
The Effects of Peripubertal THC Exposure in Neurodevelopmental Rat Models of Psychopathology
 , , , , , , , ,
, , , , , , , ,  , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Peripubertal THC Exposure in Prenatally MAM-Exposed Rats
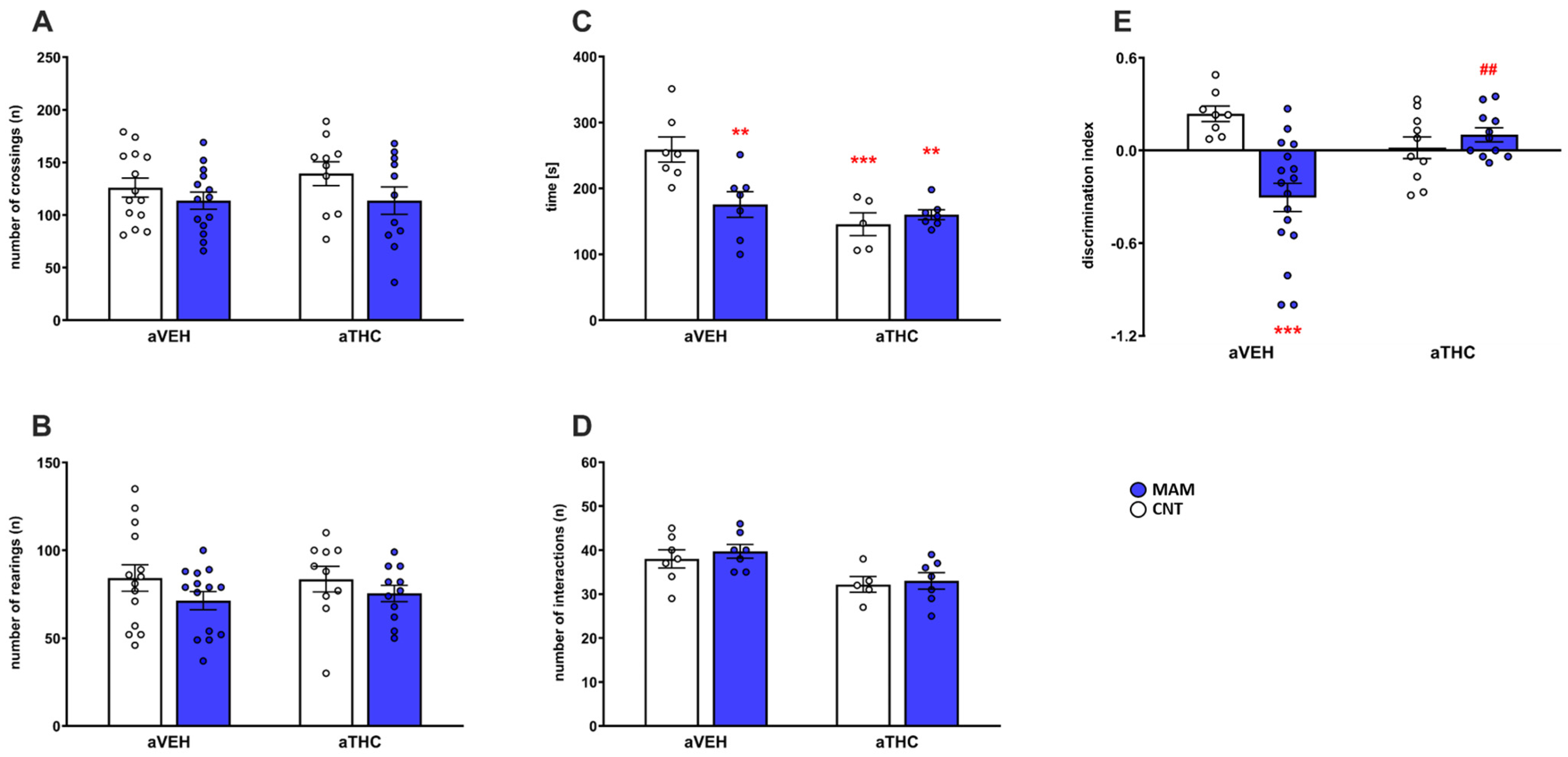
2.1.1. Behavioral Effects
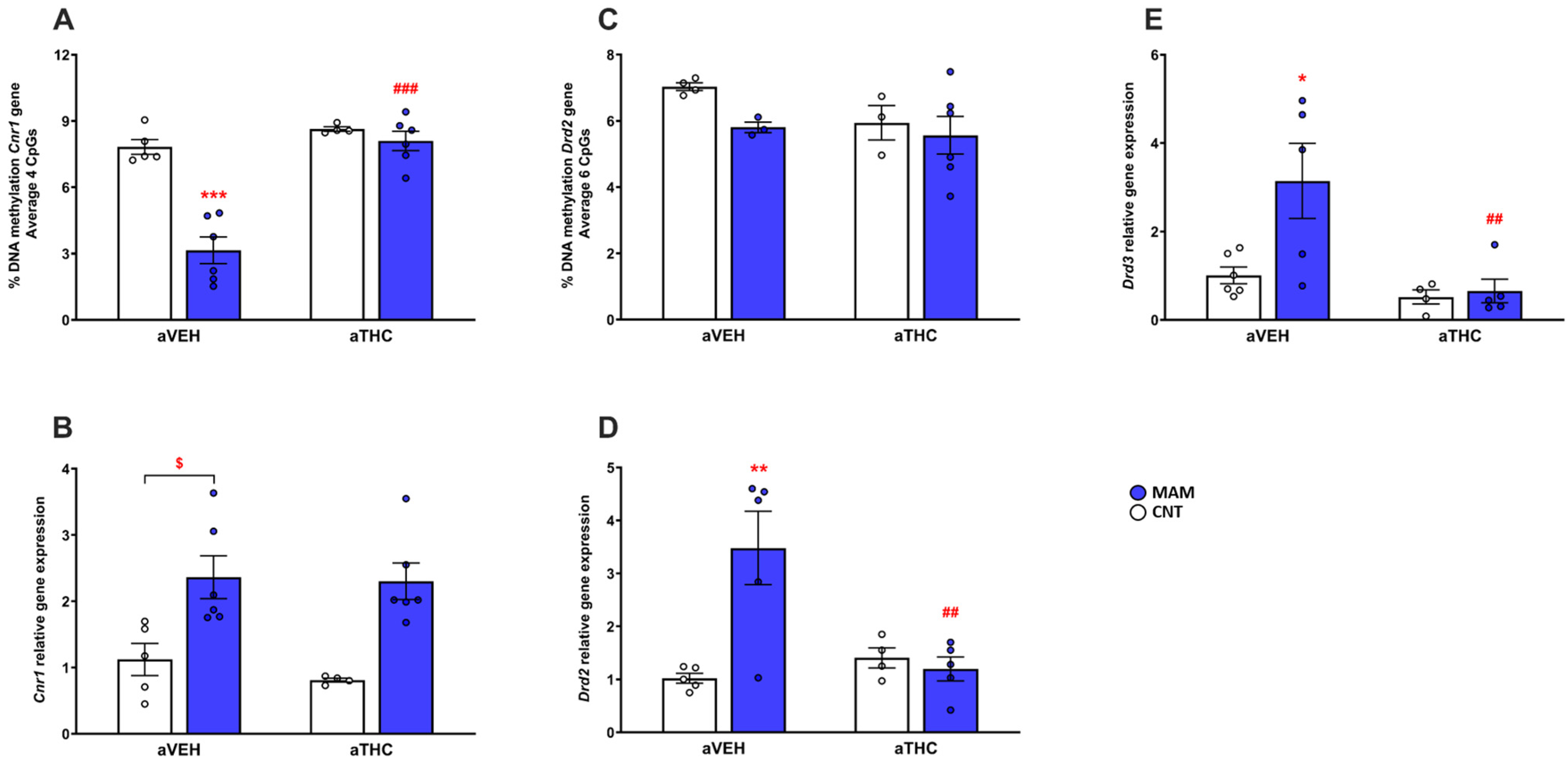
2.1.2. Molecular Effects
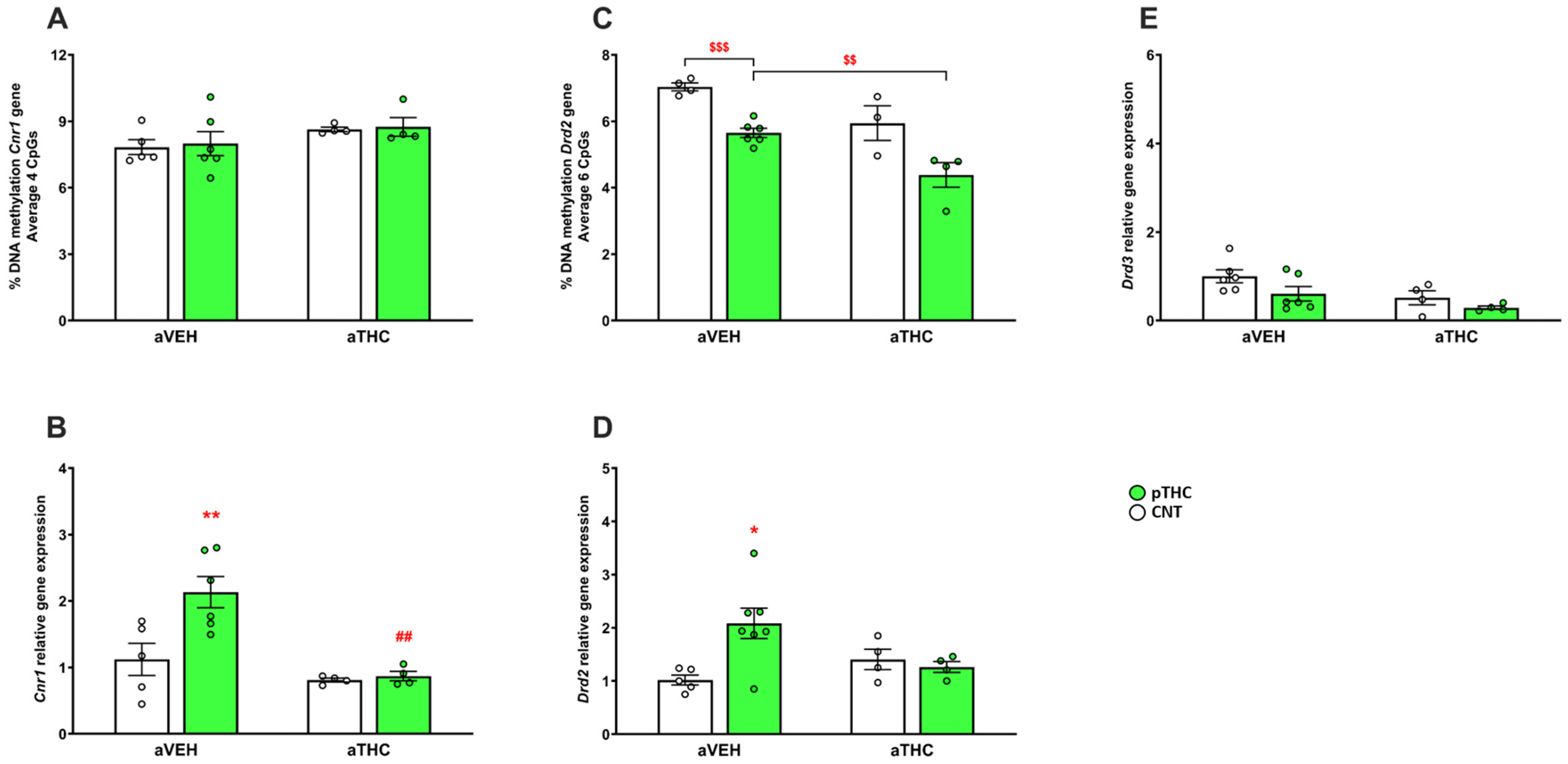
2.2. Peripubertal THC Exposure in Perinatally THC-Exposed Rats
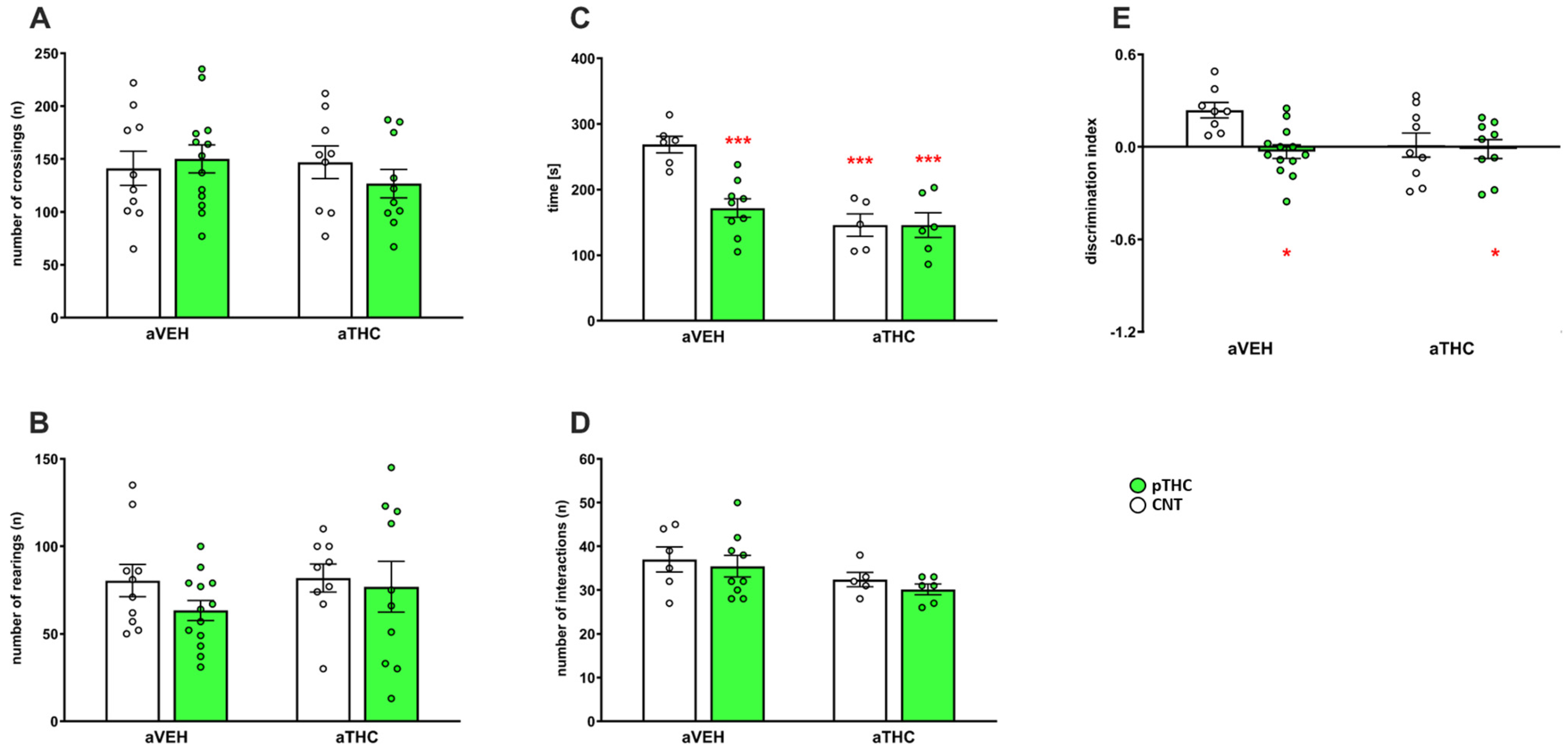
2.2.1. Behavioral Effects
2.2.2. Molecular Effects
3. Discussion
4. Materials and Methods
4.1. Animals
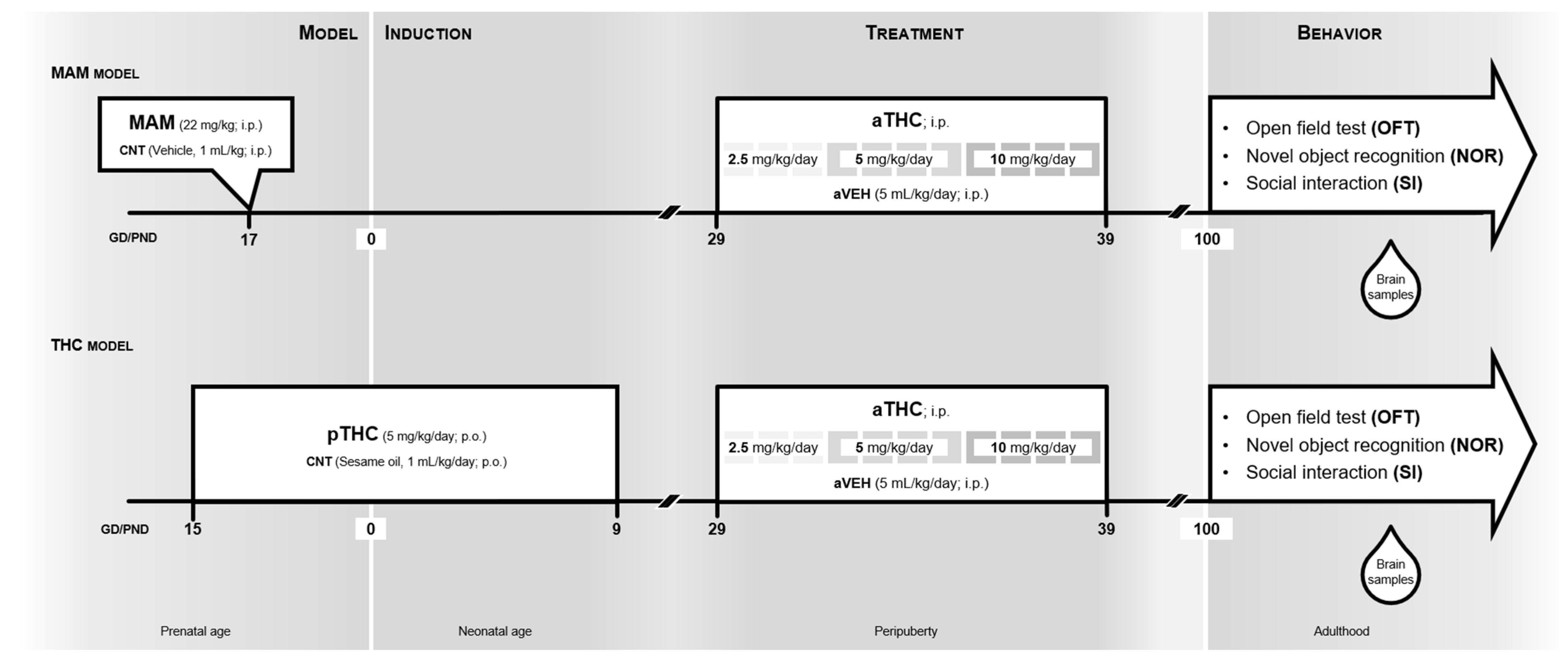
4.2. Drugs and Experimental Design
4.3. Behavioral Testing
4.3.1. Spontaneous Locomotor Activity in the Open Field Test (OFT)
4.3.2. Social Interaction (SI) Test
4.3.3. Novel Object Recognition (NOR) Test
4.4. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.5. DNA Methylation Analysis by Pyrosequencing
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Owen, M.J.; O’ Donovan, M.C.; Thapar, A.; Craddock, N. Neurodevelopmental hypothesis of schizophrenia. Br. J. Psychiatry 2011, 198, 173–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadeem, A.; Ahmad, S.F.; Al-Harbi, N.O.; Attia, S.M.; Bakheet, S.A.; Ibrahim, K.E.; Alqahtani, F.; Alqinyah, M. Nrf2 activator, sulforaphane ameliorates autism-like symptoms through suppression of Th17 related signaling and rectification of oxidant-antioxidant imbalance in periphery and brain of BTBR T+tf/J mice. Behav. Brain Res. 2019, 364, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Alshammari, M.A.; Attia, S.M. Protection by tyrosine kinase inhibitor, tyrphostin AG126, through the suppression of IL-17A, RORγt, and T-bet signaling, in the BTBR mouse model of autism. Brain Res. Bull. 2018, 142, 328–337. [Google Scholar] [CrossRef]
- Ahmad, S.F.; Ansari, M.A.; Nadeem, A.; Bakheet, S.A.; Alqahtani, F.; Alhoshani, A.R.; Alasmari, F.; Alsaleh, N.B.; Attia, S.M. 5-aminoisoquinolinone attenuates social behavior deficits and immune abnormalities in the BTBR T+ Itpr3tf/J mouse model for autism. Pharmacol. Biochem. Behav. 2020, 189, 172859. [Google Scholar] [CrossRef]
- Al-Mazroua, H.A.; Nadeem, A.; Ansari, M.A.; Attia, S.M.; Albekairi, T.H.; Bakheet, S.A.; Alobaidi, A.F.; Alhosaini, K.; Alqarni, S.A.; Ibrahim, K.E.; et al. Methylmercury chloride exposure exacerbates existing neurobehavioral and immune dysfunctions in the BTBR T+ Itpr3tf/J mouse model of autism. Immunol. Lett. 2022, 244, 19–27. [Google Scholar] [CrossRef]
- Higuera-Matas, A.; Ucha, M.; Ambrosio, E. Long-term consequences of perinatal and adolescent cannabinoid exposure on neural and psychological processes. Neurosci. Biobehav. Rev. 2015, 55, 119–146. [Google Scholar] [CrossRef] [PubMed]
- Stiles, J.; Jernigan, T.L. The Basics of Brain Development. Neuropsychol. Rev. 2010, 20, 327–348. [Google Scholar] [CrossRef] [Green Version]
- Giedd, J.N.; Blumenthal, J.; Jeffries, N.O.; Castellanos, F.X.; Liu, H.; Zijdenbos, A.; Paus, T.; Evans, A.C.; Rapoport, J.L. Brain development during childhood and adolescence: A longitudinal MRI study. Nat. Neurosci. 1999, 2, 861–863. [Google Scholar] [CrossRef]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2020, 24, 417–463. [Google Scholar] [CrossRef]
- Andersen, S.L. Trajectories of brain development: Point of vulnerability or window of opportunity? Neurosci. Biobehav. Rev. 2003, 27, 3–18. [Google Scholar] [CrossRef]
- Micale, V.; Kucerova, J.; Sulcova, A. Leading compounds for the validation of animal models of psychopathology. Cell Tissue Res. 2013, 354, 309–330. [Google Scholar] [CrossRef] [PubMed]
- Alpár, A.; Di Marzo, V.; Harkany, T. At the Tip of an Iceberg: Prenatal Marijuana and Its Possible Relation to Neuropsychiatric Outcome in the Offspring. Biol. Psychiatry 2016, 79, e33–e45. [Google Scholar] [CrossRef] [PubMed]
- Di Bartolomeo, M.; Stark, T.; Maurel, O.M.; Iannotti, F.A.; Kuchar, M.; Ruda-Kucerova, J.; Piscitelli, F.; Laudani, S.; Pekarik, V.; Salomone, S.; et al. Crosstalk between the transcriptional regulation of dopamine D2 and cannabinoid CB1 receptors in schizophrenia: Analyses in patients and in perinatal Δ9-tetrahydrocannabinol-exposed rats. Pharmacol. Res. 2021, 164, 105357. [Google Scholar] [CrossRef]
- Stark, T.; Di Martino, S.; Drago, F.; Wotjak, C.T.; Micale, V. Phytocannabinoids and schizophrenia: Focus on adolescence as a critical window of enhanced vulnerability and opportunity for treatment. Pharmacol. Res. 2021, 174, 105938. [Google Scholar] [CrossRef] [PubMed]
- Hermann, H.; Marsicano, G.; Lutz, B. Coexpression of the cannabinoid receptor type 1 with dopamine and serotonin receptors in distinct neuronal subpopulations of the adult mouse forebrain. Neuroscience 2002, 109, 451–460. [Google Scholar] [CrossRef]
- Meschler, J.P.; Howlett, A.C. Signal transduction interactions between CB1 cannabinoid and dopamine receptors in the rat and monkey striatum. Neuropharmacology 2001, 40, 918–926. [Google Scholar] [CrossRef]
- Stark, T.; Iannotti, F.A.; Di Martino, S.; Di Bartolomeo, M.; Ruda-Kucerova, J.; Piscitelli, F.; Wotjak, C.T.; D’Addario, C.; Drago, F.; Di Marzo, V.; et al. Early Blockade of CB1 Receptors Ameliorates Schizophrenia-like Alterations in the Neurodevelopmental MAM Model of Schizophrenia. Biomolecules 2022, 12, 108. [Google Scholar] [CrossRef] [PubMed]
- Ruda-Kucerova, J.; Babinska, Z.; Amchova, P.; Stark, T.; Drago, F.; Sulcova, A.; Micale, V. Reactivity to addictive drugs in the methylazoxymethanol (MAM) model of schizophrenia in male and female rats. World J. Biol. Psychiatry 2017, 18, 129–142. [Google Scholar] [CrossRef]
- D’Addario, C.; Micale, V.; Di Bartolomeo, M.; Stark, T.; Pucci, M.; Sulcova, A.; Palazzo, M.; Babinska, Z.; Cremaschi, L.; Drago, F.; et al. A preliminary study of endocannabinoid system regulation in psychosis: Distinct alterations of CNR1 promoter DNA methylation in patients with schizophrenia. Schizophr. Res. 2017, 188, 132–140. [Google Scholar] [CrossRef]
- Stark, T.; Ruda-Kucerova, J.; Iannotti, F.A.; D’Addario, C.; Di Marco, R.; Pekarik, V.; Drazanova, E.; Piscitelli, F.; Bari, M.; Babinska, Z.; et al. Peripubertal cannabidiol treatment rescues behavioral and neurochemical abnormalities in the MAM model of schizophrenia. Neuropharmacology 2019, 146, 212–221. [Google Scholar] [CrossRef]
- Stark, T.; Di Bartolomeo, M.; Di Marco, R.; Drazanova, E.; Platania, C.B.M.; Iannotti, F.A.; Ruda-Kucerova, J.; D’Addario, C.; Kratka, L.; Pekarik, V.; et al. Altered dopamine D3 receptor gene expression in MAM model of schizophrenia is reversed by peripubertal cannabidiol treatment. Biochem. Pharmacol. 2020, 177, 114004. [Google Scholar] [CrossRef] [PubMed]
- Drazanova, E.; Ruda-Kucerova, J.; Kratka, L.; Stark, T.; Kuchar, M.; Maryska, M.; Drago, F.; Starcuk, Z.; Micale, V. Different effects of prenatal MAM vs. perinatal THC exposure on regional cerebral blood perfusion detected by Arterial Spin Labelling MRI in rats. Sci. Rep. 2019, 9, 6062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kucerova, J.; Tabiova, K.; Drago, F.; Micale, V. Therapeutic Potential of Cannabinoids in Schizophrenia. Recent Pat. CNS Drug Discov. 2014, 9, 13–25. [Google Scholar] [CrossRef]
- Peters, K.Z.; Naneix, F. The role of dopamine and endocannabinoid systems in prefrontal cortex development: Adolescence as a critical period. Front. Neural Circuits 2022, 16, 939235. [Google Scholar] [CrossRef]
- Micale, V.; Di Bartolomeo, M.; Di Martino, S.; Stark, T.; Dell’Osso, B.; Drago, F.; D’Addario, C. Are the epigenetic changes predictive of therapeutic efficacy for psychiatric disorders? A translational approach towards novel drug targets. Pharmacol. Ther. 2023, 241, 108279. [Google Scholar] [CrossRef]
- Howes, O.D.; Kapur, S. The Dopamine Hypothesis of Schizophrenia: Version III--The Final Common Pathway. Schizophr. Bull. 2009, 35, 549–562. [Google Scholar] [CrossRef] [Green Version]
- Shing, N.; Walker, M.C.; Chang, P. The role of aberrant neural oscillations in the hippocampal-medial prefrontal cortex circuit in neurodevelopmental and neurological disorders. Neurobiol. Learn. Mem. 2022, 195, 107683. [Google Scholar] [CrossRef]
- Eggan, S.M.; Hashimoto, T.; Lewis, D.A. Reduced Cortical Cannabinoid 1 Receptor Messenger RNA and Protein Expression in Schizophrenia. Arch. Gen. Psychiatry 2008, 65, 772. [Google Scholar] [CrossRef] [PubMed]
- DiNieri, J.A.; Wang, X.; Szutorisz, H.; Spano, S.M.; Kaur, J.; Casaccia, P.; Dow-Edwards, D.; Hurd, Y.L. Maternal Cannabis Use Alters Ventral Striatal Dopamine D2 Gene Regulation in the Offspring. Biol. Psychiatry 2011, 70, 763–769. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Dow-Edwards, D.; Anderson, V.; Minkoff, H.; Hurd, Y.L. In utero marijuana exposure associated with abnormal amygdala dopamine D2 gene expression in the human fetus. Biol. Psychiatry 2004, 56, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Saito, A.; Ballinger, M.D.L.; Pletnikov, M.V.; Wong, D.F.; Kamiya, A. Endocannabinoid system: Potential novel targets for treatment of schizophrenia. Neurobiol. Dis. 2013, 53, 10–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruggiero, R.N.; Rossignoli, M.T.; De Ross, J.B.; Hallak, J.E.C.; Leite, J.P.; Bueno-Junior, L.S. Cannabinoids and Vanilloids in Schizophrenia: Neurophysiological Evidence and Directions for Basic Research. Front. Pharmacol. 2017, 8, 399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, M. Adolescence as a vulnerable period to alter rodent behavior. Cell Tissue Res. 2013, 354, 99–106. [Google Scholar] [CrossRef]
- Feigenson, K.A.; Kusnecov, A.W.; Silverstein, S.M. Inflammation and the two-hit hypothesis of schizophrenia. Neurosci. Biobehav. Rev. 2014, 38, 72–93. [Google Scholar] [CrossRef] [Green Version]
- Gomes, F.V.; Guimarães, F.S.; Grace, A.A. Effects of Pubertal Cannabinoid Administration on Attentional Set-Shifting and Dopaminergic Hyper-Responsivity in a Developmental Disruption Model of Schizophrenia. Int. J. Neuropsychopharmacol. 2015, 18, pyu018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, D.D.; Giuffrida, A.; Lodge, D.J. Adolescent Synthetic Cannabinoid Exposure Produces Enduring Changes in Dopamine Neuron Activity in a Rodent Model of Schizophrenia Susceptibility. Int. J. Neuropsychopharmacol. 2018, 21, 393–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecca, S.; Luchicchi, A.; Scherma, M.; Fadda, P.; Muntoni, A.L.; Pistis, M. Δ9-Tetrahydrocannabinol During Adolescence Attenuates Disruption of Dopamine Function Induced in Rats by Maternal Immune Activation. Front. Behav. Neurosci. 2019, 13, 202. [Google Scholar] [CrossRef] [PubMed]
- Ruda-Kucerova, J.; Babinska, Z.; Stark, T.; Micale, V. Suppression of Methamphetamine Self-Administration by Ketamine Pre-treatment Is Absent in the Methylazoxymethanol (MAM) Rat Model of Schizophrenia. Neurotox. Res. 2017, 32, 121–133. [Google Scholar] [CrossRef]
- Večeřa, J.; Bártová, E.; Krejčí, J.; Legartová, S.; Komůrková, D.; Rudá-Kučerová, J.; Štark, T.; Dražanová, E.; Kašpárek, T.; Šulcová, A.; et al. HDAC1 and HDAC3 underlie dynamic H3K9 acetylation during embryonic neurogenesis and in schizophrenia-like animals. J. Cell. Physiol. 2018, 233, 530–548. [Google Scholar] [CrossRef]
- Horska, K.; Kotolova, H.; Karpisek, M.; Babinska, Z.; Hammer, T.; Prochazka, J.; Stark, T.; Micale, V.; Ruda-Kucerova, J. Metabolic profile of methylazoxymethanol model of schizophrenia in rats and effects of three antipsychotics in long-acting formulation. Toxicol. Appl. Pharmacol. 2020, 406, 115214. [Google Scholar] [CrossRef]
- Kucera, J.; Horska, K.; Hruska, P.; Kuruczova, D.; Micale, V.; Ruda-Kucerova, J.; Bienertova-Vasku, J. Interacting effects of the MAM model of schizophrenia and antipsychotic treatment: Untargeted proteomics approach in adipose tissue. Progr. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 108, 110165. [Google Scholar] [CrossRef] [PubMed]
- Uttl, L.; Szczurowska, E.; Hájková, K.; Horsley, R.R.; Štefková, K.; Hložek, T.; Šíchová, K.; Balíková, M.; Kuchař, M.; Micale, v.; et al. Behavioral and Pharmacokinetic Profile of Indole-Derived Synthetic Cannabinoids JWH-073 and JWH-210 as Compared to the Phytocannabinoid Δ9-THC in Rats. Front. Neurosci. 2018, 12, 703. [Google Scholar] [CrossRef]
- Brancato, A.; Castelli, V.; Lavanco, G.; Tringali, G.; Micale, V.; Kuchar, M.; D’Amico, C.; Pizzolanti, G.; Feo, S.; Cannizzaro, C. Binge-like Alcohol Exposure in Adolescence: Behavioural, Neuroendocrine and Molecular Evidence of Abnormal Neuroplasticity… and Return. Biomedicines 2021, 9, 1161. [Google Scholar] [CrossRef]
- Molina-Holgado, F.; Amaro, A.; González, M.I.; Alvarez, F.J.; Leret, M.L. Effect of maternal Δ9-tetrahydrocannabinol on developing serotonergic system. Eur. J. Pharmacol. 1996, 316, 39–42. [Google Scholar] [CrossRef] [Green Version]
- Scherma, M.; Dessì, C.; Muntoni, A.L.; Lecca, S.; Satta, V.; Luchicchi, A.; Pistis, M.; Panlilio, L.V.; Fattore, L.; Goldberg, S.R.; et al. Adolescent Δ9-Tetrahydrocannabinol Exposure Alters WIN55,212-2 Self-Administration in Adult Rats. Neuropsychopharmacology 2016, 41, 1416–1426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, A.L.; Drago, F.; Wotjak, C.T.; Micale, V. The Dopamine and Cannabinoid Interaction in the Modulation of Emotions and Cognition: Assessing the Role of Cannabinoid CB1 Receptor in Neurons Expressing Dopamine D1 Receptors. Front. Behav. Neurosci. 2011, 5, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzian, A.L.B.; Micale, V.; Wotjak, C.T. Cannabinoid receptor type 1 receptors on GABAergic vs. glutamatergic neurons differentially gate sex-dependent social interest in mice. Eur. J. Neurosci. 2014, 40, 2293–2298. [Google Scholar] [CrossRef]
- Chiodi, V.; Domenici, M.R.; Biagini, T.; de Simone, R.; Tartaglione, A.M.; Di Rosa, M.; lo Re, O.; Mazza, T.; Micale, V.; Vinciguerra, M. Systemic depletion of histone macroH2A1.1 boosts hippocampal synaptic plasticity and social behavior in mice. FASEB J. 2021, 35, e21793. [Google Scholar] [CrossRef]
- Raffaele, M.; Kovacovicova, K.; Biagini, T.; Lo Re, O.; Frohlich, J.; Giallongo, S.; Nhan, J.D.; Giannone, A.G.; Cabibi, D.; Ivanov, M.; et al. Nociceptin/orphanin FQ opioid receptor (NOP) selective ligand MCOPPB links anxiolytic and senolytic effects. Geroscience 2022, 44, 463–483. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 4th ed.; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Drago, F.; Nicolosi, A.; Micale, V.; Lo Menzo, G. Placebo affects the performance of rats in models of depression: Is it a good control for behavioral experiments? Eur. Neuropsychopharmacol. 2001, 11, 209–213. [Google Scholar] [CrossRef]
- Tamburella, A.; Micale, V.; Navarria, A.; Drago, F. Antidepressant properties of the 5-HT4 receptor partial agonist, SL65.0155: Behavioral and neurochemical studies in rats. Progr. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 1205–1210. [Google Scholar] [CrossRef] [PubMed]
- Tamburella, A.; Micale, V.; Mazzola, C.; Salomone, S.; Drago, F. The selective norepinephrine reuptake inhibitor atomoxetine counteracts behavioral impairments in trimethyltin-intoxicated rats. Eur. J. Pharmacol. 2012, 683, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, F.A.; Henes, K.; Micale, V.; Mauch, C.P.; Takahashi, R.N.; Wotjak, C.T. Prolonged fear incubation leads to generalized avoidance behavior in mice. J. Psychiatr. Res. 2011, 45, 354–360. [Google Scholar] [CrossRef]
- Ruda-Kucerova, J.; Amchova, P.; Babinska, Z.; Dusek, L.; Micale, V.; Sulcova, A. Sex Differences in the Reinstatement of Methamphetamine Seeking after Forced Abstinence in Sprague-Dawley Rats. Front. Psychiatry 2015, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyon, E. Mutation detection using fluorescent hybridization probes and melting curve analysis. Expert Rev. Mol. Diagn. 2011, 1, 92–101. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | Behavioral Effects | Molecular Effects (PFC) | ||||||
|---|---|---|---|---|---|---|---|---|
| OFT | SI | NOR | CB1 Gene | CB1 Meth | D2 Gene | D2 Meth | D3 Gene | |
| CNT + aVEH | ↔ | ↔ | ↔ | ↔ | ↔ | ↔ | ↔ | ↔ |
| MAM + aVEH | ↔ | ↓ | ↓ | ↑ | ↓ | ↑ | ↔ | ↑ |
| MAM + aTHC | ↔ | ↓ | ↑ | ↔ | ↑ | ↓ | ↔ | ↓ |
| CNT + aTHC | ↔ | ↓ | ↔ | ↔ | ↔ | ↔ | ↔ | ↔ |
| pTHC + aVEH | ↔ | ↓ | ↓ | ↑ | ↔ | ↑ | ↓ | ↔ |
| pTHC + aTHC | ↔ | ↓ | ↓ | ↓ | ↔ | ↔ | ↓ | ↔ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Bartolomeo, M.; Stark, T.; Di Martino, S.; Iannotti, F.A.; Ruda-Kucerova, J.; Romano, G.L.; Kuchar, M.; Laudani, S.; Palivec, P.; Piscitelli, F.; et al. The Effects of Peripubertal THC Exposure in Neurodevelopmental Rat Models of Psychopathology. Int. J. Mol. Sci. 2023, 24, 3907. https://doi.org/10.3390/ijms24043907
Di Bartolomeo M, Stark T, Di Martino S, Iannotti FA, Ruda-Kucerova J, Romano GL, Kuchar M, Laudani S, Palivec P, Piscitelli F, et al. The Effects of Peripubertal THC Exposure in Neurodevelopmental Rat Models of Psychopathology. International Journal of Molecular Sciences. 2023; 24(4):3907. https://doi.org/10.3390/ijms24043907
Chicago/Turabian StyleDi Bartolomeo, Martina, Tibor Stark, Serena Di Martino, Fabio Arturo Iannotti, Jana Ruda-Kucerova, Giovanni Luca Romano, Martin Kuchar, Samuele Laudani, Petr Palivec, Fabiana Piscitelli, and et al. 2023. "The Effects of Peripubertal THC Exposure in Neurodevelopmental Rat Models of Psychopathology" International Journal of Molecular Sciences 24, no. 4: 3907. https://doi.org/10.3390/ijms24043907