

The Impact of Aerobic Exercise on HDL Quantity and Quality: A Narrative Review

Abstract
:1. Introduction
2. Structure and Functions of HDL Cholesterol
3. Aerobic Exercise
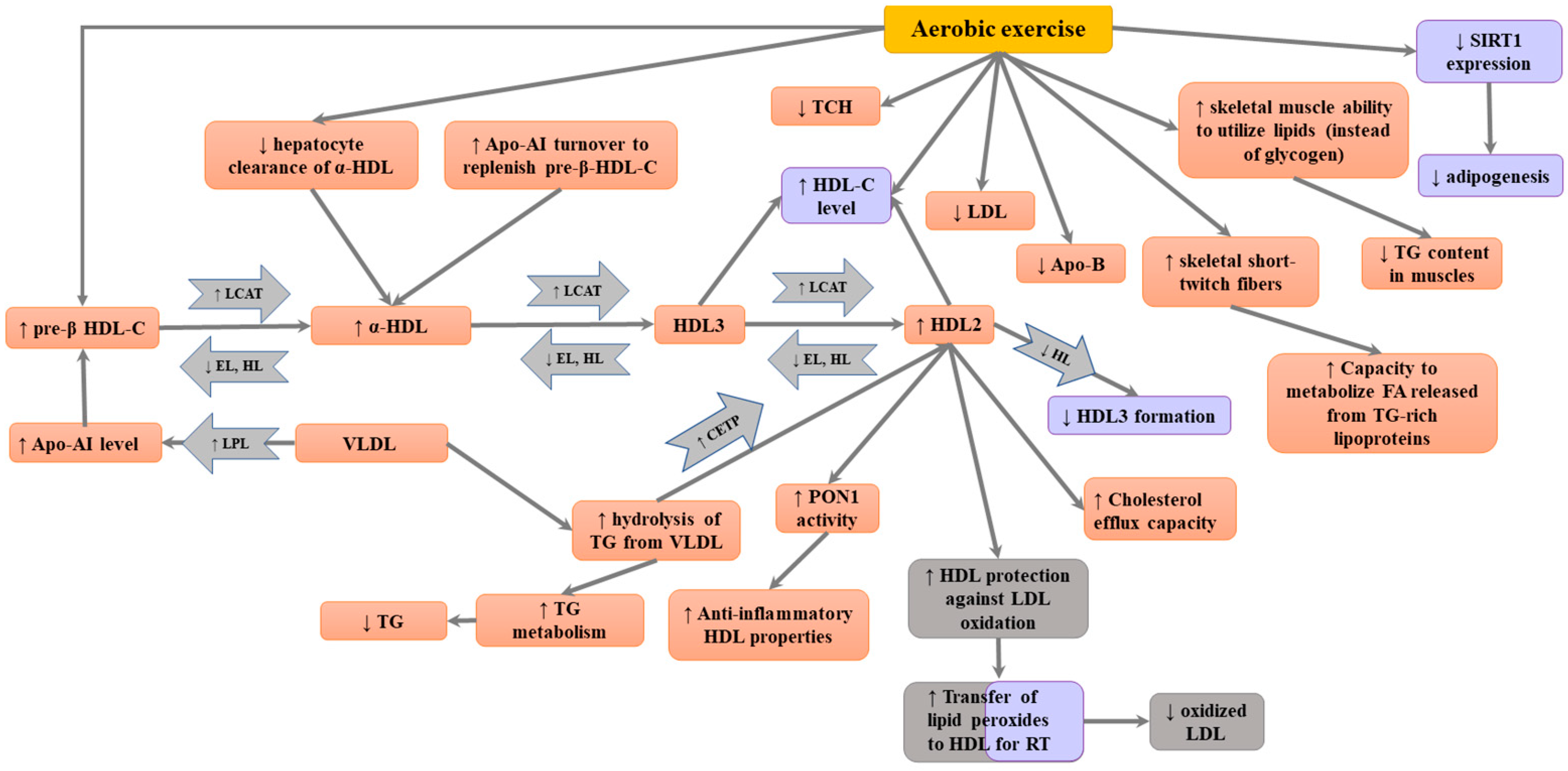
4. Impact of Aerobic Exercise on HDL Levels
5. Impact of Aerobic Exercise on HDL Composition and Function
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sirtori, C.R.; Corsini, A.; Ruscica, M. The Role of High-Density Lipoprotein Cholesterol in 2022. Curr. Atheroscler. Rep. 2022, 24, 365–377. [Google Scholar] [CrossRef]
- Cho, K.H. The Current Status of Research on High-Density Lipoproteins (HDL): A Paradigm Shift from HDL Quantity to HDL Quality and HDL Functionality. Int. J. Mol. Sci. 2022, 23, 3967. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H.; Park, H.-J.; Kim, S.-J.; Kim, J.-R. Decrease in HDL-C is associated with age and household income in adults from the Korean National Health and Nutrition Examination Survey 2017: Correlation analysis of low HDL-C and poverty. Int. J. Environ. Res. Public Health 2019, 16, 3329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.-H.; Park, H.-J.; Kim, J.-R. Decrease in serum HDL-C level is associated with elevation of blood pressure: Correlation analysis from the Korean National Health and nutrition examination survey 2017. Int. J. Environ. Res. Public Health 2020, 17, 1101. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Chen, P. The Effects of Different Exercise Modalities in the Treatment of Cardiometabolic Risk Factors in Obese Adolescents with Sedentary Behavior—A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Children 2021, 8, 1062. [Google Scholar] [CrossRef]
- Julian, V.; Bergsten, P.; Forslund, A.; Ahlstrom, H.; Ciba, I.; Dahlbom, M.; Furthner, D.; Gomahr, J.; Kullberg, J.; Maruszczak, K. Sedentary time has a stronger impact on metabolic health than moderate to vigorous physical activity in adolescents with obesity: A cross-sectional analysis of the Beta-JUDO study. Pediatr. Obes. 2022, 17, e12897. [Google Scholar] [CrossRef] [PubMed]
- Wing, R.R.; Greeno, C.G. Behavioral and psychosocial aspects of obesity and its treatment. Bailleres Clin Endocrinol Metab. 1994, 8, 689–703. [Google Scholar] [CrossRef]
- Goldbourt, U.; Yaari, S.; Medalie, J.H. Isolated low HDL cholesterol as a risk factor for coronary heart disease mortality: A 21-year follow-up of 8000 men. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couillard, C.; Després, J.-P.; Lamarche, B.; Bergeron, J.; Gagnon, J.; Leon, A.S.; Rao, D.C.; Skinner, J.S.; Wilmore, J.H.; Bouchard, C. Effects of Endurance Exercise Training on Plasma HDL Cholesterol Levels Depend on Levels of Triglycerides. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1226–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirtori, C.; Ruscica, M.; Calabresi, L.; Chiesa, G.; Giovannoni, R.; Badimon, J. HDL therapy today: From atherosclerosis, to stent compatibility to heart failure. Ann. Med. 2019, 51, 345–359. [Google Scholar] [CrossRef]
- Bartlett, J.; Predazzi, I.M.; Williams, S.M.; Bush, W.S.; Kim, Y.; Havas, S.; Toth, P.P.; Fazio, S.; Miller, M. Is Isolated Low High-Density Lipoprotein Cholesterol a Cardiovascular Disease Risk Factor? New Insights From the Framingham Offspring Study. Circ. Cardiovasc. Qual. Outcomes 2016, 9, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Gotto, A.M. Low High-Density Lipoprotein Cholesterol as a Risk Factor in Coronary Heart Disease. Circulation 2001, 103, 2213–2218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boden, W.; Probstfield, J.; Anderson, T.; Chaitman, B.; Desvignes-Nickens, P.; Koprowicz, K.; McBride, R.; Teo, K.; Weintraub, W. Niacin in patients with low HDL cholesterol levels receiving intensive statin therapy. N. Engl. J. Med. 2011, 365, 2255. [Google Scholar] [PubMed] [Green Version]
- Barter, P.J.; Caulfield, M.; Eriksson, M.; Grundy, S.M.; Kastelein, J.J.; Komajda, M.; Lopez-Sendon, J.; Mosca, L.; Tardif, J.-C.; Waters, D.D. Effects of torcetrapib in patients at high risk for coronary events. N. Engl. J. Med. 2007, 357, 2109–2122. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, G.G.; Olsson, A.G.; Ballantyne, C.M.; Barter, P.J.; Holme, I.M.; Kallend, D.; Leiter, L.A.; Leitersdorf, E.; McMurray, J.J.; Shah, P.K. Rationale and design of the dal-OUTCOMES trial: Efficacy and safety of dalcetrapib in patients with recent acute coronary syndrome. Am. Heart J. 2009, 158, 896–901.e893. [Google Scholar] [CrossRef]
- Ruiz-Ramie, J.J.; Barber, J.L.; Sarzynski, M.A. Effects of exercise on HDL functionality. Curr. Opin. Lipidol. 2019, 30, 16–23. [Google Scholar] [CrossRef]
- Kodama, S.; Tanaka, S.; Saito, K.; Shu, M.; Sone, Y.; Onitake, F.; Suzuki, E.; Shimano, H.; Yamamoto, S.; Kondo, K. Effect of aerobic exercise training on serum levels of high-density lipoprotein cholesterol: A meta-analysis. Arch. Intern. Med. 2007, 167, 999–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durstine, J.L.; Grandjean, P.W.; Davis, P.G.; Ferguson, M.A.; Alderson, N.L.; DuBose, K.D. Blood lipid and lipoprotein adaptations to exercise. Sport. Med. 2001, 31, 1033–1062. [Google Scholar] [CrossRef] [PubMed]
- Camont, L.; Lhomme, M.; Rached, F.; Le Goff, W.; Nègre-Salvayre, A.; Salvayre, R.; Calzada, C.; Lagarde, M.; Chapman, M.J.; Kontush, A. Small, dense high-density lipoprotein-3 particles are enriched in negatively charged phospholipids: Relevance to cellular cholesterol efflux, antioxidative, antithrombotic, anti-inflammatory, and antiapoptotic functionalities. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2715–2723. [Google Scholar] [CrossRef] [Green Version]
- Rosenson, R.S.; Brewer, H.B., Jr.; Chapman, M.J.; Fazio, S.; Hussain, M.M.; Kontush, A.; Krauss, R.M.; Otvos, J.D.; Remaley, A.T.; Schaefer, E.J. HDL measures, particle heterogeneity, proposed nomenclature, and relation to atherosclerotic cardiovascular events. Clin. Chem. 2011, 57, 392–410. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.T.; Krauss, R.M.; Vranizan, K.M.; Stefanick, M.L.; Wood, P.; Lindgren, F. Associations of lipoproteins and apolipoproteins with gradient gel electrophoresis estimates of high density lipoprotein subfractions in men and women. Arterioscler. Thromb. J. Vasc. Biol. 1992, 12, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, N.P.; Martin, S.E.; Crouse, S.F. Acute exercise and training alter blood lipid and lipoprotein profiles differently in overweight and obese men and women. Obesity 2012, 20, 1618–1627. [Google Scholar] [CrossRef] [PubMed]
- Cuchel, M.; Rader, D.J. Macrophage reverse cholesterol transport: Key to the regression of atherosclerosis? Circulation 2006, 113, 2548–2555. [Google Scholar] [CrossRef] [PubMed]
- Woudberg, N.J.; Mendham, A.E.; Katz, A.A.; Goedecke, J.H.; Lecour, S. Exercise intervention alters HDL subclass distribution and function in obese women. Lipids Health Dis. 2018, 17, 232. [Google Scholar] [CrossRef] [Green Version]
- Hafiane, A.; Gianopoulos, I.; Sorci-Thomas, M.G.; Daskalopoulou, S.S. Current models of apolipoprotein AI lipidation by adenosine triphosphate binding cassette transporter A1. Curr. Opin. Lipidol. 2022, 33, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Pownall, H.J.; Morrisett, J.D.; Sparrow, J.T.; Smith, L.C.; Shepherd, J.; Jackson, R.L.; Gotto, A.M. A review of the unique features of HDL apoproteins. Lipids 1979, 14, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovich, O.; Ozerova, I.; Olfer’ev, A.; Serdyuk, A.; Metel’skaya, V.; Perova, N. Association of serum apolipoprotein A-II concentration with combined hyperlipidemia and impaired glucose tolerance. Bull. Exp. Biol. Med. 2006, 141, 678–681. [Google Scholar] [CrossRef]
- Ribas, V.; Sánchez-Quesada, J.L.; Antón, R.; Camacho, M.; Julve, J.; Escolà-Gil, J.C.; Vila, L.s.; Ordóñez-Llanos, J.; Blanco-Vaca, F. Human apolipoprotein A-II enrichment displaces paraoxonase from HDL and impairs its antioxidant properties: A new mechanism linking HDL protein composition and antiatherogenic potential. Circ. Res. 2004, 95, 789–797. [Google Scholar] [CrossRef]
- de Beer, M.C.; Durbin, D.M.; Cai, L.; Mirocha, N.; Jonas, A.; Webb, N.R.; de Beer, F.C.; van der Westhuyzen, D.R. Apolipoprotein A-II modulates the binding and selective lipid uptake of reconstituted high density lipoprotein by scavenger receptor BI. J. Biol. Chem. 2001, 276, 15832–15839. [Google Scholar] [CrossRef] [Green Version]
- Park, K.-H.; Shin, D.-G.; Kim, J.-R.; Cho, K.-H. Senescence-related truncation and multimerization of apolipoprotein AI in high-density lipoprotein with an elevated level of advanced glycated end products and cholesteryl ester transfer activity. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2010, 65, 600–610. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-H. Synthesis of reconstituted high density lipoprotein (rHDL) containing apoA-I and apoC-III: The functional role of apoC-III in rHDL. Mol. Cells 2009, 27, 291–297. [Google Scholar] [CrossRef]
- Park, K.-H.; Shin, D.-G.; Cho, K.-H. Dysfunctional lipoproteins from young smokers exacerbate cellular senescence and atherogenesis with smaller particle size and severe oxidation and glycation. Toxicol. Sci. 2014, 140, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zewinger, S.; Reiser, J.; Jankowski, V.; Alansary, D.; Hahm, E.; Triem, S.; Klug, M.; Schunk, S.J.; Schmit, D.; Kramann, R. Apolipoprotein C3 induces inflammation and organ damage by alternative inflammasome activation. Nat. Immunol. 2020, 21, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.; Wang, X.; Noffsinger, V.P.; Jennings, D.; de Beer, M.C.; de Beer, F.C.; Tannock, L.R.; Webb, N.R. Serum amyloid A is not incorporated into HDL during HDL biogenesis. J. Lipid Res. 2020, 61, 328–337. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.-H.; de Beer, M.C.; Wroblewski, J.M.; Webb, N.R.; de Beer, F.C. SAA does not induce cytokine production in physiological conditions. Cytokine 2013, 61, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, P.G.; Thompson, J.C.; Webb, N.R.; de Beer, F.C.; King, V.L.; Tannock, L.R. Serum amyloid A, but not C-reactive protein, stimulates vascular proteoglycan synthesis in a pro-atherogenic manner. Am. J. Pathol. 2008, 173, 1902–1910. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Wu, T.; Qin, W.; An, C.; Wang, Z.; Zhang, M.; Zhang, Y.; Zhang, C.; An, F. Serum amyloid A directly accelerates the progression of atherosclerosis in apolipoprotein E-deficient mice. Mol. Med. 2011, 17, 1357–1364. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, S.D.; Shim, J.W.; Lee, S.Y.; Lee, H.; Cho, K.-H.; Yun, J.; Bae, Y.-S. Serum amyloid A induces CCL2 production via formyl peptide receptor-like 1-mediated signaling in human monocytes. J. Immunol. 2008, 181, 4332–4339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sponder, M.; Kopecky, C.; Campean, I.-A.; Emich, M.; Fritzer-Szekeres, M.; Litschauer, B.; Graf, S.; Säemann, M.D.; Strametz-Juranek, J. Sports and HDL-Quality Reflected By Serum Amyloid A and Surfactant Protein, B. Int. J. Med. Sci. 2017, 14, 1040–1048. [Google Scholar] [CrossRef] [Green Version]
- Webb, N.R. High-density lipoproteins and serum amyloid A (SAA). Curr. Atheroscler. Rep. 2021, 23, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Dullaart, R.P.; de Boer, J.F.; Annema, W.; Tietge, U.J. The inverse relation of HDL anti-oxidative functionality with serum amyloid A is lost in metabolic syndrome subjects. Obesity 2013, 21, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Nofer, J.-R.; Kehrel, B.; Fobker, M.; Levkau, B.; Assmann, G.; von Eckardstein, A. HDL and arteriosclerosis: Beyond reverse cholesterol transport. Atherosclerosis 2002, 161, 1–16. [Google Scholar] [CrossRef]
- Barter, P.; Rye, K.-A. High density lipoproteins and coronary heart disease. Atherosclerosis 1996, 121, 1–12. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, J.-R.; Lee, I.-C.; Kwon, H.-J. Native high-density lipoproteins (HDL) with higher paraoxonase exerts a potent antiviral effect against SARS-CoV-2 (COVID-19), while glycated HDL lost the antiviral activity. Antioxidants 2021, 10, 209. [Google Scholar] [CrossRef]
- Chen, C.; Khismatullin, D.B. Oxidized low-density lipoprotein contributes to atherogenesis via co-activation of macrophages and mast cells. PLoS ONE 2015, 10, e0123088. [Google Scholar] [CrossRef]
- Holvoet, P.; Keyzer, D.D.; Jacobs, D., Jr. Oxidized LDL and the metabolic syndrome. Future Lipidol. 2008, 3, 637–649. [Google Scholar] [CrossRef] [PubMed]
- Soran, H.; Schofield, J.D.; Durrington, P.N. Antioxidant properties of HDL. Front. Pharm. 2015, 6, 222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahotupa, M.; Suomela, J.-P.; Vuorimaa, T.; Vasankari, T. Lipoprotein-specific transport of circulating lipid peroxides. Ann. Med. 2010, 42, 521–529. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.A.; Hutchinson, R.; Morrisett, J.; Boerwinkle, E.; Davis, C.E.; Gotto, A.M., Jr.; Patsch, W. Plasma lipid, lipoprotein cholesterol, and apoprotein distributions in selected US communities. The Atherosclerosis Risk in Communities (ARIC) Study. Arter. Thromb. 1993, 13, 1139–1158. [Google Scholar] [CrossRef] [PubMed]
- Pascot, A.; Lemieux, I.; Bergeron, J.; Tremblay, A.; Nadeau, A.; Prud’homme, D.; Couillard, C.; Lamarche, B.; Després, J.P. HDL particle size: A marker of the gender difference in the metabolic risk profile. Atherosclerosis 2002, 160, 399–406. [Google Scholar] [CrossRef]
- Li, Z.; McNamara, J.R.; Fruchart, J.C.; Luc, G.; Bard, J.M.; Ordovas, J.M.; Wilson, P.W.; Schaefer, E.J. Effects of gender and menopausal status on plasma lipoprotein subspecies and particle sizes. J. Lipid. Res. 1996, 37, 1886–1896. [Google Scholar] [CrossRef]
- Brinton, E.A.; Eisenberg, S.; Breslow, J.L. Human HDL cholesterol levels are determined by apoA-I fractional catabolic rate, which correlates inversely with estimates of HDL particle size. Effects of gender, hepatic and lipoprotein lipases, triglyceride and insulin levels, and body fat distribution. Arter. Thromb. 1994, 14, 707–720. [Google Scholar] [CrossRef] [Green Version]
- Brinton, E.A.; Eisenberg, S.; Breslow, J.L. Elevated high density lipoprotein cholesterol levels correlate with decreased apolipoprotein A-I and A-II fractional catabolic rate in women. J. Clin. Investig. 1989, 84, 262–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, G.A.; Pineda-Torra, I.; Ciurtin, C.; Jury, E.C. Sex Differences in Lipid Metabolism: Implications for Systemic Lupus Erythematosus and Cardiovascular Disease Risk. Front. Med. 2022, 9, 1492. [Google Scholar] [CrossRef] [PubMed]
- Stadler, J.T.; Lackner, S.; Mörkl, S.; Trakaki, A.; Scharnagl, H.; Borenich, A.; Wonisch, W.; Mangge, H.; Zelzer, S.; Meier-Allard, N.; et al. Obesity Affects HDL Metabolism, Composition and Subclass Distribution. Biomedicines 2021, 9, 242. [Google Scholar] [CrossRef]
- Franssen, R.; Monajemi, H.; Stroes, E.S.; Kastelein, J.J. Obesity and dyslipidemia. Med. Clin. N. Am. 2011, 95, 893–902. [Google Scholar] [CrossRef]
- Radeau, T.; Lau, P.; Robb, M.; McDonnell, M.; Ailhaud, G.; McPherson, R. Cholesteryl ester transfer protein (CETP) mRNA abundance in human adipose tissue: Relationship to cell size and membrane cholesterol content. J. Lipid. Res. 1995, 36, 2552–2561. [Google Scholar] [CrossRef]
- Ebenbichler, C.F.; Laimer, M.; Kaser, S.; Ritsch, A.; Sandhofer, A.; Weiss, H.; Aigner, F.; Patsch, J.R. Relationship between cholesteryl ester transfer protein and atherogenic lipoprotein profile in morbidly obese women. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1465–1469. [Google Scholar] [CrossRef]
- Kappelle, P.J.; de Boer, J.F.; Perton, F.G.; Annema, W.; de Vries, R.; Dullaart, R.P.; Tietge, U.J. Increased LCAT activity and hyperglycaemia decrease the antioxidative functionality of HDL. Eur. J. Clin. Investig. 2012, 42, 487–495. [Google Scholar] [CrossRef]
- Baragetti, A.; Ossoli, A.; Strazzella, A.; Simonelli, S.; Baragetti, I.; Grigore, L.; Pellegatta, F.; Catapano, A.L.; Norata, G.D.; Calabresi, L. Low Plasma Lecithin: Cholesterol Acyltransferase (LCAT) Concentration Predicts Chronic Kidney Disease. J. Clin. Med. 2020, 9, 2289. [Google Scholar] [CrossRef]
- Kaiser, T.; Kinny-Köster, B.; Bartels, M.; Berg, T.; Scholz, M.; Engelmann, C.; Seehofer, D.; Becker, S.; Ceglarek, U.; Thiery, J. Cholesterol esterification in plasma as a biomarker for liver function and prediction of mortality. BMC Gastroenterol. 2017, 17, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Darabi, M.; Tubeuf, E.; Canicio, A.; Lhomme, M.; Frisdal, E.; Lanfranchi-Lebreton, S.; Matheron, L.; Rached, F.; Ponnaiah, M.; et al. Free cholesterol transfer to high-density lipoprotein (HDL) upon triglyceride lipolysis underlies the U-shape relationship between HDL-cholesterol and cardiovascular disease. Eur. J. Prev. Cardiol. 2020, 27, 1606–1616. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Toth, P.P.; Kris-Etherton, P.M.; Abate, N.; Aronne, L.J.; Brown, W.V.; Gonzalez-Campoy, J.M.; Jones, S.R.; Kumar, R.; La Forge, R.; et al. Obesity, adiposity, and dyslipidemia: A consensus statement from the National Lipid Association. J. Clin. Lipidol. 2013, 7, 304–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, W.S.; Heink, A.; Sexmith, H.; Dolan, L.M.; Gordon, S.M.; Otvos, J.D.; Melchior, J.T.; Elder, D.A.; Khoury, J.; Geh, E.; et al. Obesity is associated with an altered HDL subspecies profile among adolescents with metabolic disease. J. Lipid. Res. 2017, 58, 1916–1923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davidson, W.S.; Inge, T.H.; Sexmith, H.; Heink, A.; Elder, D.; Hui, D.Y.; Melchior, J.T.; Kelesidis, T.; Shah, A.S. Weight loss surgery in adolescents corrects high-density lipoprotein subspecies and their function. Int. J. Obesity 2017, 41, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Osto, E.; Doytcheva, P.; Corteville, C.; Bueter, M.; Dörig, C.; Stivala, S.; Buhmann, H.; Colin, S.; Rohrer, L.; Hasballa, R.; et al. Rapid and body weight-independent improvement of endothelial and high-density lipoprotein function after Roux-en-Y gastric bypass: Role of glucagon-like peptide-1. Circulation 2015, 131, 871–881. [Google Scholar] [CrossRef] [Green Version]
- Nakhjavani, M.L.M.; Nargesi, A.A.; Mostafavi, E.; Mirmiranpour, H.; Esteghamati, A. Gender-related differences in HDL structure with the progression of microalbuminuria in patients with type 2 diabetes. J. Diabetes Metab. Disord. Control 2015, 2, 110–115. [Google Scholar] [CrossRef] [Green Version]
- Eriksson, M.; Zethelius, B.; Eeg-Olofsson, K.; Nilsson, P.M.; Gudbjörnsdottir, S.; Cederholm, J.; Eliasson, B. Blood lipids in 75,048 type 2 diabetic patients: A population-based survey from the Swedish National diabetes register. Eur. J. Cardiovasc. Prev. Rehabil. 2011, 18, 97–105. [Google Scholar] [CrossRef]
- Carnevale Schianca, G.P.; Fra, G.P.; Colli, E.; Bigliocca, M.; Mella, R.; Scaglia, E.; Bartoli, E. Sex differences in lipid profiles in relation to the progression of glucose abnormalities. J. Diabetes 2012, 4, 95–101. [Google Scholar] [CrossRef]
- Nakhjavani, M.; Morteza, A.; Karimi, R.; Banihashmi, Z.; Esteghamati, A. Diabetes induces gender gap on LCAT levels and activity. Life Sci. 2013, 92, 51–54. [Google Scholar] [CrossRef]
- Magge, S.N.; Prasad, D.; Koren, D.; Gallagher, P.R.; Mohler, E.R., 3rd; Stettler, N.; Levitt Katz, L.E.; Rader, D.J. Prediabetic obese adolescents have a more atherogenic lipoprotein profile compared with normoglycemic obese peers. J. Pediatr. 2012, 161, 881–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borggreve, S.E.; De Vries, R.; Dullaart, R.P. Alterations in high-density lipoprotein metabolism and reverse cholesterol transport in insulin resistance and type 2 diabetes mellitus: Role of lipolytic enzymes, lecithin:cholesterol acyltransferase and lipid transfer proteins. Eur. J. Clin. Investig. 2003, 33, 1051–1069. [Google Scholar] [CrossRef]
- Mendivil, C.O.; Furtado, J.; Morton, A.M.; Wang, L.; Sacks, F.M. Novel Pathways of Apolipoprotein A-I Metabolism in High-Density Lipoprotein of Different Sizes in Humans. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 156–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curioni, C.; Lourenco, P. Long-term weight loss after diet and exercise: A systematic review. Int. J. Obes. 2005, 29, 1168–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soci, U.P.R.; Melo, S.F.S.; Gomes, J.L.P.; Silveira, A.C.; Nóbrega, C.; Oliveira, E.M.d. Exercise training and epigenetic regulation: Multilevel modification and regulation of gene expression. Exerc. Cardiovasc. Dis. Prev. Treat. 2017, 1000, 281–322. [Google Scholar]
- Slentz, C.A.; Duscha, B.D.; Johnson, J.L.; Ketchum, K.; Aiken, L.B.; Samsa, G.P.; Houmard, J.A.; Bales, C.W.; Kraus, W.E. Effects of the amount of exercise on body weight, body composition, and measures of central obesity: STRRIDE—A randomized controlled study. Arch. Intern. Med. 2004, 164, 31–39. [Google Scholar] [CrossRef]
- Ohta, M.; Nanri, H.; Matsushima, Y.; Sato, Y.; Ikeda, M. Blood pressure-lowering effects of lifestyle modification: Possible involvement of nitric oxide bioavailability. Hypertens. Res. 2005, 28, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Muscella, A.; Stefàno, E.; Lunetti, P.; Capobianco, L.; Marsigliante, S. The Regulation of Fat Metabolism During Aerobic Exercise. Biomolecules 2020, 10, 1699. [Google Scholar] [CrossRef]
- Halabchi, F.; Alizadeh, Z.; Sahraian, M.A.; Abolhasani, M. Exercise prescription for patients with multiple sclerosis; potential benefits and practical recommendations. BMC Neurol. 2017, 17, 185. [Google Scholar] [CrossRef] [Green Version]
- Dowman, L.M.; McDonald, C.F.; Hill, C.J.; Lee, A.L.; Barker, K.; Boote, C.; Glaspole, I.; Goh, N.S.; Southcott, A.M.; Burge, A.T. The evidence of benefits of exercise training in interstitial lung disease: A randomised controlled trial. Thorax 2017, 72, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Tambosco, L.; Percebois-Macadré, L.; Rapin, A.; Nicomette-Bardel, J.; Boyer, F.-C. Effort training in Parkinson’s disease: A systematic review. Ann. Phys. Rehabil. Med. 2014, 57, 79–104. [Google Scholar] [CrossRef] [Green Version]
- US Department of Health and Human Services; Centers for Disease Control and Prevention; National Center for Chronic Disease Prevention and Health Promotion; The President’s Council on Physical Fitness and Sports. Physical Activity and Health: A Report of the Surgeon General; U.S. Department of Health and Human Services: Atlanta, GA, USA, 1996.
- Mann, S.; Beedie, C.; Jimenez, A. Differential Effects of Aerobic Exercise, Resistance Training and Combined Exercise Modalities on Cholesterol and the Lipid Profile: Review, Synthesis and Recommendations. Sport. Med. 2014, 44, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.; Qi, H.; He, X.; Lu, Y.; Yang, C.; Li, F.; Wang, L.; Qiang, D.; Shui, K.; Zhou, L.; et al. Aerobic Exercise for a Duration of 90 min or Longer Per Week may Reduce the Atherogenic Index of Plasma. Sci. Rep. 2018, 8, 1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, M.; Moyna, N.M.; Zderic, T.W.; O’Gorman, D.J.; McCaffrey, N.; Carson, B.P.; Hamilton, M.T. Lipoprotein particle distribution and skeletal muscle lipoprotein lipase activity after acute exercise. Lipids Health Dis. 2012, 11, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, S.; Zhong, J.; Sun, C.; Zhang, J. Effects of aerobic exercise on TC, HDL-C, LDL-C and TG in patients with hyperlipidemia: A protocol of systematic review and meta-analysis. Medicine 2021, 100, e25103. [Google Scholar] [CrossRef]
- Rothenbacher, D.; Koenig, W.; Brenner, H. Lifetime physical activity patterns and risk of coronary heart disease. Heart 2006, 92, 1319–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virani, S.S.; Alonso, A.; Aparicio, H.J.; Benjamin, E.J.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Cheng, S.; Delling, F.N.; et al. Heart Disease and Stroke Statistics—2021 update: A report from the American Heart Association. Circulation 2021, 143, e254–e743. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services. Physical Activity Guidelines for Americans, 2nd ed.; U.S. Department of Health and Human Services: Washington, DC, USA, 2018.
- Argani, N.; Sharifi, G.; Golshahi, J. Comparison of the effect of different intensity exercise on a bicycle ergometer on postprandial lipidemia in type II diabetic patients. ARYA Atheroscler. 2014, 10, 147. [Google Scholar] [PubMed]
- Sarzynski, M.A.; Burton, J.; Rankinen, T.; Blair, S.N.; Church, T.S.; Després, J.-P.; Hagberg, J.M.; Landers-Ramos, R.; Leon, A.S.; Mikus, C.R. The effects of exercise on the lipoprotein subclass profile: A meta-analysis of 10 interventions. Atherosclerosis 2015, 243, 364–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casella-Filho, A.; Chagas, A.C.; Maranhão, R.C.; Trombetta, I.C.; Cesena, F.H.; Silva, V.M.; Tanus-Santos, J.E.; Negrão, C.E.; da Luz, P.L. Effect of exercise training on plasma levels and functional properties of high-density lipoprotein cholesterol in the metabolic syndrome. Am. J. Cardiol. 2011, 107, 1168–1172. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, I.; Iborra, R.T.; Neves, M.; Lottenberg, S.A.; Charf, A.M.; Nunes, V.S.; Negrao, C.E.; Nakandakare, E.R.; Quintão, E.; Passarelli, M. HDL atheroprotection by aerobic exercise training in type 2 diabetes mellitus. Med. Sci. Sport. Exerc. 2008, 40, 779–786. [Google Scholar] [CrossRef]
- Ferguson, M.A.; Alderson, N.L.; Trost, S.G.; Essig, D.A.; Burke, J.R.; Durstine, J.L. Effects of four different single exercise sessions on lipids, lipoproteins, and lipoprotein lipase. J. Appl. Physiol. 1998, 85, 1169–1174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cobbold, C. Battle of the sexes: Which is better for you, high-or low-intensity exercise? J. Sport Health Sci. 2018, 7, 429. [Google Scholar] [CrossRef] [PubMed]
- Harms, C.A. Does gender affect pulmonary function and exercise capacity? Respir. Physiol. Neurobiol. 2006, 151, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, N.; Tomkinson, G.; Ekelund, U. Aerobic fitness and its relationship to sport, exercise training and habitual physical activity during youth. Br. J. Sports Med. 2011, 45, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Despres, J.P.; Pouliot, M.C.; Moorjani, S.; Nadeau, A.; Tremblay, A.; Lupien, P.; Theriault, G.; Bouchard, C. Loss of abdominal fat and metabolic response to exercise training in obese women. Am. J. Physiol.-Endocrinol. Metab. 1991, 261, E159–E167. [Google Scholar] [CrossRef] [PubMed]
- Hardman, A. Physical activity, obesity and blood lipids. Int. J. Obes. 1999, 23, S64–S71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Folsom, A.R.; Arnett, D.K.; Hutchinson, R.G.; Liao, F.; Clegg, L.X.; Cooper, L.S. Physical activity and incidence of coronary heart disease in middle-aged women and men. Med. Sci. Sport. Exerc. 1997, 29, 901–909. [Google Scholar] [CrossRef]
- Aadahl, M.; von Huth Smith, L.; Pisinger, C.; Toft, U.N.; Glümer, C.; Borch-Johnsen, K.; Jørgensen, T. Five-year change in physical activity is associated with changes in cardiovascular disease risk factors: The Inter99 study. Prev. Med. 2009, 48, 326–331. [Google Scholar] [CrossRef]
- Zmuda, J.M.; Yurgalevitch, S.M.; Flynn, M.M.; Bausserman, L.L.; Saratelli, A.; Spannaus-Martin, D.J.; Herbert, P.N.; Thompson, P.D. Exercise training has little effect on HDL levels and metabolism in men with initially low HDL cholesterol. Atherosclerosis 1998, 137, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.T.; Stefanick, M.L.; Vranizan, K.M.; Wood, P.D. The effects of weight loss by exercise or by dieting on plasma high-density lipoprotein (HDL) levels in men with low, intermediate, and normal-to-high HDL at baseline. Metabolism 1994, 43, 917–924. [Google Scholar] [CrossRef] [Green Version]
- Välimäki, I.A.; Vuorimaa, T.; Ahotupa, M.; Vasankari, T.J. Strenuous physical exercise accelerates the lipid peroxide clearing transport by HDL. Eur. J. Appl. Physiol. 2016, 116, 1683–1691. [Google Scholar] [CrossRef]
- Lira, F.S.; Zanchi, N.E.; Lima-Silva, A.E.; Pires, F.O.; Bertuzzi, R.C.; Caperuto, E.C.; Kiss, M.A.; Seelaender, M.; Santos, R.V. Is acute supramaximal exercise capable of modulating lipoprotein profile in healthy men? Eur. J. Clin. Investig. 2010, 40, 759–765. [Google Scholar] [CrossRef]
- Leaf, D.A. The effect of physical exercise on reverse cholesterol transport. Metabolism 2003, 52, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Naharudin, M.N.; Yusof, A. The effect of 10 days of energy-deficit diet and high-intensity exercise training on the plasma high-density-lipoprotein (HDL) level among healthy collegiate males. Eur. J. Sport Sci. 2022, 22, 826–835. [Google Scholar] [CrossRef] [PubMed]
- Lira, F.S.; Zanchi, N.E.; Lima-Silva, A.E.; Pires, F.O.; Bertuzzi, R.C.; Santos, R.V.; Caperuto, E.C.; Kiss, M.A.; Seelaender, M. Acute high-intensity exercise with low energy expenditure reduced LDL-c and total cholesterol in men. Eur. J. Appl. Physiol. 2009, 107, 203–210. [Google Scholar] [CrossRef]
- Antero Kesaniemi, Y.; Danforth, E.; Jensen, M.D.; Kopelman, P.G.; Lefèbvre, P.; Reeder, B.A. Dose-response issues concerning physical activity and health: An evidence-based symposium. Med. Sci. Sport. Exerc. 2001, 33, S351–S358. [Google Scholar] [CrossRef] [PubMed]
- Banz, W.J.; Maher, M.A.; Thompson, W.G.; Bassett, D.R.; Moore, W.; Ashraf, M.; Keefer, D.J.; Zemel, M.B. Effects of resistance versus aerobic training on coronary artery disease risk factors. Exp. Biol. Med. 2003, 228, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, Y.; Nogami, Y. Response of Lipids and Lipoproteins to Regular Aquatic Endurance Exercise: A Meta-Analysis of Randomized Controlled Trials. J. Atheroscler. Thromb. 2019, 26, 14–30. [Google Scholar] [CrossRef] [Green Version]
- Nybo, L.; Sundstrup, E.; Jakobsen, M.D.; Mohr, M.; Hornstrup, T.; Simonsen, L.; Bülow, J.; Randers, M.B.; Nielsen, J.J.; Aagaard, P.; et al. High-intensity training versus traditional exercise interventions for promoting health. Med. Sci. Sports Exerc. 2010, 42, 1951–1958. [Google Scholar] [CrossRef] [Green Version]
- Wood, G.; Murrell, A.; Van Der Touw, T.; Smart, N. HIIT is not superior to MICT in altering blood lipids: A systematic review and meta-analysis. BMJ Open Sport Exerc. Med. 2019, 5, e000647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, A.C.; Haskell, W.L.; Young, D.R.; Oka, R.K.; Stefanick, M.L. Long-term effects of varying intensities and formats of physical activity on participation rates, fitness, and lipoproteins in men and women aged 50 to 65 years. Circulation 1995, 91, 2596–2604. [Google Scholar] [CrossRef]
- Bouaziz, W.; Vogel, T.; Schmitt, E.; Kaltenbach, G.; Geny, B.; Lang, P.O. Health benefits of aerobic training programs in adults aged 70 and over: A systematic review. Arch. Gerontol. Geriatr. 2017, 69, 110–127. [Google Scholar] [CrossRef] [PubMed]
- Berglund, I.; Vesterbekkmo, E.K.; Retterstøl, K.; Anderssen, S.A.; Fiatarone Singh, M.A.; Helge, J.W.; Lydersen, S.; Wisløff, U.; Stensvold, D. The Long-term Effect of Different Exercise Intensities on High-Density Lipoprotein Cholesterol in Older Men and Women Using the Per Protocol Approach: The Generation 100 Study. Mayo Clin. Proc. Innov. Qual. Outcomes 2021, 5, 859–871. [Google Scholar] [CrossRef]
- Dunn, A.L.; Marcus, B.H.; Kampert, J.B.; Garcia, M.E.; Kohl III, H.W.; Blair, S.N. Reduction in cardiovascular disease risk factors: 6-month results from ProjectActive. Prev. Med. 1997, 26, 883–892. [Google Scholar] [CrossRef] [PubMed]
- LeMura, L.M.; von Duvillard, S.P.; Andreacci, J.; Klebez, J.M.; Chelland, S.A.; Russo, J. Lipid and lipoprotein profiles, cardiovascular fitness, body composition, and diet during and after resistance, aerobic and combination training in young women. Eur. J. Appl. Physiol. 2000, 82, 451–458. [Google Scholar] [CrossRef]
- Kraus, W.E.; Houmard, J.A.; Duscha, B.D.; Knetzger, K.J.; Wharton, M.B.; McCartney, J.S.; Bales, C.W.; Henes, S.; Samsa, G.P.; Otvos, J.D. Effects of the amount and intensity of exercise on plasma lipoproteins. N. Engl. J. Med. 2002, 347, 1483–1492. [Google Scholar] [CrossRef]
- Leon, A.; Gaskill, S.; Rice, T.; Bergeron, J.; Gagnon, J.; Rao, D.; Skinner, J.; Wilmore, J.; Bouchard, C. Variability in the response of HDL cholesterol to exercise training in the HERITAGE Family Study. Int. J. Sport. Med. 2002, 23, 1–9. [Google Scholar] [CrossRef]
- Siri-Tarino, P.W. Effects of diet on high-density lipoprotein cholesterol. Curr. Atheroscler. Rep. 2011, 13, 453–460. [Google Scholar] [CrossRef]
- Volek, J.S.; Sharman, M.J.; Forsythe, C.E. Modification of lipoproteins by very low-carbohydrate diets. J. Nutr. 2005, 135, 1339–1342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Earnest, C.P.; Artero, E.G.; Sui, X.; Lee, D.-c.; Church, T.S.; Blair, S.N. Maximal estimated cardiorespiratory fitness, cardiometabolic risk factors, and metabolic syndrome in the aerobics center longitudinal study. Mayo Clin. Proc. 2013, 88, 259–270. [Google Scholar] [CrossRef] [Green Version]
- Calabresi, L.; Franceschini, G. Lecithin: Cholesterol acyltransferase, high-density lipoproteins, and atheroprotection in humans. Trends Cardiovasc. Med. 2010, 20, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Riedl, I.; Yoshioka, M.; Nishida, Y.; Tobina, T.; Paradis, R.; Shono, N.; Tanaka, H.; St-Amand, J. Regulation of skeletal muscle transcriptome in elderly men after 6 weeks of endurance training at lactate threshold intensity. Exp. Gerontol. 2010, 45, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Physical Activity Guidelines Advisory Committee report, 2008. To the Secretary of Health and Human Services. Part A: Executive summary. Nutr. Rev. 2009, 67, 114–120. [CrossRef]
- Vanhees, L.; Geladas, N.; Hansen, D.; Kouidi, E.; Niebauer, J.; Reiner, Ž.; Cornelissen, V.; Adamopoulos, S.; Prescott, E.; Börjesson, M. Importance of characteristics and modalities of physical activity and exercise in the management of cardiovascular health in individuals with cardiovascular risk factors: Recommendations from the EACPR (Part II). Eur. J. Prev. Cardiol. 2012, 19, 1005–1033. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-H. High-Density Lipoproteins as Biomarkers and Therapeutic Tools; Impacts of Lifestyle, Diseases, and Environmental Stressors on HDL; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1. [Google Scholar]
- Nye, E.R.; Carlson, K.; Kirstein, P.; Rössner, S. Changes in high density lipoprotein subfractions and other lipoproteins induced by exercise. Clin. Chim. Acta 1981, 113, 51–57. [Google Scholar] [CrossRef]
- Patsch, J.R.; Prasad, S.; Gotto, A.M., Jr.; Bengtsson-Olivecrona, G. Postprandial lipemia. A key for the conversion of high density lipoprotein2 into high density lipoprotein3 by hepatic lipase. J. Clin. Investig. 1984, 74, 2017–2023. [Google Scholar] [CrossRef] [Green Version]
- Tikkanen, H.O.; Näveri, H.; Härkönen, M. Skeletal muscle fiber distribution influences serum high-density lipoprotein cholesterol level. Atherosclerosis 1996, 120, 1–5. [Google Scholar] [CrossRef]
- Sarzynski, M.A.; Ruiz-Ramie, J.J.; Barber, J.L.; Slentz, C.A.; Apolzan, J.W.; McGarrah, R.W.; Harris, M.N.; Church, T.S.; Borja, M.S.; He, Y. Effects of increasing exercise intensity and dose on multiple measures of HDL (high-density lipoprotein) function. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 943–952. [Google Scholar] [CrossRef] [Green Version]
- Jafari, M.; Leaf, D.A.; MacRae, H.; Kasem, J.; O’Conner, P.; Pullinger, C.; Malloy, M.; Kane, J.P. The effects of physical exercise on plasma prebeta-1 high-density lipoprotein. Metabolism 2003, 52, 437–442. [Google Scholar] [CrossRef]
- Hennessy, L.K.; Kunitake, S.T.; Kane, J.P. Apolipoprotein A-I-containing lipoproteins, with or without apolipoprotein A-II, as progenitors of pre-beta high-density lipoprotein particles. Biochemistry 1993, 32, 5759–5765. [Google Scholar] [CrossRef] [PubMed]
- Frey, I.; Baumstark, M.W.; Berg, A.; Keul, J. Influence of acute maximal exercise on lecithin:cholesterol acyltransferase activity in healthy adults of differing aerobic performance. Eur. J. Appl. Physiol. Occup. Physiol. 1991, 62, 31–35. [Google Scholar] [CrossRef]
- Trejo-Gutierrez, J.F.; Fletcher, G. Impact of exercise on blood lipids and lipoproteins. J. Clin. Lipidol. 2007, 1, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Kuusi, T.; Saarinen, P.; Nikkilä, E.A. Evidence for the role of hepatic endothelial lipase in the metabolism of plasma high density lipoprotein2 in man. Atherosclerosis 1980, 36, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Thompson, P.D.; Cullinane, E.M.; Sady, S.P.; Flynn, M.M.; Chenevert, C.B.; Herbert, P.N. High density lipoprotein metabolism in endurance athletes and sedentary men. Circulation 1991, 84, 140–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiainen, S.; Luoto, R.; Ahotupa, M.; Raitanen, J.; Vasankari, T. 6-mo aerobic exercise intervention enhances the lipid peroxide transport function of HDL. Free. Radic. Res. 2016, 50, 1279–1285. [Google Scholar] [CrossRef]
- Lee, H.; Park, J.E.; Choi, I.; Cho, K.H. Enhanced functional and structural properties of high-density lipoproteins from runners and wrestlers compared to throwers and lifters. BMB Rep. 2009, 42, 605–610. [Google Scholar] [CrossRef] [Green Version]
- Sang, H.; Yao, S.; Zhang, L.; Li, X.; Yang, N.; Zhao, J.; Zhao, L.; Si, Y.; Zhang, Y.; Lv, X. Walk-run training improves the anti-inflammation properties of high-density lipoprotein in patients with metabolic syndrome. J. Clin. Endocrinol. Metab. 2015, 100, 870–879. [Google Scholar] [CrossRef] [Green Version]
- Roberts, C.K.; Ng, C.; Hama, S.; Eliseo, A.J.; Barnard, R.J. Effect of a short-term diet and exercise intervention on inflammatory/anti-inflammatory properties of HDL in overweight/obese men with cardiovascular risk factors. J. Appl. Physiol. 2006, 101, 1727–1732. [Google Scholar] [CrossRef] [Green Version]
- Woudberg, N.J.; Goedecke, J.H.; Blackhurst, D.; Frias, M.; James, R.; Opie, L.H.; Lecour, S. Association between ethnicity and obesity with high-density lipoprotein (HDL) function and subclass distribution. Lipids. Health Dis. 2016, 15, 92. [Google Scholar] [CrossRef] [Green Version]
- Cho, K.-H.; Nam, H.-S.; Kang, D.-J.; Zee, S.; Park, M.-H. Enhancement of High-Density Lipoprotein (HDL) Quantity and Quality by Regular and Habitual Exercise in Middle-Aged Women with Improvements in Lipid and Apolipoprotein Profiles: Larger Particle Size and Higher Antioxidant Ability of HDL. Int. J. Mol. Sci. 2023, 24, 1151. [Google Scholar] [CrossRef]
- Crouse, S.F.; O’Brien, B.C.; Grandjean, P.W.; Lowe, R.C.; Rohack, J.J.; Green, J.S.; Tolson, H. Training intensity, blood lipids, and apolipoproteins in men with high cholesterol. J. Appl. Physiol. 1997, 82, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Altena, T.S.; Michaelson, J.L.; Ball, S.D.; Thomas, T.R. Single sessions of intermittent and continuous exercise and postprandial lipemia. Med. Sci. Sport. Exerc. 2004, 36, 1364–1371. [Google Scholar] [CrossRef] [Green Version]
- Shah, A.S.; Tan, L.; Long, J.L.; Davidson, W.S. Proteomic diversity of high density lipoproteins: Our emerging understanding of its importance in lipid transport and beyond1. J. Lipid. Res. 2013, 54, 2575–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kontush, A.; Lhomme, M.; Chapman, M.J. Unraveling the complexities of the HDL lipidome1. J. Lipid Res. 2013, 54, 2950–2963. [Google Scholar] [CrossRef] [Green Version]
- Vinetti, G.; Mozzini, C.; Desenzani, P.; Boni, E.; Bulla, L.; Lorenzetti, I.; Romano, C.; Pasini, A.; Cominacini, L.; Assanelli, D. Supervised exercise training reduces oxidative stress and cardiometabolic risk in adults with type 2 diabetes: A randomized controlled trial. Sci. Rep. 2015, 5, 9238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Rowland, T.; Mattel, L.; Vanderburgh, P.; Manos, T.; Charkoudian, N. The influence of short-term aerobic training on blood lipids in healthy 10-12 year old children. Int. J. Sport. Med. 1996, 17, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.S.-C.; Xie, Y.J.; Woo, J.; Kwok, T.C.-Y. Effects of Tai Chi and walking exercises on weight loss, metabolic syndrome parameters, and bone mineral density: A cluster randomized controlled trial. Evid.-Based Complement. Altern. Med. 2015, 2015, 976123. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Type of Exercise | Studied Population | Intensity | Duration | Additional Benefits | Ref. |
|---|---|---|---|---|---|
| On a cycle ergometer (universal aerobicycle); | 200 healthy and sedentary white men (79 fathers and 121 sons) with baseline: (1) Low TG and high HDL cholesterol (normolipidemia); (2) Low TG and low HDL cholesterol (isolated low HDL cholesterol); (3) High TG and high HDL cholesterol (isolated high TGs); and (4) High TGs and low HDL cholesterol (high TG/low HDL cholesterol). | 2 weeks at an HR associated with 55% VO2max for 30 min increased every 2 weeks until the 14th week to HR associated with 75% of their initial VO2max for 50 min and maintained for the next 6 weeks. | 60 sessions within 21 weeks |
| [9] |
| Various | 4039 men and women (30–60 years) completed the study. | High-intensity intervention physical activity groups: <2 h/week, 2–3.9 h/week, 4–6.9 h/week and ≥7 h/week. | Follow-up: 5 years |
| [101] |
| Supervised endurance exercise training without weight loss Walking, jogging and stationary cycling | 17 men aged 26–49 years with: (1) initially low HDL-C levels (<40 mg/dL, N = 7); (2) Normal (>44 mg/dL, N = 10) HDL-C levels on a controlled diet. | Four supervised exercise sessions/week at 60–80% of the subjects’ measured maximal HR. | 12 months |
| [102] |
| Calisthenics, walking, jogging or running | Overweight men aged 30–59 years randomized to: (1) Running (N = 46); (2) Caloric restriction (N = 42); (3) Remain sedentary, non-dieting controls (N = 42). | At the beginning, exercise for 25 min three times/week at 60% to 80% of maximal HR increased to 40 to 50 min 4 days or more per week. | l-year study |
| [103] |
| A maximal-velocity incremented continuous treadmill run (Telineyhtymä, Kotka, Finland) at 1 slope until exhaustion | 24 male national top-level endurance runners; 12 middle-distance runners; and 12 marathon runners. | High intensity. | Training and competing in middle and long distance running regularly for more than 5 years. | Immediate effects
| [104] |
| Cycling | 20 recreationally active males allocated to: (1) Energy-deficit diet; (2) control group. | High-intensity cycling at 90% VO2peak at a constant workload for 8 min, 2 h after breakfast, every second day/10 days |
| [107] | |
| Aquatic exercise | Meta-analysis of 11 articles on 12 studies: (1) Exercise group (n = 208); (2) control group (n = 185). | Exercise session ranged from 15 to 60 min. The frequency of exercise was one to four sessions per week. | Duration of intervention ranged from 6 to 24 weeks (median: 12 weeks) |
| [111] |
| Running | 36 untrained men (mean age, 31 years) assigned to: (1) Intense interval running (n = 8); (2) Heavy-resistance strength training group (n = 8); (3) Prolonged moderate-intensity continuous running (n = 9); (4) No physical training (n = 11). | (1) Intense training; (2) Strength training; (3) Moderate-intensity training. | 12 weeks |
| [112] |
| Walking–jogging, stationary cycles and treadmills | 120 woman and 149 men who were sedentary, aged 50–56, free of CAD and not treated for hypertension or hyperlipidemia assigned to: (1) Higher-intensity, group-based exercise training; (2) Higher-intensity, home-based exercise training; (3) Lower-intensity, home-based exercise training; (4) A 1-year delayed treatment control (exercise training program during the second year). | (1,2) Exercise intensity gradually increased over the initial 6-week period to 73% to 88% of the peak heart rate; (3) Exercise intensity based on 60% to 73% of heart rate. | (1,2) 60 min training, 3 trainings/week (3) five 30-min exercise sessions per week |
| [114] |
| Walking, running, cycling and spinning+ | 1567 older adults (790 (50.4%) women) assigned to: (1) Control (n = 412); (2) HIIT (n = 142); (3) MICT (n = 119). | (2) Two weekly sessions of high-intensity training (10 min warmup followed by 4 × 4 min intervals at ~90% of peak heart rate); (3) Two weekly sessions of moderate-intensity training (50 min of continuous work at ∼70% of peak heart rate). | 5 years |
| [116] |
| Walking and exercises | 116 initially sedentary men and 119 women randomly assigned to: (1) 6-month lifestyle physical activity counselling intervention; or (2) 6-month gymnasium-based structured program. | (1) At least 30 min of moderate-intensity physical activity on most days of the week; (2) Traditional exercise with an intensity of 50–85% of maximal aerobic power and session durations of of 20–60 min for 3 to 5 days a week. | 6 months |
| [117] |
| Training on a Nautilus machine Cycling ergometry, rowing ergometry and treadmill walking/jogging | 48 sedentary, healthy women (mean age, 20.4 years) assigned to: (1) Control group (n = 12); (2) Aerobic training (n = 12); (3) Resistance training (n =12); (4) Cross training (combined aerobic and resistance training, n = 12). | (2) Cycling ergometry, rowing ergometry, treadmill walking/jogging three times/week (10 min warmup, 30 min exercise at 70–75% HRmax and 10 min cool-down); (3) Training on Nautilus machine 3 times/week, 2–3 sets of 8–10 repetitions at 60–70% of established RM; (4) Combination of the above. | 16 weeks of training, 6 weeks of detraining |
| [118] |
| 111 sedentary, overweight men and women with mild-to-moderate dyslipidemia assigned to: (1) Control group; (2) High-volume high-intensity exercise; (3) Low-volume high-intensity exercise; (4) Low-volume moderate-intensity exercise. | (2) Caloric equivalent of jogging 20 mi (32.0 km)/week at 65 to 80% of VO2max; (3) Equivalent of jogging 12 mi (19.2 km)/week at 65 to 80% of VO2max; (4) Equivalent of walking 12 mi per week at 40% to 55% of VO2max). | 6 months (1) 8 months (2) |
| [119] | |
| Exercises on treadmill; land-based or aquatically based treadmill | Overweight/obese men (n = 10) and women (n = 8). | Acute exercise session on a treadmill (70% VO2max, 400 kcal energy expenditure); Endurance exercise training (land-based or aquatic-based treadmill): 3 sessions/week, progressing to 500 kcal/session. | 12 weeks |
| [22] |
| Various | Meta-analysis of data of 1555 adults from 6 studies encompassing 10 distinct exercise programs: APOE (N = 106), DREW (N = 385), GERS (N = 79), HERITAGE (N = 715), STRRIDE I (N = 168) and II (N = 102). | Various. | Various |
| [91] |
| Various | 98 subjects (38 females, 60 males), aged 30–65 years with at least one CAD risk factor assigned to four groups: (1) Performance ≤99% at baseline, performance gain ≤7.9%; (2) Performance ≤99% at baseline, performance gain >7.9%; (3) Performance >100% at baseline, performance gain ≤7.9%; (4) Performance >100% at baseline, performance gain >7.9%. | At least 75 min/week of vigorous or 150 min/week of moderate-intensity endurance training. | 8 months |
| [39] |
| Bicycle ergometers | 30 sedentary subjects (20 with and 10 without MS). | Moderate-intensity exercise training on bicycle ergometers. | 3 months |
| [92] |
| Running, throwing hammer, wrestling and weightlifting | National-class male athletes: representatives in running (1500 m middle distance, n = 10), throwing (hammer, n = 10), wrestling (n = 10) and weightlifting (n = 8). Age- and gender-matched sedentary reference subjects (n = 14). | Athletes: exercising at least 8 h per day; Reference: regular exercise with moderate intensity less than 1 h/week. | At least 6 years, |
| [140] |
| Walk/run training | 35 patients with MS assigned to: (1) Control group (n = 12); (2) Exercise group (n = 27). | 10-week walk/run training program. | 10 weeks |
| [141] |
| Aerobic dance, running, skipping, and stepping Bands and free weights; Squats, lunges, bicep curls, push-ups and shoulder presses | 32 obese black South African women aged 20–35 years assigned to: (1) Exercise (combined aerobic and resistance exercise) (n = 20); or (2) Control (<1 session of <20 min per week) (n = 15). | Aerobic and resistance exercise training of moderate–vigorous intensity (75–80% peak heart rate, HR peak) for 40 to 60 min 4 days/week supervised by a trained exercise physiologist. Prescribed intensity of 60% to 70% HRpeak. | 12-weeks |
| [24] |
| Walking or Nordic walking, jogging, cycling, swimming, skiing, aerobics or other gymnastic exercise | 161 sedentary women aged 43–63 years with no current use of hormone therapy assigned to: (1) Exercise group; (2) Control group. | Unsupervised aerobic training 4 times/week, 50 min of the exercise each time, 64–80% of the maximal HR. | 6 months |
| [139] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Franczyk, B.; Gluba-Brzózka, A.; Ciałkowska-Rysz, A.; Ławiński, J.; Rysz, J. The Impact of Aerobic Exercise on HDL Quantity and Quality: A Narrative Review. Int. J. Mol. Sci. 2023, 24, 4653. https://doi.org/10.3390/ijms24054653
Franczyk B, Gluba-Brzózka A, Ciałkowska-Rysz A, Ławiński J, Rysz J. The Impact of Aerobic Exercise on HDL Quantity and Quality: A Narrative Review. International Journal of Molecular Sciences. 2023; 24(5):4653. https://doi.org/10.3390/ijms24054653
Chicago/Turabian StyleFranczyk, Beata, Anna Gluba-Brzózka, Aleksandra Ciałkowska-Rysz, Janusz Ławiński, and Jacek Rysz. 2023. "The Impact of Aerobic Exercise on HDL Quantity and Quality: A Narrative Review" International Journal of Molecular Sciences 24, no. 5: 4653. https://doi.org/10.3390/ijms24054653