (Takara, Kusatsu, Japan). GAPDH was used as a reference gene. The primer sequences were designed using Prime Primer 6.0 (Table 1). A 10 μL reaction system (5 μL of SYBR®Premix Ex Taq™ II, 3 μL of H2O, 1 μL of cDNA, 0.4 μL each of forward and reverse primers, and 0.2 μL of ROX) was established. The PCR conditions were as follows: 95 °C, 5 min; 94 °C, 2 min; 40 cycles (95 °C, 10 s, Tm, 15 s, 72 °C, 15 s); 72 °C, 10 min. To establish the melting curve of the qRT-PCR product, the primers were heated slowly with a gradual increase of 1 °C every 5 s from 72 °C to 99 °C. The data were analyzed using the 2−ΔΔCT method.
5. Conclusions
Significant differences in H3K4me1 modification between queen and worker larvae were observed during caste differentiation in honey bees. Chromatin modification can regulate the transcription of the genes that determine the caste. Furthermore, H3K4me1 modification was closely involved in the regulation of the development of worker larvae, and may be an important modification in the worker development pathway. These findings clearly establish the contribution of histone methylation to honey bee development, and may contribute to further research on caste differentiation and developmental plasticity.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms24076217/s1.
Author Contributions
Conceptualization, Y.Z., Z.L., Z.W., X.H. and Z.Z.; methodology, Y.Z.; software, Y.Z.; validation, Z.W. and Z.Z.; formal analysis, Y.Z. and Z.L.; investigation, Z.Z.; resources, Z.Z.; data curation, Y.Z. and Z.L.; writing—original draft preparation, Y.Z.; writing—review and editing, Y.Z. and Z.L.; visualization, Y.Z. and Z.L.; supervision, Z.Z.; project administration, Z.Z.; funding acquisition, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (32172790, 31872432) and the Earmarked Fund for China Agriculture Research System (CARS-44-KXJ15).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Raw sequencing reads for RNA-seq are available at SRA; accession: PRJNA770835. Raw sequencing reads for ChIP-seq are available at SRAl accession: PRJNA891375.
Acknowledgments
Thanks to Nogogene for providing the raw data analysis.
Conflicts of Interest
The authors declare that there are no conflict of interest.
References
- Weaver, N. Physiology of caste determination. Annu. Rev. 1966, 11, 79–102. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D.; Michener, C.D. The social Behavior of the Bees: A Comparative Study; Harvard University Press: Cambridge, MA, USA, 1974. [Google Scholar]
- Elango, N.; Hunt, B.G.; Goodisman, M.A.; Yi, S.V. DNA methylation is widespread and associated with differential gene expression in castes of the honeybee, Apis mellifera. Proc. Natl. Acad. Sci. USA 2009, 106, 11206–11211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, D. Honeybees and cell lines as models of DNA methylation and aging in response to diet. Exp. Gerontol. 2013, 48, 614–619. [Google Scholar] [CrossRef] [PubMed]
- Yagound, B.; Remnant, E.J.; Buchmann, G.; Oldroyd, B.P. Intergenerational transfer of DNA methylation marks in the honey bee. Proc. Natl. Acad. Sci. USA 2020, 117, 32519–32527. [Google Scholar] [CrossRef] [PubMed]
- Yagound, B.; Smith, N.M.; Buchmann, G.; Oldroyd, B.P.; Remnant, E.J. Unique DNA methylation profiles are associated with cis-variation in honey bees. Genome Biol. Evol. 2019, 11, 2517–2530. [Google Scholar] [CrossRef]
- Shi, Y.Y.; Yan, W.Y.; Huang, Z.Y.; Wang, Z.L.; Wu, X.B.; Zeng, Z.J. Genomewide analysis indicates that queen larvae have lower methylation levels in the honey bee (Apis mellifera). Nat. 2013, 100, 193–197. [Google Scholar] [CrossRef]
- Wheeler, D.; Buck, N.; Evans, J. Expression of insulin/insulin-like signalling and TOR pathway genes in honey bee caste determination. Insect Mol. Biol. 2014, 23, 113–121. [Google Scholar] [CrossRef]
- Patel, A.; Fondrk, M.K.; Kaftanoglu, O.; Emore, C.; Hunt, G.; Frederick, K.; Amdam, G.V. The making of a queen: TOR pathway is a key player in diphenic caste development. PLoS ONE 2007, 2, e509. [Google Scholar] [CrossRef] [Green Version]
- He, X.J.; Wei, H.; Jiang, W.J.; Liu, Y.B.; Wu, X.B.; Zeng, Z. Honeybee (Apis mellifera) maternal effect causes alternation of DNA methylation regulating queen development. Sociobiology 2021, 68, e5935. [Google Scholar] [CrossRef]
- Wojciechowski, M.; Lowe, R.; Maleszka, J.; Conn, D.; Maleszka, R.; Hurd, P.J. Phenotypically distinct female castes in honey bees are defined by alternative chromatin states during larval development. Genome Res. 2018, 28, 1532–1542. [Google Scholar] [CrossRef] [Green Version]
- Kucharski, R.; Maleszka, J.; Foret, S.; Maleszka, R. Nutritional control of reproductive status in honeybees via DNA methylation. Science 2008, 319, 1827–1830. [Google Scholar] [CrossRef] [Green Version]
- Foret, S.; Kucharski, R.; Pellegrini, M.; Feng, S.; Jacobsen, S.E.; Robinson, G.E.; Maleszka, R. DNA methylation dynamics, metabolic fluxes, gene splicing, and alternative phenotypes in honey bees. Proc. Natl. Acad. Sci. USA 2012, 109, 4968–4973. [Google Scholar] [CrossRef] [Green Version]
- Murray, K. The occurrence of iε-N-methyl lysine in histones. Biochemistry 1964, 3, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Byvoet, P.; Shepherd, G.; Hardin, J.; Noland, B. The distribution and turnover of labeled methyl groups in histone fractions of cultured mammalian cells. Arch. Biochem. Biophys. 1972, 148, 558–567. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissenberg, J.C.; Shilatifard, A. Histone H3 lysine 4 (H3K4) methylation in development and differentiation. Dev. Biol. 2010, 339, 240–249. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, R.A. Histone tails: Directing the chromatin response to DNA damage. FEBS Lett. 2011, 585, 2883–2890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nottke, A.; Colaiácovo, M.P.; Shi, Y. Developmental roles of the histone lysine demethylases. Development 2009, 136, 879–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, M.T.; Helin, K. Histone demethylases in development and disease. Trends Cell Biol. 2010, 20, 662–671. [Google Scholar] [CrossRef]
- Sarg, B.; Koutzamani, E.; Helliger, W.; Rundquist, I.; Lindner, H.H. Postsynthetic trimethylation of histone H4 at lysine 20 in mammalian tissues is associated with aging. J. Biol. Chem. 2002, 277, 39195–39201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maures, T.J.; Greer, E.L.; Hauswirth, A.G.; Brunet, A. The H3K27 demethylase UTX-1 regulates C. elegans lifespan in a germline-independent, insulin-dependent manner. Aging Cell 2011, 10, 980–990. [Google Scholar] [CrossRef] [Green Version]
- Siebold, A.P.; Banerjee, R.; Tie, F.; Kiss, D.L.; Moskowitz, J.; Harte, P.J. Polycomb Repressive Complex 2 and Trithorax modulate Drosophila longevity and stress resistance. Proc. Natl. Acad. Sci. USA 2010, 107, 169–174. [Google Scholar] [CrossRef] [Green Version]
- Guttman, M.; Amit, I.; Garber, M.; French, C.; Lin, M.F.; Feldser, D.; Huarte, M.; Zuk, O.; Carey, B.W.; Cassady, J.P. Chromatin signature reveals over a thousand highly conserved large non-coding RNAs in mammals. Nature 2009, 458, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Greer, E.L.; Maures, T.J.; Ucar, D.; Hauswirth, A.G.; Mancini, E.; Lim, J.P.; Benayoun, B.A.; Shi, Y.; Brunet, A. Transgenerational epigenetic inheritance of longevity in Caenorhabditis elegans. Nature 2011, 479, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Barchuk, A.R.; Cristino, A.S.; Kucharski, R.; Costa, L.F.; Simões, Z.L.; Maleszka, R. Molecular determinants of caste differentiation in the highly eusocial honeybee Apis mellifera. BMC Dev. Biol. 2007, 7, 70. [Google Scholar] [CrossRef] [Green Version]
- Waddington, C.D. The Strategy of the Genes. A Discussion of Some Aspects of Theoretical Biology; Allen and Unwin: London, UK, 1957. [Google Scholar]
- Local, A.; Huang, H.; Albuquerque, C.P.; Singh, N.; Lee, A.Y.; Wang, W.; Wang, C.; Hsia, J.E.; Shiau, A.K.; Ge, K. Identification of H3K4me1-associated proteins at mammalian enhancers. Nat. Genet. 2018, 50, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Simola, D.F.; Ye, C.; Mutti, N.S.; Dolezal, K.; Bonasio, R.; Liebig, J.; Reinberg, D.; Berger, S.L. A chromatin link to caste identity in the carpenter ant Camponotus floridanus. Genome Res. 2013, 23, 486–496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, K.; Ding, Y.; Malkaram, S.; Riethoven, J.-J.M.; Liu, R.; Yang, J.; Laczko, P.; Chen, H.; Xia, Y.; Ladunga, I. Dynamic changes in genome-wide histone H3 lysine 4 methylation patterns in response to dehydration stress in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; He, X.J.; Barron, A.; Li, Z.; Jin, M.J.; Wang, Z.; Huang, Q.; Zhang, L.Z.; Wu, X.; Yan, W.Y. The diverging epigenomic landscapes of honeybee queens and workers revealed by multiomic sequencing. Insect Biochem. Mol. Biol. 2023, 155, 103929. [Google Scholar] [CrossRef]
- Bomtorin, A.D.; Mackert, A.; Rosa, G.C.C.; Moda, L.M.; Martins, J.R.; Bitondi, M.M.G.; Hartfelder, K.; Simões, Z.L.P. Juvenile hormone biosynthesis gene expression in the corpora allata of honey bee (Apis mellifera L.) female castes. PLoS ONE 2014, 9, e86923. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Wang, L.; Zhao, Y.; Wang, Y.; Chen, C.; Hu, Y.; Zhu, Y.; Sun, H.; Cheng, Y.; Sun, Q. Single-cell transcriptomic analysis of honeybee brains identifies vitellogenin as caste differentiation-related factor. Iscience 2022, 25, 104643. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Huang, Z.Y.; Liu, F.; Li, Z.; Yan, L.; Zhang, S.; Chen, S.; Zhong, B.; Su, S. Molecular cloning and characterization of juvenile hormone acid methyltransferase in the honey bee, Apis mellifera, and its differential expression during caste differentiation. PLoS ONE 2013, 8, e68544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martins, J.R.; Nunes, F.M.; Cristino, A.S.; Simões, Z.L.; Bitondi, M.M. The four hexamerin genes in the honey bee: Structure, molecular evolution and function deduced from expression patterns in queens, workers and drones. BMC Mol. Biol. 2010, 11, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lago, D.C.; Humann, F.C.; Barchuk, A.R.; Abraham, K.J.; Hartfelder, K. Differential gene expression underlying ovarian phenotype determination in honey bee, Apis mellifera L., caste development. Insect Biochem. Mol. Biol. 2016, 79, 1–12. [Google Scholar] [CrossRef]
- Wang, M.; Xiao, Y.; Li, Y.; Wang, X.; Qi, S.; Wang, Y.; Zhao, L.; Wang, K.; Peng, W.; Luo, G.-Z. RNA m6A modification functions in larval development and caste differentiation in honeybee (Apis mellifera). Cell Rep. 2021, 34, 108580. [Google Scholar] [CrossRef]
- Mutti, N.S.; Dolezal, A.G.; Wolschin, F.; Mutti, J.S.; Gill, K.S.; Amdam, G.V. IRS and TOR nutrient-signaling pathways act via juvenile hormone to influence honey bee caste fate. J. Exp. Biol. 2011, 214, 3977–3984. [Google Scholar] [CrossRef] [Green Version]
- Ingham, P.; Whittle, R. Trithorax: A new homoeotic mutation of Drosophila melanogaster causing transformations of abdominal and thoracic imaginal segments. Mol. Gen. Genet. 1980, 179, 607–614. [Google Scholar] [CrossRef]
- Byrd, K.N.; Shearn, A. ASH1, a Drosophila trithorax group protein, is required for methylation of lysine 4 residues on histone H3. Proc. Natl. Acad. Sci. USA 2003, 100, 11535–11540. [Google Scholar] [CrossRef] [Green Version]
- Ancelin, K.; Syx, L.; Borensztein, M.; Ranisavljevic, N.; Vassilev, I.; Briseno-Roa, L.; Liu, T.; Metzger, E.; Servant, N.; Barillot, E. Maternal LSD1/KDM1A is an essential regulator of chromatin and transcription landscapes during zygotic genome activation. Elife 2016, 5, e08851. [Google Scholar] [CrossRef]
- Andreu-Vieyra, C.V.; Chen, R.; Agno, J.E.; Glaser, S.; Anastassiadis, K.; Stewart, A.F.; Matzuk, M.M. MLL2 is required in oocytes for bulk histone 3 lysine 4 trimethylation and transcriptional silencing. PLoS Biol. 2010, 8, e1000453. [Google Scholar] [CrossRef]
- Dodge, J.E.; Kang, Y.-K.; Beppu, H.; Lei, H.; Li, E. Histone H3-K9 methyltransferase ESET is essential for early development. Mol. Cell. Biol. 2004, 24, 2478–2486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jambhekar, A.; Dhall, A.; Shi, Y. Roles and regulation of histone methylation in animal development. Nat. Rev. Mol. Cell Biol. 2019, 20, 625–641. [Google Scholar] [CrossRef]
- Arney, K.L.; Bao, S.; Bannister, A.J.; Kouzarides, T.; Surani, M.A. Histone methylation defines epigenetic asymmetry in the mouse zygote. Int. J. Dev. Biol. 2002, 46, 317–320. [Google Scholar] [PubMed]
- Suzuki, M.M.; Bird, A. DNA methylation landscapes: Provocative insights from epigenomics. Nat. Rev. Genet. 2008, 9, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Glastad, K.M.; Hunt, B.G.; Goodisman, M.A. Epigenetics in insects: Genome regulation and the generation of phenotypic diversity. Annu. Rev. Entomol. 2019, 64, e203. [Google Scholar] [CrossRef] [Green Version]
- Sharifi-Zarchi, A.; Gerovska, D.; Adachi, K.; Totonchi, M.; Pezeshk, H.; Taft, R.J.; Schöler, H.R.; Chitsaz, H.; Sadeghi, M.; Baharvand, H. DNA methylation regulates discrimination of enhancers from promoters through a H3K4me1-H3K4me3 seesaw mechanism. BMC Genom. 2017, 18, 964. [Google Scholar] [CrossRef]
- He, X.J.; Barron, A.B.; Yang, L.; Chen, H.; He, Y.Z.; Zhang, L.Z.; Huang, Q.; Wang, Z.L.; Wu, X.B.; Yan, W.Y. Extent and complexity of RNA processing in honey bee queen and worker caste development. Iscience 2022, 25, 104301. [Google Scholar] [CrossRef]
- Martins, J.R.; Nunes, F.M.F.; Simões, Z.L.P.; Bitondi, M.M.G. A honeybee storage protein gene, hex 70a, expressed in developing gonads and nutritionally regulated in adult fat body. J. Insect Physiol. 2008, 54, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.R.; Anhezini, L.; Dallacqua, R.P.; Simoes, Z.L.; Bitondi, M.M. A honey bee hexamerin, HEX 70a, is likely to play an intranuclear role in developing and mature ovarioles and testioles. PLoS ONE 2011, 6, e29006. [Google Scholar] [CrossRef] [Green Version]
- Herz, H.-M.; Mohan, M.; Garruss, A.S.; Liang, K.; Takahashi, Y.-h.; Mickey, K.; Voets, O.; Verrijzer, C.P.; Shilatifard, A. Enhancer-associated H3K4 monomethylation by Trithorax-related, the Drosophila homolog of mammalian Mll3/Mll4. Genes Dev. 2012, 26, 2604–2620. [Google Scholar] [CrossRef] [Green Version]
- Dalvai, M.; Bellucci, L.; Fleury, L.; Lavigne, A.; Moutahir, F.; Bystricky, K. H2A. Z-dependent crosstalk between enhancer and promoter regulates Cyclin D1 expression. Oncogene 2013, 32, 4243–4251. [Google Scholar] [CrossRef] [Green Version]
- Dalvai, M.; Fleury, L.; Bellucci, L.; Kocanova, S.; Bystricky, K. TIP48/Reptin and H2A. Z requirement for initiating chromatin remodeling in estrogen-activated transcription. PLoS Genet. 2013, 9, e1003387. [Google Scholar] [CrossRef] [PubMed]
- Mora, A.; Sandve, G.K.; Gabrielsen, O.S.; Eskeland, R. In the loop: Promoter–enhancer interactions and bioinformatics. Brief. Bioinform. 2016, 17, 980–995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, R.M.; Soto, X.; Chen, Y.; Zorn, A.M.; Amaya, E. spib is required for primitive myeloid development in Xenopus. Blood J. Am. Soc. Hematol. 2008, 112, 2287–2296. [Google Scholar] [CrossRef] [Green Version]
- Willis, S.N.; Tellier, J.; Liao, Y.; Trezise, S.; Light, A.; O’Donnell, K.; Garrett-Sinha, L.A.; Shi, W.; Tarlinton, D.M.; Nutt, S.L. Environmental sensing by mature B cells is controlled by the transcription factors PU. 1 and SpiB. Nat. Commun. 2017, 8, 1426. [Google Scholar] [CrossRef] [PubMed]
- Lugus, J.J.; Chung, Y.S.; Mills, J.C.; Kim, S.-I.; Grass, J.A.; Kyba, M.; Doherty, J.M.; Bresnick, E.H.; Choi, K. GATA2 functions at multiple steps in hemangioblast development and differentiation. Development 2007, 134, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazenwadel, J.; Betterman, K.L.; Chong, C.-E.; Stokes, P.H.; Lee, Y.K.; Secker, G.A.; Agalarov, Y.; Demir, C.S.; Lawrence, D.M.; Sutton, D.L. GATA2 is required for lymphatic vessel valve development and maintenance. J. Clin. Investig. 2015, 125, 2979–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Q.-Z.; Wu, X.-B.; Guan, C.; Zeng, Z.-J. A new method of queen rearing without grafting larvae. Am. Bee J. 2013, 153, 1279–1280. [Google Scholar]
- Zhang, Y.; Li, Z.; Wang, Z.-L.; Zhang, L.-Z.; Zeng, Z.-J. A Comparison of RNA Interference via Injection and Feeding in Honey Bees. Insects 2022, 13, 928. [Google Scholar] [CrossRef]
Figure 1.
(A) Bar plot showing the number of unique H3K4me1 peaks in queen and worker larvae. (B,C) Bar plot showing the percentage of unique H3K4me1 ChIP-seq peaks within promoters, introns, and exons in 2Q vs. 2W and 4Q vs. 4W. (D) Bar plot showing the number of differential H3K4me1 peaks in queen and worker larvae. (E,F) Bar plot showing the percentage of differential H3K4me1 ChIP-seq peaks within promoters, introns, and exons in 2Q vs. 2W and 4Q vs. 4W.
Figure 1.
(A) Bar plot showing the number of unique H3K4me1 peaks in queen and worker larvae. (B,C) Bar plot showing the percentage of unique H3K4me1 ChIP-seq peaks within promoters, introns, and exons in 2Q vs. 2W and 4Q vs. 4W. (D) Bar plot showing the number of differential H3K4me1 peaks in queen and worker larvae. (E,F) Bar plot showing the percentage of differential H3K4me1 ChIP-seq peaks within promoters, introns, and exons in 2Q vs. 2W and 4Q vs. 4W.

Figure 2.
(A) Venn diagram showing overlap of differentially H3K4me1-modified genes and DEGs between queens and workers. (B) Volcano plot of the difference in enrichment between queens and workers against the negative log p-value for the H3K4me1 signal. Red areas indicate that the H3K4me1 modification is up-regulated in queens. Blue areas indicate that the H3K4me1 modification is up-regulated in workers. Gray indicates a lack of a significant difference (p > 0.05). (C) Scatter plots of the significant differences in expression, determined by ChIP-seq, between queen and worker larvae (x-axis) against the Log2FC of transcript expression between queen and worker larvae (y-axis). Red indicates that the differences determined by RNA-seq and ChIP-seq are in agreement. Blue indicates that the opposite patterns were obtained by RNA-seq and ChIP-seq. Gray indicates that there is no significant difference (p > 0.05).
Figure 2.
(A) Venn diagram showing overlap of differentially H3K4me1-modified genes and DEGs between queens and workers. (B) Volcano plot of the difference in enrichment between queens and workers against the negative log p-value for the H3K4me1 signal. Red areas indicate that the H3K4me1 modification is up-regulated in queens. Blue areas indicate that the H3K4me1 modification is up-regulated in workers. Gray indicates a lack of a significant difference (p > 0.05). (C) Scatter plots of the significant differences in expression, determined by ChIP-seq, between queen and worker larvae (x-axis) against the Log2FC of transcript expression between queen and worker larvae (y-axis). Red indicates that the differences determined by RNA-seq and ChIP-seq are in agreement. Blue indicates that the opposite patterns were obtained by RNA-seq and ChIP-seq. Gray indicates that there is no significant difference (p > 0.05).

Figure 3.
Negative log2 p-values for the honey bee caste differentiation-related KEGG pathways that are enriched with unique H3K4me1-related genes in 2Q (A), 2W (B), 4Q (C), and 4W (D). Pathways marked in red font are pathways associated with caste differentiation, while black font is not. (E) KEGG pathway enrichment analysis of differentially H3K4me1 peak-related genes in 2Q vs. 2W. (F) KEGG pathway enrichment of differentially H3K4me1 peak-related genes in 4Q vs. 4W.
Figure 3.
Negative log2 p-values for the honey bee caste differentiation-related KEGG pathways that are enriched with unique H3K4me1-related genes in 2Q (A), 2W (B), 4Q (C), and 4W (D). Pathways marked in red font are pathways associated with caste differentiation, while black font is not. (E) KEGG pathway enrichment analysis of differentially H3K4me1 peak-related genes in 2Q vs. 2W. (F) KEGG pathway enrichment of differentially H3K4me1 peak-related genes in 4Q vs. 4W.

Figure 4.
Function of H3K4me1 modification in caste differentiation by modulating the expression levels and modification abundance of genes. (A) Juvenile hormone esterase (JHe; LOC406066). The gene in this region reached genome-wide significance (p ≤ 0.01) at both the 2nd and 4th instars, and had a greater than two-fold difference in H3K4me1 modification. (B) RNA expression levels and H3K4me1 abundance of differentially expressed candidate caste-differentiation-related transcripts are shown. Expression levels are expressed as mean ± SEM relative to a reference gene (GAPDH; LOC726445) in three replications. Fold enrichment is the difference in abundance between queen larvae and worker larvae; green denotes a higher fold enrichment, while purple denotes a lower fold enrichment. Differences in relative expression levels were analyzed using t-tests.
Figure 4.
Function of H3K4me1 modification in caste differentiation by modulating the expression levels and modification abundance of genes. (A) Juvenile hormone esterase (JHe; LOC406066). The gene in this region reached genome-wide significance (p ≤ 0.01) at both the 2nd and 4th instars, and had a greater than two-fold difference in H3K4me1 modification. (B) RNA expression levels and H3K4me1 abundance of differentially expressed candidate caste-differentiation-related transcripts are shown. Expression levels are expressed as mean ± SEM relative to a reference gene (GAPDH; LOC726445) in three replications. Fold enrichment is the difference in abundance between queen larvae and worker larvae; green denotes a higher fold enrichment, while purple denotes a lower fold enrichment. Differences in relative expression levels were analyzed using t-tests.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primer sequences for quantitative qRT-PCR.
| Genes | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | GCTGGTTTCATCGATGGTTT | ACGATTTCGACCACCGTAAC |
| JHe | CTTTTCTCGCTTCCACAACC | TCCTGGTCCAGCAATGTGTA |
| VG | AAGACCAATCCACCGTTGAG | TGGTTCACGCTCCTAGCTTT |
| JHAMT | GGATTTGCCCAAAGACACAT | CGAGGATTCGCGTACAATTT |
| Hex70a | GAGGGTCAAGCATGGAACAT | GTTGTTCTTCGCCCAGAGAG |
| Hsp90 | CTGAGAGTGACGCGAAGCTA | CTCCGGCATCTTTTCACAAT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Li, Z.; He, X.; Wang, Z.; Zeng, Z. H3K4me1 Modification Functions in Caste Differentiation in Honey Bees. Int. J. Mol. Sci. 2023, 24, 6217. https://doi.org/10.3390/ijms24076217
AMA Style
Zhang Y, Li Z, He X, Wang Z, Zeng Z. H3K4me1 Modification Functions in Caste Differentiation in Honey Bees. International Journal of Molecular Sciences. 2023; 24(7):6217. https://doi.org/10.3390/ijms24076217
Chicago/Turabian StyleZhang, Yong, Zhen Li, Xujiang He, Zilong Wang, and Zhijiang Zeng. 2023. "H3K4me1 Modification Functions in Caste Differentiation in Honey Bees" International Journal of Molecular Sciences 24, no. 7: 6217. https://doi.org/10.3390/ijms24076217
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.