
Parasite and Pesticide Impacts on the Bumblebee (Bombus terrestris) Haemolymph Proteome

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Whole-Organism Metrics
2.1.1. Survival
2.1.2. Weight Change
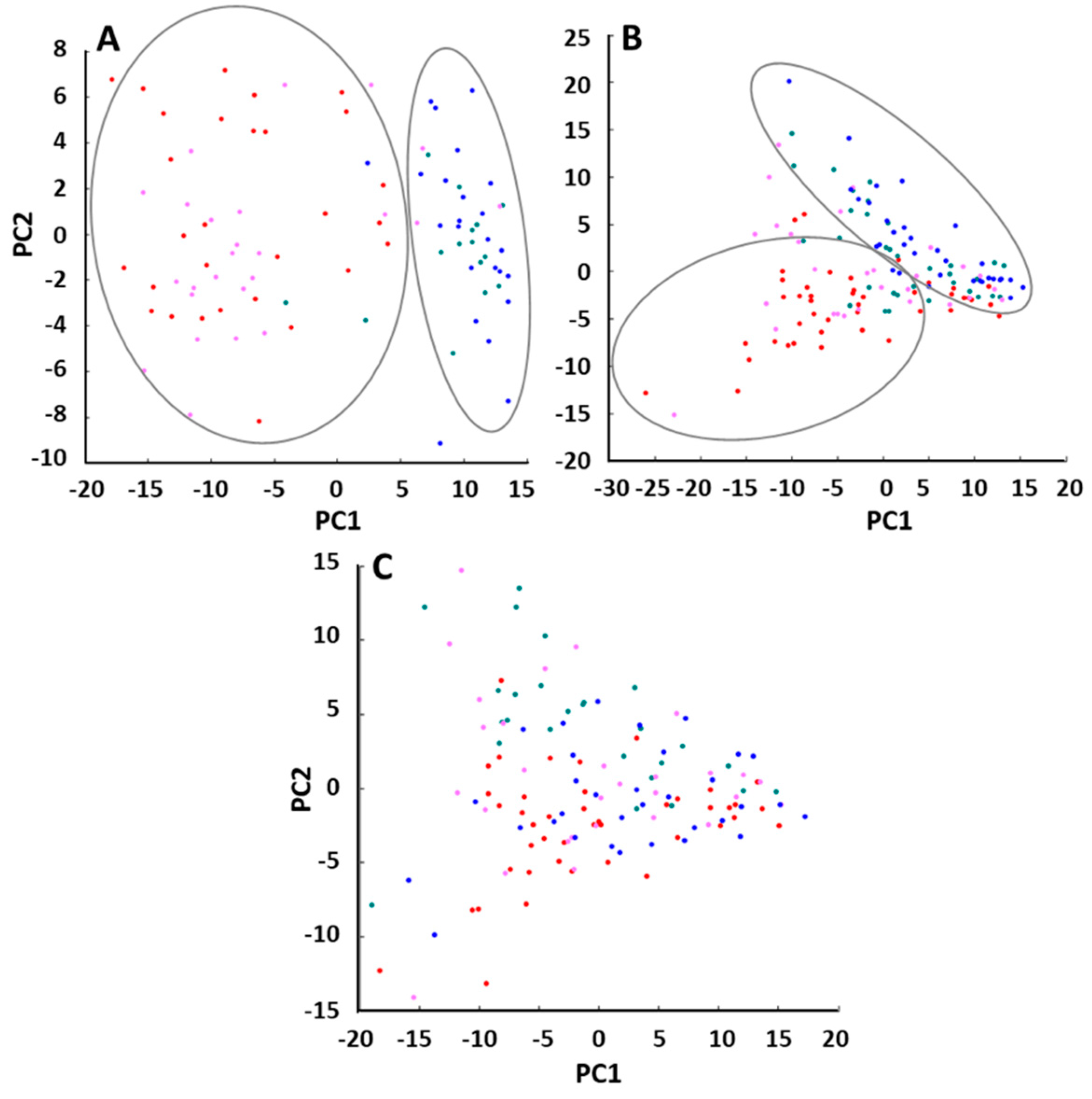
2.2. Molecular Mass Fingerprints (MFPs)
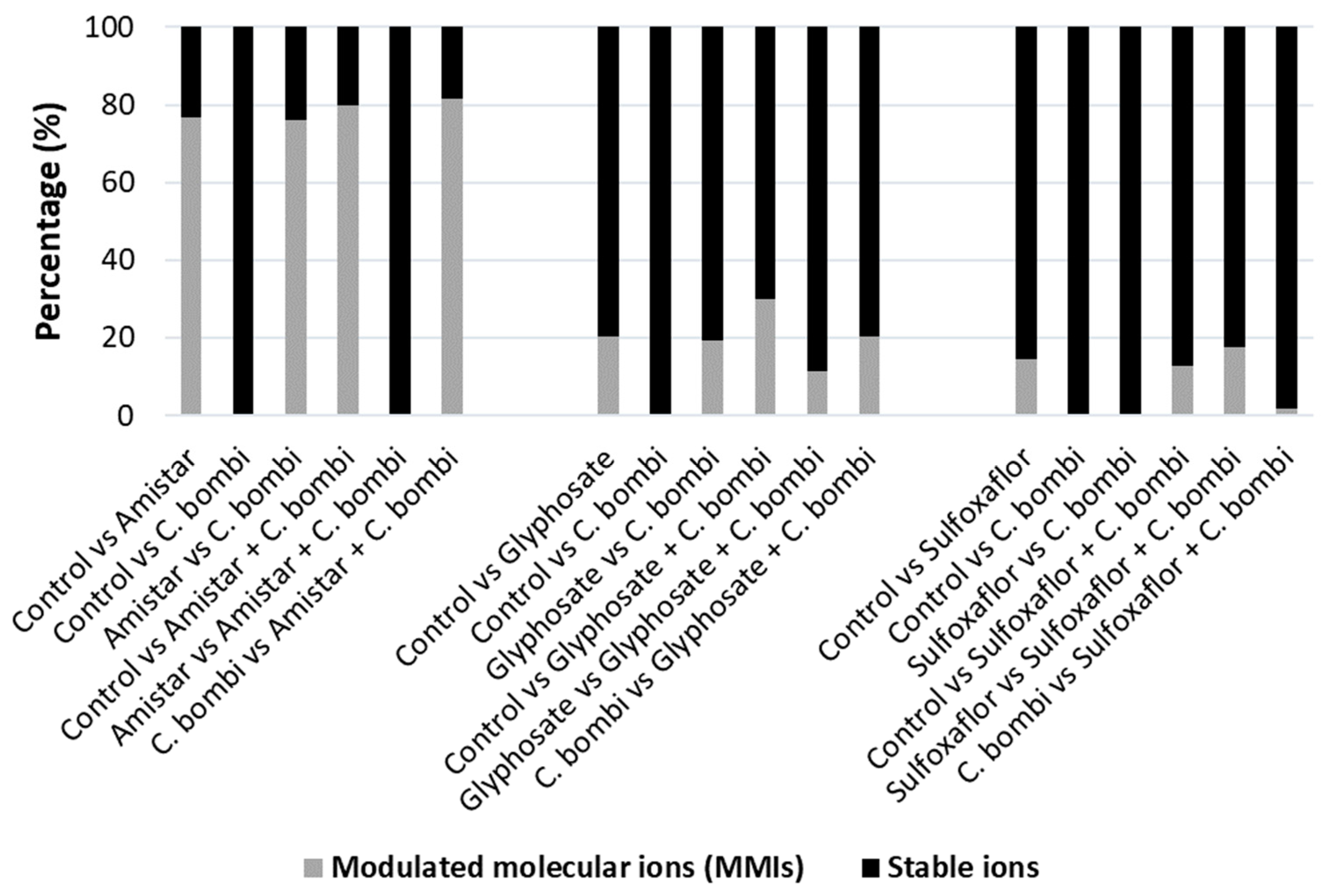
2.3. Modulated Molecular Ions (MMIs) Following Amistar, Sulfoxaflor, and Glyphosate Exposure and Co-Infection with C. bombi
2.4. Variation in Three Bee Immune Peptides—Apidaecin, Abaecin, and Chymotrypsin Inhibitor—Following Pesticide Exposure
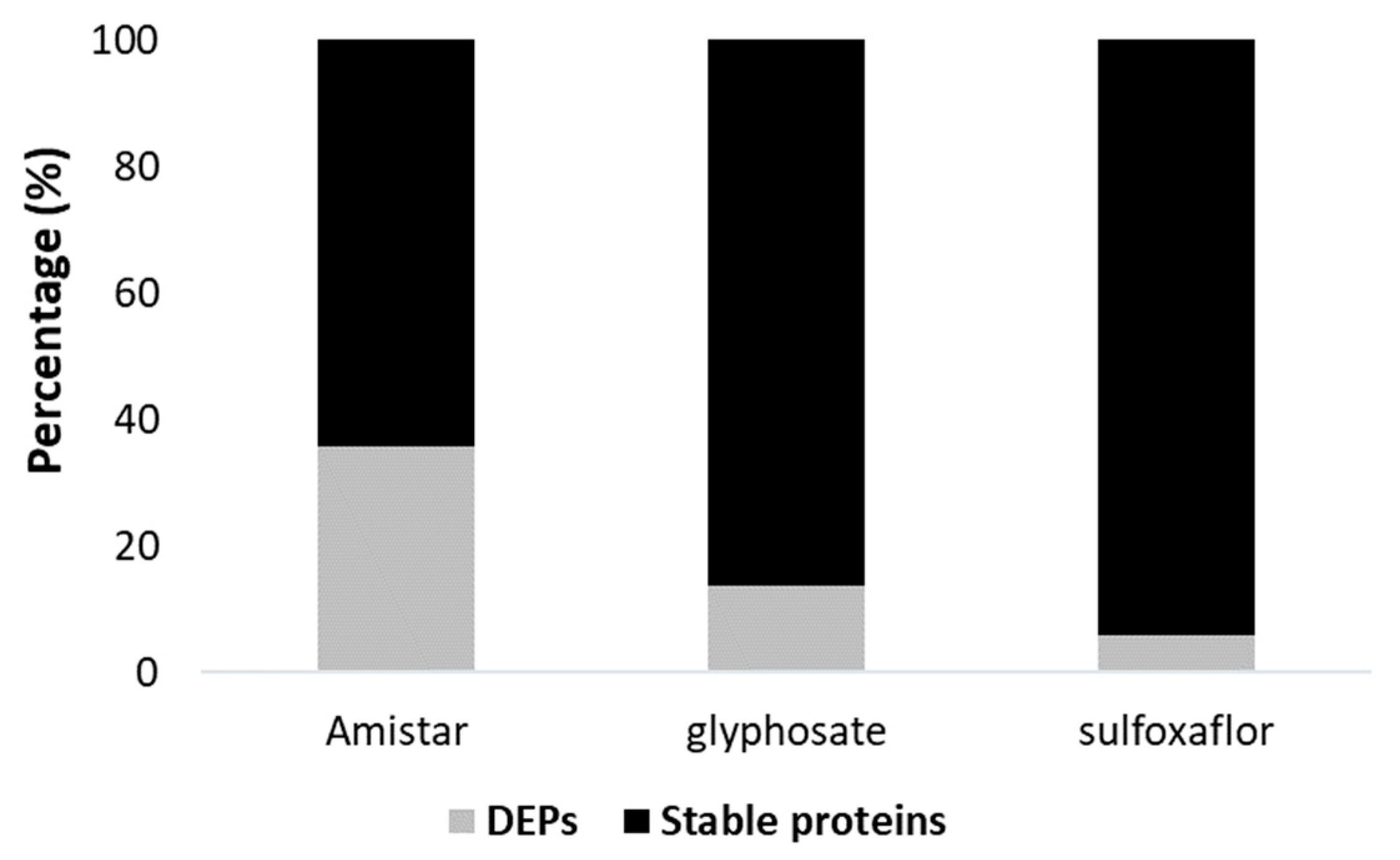
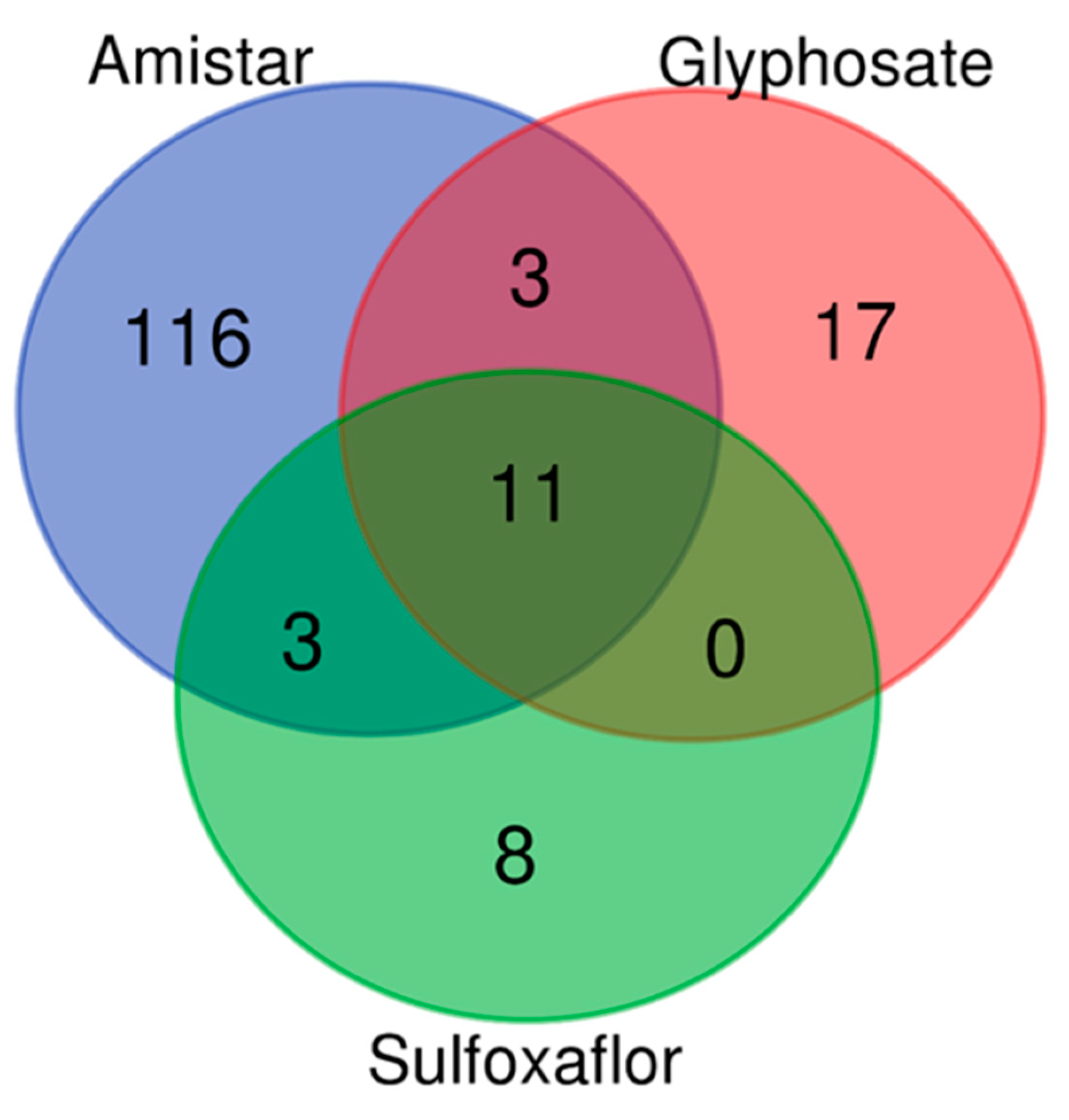
2.5. Protein Quantity Variations Following Pesticide and Parasite Exposure Demonstrated by Differential Bottom-Up Proteomics
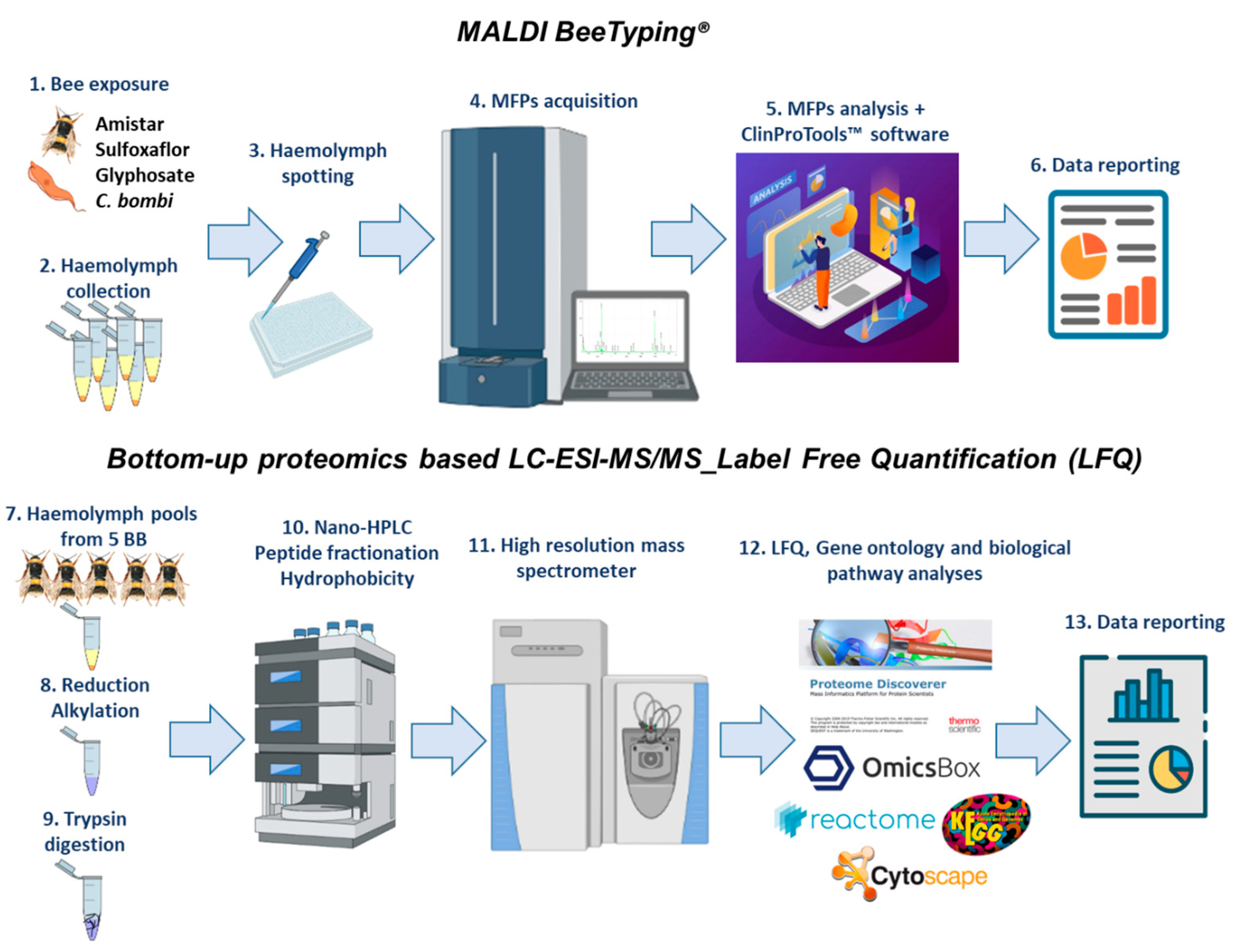
3. Material and Methods
3.1. Bees
3.2. Pesticides
3.3. Parasites
3.4. Exposure
3.5. Metrics
3.6. Haemolymph Extraction
3.7. Batches
3.8. Haemolymph Analyses: Chemicals and Reagents
3.9. Haemolymph Preparation for MALDI Molecular Mass Fingerprint (MFP)
3.10. MALDI BeeTyping® Acquisition
3.11. Data Processing and Statistical Analyses
3.12. Bottom-Up Proteomics-Based Nano LC-MS/MS
3.13. Label-Free Quantification (LFQ)
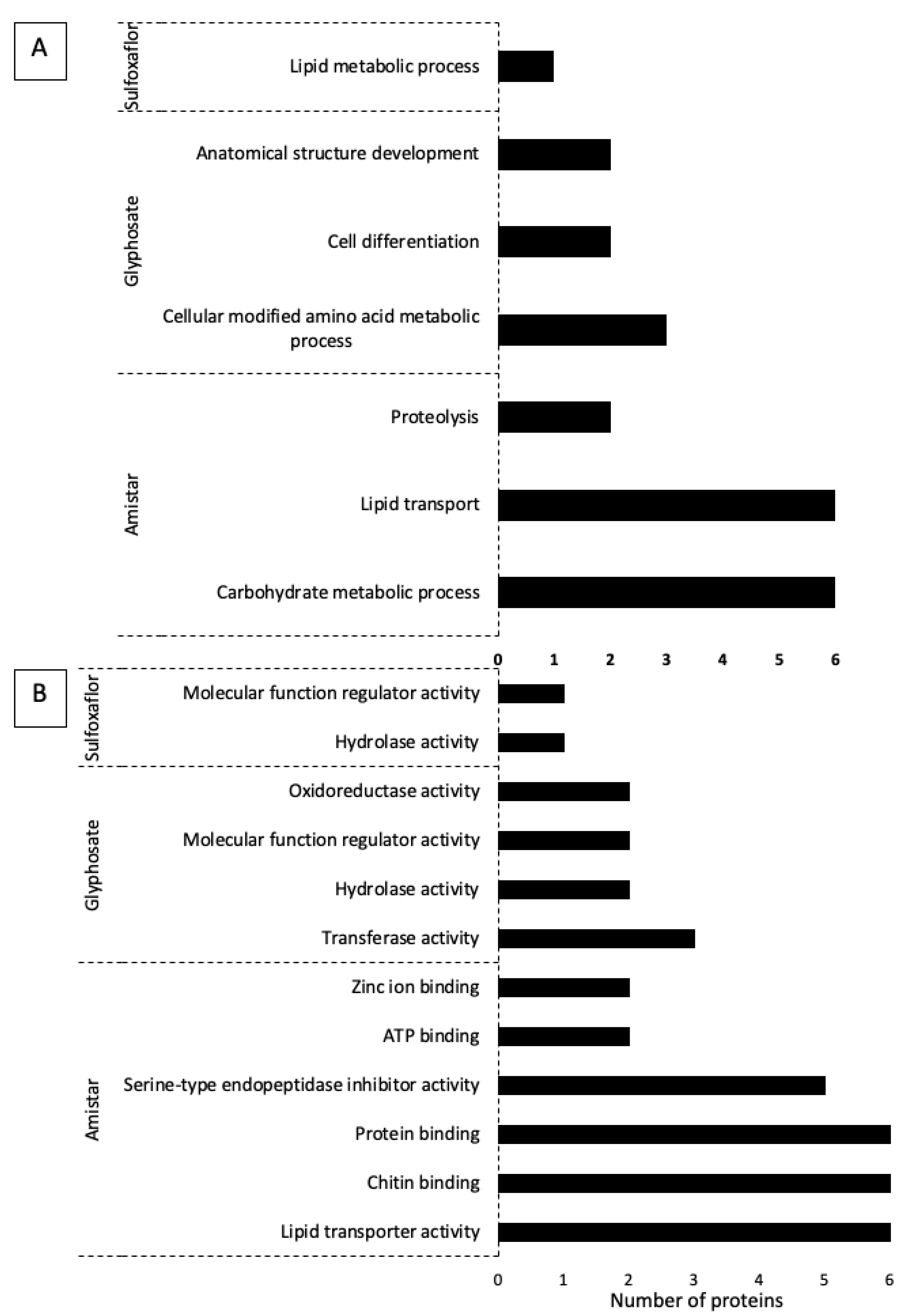
3.14. Functional Annotation: Gene Ontology and Pathways Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding Pollinators and Their Values to Human Well-Being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [Green Version]
- European Commission; Directorate General for the Environment; IUCN (International Union for Conservation of Nature). European Red List of Bees; Publications Office: Luxembourg, LU, USA, 2014.
- IPBES. The Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; Zenodo: Online, 2016. [Google Scholar]
- Hemberger, J.; Crossley, M.S.; Gratton, C. Historical Decrease in Agricultural Landscape Diversity Is Associated with Shifts in Bumble Bee Species Occurrence. Ecol. Lett. 2021, 24, 1800–1813. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed Coating with a Neonicotinoid Insecticide Negatively Affects Wild Bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef]
- McArt, S.H.; Fersch, A.A.; Milano, N.J.; Truitt, L.L.; Böröczky, K. High Pesticide Risk to Honey Bees despite Low Focal Crop Pollen Collection during Pollination of a Mass Blooming Crop. Sci. Rep. 2017, 7, 46554. [Google Scholar] [CrossRef] [Green Version]
- Woodcock, B.A.; Isaac, N.J.B.; Bullock, J.M.; Roy, D.B.; Garthwaite, D.G.; Crowe, A.; Pywell, R.F. Impacts of Neonicotinoid Use on Long-Term Population Changes in Wild Bees in England. Nat. Commun. 2016, 7, 12459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Straub, L.; Strobl, V.; Yañez, O.; Albrecht, M.; Brown, M.J.F.; Neumann, P. Do Pesticide and Pathogen Interactions Drive Wild Bee Declines? Int. J. Parasitol. Parasites Wildl. 2022, 18, 232–243. [Google Scholar] [CrossRef]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic Exposure to Neonicotinoids Reduces Honey Bee Health near Corn Crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tantillo, G.; Bottaro, M.; Di Pinto, A.; Martella, V.; Di Pinto, P.; Terio, V. Virus Infections of Honeybees Apis mellifera. Ital. J. Food Saf. 2015, 4, 5364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesovnik, T.; Cizelj, I.; Zorc, M.; Citar, M.; Bozic, J.; Glavan, G.; Narat, M. Immune Related Gene Expression in Worker Honey Bee (Apis mellifera carnica) Pupae Exposed to Neonicotinoid Thiamethoxam and Varroa Mites (Varroa Destructor). PLoS ONE 2017, 12, e0187079. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Baer, B. Consequences of a Short Time Exposure to a Sublethal Dose of Flupyradifurone (Sivanto) Pesticide Early in Life on Survival and Immunity in the Honeybee (Apis mellifera). Sci. Rep. 2019, 9, 19753. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.D.; Fountain, M.T.; Brown, M.J.F. Varietal and Seasonal Differences in the Effects of Commercial Bumblebees on Fruit Quality in Strawberry Crops. Agric. Ecosyst. Environ. 2019, 281, 124–133. [Google Scholar] [CrossRef]
- Koziy, R.V.; Wood, S.C.; Kozii, I.V.; van Rensburg, C.J.; Moshynskyy, I.; Dvylyuk, I.; Simko, E. Deformed Wing Virus Infection in Honey Bees (Apis mellifera L.). Vet. Pathol. 2019, 56, 636–641. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; Ahmed, H.R.; El-Wahed, A.A.A.; Saeed, A.; Algethami, A.F.; Attia, N.F.; Guo, Z.; Musharraf, S.G.; Khatib, A.; Alsharif, S.M.; et al. Bee Stressors from an Immunological Perspective and Strategies to Improve Bee Health. Vet. Sci. 2022, 9, 199. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, B.; Wu, H.; Cao, L.; Zhang, Y.; Li, W.; Han, R. Immune Response and Hemolymph Microbiota of Apis mellifera and Apis cerana After the Challenge With Recombinant Varroa Toxic Protein. J. Econ. Entomol. 2021, 114, 1310–1320. [Google Scholar] [CrossRef] [PubMed]
- Le Conte, Y.; Navajas, M. Climate Change: Impact on Honey Bee Populations and Diseases. Rev. Sci. Tech. Int. Off. Epizoot. 2008, 27, 485–497, 499–510. [Google Scholar] [CrossRef]
- Grassl, J.; Holt, S.; Cremen, N.; Peso, M.; Hahne, D.; Baer, B. Synergistic Effects of Pathogen and Pesticide Exposure on Honey Bee (Apis mellifera) Survival and Immunity. J. Invertebr. Pathol. 2018, 159, 78–86. [Google Scholar] [CrossRef]
- Havard, T.; Laurent, M.; Chauzat, M.-P. Impact of Stressors on Honey Bees (Apis mellifera; Hymenoptera: Apidae): Some Guidance for Research Emerge from a Meta-Analysis. Diversity 2019, 12, 7. [Google Scholar] [CrossRef] [Green Version]
- Breda, D.; Frizzera, D.; Giordano, G.; Seffin, E.; Zanni, V.; Annoscia, D.; Topping, C.J.; Blanchini, F.; Nazzi, F. A Deeper Understanding of System Interactions Can Explain Contradictory Field Results on Pesticide Impact on Honey Bees. Nat. Commun. 2022, 13, 5720. [Google Scholar] [CrossRef] [PubMed]
- Schöning, C.; Gisder, S.; Geiselhardt, S.; Kretschmann, I.; Bienefeld, K.; Hilker, M.; Genersch, E. Evidence for Damage-Dependent Hygienic Behaviour towards Varroa Destructor -Parasitised Brood in the Western Honey Bee, Apis mellifera. J. Exp. Biol. 2012, 215, 264–271. [Google Scholar] [CrossRef] [Green Version]
- Houdelet, C.; Sinpoo, C.; Chantaphanwattana, T.; Voisin, S.N.; Bocquet, M.; Chantawannakul, P.; Bulet, P. Proteomics of Anatomical Sections of the Gut of Nosema -Infected Western Honeybee (Apis mellifera) Reveals Different Early Responses to Nosema Spp. Isolates. J. Proteome Res. 2021, 20, 804–817. [Google Scholar] [CrossRef]
- Vázquez, D.E.; Ilina, N.; Pagano, E.A.; Zavala, J.A.; Farina, W.M. Glyphosate Affects the Larval Development of Honey Bees Depending on the Susceptibility of Colonies. PLoS ONE 2018, 13, e0205074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicewicz, Ł.; Nicewicz, A.W.; Kafel, A.; Nakonieczny, M. Set of Stress Biomarkers as a Practical Tool in the Assessment of Multistress Effect Using Honeybees from Urban and Rural Areas as a Model Organism: A Pilot Study. Environ. Sci. Pollut. Res. 2021, 28, 9084–9096. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, D.E.; Latorre-Estivalis, J.M.; Ons, S.; Farina, W.M. Chronic Exposure to Glyphosate Induces Transcriptional Changes in Honey Bee Larva: A Toxicogenomic Study. Environ. Pollut. 2020, 261, 114148. [Google Scholar] [CrossRef]
- Guo, Y.; Zhang, Z.; Zhuang, M.; Wang, L.; Li, K.; Yao, J.; Yang, H.; Huang, J.; Hao, Y.; Ying, F.; et al. Transcriptome Profiling Reveals a Novel Mechanism of Antiviral Immunity Upon Sacbrood Virus Infection in Honey Bee Larvae (Apis cerana). Front. Microbiol. 2021, 12, 615893. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Powell, J.E.; Moran, N.A. Glyphosate Induces Immune Dysregulation in Honey Bees. Anim. Microbiome 2022, 4, 16. [Google Scholar] [CrossRef]
- Haas, J.; Hayward, A.; Buer, B.; Maiwald, F.; Nebelsiek, B.; Glaubitz, J.; Bass, C.; Nauen, R. Phylogenomic and Functional Characterization of an Evolutionary Conserved Cytochrome P450-Based Insecticide Detoxification Mechanism in Bees. Proc. Natl. Acad. Sci. USA 2022, 119, e2205850119. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.M.; Mao, W.; Pollock, H.S.; Niu, G.; Schuler, M.A.; Berenbaum, M.R. Ecologically Appropriate Xenobiotics Induce Cytochrome P450s in Apis mellifera. PLoS ONE 2012, 7, e31051. [Google Scholar] [CrossRef]
- Hu, Y.-T.; Wu, T.-C.; Yang, E.-C.; Wu, P.-C.; Lin, P.-T.; Wu, Y.-L. Regulation of Genes Related to Immune Signaling and Detoxification in Apis mellifera by an Inhibitor of Histone Deacetylation. Sci. Rep. 2017, 7, 41255. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Li, G.; Guo, D.; Wang, Y.; Liu, Q.; Gao, Z.; Wang, H.; Liu, Z.; Guo, X.; Xu, B. Transcriptomic and Metabolomic Landscape of the Molecular Effects of Glyphosate Commercial Formulation on Apis mellifera ligustica and Apis cerana cerana. Sci. Total Environ. 2020, 744, 140819. [Google Scholar] [CrossRef]
- Słowińska, M.; Nynca, J.; Bąk, B.; Wilde, J.; Siuda, M.; Ciereszko, A. 2D-DIGE Proteomic Analysis Reveals Changes in Haemolymph Proteome of 1-Day-Old Honey Bee (Apis mellifera) Workers in Response to Infection with Varroa Destructor Mites. Apidologie 2019, 50, 632–656. [Google Scholar] [CrossRef] [Green Version]
- Zaluski, R.; Bittarello, A.C.; Vieira, J.C.S.; Braga, C.P.; Padilha, P.d.M.; Fernandes, M.d.S.; Bovi, T.d.S.; Orsi, R.d.O. Modification of the Head Proteome of Nurse Honeybees (Apis mellifera) Exposed to Field-Relevant Doses of Pesticides. Sci. Rep. 2020, 10, 2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roat, T.C.; dos Santos-Pinto, J.R.A.; dos Santos, L.D.; Santos, K.S.; Malaspina, O.; Palma, M.S. Modification of the Brain Proteome of Africanized Honeybees (Apis mellifera) Exposed to a Sub-lethal Doses of the Insecticide Fipronil. Ecotoxicology 2014, 23, 1659–1670. [Google Scholar] [CrossRef] [PubMed]
- Lundin, O.; Rundlöf, M.; Smith, H.G.; Fries, I.; Bommarco, R. Neonicotinoid Insecticides and Their Impacts on Bees: A Systematic Review of Research Approaches and Identification of Knowledge Gaps. PLoS ONE 2015, 10, e0136928. [Google Scholar] [CrossRef] [Green Version]
- Yordanova, M.; Evison, S.E.F.; Gill, R.J.; Graystock, P. The Threat of Pesticide and Disease Co-Exposure to Managed and Wild Bee Larvae. Int. J. Parasitol. Parasites Wildl. 2022, 17, 319–326. [Google Scholar] [CrossRef]
- Cullen, M.G.; Thompson, L.J.; Carolan, J.C.; Stout, J.C.; Stanley, D.A. Fungicides, Herbicides and Bees: A Systematic Review of Existing Research and Methods. PLoS ONE 2019, 14, e0225743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michener, C.D. The Bees of the World; JHU Press: Baltimore, MD, USA, 2000; ISBN 978-0-8018-6133-8. [Google Scholar]
- Dafni, A.; Kevan, P.; Gross, C.L.; Goka, K. Bombus terrestris, Pollinator, Invasive and Pest: An Assessment of Problems Associated with Its Widespread Introductions for Commercial Purposes. Appl. Entomol. Zool. 2010, 45, 101–113. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef] [PubMed]
- Shykoff, J.A.; Schmid-Hempel, P. Incidence and Effects of Four Parasites in Natural Populations of Bumble Bees in Switzerland. Apidologie 1991, 22, 117–125. [Google Scholar] [CrossRef]
- Korner, P.; Schmid-Hempel, P. Correlates of Parasite Load in Bumblebees in an Alpine Habitat. Entomol. Sci. 2005, 8, 151–160. [Google Scholar] [CrossRef]
- Rutrecht, S.T.; Brown, M.J. The Life-History Impact and Implications of Multiple Parasites for Bumble Bee Queens. Int. J. Parasitol. 2008, 38, 799–808. [Google Scholar] [CrossRef]
- Jones, C.M.; Brown, M.J.F. Parasites and Genetic Diversity in an Invasive Bumblebee. J. Anim. Ecol. 2014, 83, 1428–1440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siviter, H.; Scott, A.; Pasquier, G.; Pull, C.D.; Brown, M.J.F.; Leadbeater, E. No Evidence for Negative Impacts of Acute Sulfoxaflor Exposure on Bee Olfactory Conditioning or Working Memory. PeerJ 2019, 7, e7208. [Google Scholar] [CrossRef] [Green Version]
- Siviter, H.; Brown, M.J.F.; Leadbeater, E. Sulfoxaflor Exposure Reduces Bumblebee Reproductive Success. Nature 2018, 561, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, O.P.; Straw, E.A.; Linguadoca, A.; Brown, M.J.F. No Effect of Dual Exposure to Sulfoxaflor and a Trypanosome Parasite on Bumblebee Olfactory Learning. Sci. Rep. 2022, 12, 8611. [Google Scholar] [CrossRef] [PubMed]
- Straw, E.A.; Brown, M.J.F. Co-Formulant in a Commercial Fungicide Product Causes Lethal and Sub-Lethal Effects in Bumble Bees. Sci. Rep. 2021, 11, 21653. [Google Scholar] [CrossRef]
- Duke, S.O.; Powles, S.B. Glyphosate: A Once-in-a-Century Herbicide: Glyphosate: A Once-in-a-Century Herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef]
- Benbrook, C.M. Trends in Glyphosate Herbicide Use in the United States and Globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef] [Green Version]
- Battisti, L.; Potrich, M.; Sampaio, A.R.; de Castilhos Ghisi, N.; Costa-Maia, F.M.; Abati, R.; Dos Reis Martinez, C.B.; Sofia, S.H. Is Glyphosate Toxic to Bees? A Meta-Analytical Review. Sci. Total Environ. 2021, 767, 145397. [Google Scholar] [CrossRef]
- Straw, E.A.; Brown, M.J.F. No Evidence of Effects or Interaction between the Widely Used Herbicide, Glyphosate, and a Common Parasite in Bumble Bees. PeerJ 2021, 9, e12486. [Google Scholar] [CrossRef]
- Motta, E.V.S.; Raymann, K.; Moran, N.A. Glyphosate Perturbs the Gut Microbiota of Honey Bees. Proc. Natl. Acad. Sci. USA 2018, 115, 10305–10310. [Google Scholar] [CrossRef] [Green Version]
- Thompson, H.M.; Levine, S.L.; Doering, J.; Norman, S.; Manson, P.; Sutton, P.; von Mérey, G. Evaluating Exposure and Potential Effects on Honeybee Brood (Apis mellifera) Development Using Glyphosate as an Example. Integr. Environ. Assess. Manag. 2014, 10, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Odemer, R.; Rosenkranz, P. Chronic Exposure to a Neonicotinoid Pesticide and a Synthetic Pyrethroid in Full-Sized Honey Bee Colonies. J. Apic. Res. 2020, 59, 2–11. [Google Scholar] [CrossRef]
- Almasri, H.; Tavares, D.A.; Pioz, M.; Sené, D.; Tchamitchian, S.; Cousin, M.; Brunet, J.-L.; Belzunces, L.P. Mixtures of an Insecticide, a Fungicide and a Herbicide Induce High Toxicities and Systemic Physiological Disturbances in Winter Apis mellifera Honey Bees. Ecotoxicol. Environ. Saf. 2020, 203, 111013. [Google Scholar] [CrossRef]
- Brown, M.J.F.; Loosli, R.; Schmid-Hempel, P. Condition-Dependent Expression of Virulence in a Trypanosome Infecting Bumblebees. Oikos 2000, 91, 421–427. [Google Scholar] [CrossRef]
- Brown, M.J.; Moret, Y.; Schmid-Hempel, P. Activation of Host Constitutive Immune Defence by an Intestinal Trypanosome Parasite of Bumble Bees. Parasitology 2003, 126, 253–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gegear, R.J.; Otterstatter, M.C.; Thomson, J.D. Bumble-Bee Foragers Infected by a Gut Parasite Have an Impaired Ability to Utilize Floral Information. Proc. R. Soc. B Biol. Sci. 2006, 273, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Otterstatter, M.C.; Thomson, J.D. Contact Networks and Transmission of an Intestinal Pathogen in Bumble Bee (Bombus impatiens) Colonies. Oecologia 2007, 154, 411–421. [Google Scholar] [CrossRef]
- Otterstatter, M.C.; Thomson, J.D. Within-Host Dynamics of an Intestinal Pathogen of Bumble Bees. Parasitology 2006, 133, 749–761. [Google Scholar] [CrossRef]
- Yourth, C.P.; Brown, M.J.F.; Schmid-Hempel, P. Effects of Natal and Novel Crithidia bombi (Trypanosomatidae) Infections on Bombus terrestris Hosts. Insectes Sociaux 2008, 55, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Arafah, K.; Voisin, S.N.; Masson, V.; Alaux, C.; Le Conte, Y.; Bocquet, M.; Bulet, P. MALDI–MS Profiling to Address Honey Bee Health Status under Bacterial Challenge through Computational Modeling. Proteomics 2019, 19, 1900268. [Google Scholar] [CrossRef]
- Houdelet, C.; Bocquet, M.; Bulet, P. Matrix-assisted Laser Desorption/Ionization Mass Spectrometry Biotyping, an Approach for Deciphering and Assessing the Identity of the Honeybee Pathogen Nosema. Rapid Commun. Mass Spectrom. 2021, 35, e8980. [Google Scholar] [CrossRef]
- Ober, C.; Chupp, G.L. The Chitinase and Chitinase-like Proteins: A Review of Genetic and Functional Studies in Asthma and Immune-Mediated Diseases. Curr. Opin. Allergy Clin. Immunol. 2009, 9, 401–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, D.Q.D.; Winzerling, J.J. Insect Ferritins: Typical or Atypical? Biochim. Biophys. Acta BBA-Gen. Subj. 2010, 1800, 824–833. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.H.; Hu, Y.; Xing, L.S.; Jiang, H.; Hu, S.N.; Raikhel, A.S.; Zou, Z. A Critical Role for CLSP2 in the Modulation of Antifungal Immune Response in Mosquitoes. PLoS Pathog. 2015, 11, e1004931. [Google Scholar] [CrossRef] [Green Version]
- Park, H.G.; Lee, K.S.; Kim, B.Y.; Yoon, H.J.; Choi, Y.S.; Lee, K.Y.; Wan, H.; Li, J.; Jin, B.R. Honeybee (Apis cerana) Vitellogenin Acts as an Antimicrobial and Antioxidant Agent in the Body and Venom. Dev. Comp. Immunol. 2018, 85, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zhou, W.; Huang, B.; Gao, M.; Li, Q.; Tao, Y.; Wang, Z. Molecular and Functional Characterization of Peptidoglycan Recognition Proteins OfPGRP-A and OfPGRP-B in Ostrinia furnacalis (Lepidoptera: Crambidae). Insects 2022, 13, 417. [Google Scholar] [CrossRef]
- Kodrík, D.; Ibrahim, E.; Gautam, U.K.; Frydrychová, R.Č.; Bednářová, A.; Krištůfek, V.; Jedlička, P. Changes in Vitellogenin Expression Caused by Nematodal and Fungal Infections in Insects. J. Exp. Biol. 2019, 222, jeb.202853. [Google Scholar] [CrossRef] [Green Version]
- Eichelberger, K.R.; Goldman, W.E. Manipulating Neutrophil Degranulation as a Bacterial Virulence Strategy. PLOS Pathog. 2020, 16, e1009054. [Google Scholar] [CrossRef]
- El-Benna, J.; Hurtado-Nedelec, M.; Gougerot-Pocidalo, M.-A.; Dang, P.M.-C. Effects of Venoms on Neutrophil Respiratory Burst: A Major Inflammatory Function. J. Venom. Anim. Toxins Trop. Dis. 2021, 27, e20200179. [Google Scholar] [CrossRef]
- Gierlikowska, B.; Stachura, A.; Gierlikowski, W.; Demkow, U. Phagocytosis, Degranulation and Extracellular Traps Release by Neutrophils—The Current Knowledge, Pharmacological Modulation and Future Prospects. Front. Pharmacol. 2021, 12, 666732. [Google Scholar] [CrossRef]
- Baer, B.; Schmid-Hempel, P. Experimental Variation in Polyandry Affects Parasite Loads and Fitness in a Bumble-Bee. Nature 1999, 397, 151–154. [Google Scholar] [CrossRef]
- Ruiz-González, M.X.; Kelly, M.; Moret, Y.; Brown, M.J.F. Parasite Resistance and Immunity across Female Castes in a Social Insect. Behav. Ecol. Sociobiol. 2022, 76, 56. [Google Scholar] [CrossRef]
- OECD. Test No. 247: Bumblebee, Acute Oral Toxicity Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Berlin, Germany, 2017; ISBN 978-92-64-28412-8. [Google Scholar]
- Masson, V.; Arafah, K.; Voisin, S.; Bulet, P. Comparative Proteomics Studies of Insect Cuticle by Tandem Mass Spectrometry: Application of a Novel Proteomics Approach to the Pea Aphid Cuticular Proteins. Proteomics 2018, 18, 1700368. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pairwise Comparison | Apidaecin | Abaecin | Chymotrypsin Inhibitor |
|---|---|---|---|
| Control vs. Amistar | 277 | 216 | 47 |
| Control vs. C. bombi | 104 | NA | 134 |
| Amistar vs. C. bombi | 38 | 38 | 280 |
| Control vs. Amistar + C. bombi | 295 | 238 | 29 |
| Amistar vs. Amistar + C. bombi | 107 | 110 | 60 |
| C. bombi vs. Amistar + C. bombi | 283 | 289 | 21 |
| Control vs. glyphosate | 111 | 67 | 123 |
| Control vs. C. bombi | 108 | 98 | 119 |
| Glyphosate vs. C. bombi | 97 | 149 | 97 |
| Control vs. glyphosate + C. bombi | 125 | 65 | 120 |
| Glyphosate vs. glyphosate + C. bombi | 112 | 96 | 98 |
| C. bombi vs. glyphosate + C. bombi | 115 | 65 | 101 |
| Control vs. sulfoxaflor | 98 | 116 | 123 |
| Sulfoxaflor vs. C. bombi | 111 | 86 | 96 |
| Control vs. sulfoxaflor + C. bombi | 125 | 90 | 105 |
| Sulfoxaflor vs. sulfoxaflor + C. bombi | 128 | 78 | 85 |
| C. bombi vs. sulfoxaflor + C. bombi | 116 | 92 | 89 |
| Condition | Pathway | Protein Number |
|---|---|---|
| Amistar | Neutrophil degranulation | 13 |
| Digestion of dietary carbohydrate | 5 | |
| Platelet degranulation | 5 | |
| Amoebiasis | 5 | |
| Common pathway of fibrin clot formation | 4 | |
| Glucocorticoid biosynthesis | 4 | |
| ECM proteoglycans | 4 | |
| COPII-mediated vesicle transport | 4 | |
| Cargo concentration in the ER | 4 | |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | 4 | |
| Intrinsic pathway of fibrin clot formation | 4 | |
| Post-translational protein phosphorylation | 4 | |
| mRNA splicing—major pathway | 2 | |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | 2 | |
| VLDL assembly | 2 | |
| Glyphosate | Neutrophil degranulation | 3 |
| Arginine and proline metabolism | 3 | |
| Post-translational protein phosphorylation | 2 | |
| Platelet degranulation | 2 | |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | 2 | |
| Creatine metabolism | 1 | |
| Intrinsic pathway of fibrin clot formation | 1 | |
| Apoptotic cleavage of cell adhesion proteins | 1 | |
| Common pathway of fibrin clot formation | 1 | |
| Integrin cell surface interactions | 1 | |
| Creatine metabolism | 1 | |
| VEGFR2-mediated vascular permeability | 1 | |
| Adherens junctions interactions | 1 | |
| Glucocorticoid biosynthesis | 1 | |
| Antimicrobial peptides | 1 | |
| Sulfoxaflor | COPII-mediated vesicle transport | 1 |
| Defective C1GALT1C1 causes TNPS | 1 | |
| Glycosphingolipid metabolism | 1 | |
| Post-translational protein phosphorylation | 1 | |
| Platelet degranulation | 1 | |
| Intrinsic pathway of fibrin clot formation | 1 | |
| Termination of O-glycan biosynthesis | 1 | |
| Defective GALNT12 causes CRCS1 | 1 | |
| Common pathway of fibrin clot formation | 1 | |
| Glucocorticoid biosynthesis | 1 | |
| Dectin-2 family | 1 | |
| Association of TriC/CCT with target proteins during biosynthesis | 1 | |
| Cargo concentration in the ER | 1 | |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | 1 | |
| Defective GALNT3 causes HFTC | 1 |
| Control (Ctrl) (n = 31, 30) | Amistar Only 200 μg (Equal to 0.8 µL of Pure Amistar) (n = 29, 16) | Dimethoate (Ctrl+)4 μg (n = 29, 0) |
| C. bombi only 10,000 cells (n = 29, 28) | Amistar and C. bombi 200 μg (equal to 0.8 µL of pure Amistar) 10,000 cells (n = 32, 26) |
| Control (Ctrl) (n = 46, 45) | Glyphosate 200 μg (n = 41, 40) | Sulfoxaflor 0.06 μg (n = 36, 32) | Dimethoate (Ctrl+) 4 μg (n = 36, 0) |
| C. bombi 10,000 cells (n = 36, 31) | Glyphosate and C. bombi 200 μg 10,000 cells (n = 35, 35) | Sulfoxaflor and C. bombi 0.06 μg 10,000 cells (n = 44, 39) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Askri, D.; Straw, E.A.; Arafah, K.; Voisin, S.N.; Bocquet, M.; Brown, M.J.F.; Bulet, P. Parasite and Pesticide Impacts on the Bumblebee (Bombus terrestris) Haemolymph Proteome. Int. J. Mol. Sci. 2023, 24, 5384. https://doi.org/10.3390/ijms24065384
Askri D, Straw EA, Arafah K, Voisin SN, Bocquet M, Brown MJF, Bulet P. Parasite and Pesticide Impacts on the Bumblebee (Bombus terrestris) Haemolymph Proteome. International Journal of Molecular Sciences. 2023; 24(6):5384. https://doi.org/10.3390/ijms24065384
Chicago/Turabian StyleAskri, Dalel, Edward A. Straw, Karim Arafah, Sébastien N. Voisin, Michel Bocquet, Mark J. F. Brown, and Philippe Bulet. 2023. "Parasite and Pesticide Impacts on the Bumblebee (Bombus terrestris) Haemolymph Proteome" International Journal of Molecular Sciences 24, no. 6: 5384. https://doi.org/10.3390/ijms24065384