
The Effects of Neuropeptide B on Proliferation and Differentiation of Porcine White Preadipocytes into Mature Adipocytes
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
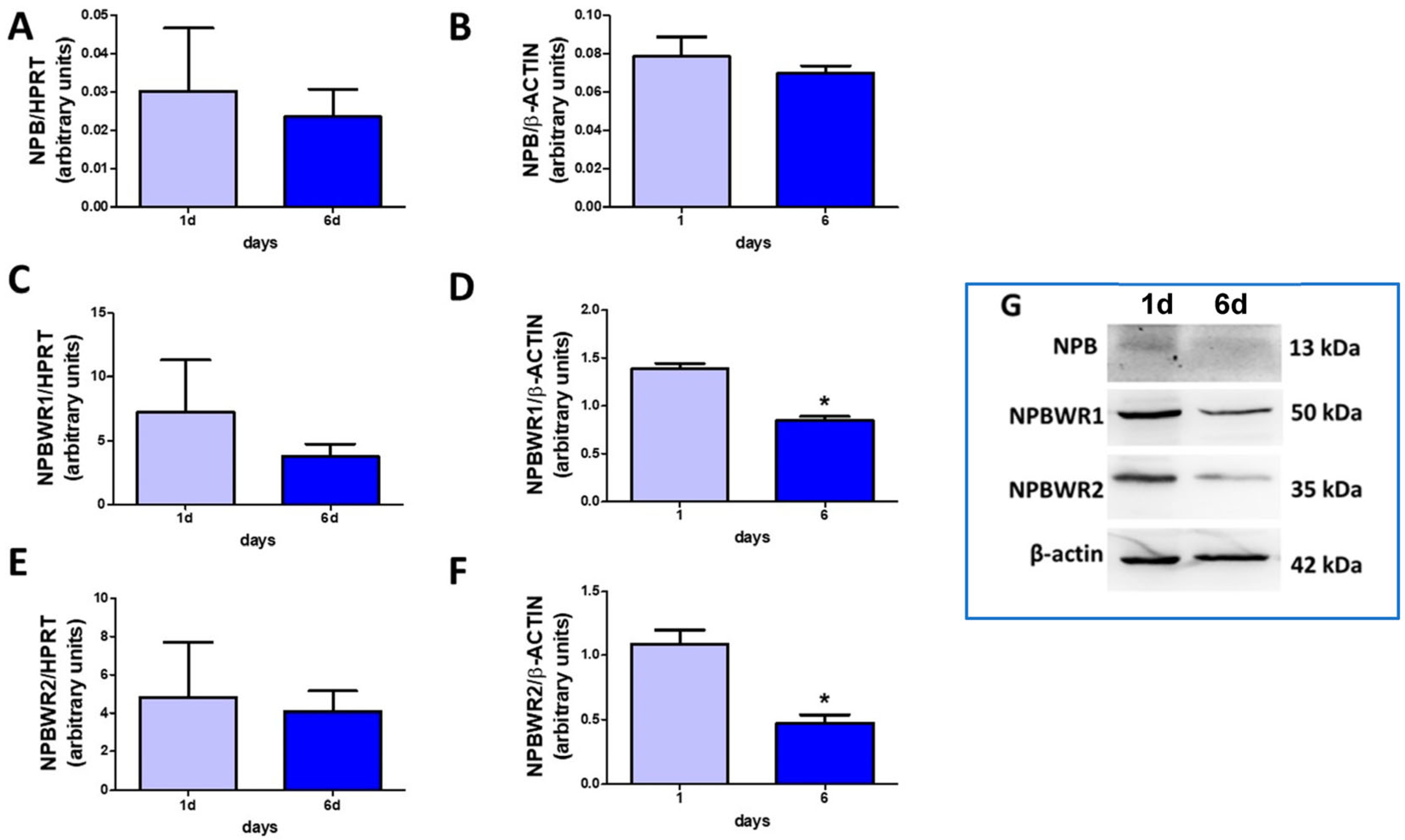
2.1. NPB and Their Receptors Are Present in Porcine Preadipocytes
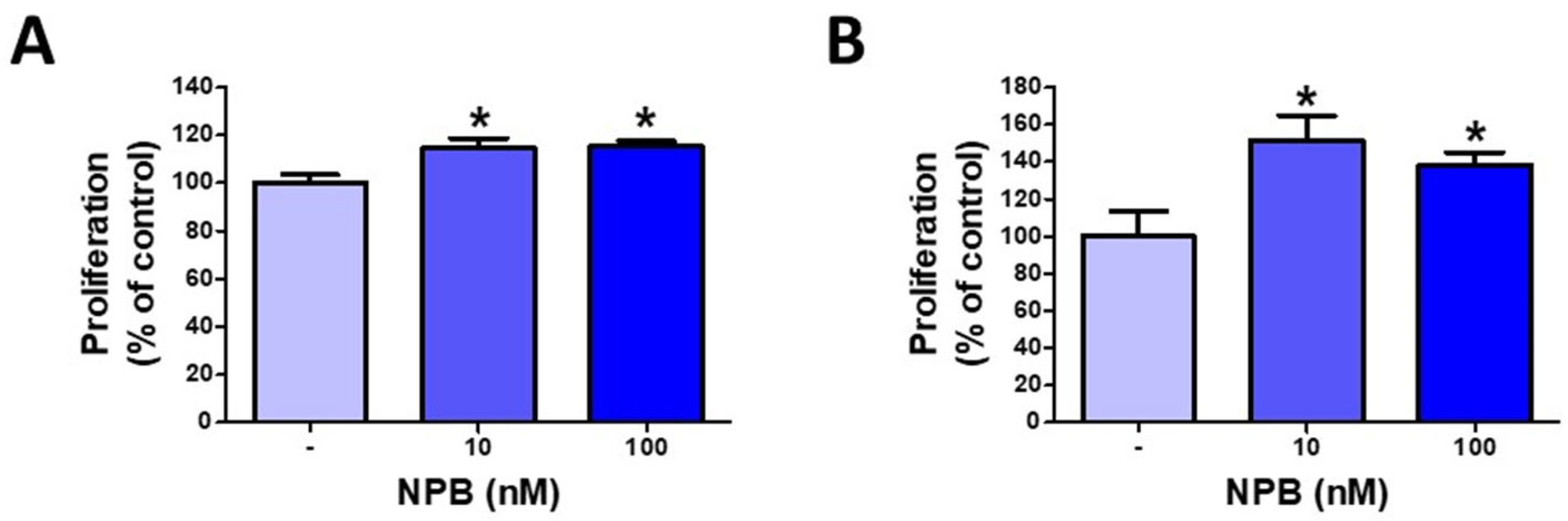
2.2. NPB Promotes Proliferation of Porcine Preadipocytes
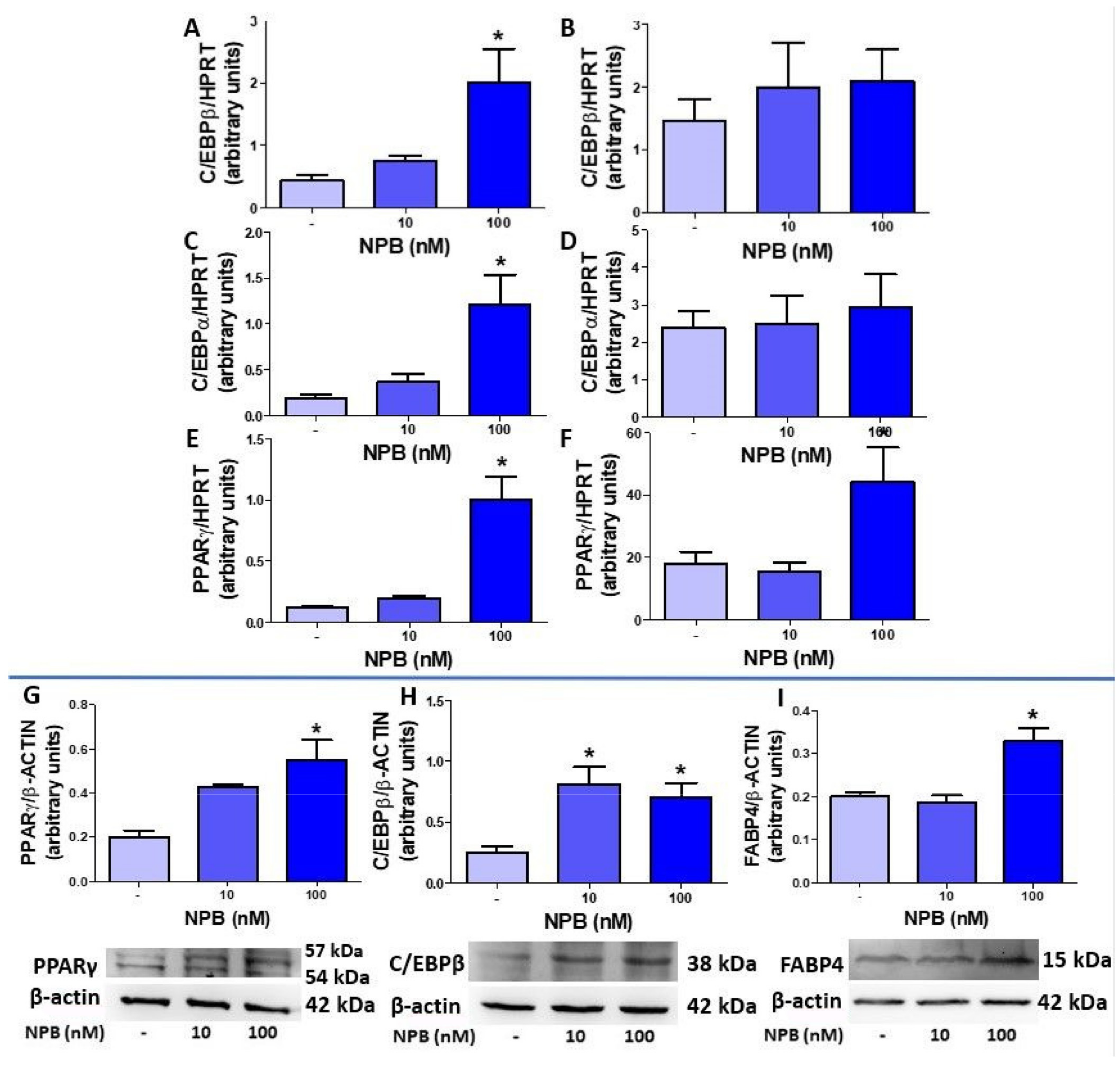
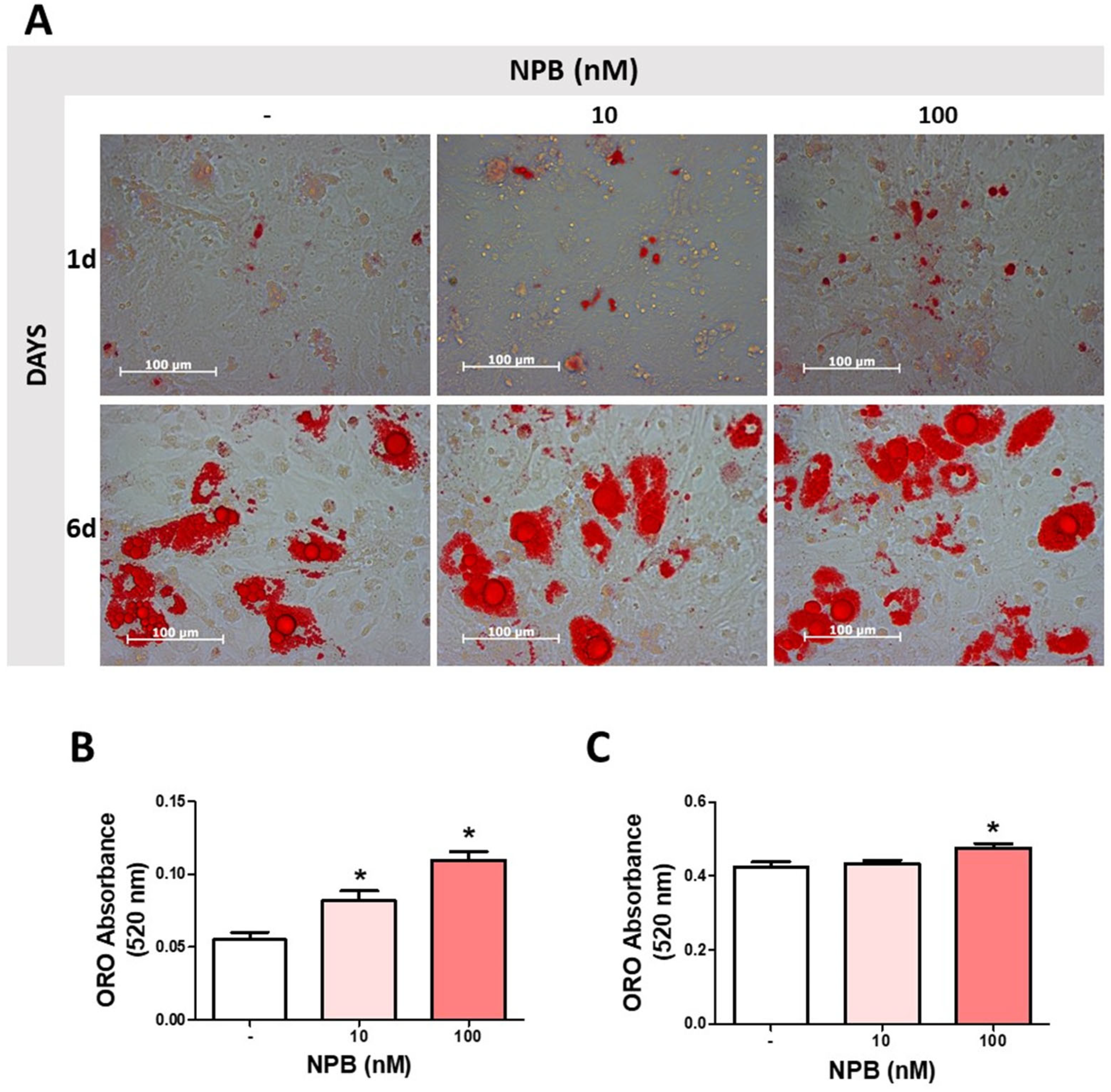
2.3. NPB Stimulates Differentiation of Porcine Preadipocytes into Adipocytes
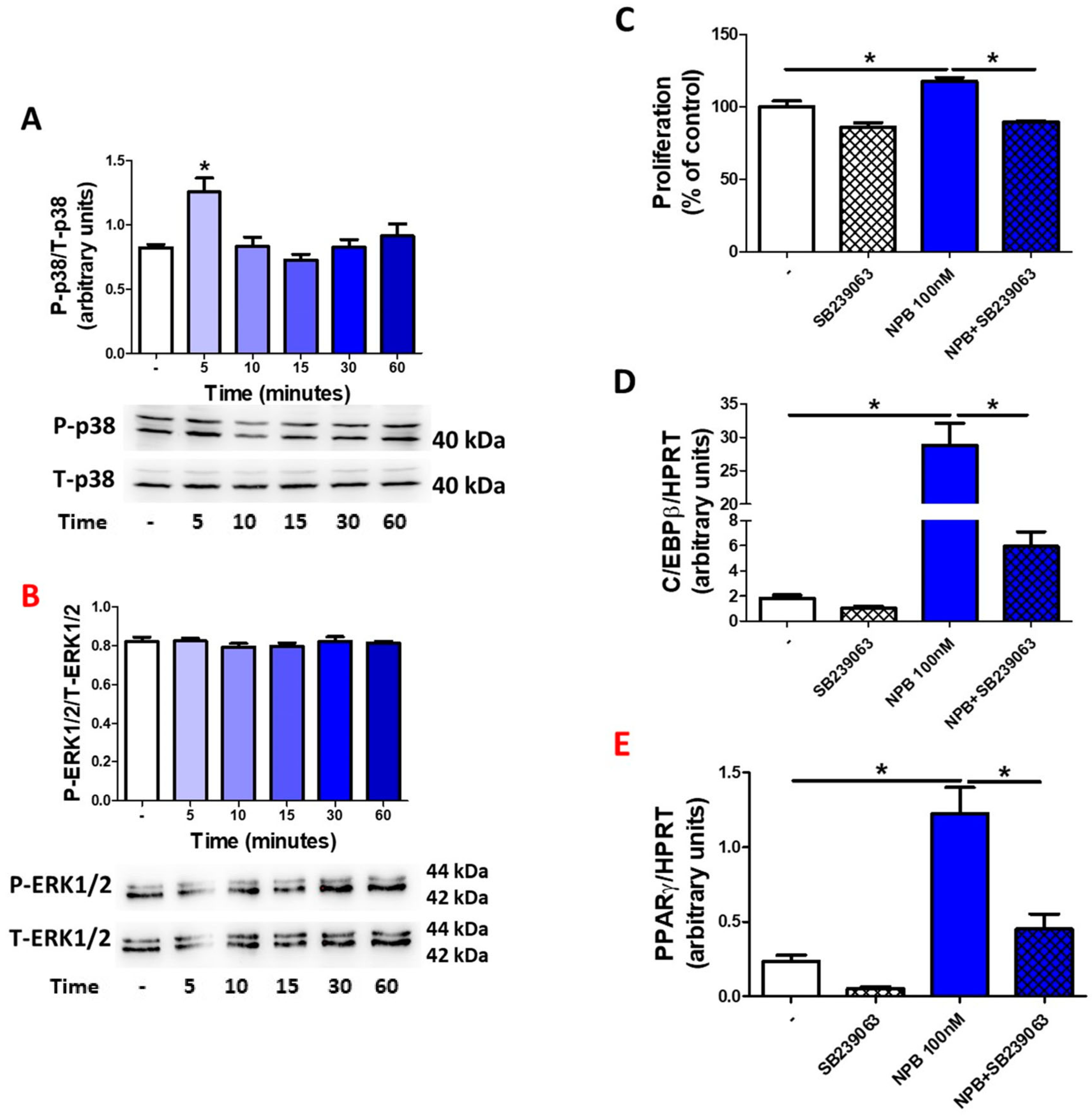
2.4. NPB Modulates Adipogenesis in Porcine Preadipocytes via p38-Dependent Mechanism
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Isolation and Culture
4.3. Cell Proliferation Assay
4.4. Differentiation of White Preadipocytes
4.5. Oil Red O (ORO) Staining and Microphotographs
4.6. Reverse Transcription and Real-Time qPCR
4.7. Western Blot Immunodetection
4.8. Statistical Analysis Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Brezillon, S.; Lannoy, V.; Franssen, J.-D.; le Poul, E.; Dupriez, V.; Lucchetti, J.; Detheux, M.; Parmentier, M. Identification of Natural Ligands for the Orphan G Protein-Coupled Receptors GPR7 and GPR8. J. Biol. Chem. 2003, 278, 776–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujii, R.; Yoshida, H.; Fukusumi, S.; Habata, Y.; Hosoya, M.; Kawamata, Y.; Yano, T.; Hinuma, S.; Kitada, C.; Asami, T.; et al. Identification of a Neuropeptide Modified with Bromine as an Endogenous Ligand for GPR7. J. Biol. Chem. 2002, 277, 34010–34016. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Yoshida, T.; Miyamoto, N.; Motoike, T.; Kurosu, H.; Shibata, K.; Yamanaka, A.; Williams, S.C.; Richardson, J.A.; Tsujino, N.; et al. Characterization of a Family of Endogenous Neuropeptide Ligands for the G Protein-Coupled Receptors GPR7 and GPR8. Proc. Natl. Acad. Sci. USA 2003, 100, 6251–6256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dowd, B.F.; Scheideler, M.A.; Nguyen, T.; Cheng, R.; Rasmussen, J.S.; Marchese, A.; Zastawny, R.; Heng, H.H.Q.; Tsui, L.-C.; Shi, X.; et al. The Cloning and Chromosomal Mapping of Two Novel Human Opioid-Somatostatin-like Receptor Genes, GPR7 and GPR8, Expressed in Discrete Areas of the Brain. Genomics 1995, 28, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.; Su, J.; Zheng, L.; Jin, M.; Hou, Y.; Ma, Z.; Guo, T.; Zhu, S.; Ma, X.; Ahmed, E.; et al. Cloning and Distribution of Neuropeptide W and Its Receptors in Pigs. Res. Vet. Sci. 2015, 101, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Ma, Z.; Suo, C.; Cheng, L.; Su, J.; Lei, Z. Cloning and MRNA Expression of NPB and Its Effect on Hormone Secretion of the Reproductive Cells in the Pig. Gen. Comp. Endocrinol. 2018, 261, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Bu, G.; Lin, D.; Cui, L.; Huang, L.; Lv, C.; Huang, S.; Wan, Y.; Fang, C.; Li, J.; Wang, Y. Characterization of Neuropeptide B (NPB), Neuropeptide W (NPW), and Their Receptors in Chickens: Evidence for NPW Being a Novel Inhibitor of Pituitary GH and Prolactin Secretion. Endocrinology 2016, 157, 3562–3576. [Google Scholar] [CrossRef]
- Hiraki, T.; Nakasone, K.; Hosono, K.; Kawabata, Y.; Nagahama, Y.; Okubo, K. Neuropeptide B Is Female-Specifically Expressed in the Telencephalic and Preoptic Nuclei of the Medaka Brain. Endocrinology 2014, 155, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Mazzocchi, G.; Rebuffat, P.; Ziolkowska, A.; Rossi, G.P.; Malendowicz, L.K.; Nussdorfer, G.G. G Protein Receptors 7 and 8 Are Expressed in Human Adrenocortical Cells, and Their Endogenous Ligands Neuropeptides B and w Enhance Cortisol Secretion by Activating Adenylate Cyclase- and Phospholipase C-Dependent Signaling Cascades. J. Clin. Endocrinol. Metab. 2005, 90, 3466–3471. [Google Scholar] [CrossRef] [Green Version]
- Billert, M.; Sassek, M.; Wojciechowicz, T.; Jasaszwili, M.; Strowski, M.; Nowak, K.; Skrzypski, M. Neuropeptide B Stimulates Insulin Secretion and Expression but Not Proliferation in Rat Insulin-producing INS-1E Cells. Mol. Med. Rep. 2019, 20, 2030–2038. [Google Scholar] [CrossRef]
- Hiraki-Kajiyama, T.; Yamashita, J.; Yokoyama, K.; Kikuchi, Y.; Nakajo, M.; Miyazoe, D.; Nishiike, Y.; Ishikawa, K.; Hosono, K.; Kawabata-Sakata, Y.; et al. Neuropeptide B Mediates Female Sexual Receptivity in Medaka Fish, Acting in a Female-Specific but Reversible Manner. eLife 2019, 8, e39495. [Google Scholar] [CrossRef]
- Bu, G.; Lin, Y.; Liu, J.; Yu, P.; Yong, T.; Yang, M.; Huang, L.; Han, X.; Du, X.; Kong, F.; et al. Evidence for Progesterone Acting as an Inhibitor of Stress Axis via Stimulating Pituitary Neuropeptide B/W Receptor 2 (NPBWR2) Expression in Chickens. J. Steroid Biochem. Mol. Biol. 2022, 226, 106218. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.A.; Beuckmann, C.T.; Williams, S.C.; Sinton, C.M.; Motoike, T.; Richardson, J.A.; Hammer, R.E.; Garry, M.G.; Yanagisawa, M. Neuropeptide B-Deficient Mice Demonstrate Hyperalgesia in Response to Inflammatory Pain. Proc. Natl. Acad. Sci. USA 2005, 102, 9942–9947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wojciechowicz, T.; Billert, M.; Dhandapani, P.; Szczepankiewicz, D.; Wasielewski, O.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Neuropeptide B Promotes Proliferation and Differentiation of Rat Brown Primary Preadipocytes. FEBS Open Bio 2021, 11, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding Adipocyte Differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [CrossRef] [Green Version]
- Lefterova, M.I.; Lazar, M.A. New Developments in Adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Brun, R.P.; Kim, J.B.; Hu, E.; Altiok, S.; Spiegelman, B.M. Adipocyte Differentiation: A Transcriptional Regulatory Cascade. Curr. Opin. Cell Biol. 1996, 8, 826–832. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.D.; Lin, F.T.; MacDougald, O.A.; Vasseur-Cognet, M. Control of Adipocyte Differentiation by CCAAT/Enhancer Binding Protein Alpha (C/EBP Alpha). Int. J. Obes. Relat. Metab. Disord. 1996, 20 (Suppl. S3), S91–S96. [Google Scholar]
- Hwang, C.S.; Mandrup, S.; MacDougald, O.A.; Geiman, D.E.; Lane, M.D. Transcriptional Activation of the Mouse Obese (Ob) Gene by CCAAT/Enhancer Binding Protein Alpha. Proc. Natl. Acad. Sci. USA 1996, 93, 873–877. [Google Scholar] [CrossRef] [Green Version]
- Kaestner, K.H.; Christy, R.J.; Lane, M.D. Mouse Insulin-Responsive Glucose Transporter Gene: Characterization of the Gene and Trans-Activation by the CCAAT/Enhancer Binding Protein. Proc. Natl. Acad. Sci. USA 1990, 87, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Aratani, Y.; Kitagawa, Y. Enhanced Synthesis and Secretion of Type IV Collagen and Entactin during Adipose Conversion of 3T3-L1 Cells and Production of Unorthodox Laminin Complex. J. Biol. Chem. 1988, 263, 16163–16169. [Google Scholar] [CrossRef]
- Spiegelman, B.M.; Frank, M.; Green, H. Molecular Cloning of MRNA from 3T3 Adipocytes. Regulation of MRNA Content for Glycerophosphate Dehydrogenase and Other Differentiation-Dependent Proteins during Adipocyte Development. J. Biol. Chem. 1983, 258, 10083–10089. [Google Scholar] [CrossRef] [PubMed]
- Paulauskis, J.D.; Sul, H.S. Cloning and Expression of Mouse Fatty Acid Synthase and Other Specific MRNAs. Developmental and Hormonal Regulation in 3T3-L1 Cells. J. Biol. Chem. 1988, 263, 7049–7054. [Google Scholar] [CrossRef] [PubMed]
- Bäck, K.; Arnqvist, H.J. Changes in Insulin and IGF-I Receptor Expression during Differentiation of Human Preadipocytes. Growth Horm. IGF Res. 2009, 19, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Cargnello, M.; Roux, P.P. Activation and Function of the MAPKs and Their Substrates, the MAPK-Activated Protein Kinases. Microbiol. Mol. Biol. Rev. 2011, 75, 50–83. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowicz, T.; Szczepankiewicz, D.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. Neuropeptide B Promotes Differentiation of Rodent White Preadipocytes into Mature Adipocytes. Mol. Cell. Endocrinol. 2023, 562, 111850. [Google Scholar] [CrossRef]
- Han, J.; Wu, J.; Silke, J. An Overview of Mammalian P38 Mitogen-Activated Protein Kinases, Central Regulators of Cell Stress and Receptor Signaling. F1000Research 2020, 9, 653. [Google Scholar] [CrossRef]
- Raingeaud, J.; Gupta, S.; Rogers, J.S.; Dickens, M.; Han, J.; Ulevitch, R.J.; Davis, R.J. Pro-Inflammatory Cytokines and Environmental Stress Cause P38 Mitogen-Activated Protein Kinase Activation by Dual Phosphorylation on Tyrosine and Threonine. J. Biol. Chem. 1995, 270, 7420–7426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabiee, A.; Schwämmle, V.; Sidoli, S.; Dai, J.; Rogowska-Wrzesinska, A.; Mandrup, S.; Jensen, O.N. Nuclear Phosphoproteome Analysis of 3T3-L1 Preadipocyte Differentiation Reveals System-Wide Phosphorylation of Transcriptional Regulators. Proteomics 2017, 17, 1600248. [Google Scholar] [CrossRef] [PubMed]
- Hata, K.; Nishimura, R.; Ikeda, F.; Yamashita, K.; Matsubara, T.; Nokubi, T.; Yoneda, T. Differential Roles of Smad1 and P38 Kinase in Regulation of Peroxisome Proliferator-Activating Receptor γ during Bone Morphogenetic Protein 2-Induced Adipogenesis. Mol. Biol. Cell 2003, 14, 545–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Younce, C.; Kolattukudy, P. MCP-1 Induced Protein Promotes Adipogenesis via Oxidative Stress, Endoplasmic Reticulum Stress and Autophagy. Cell. Physiol. Biochem. 2012, 30, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Bahmad, H.F.; Daouk, R.; Azar, J.; Sapudom, J.; Teo, J.C.M.; Abou-Kheir, W.; Al-Sayegh, M. Modeling Adipogenesis: Current and Future Perspective. Cells 2020, 9, 2326. [Google Scholar] [CrossRef]
- Walters, E.M.; Prather, R.S. Advancing Swine Models for Human Health and Diseases. Mo. Med. 2013, 110, 212–215. [Google Scholar]
- Xing, K.; Liu, H.; Zhang, F.; Liu, Y.; Shi, Y.; Ding, X.; Wang, C. Identification of Key Genes Affecting Porcine Fat Deposition Based on Co-Expression Network Analysis of Weighted Genes. J. Anim. Sci. Biotechnol. 2021, 12, 100. [Google Scholar] [CrossRef]
- Font-Nieves, M.; Sans-Fons, M.G.; Gorina, R.; Bonfill-Teixidor, E.; Salas-Pérdomo, A.; Márquez-Kisinousky, L.; Santalucia, T.; Planas, A.M. Induction of COX-2 Enzyme and Down-Regulation of COX-1 Expression by Lipopolysaccharide (LPS) Control Prostaglandin E2 Production in Astrocytes. J. Biol. Chem. 2012, 287, 6454–6468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, E.D.; Walkey, C.J.; Puigserver, P.; Spiegelman, B.M. Transcriptional Regulation of Adipogenesis. Genes Dev. 2000, 14, 1293–1307. [Google Scholar] [CrossRef]
- Baglioni, S.; Cantini, G.; Poli, G.; Francalanci, M.; Squecco, R.; di Franco, A.; Borgogni, E.; Frontera, S.; Nesi, G.; Liotta, F.; et al. Functional Differences in Visceral and Subcutaneous Fat Pads Originate from Differences in the Adipose Stem Cell. PLoS ONE 2012, 7, e36569. [Google Scholar] [CrossRef]
- Ibrahim, M.M. Subcutaneous and Visceral Adipose Tissue: Structural and Functional Differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef]
- Furuhashi, M.; Hotamisligil, G.S. Fatty Acid-Binding Proteins: Role in Metabolic Diseases and Potential as Drug Targets. Nat. Rev. Drug Discov. 2008, 7, 489–503. [Google Scholar] [CrossRef] [Green Version]
- Woolf, P.J.; Linderman, J.J. Untangling Ligand Induced Activation and Desensitization of G-Protein–Coupled Receptors. Biophys. J. 2003, 84, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bost, F.; Aouadi, M.; Caron, L.; Even, P.; Belmonte, N.; Prot, M.; Dani, C.; Hofman, P.; Pagès, G.; Pouysségur, J.; et al. The Extracellular Signal-Regulated Kinase Isoform ERK1 Is Specifically Required for in Vitro and in Vivo Adipogenesis. Diabetes 2005, 54, 402–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donzelli, E.; Lucchini, C.; Ballarini, E.; Scuteri, A.; Carini, F.; Tredici, G.; Miloso, M. ERK1 and ERK2 Are Involved in Recruitment and Maturation of Human Mesenchymal Stem Cells Induced to Adipogenic Differentiation. J. Mol. Cell Biol. 2011, 3, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leiva, M.; Matesanz, N.; Pulgarín-Alfaro, M.; Nikolic, I.; Sabio, G. Uncovering the Role of P38 Family Members in Adipose Tissue Physiology. Front. Endocrinol. 2020, 11, 572089. [Google Scholar] [CrossRef]
- Guindi, C.; Cloutier, A.; Gaudreau, S.; Zerif, E.; McDonald, P.P.; Tatsiy, O.; Asselin, C.; Dupuis, G.; Gris, D.; Amrani, A. Role of the P38 MAPK/C/EBPβ Pathway in the Regulation of Phenotype and IL-10 and IL-12 Production by Tolerogenic Bone Marrow-Derived Dendritic Cells. Cells 2018, 7, 256. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Jin, B.; Li, Y.-P. C/EBPβ Mediates Tumour-Induced Ubiquitin Ligase Atrogin1/MAFbx Upregulation and Muscle Wasting. EMBO J. 2011, 30, 4323–4335. [Google Scholar] [CrossRef] [Green Version]
- Bradley, M.N.; Zhou, L.; Smale, S.T. C/EBPbeta Regulation in Lipopolysaccharide-Stimulated Macrophages. Mol. Cell Biol. 2003, 23, 4841–4858. [Google Scholar] [CrossRef] [Green Version]
- Engelman, J.A.; Lisanti, M.P.; Scherer, P.E. Specific Inhibitors of P38 Mitogen-Activated Protein Kinase Block 3T3-L1 Adipogenesis. J. Biol. Chem. 1998, 273, 32111–32120. [Google Scholar] [CrossRef] [Green Version]
- Ando, Y.; Sato, F.; Fukunaga, H.; Iwasaki, Y.; Chiba, Y.; Tebakari, M.; Daigo, Y.; Kawashima, J.; Kamei, J. Placental Extract Suppresses Differentiation of 3T3-L1 Preadipocytes to Mature Adipocytes via Accelerated Activation of P38 MAPK during the Early Phase of Adipogenesis. Nutr. Metab. 2019, 16, 32. [Google Scholar] [CrossRef]
- Wu, W.; Xu, K.; Li, M.; Zhang, J.; Wang, Y. MicroRNA-29b/29c Targeting CTRP6 Influences Porcine Adipogenesis via the AKT/PKA/MAPK Signalling Pathway. Adipocyte 2021, 10, 264–274. [Google Scholar] [CrossRef]
- Albrecht, W. Which Concentrations Are Optimal for in Vitro Testing? EXCLI J. 2020, 19, 1172–1173. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, M.; Segaloff, D.L. Effects of Collagenase on the Structure of the Lutropin/Choriogonadotropin Receptor. J. Biol. Chem. 1986, 261, 3807–3815. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowicz, T.; Skrzypski, M.; Szczepankiewicz, D.; Hertig, I.; Kołodziejski, P.A.; Billert, M.; Strowski, M.Z.; Nowak, K.W. Original Research: Orexins A and B Stimulate Proliferation and Differentiation of Porcine Preadipocytes. Exp. Biol. Med. 2016, 241, 1786–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, T.G.; White, M.E.; Wolverton, C.K. Glucocorticoids and the Differentiation of Porcine Preadipocytes4. J. Anim. Sci. 1989, 67, 2222–2229. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Left Primer (5′-3′) | Right Primer (5′-3′) | NCBI Reference Sequence |
|---|---|---|---|
| NPB NPBWR1 NPBWR2 C/EBPα C/EBPβ PPARγ HPRT | gatgtcttcctgtccctccg caacatagccgacttcctgc gacacaacatcaccttcccg ctcaccgctccgattcctac tccgatctcttctccgacga ttccatgctgtcatgggtgaaa cattcctatgactgtagattt | gaaaagacggggcttggtac agaggctggagaaggtgttg gcagatgagggagtacacca tccttctattgcgggggaga caggctcacgtagccgtatt accatggtcacctcttgtgaa ctttggattatgctgcttg | XM_021066433.1 XM_003355060.4 XM_021077700.1 XM_003127015.4 AB569088.1 XM_013981981.2 NM_001032376 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojciechowicz, T.; Kolodziejski, P.A.; Billert, M.; Strowski, M.Z.; Nowak, K.W.; Skrzypski, M. The Effects of Neuropeptide B on Proliferation and Differentiation of Porcine White Preadipocytes into Mature Adipocytes. Int. J. Mol. Sci. 2023, 24, 6096. https://doi.org/10.3390/ijms24076096
Wojciechowicz T, Kolodziejski PA, Billert M, Strowski MZ, Nowak KW, Skrzypski M. The Effects of Neuropeptide B on Proliferation and Differentiation of Porcine White Preadipocytes into Mature Adipocytes. International Journal of Molecular Sciences. 2023; 24(7):6096. https://doi.org/10.3390/ijms24076096
Chicago/Turabian StyleWojciechowicz, Tatiana, Paweł A. Kolodziejski, Maria Billert, Mathias Z. Strowski, Krzysztof W. Nowak, and Marek Skrzypski. 2023. "The Effects of Neuropeptide B on Proliferation and Differentiation of Porcine White Preadipocytes into Mature Adipocytes" International Journal of Molecular Sciences 24, no. 7: 6096. https://doi.org/10.3390/ijms24076096