
Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
2.1. Isolation and Host Range Analysis of Phages
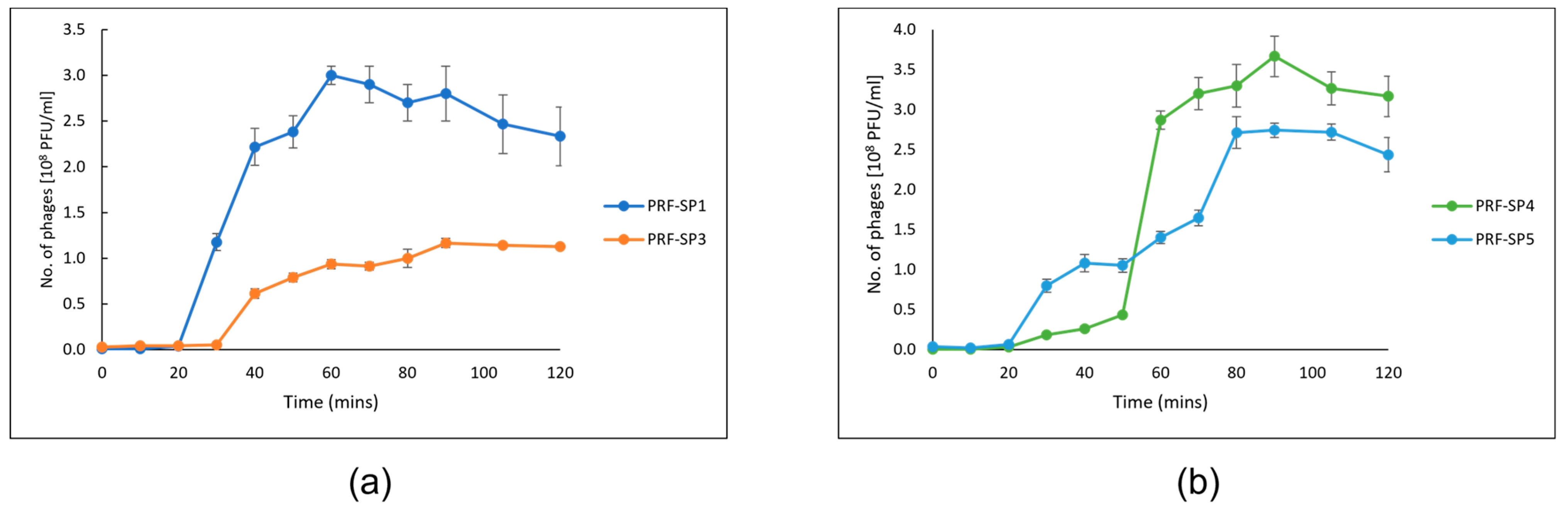
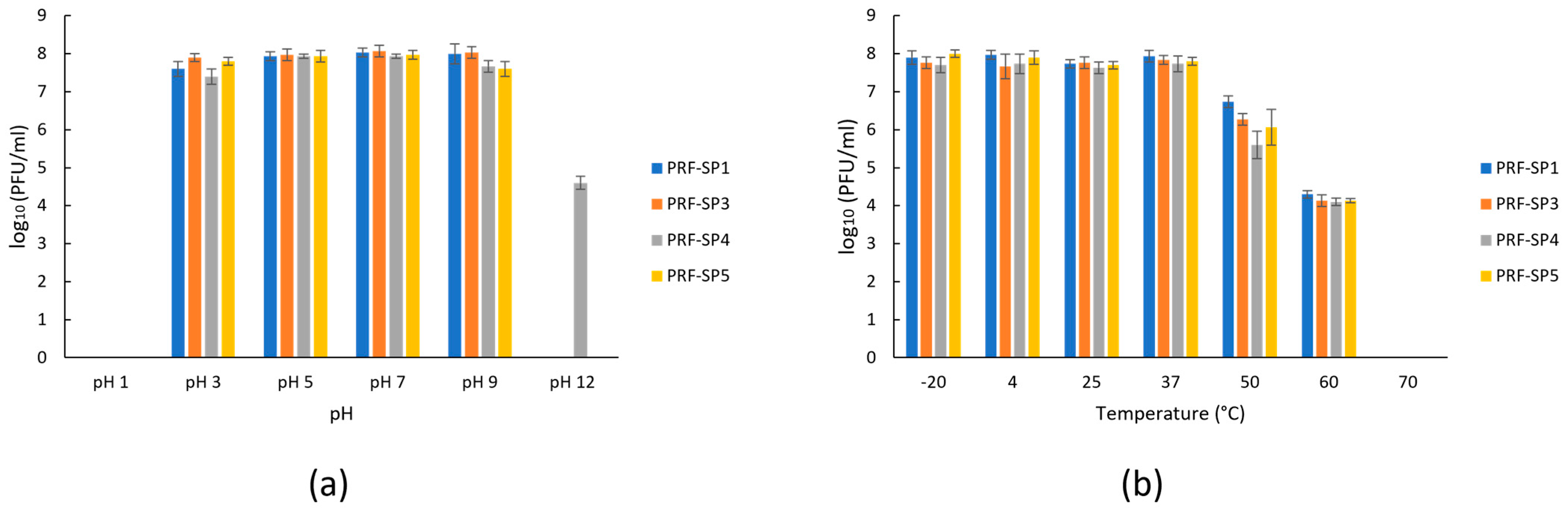
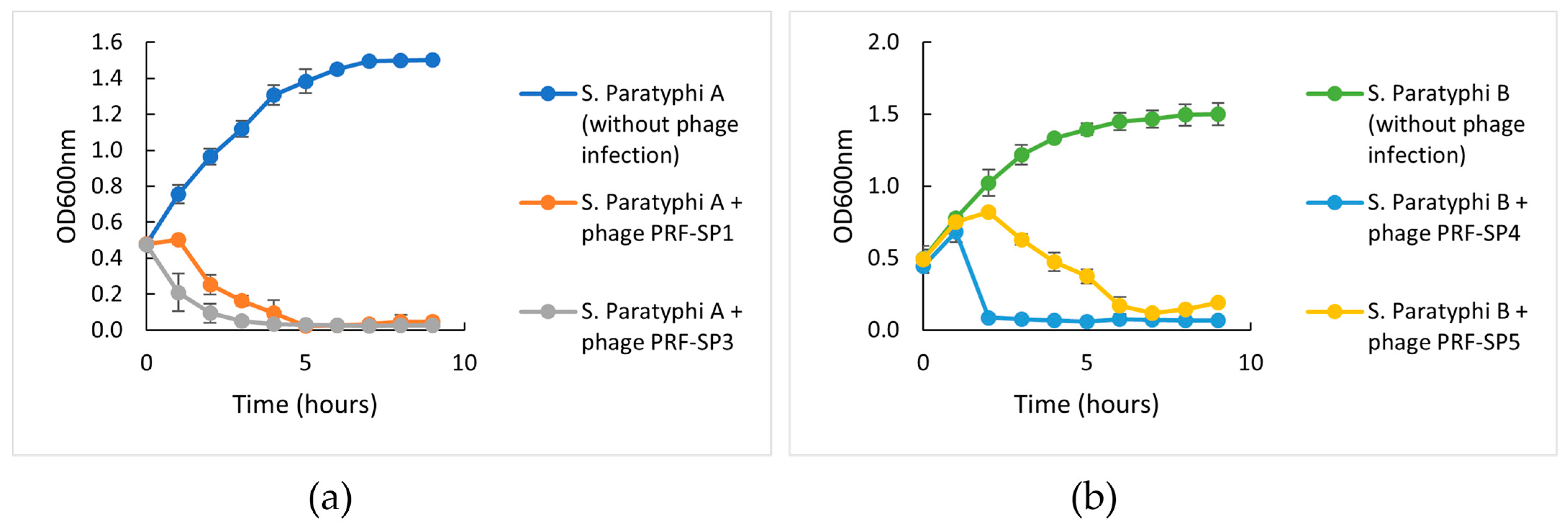
2.2. Phenotypic Characteristics of Phages
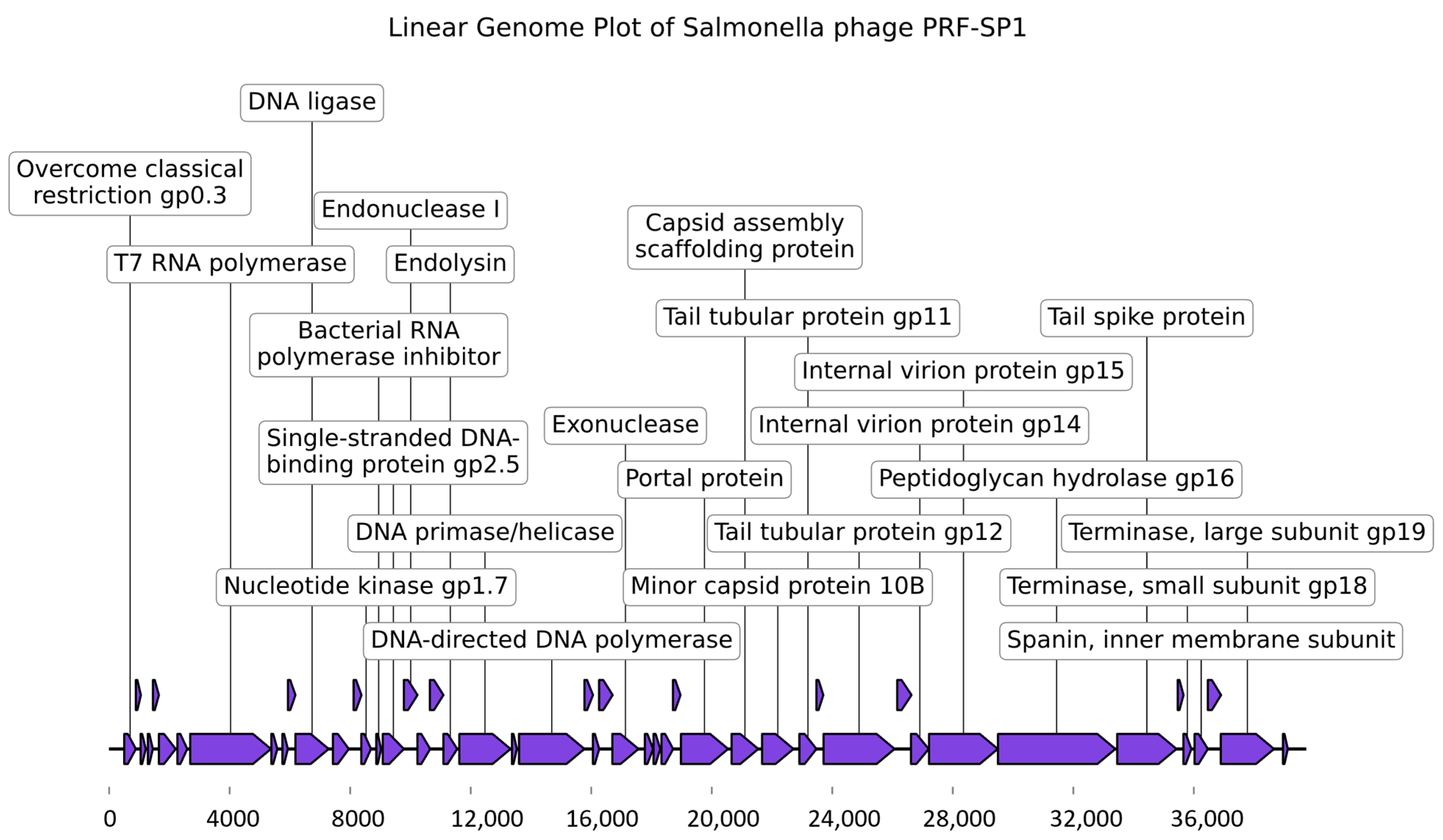
2.3. Genome Characterization
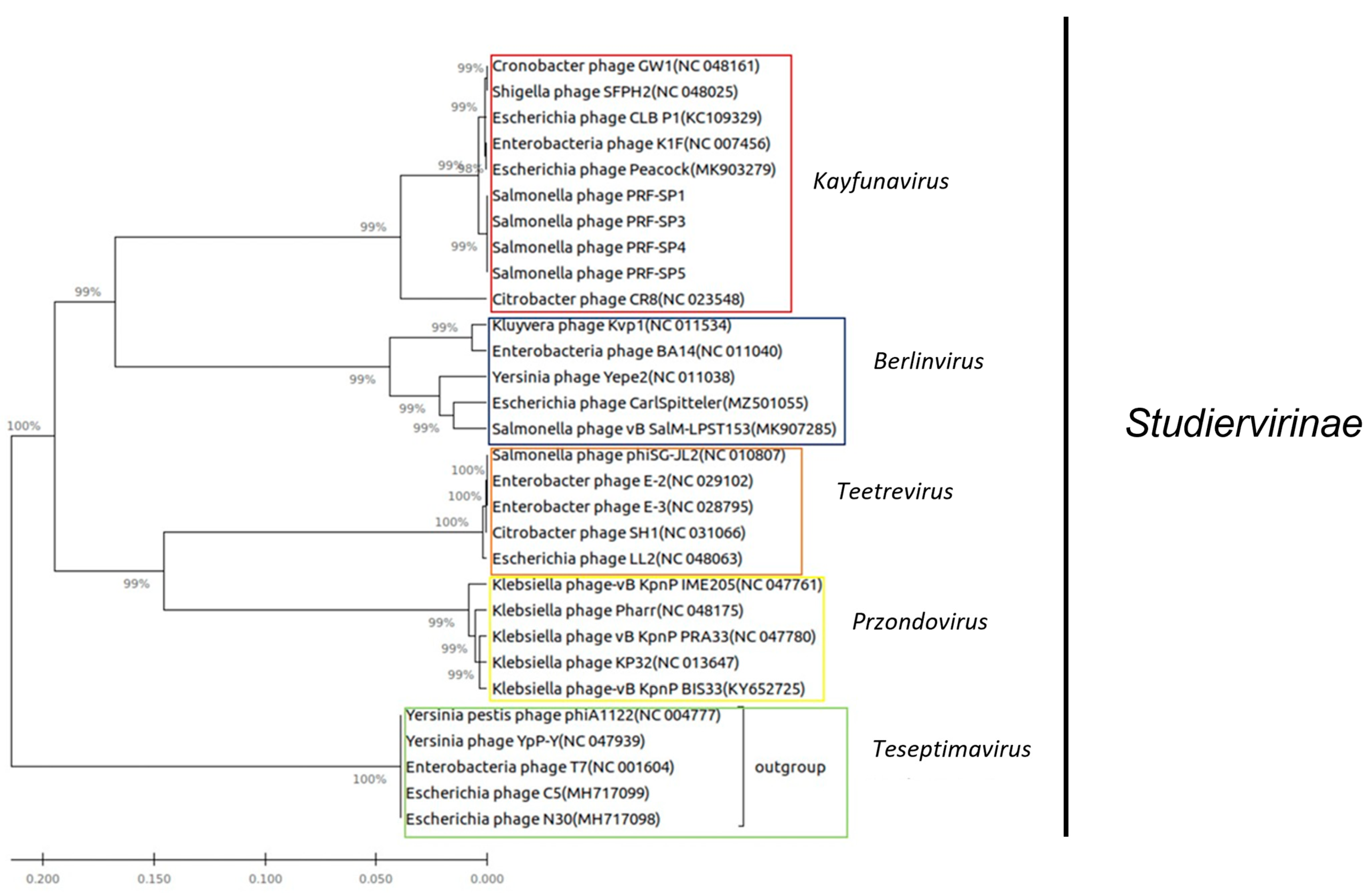
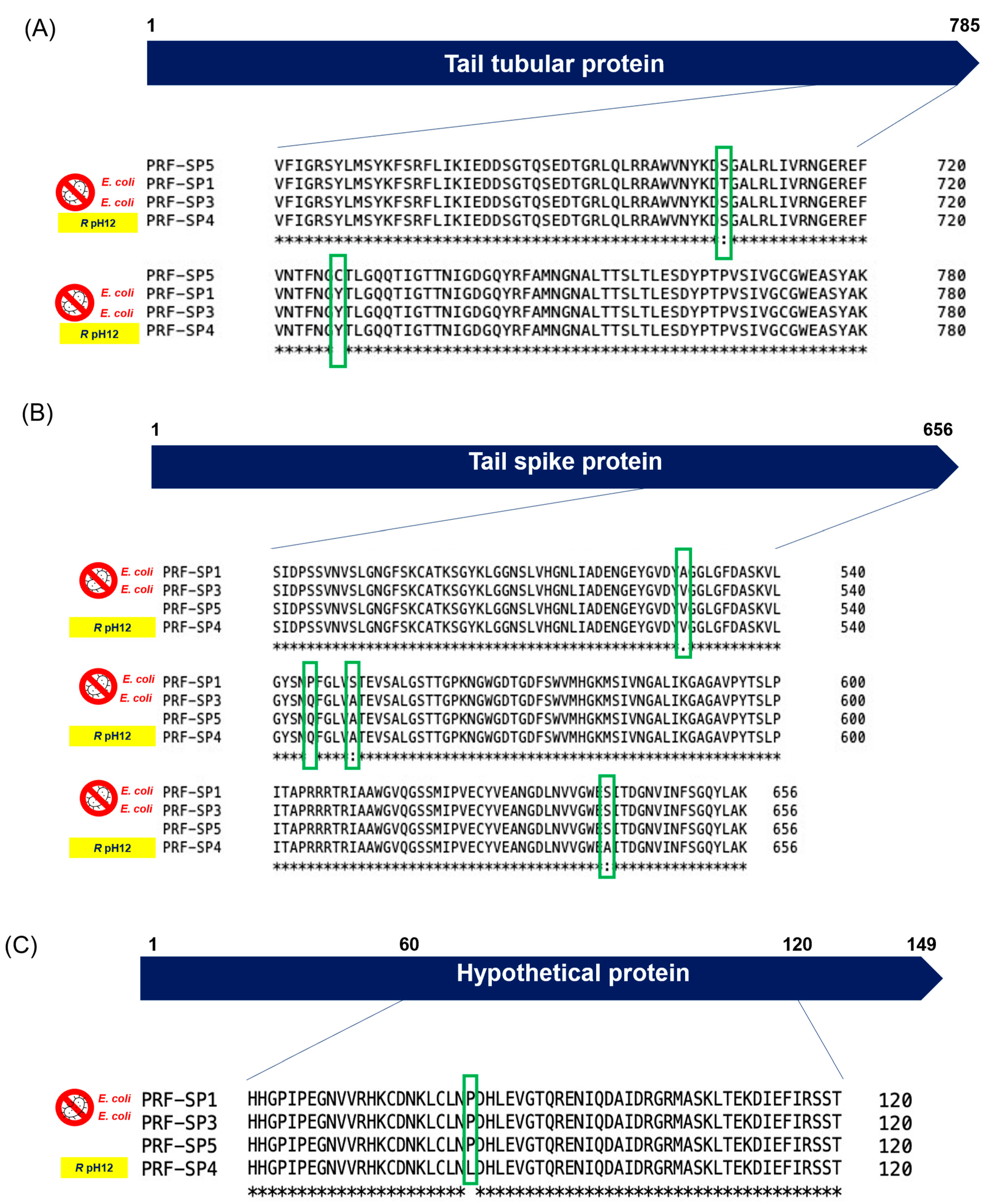
2.4. Comparative Genomic Analysis of Phages
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Culture Conditions
4.2. Sample Collection
4.3. Isolation of Bacteriophages from Soil and Water Samples
4.4. Purification of Bacteriophages
4.5. Propagation and Determination of Phage Titer
4.6. Host Range of Phages
4.7. Determination of Multiplicity of Infection (MOI)
4.8. One-Step Growth Curve
4.9. Transmission Electron Microscopy
4.10. Phage Stability at Different Temperatures and pH Values
4.11. Host Challenge Test
4.12. Genome Sequencing and Comparative Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Popa, G.L.; Papa, M.I. Salmonella spp. Infection—A continuous threat worldwide. Germs 2021, 11, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, Clinical Presentation, Laboratory Diagnosis, Antimicrobial Resistance, and Antimicrobial Management of Invasive Salmonella Infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- Fusheini, A.; Gyawu, S.K. Prevalence of Typhoid and Paratyphoid Fever in the Hohoe Municipality of the Volta Region, Ghana: A Five-Year Retrospective Trend Analysis. Ann. Glob. Health 2020, 86, 111. [Google Scholar] [CrossRef] [PubMed]
- Diep, B.; Barretto, C.; Portmann, A.-C.; Fournier, C.; Karczmarek, A.; Voets, G.; Li, S.; Deng, X.; Klijn, A. Salmonella Serotyping; Comparison of the Traditional Method to a Microarray-Based Method and an in silico Platform Using Whole Genome Sequencing Data. Front. Microbiol. 2019, 10, 2554. [Google Scholar] [CrossRef] [PubMed]
- Makalatia, K.; Kakabadze, E.; Wagemans, J.; Grdzelishvili, N.; Bakuradze, N.; Natroshvili, G.; Macharashvili, N.; Sedrakyan, A.; Arakelova, K.; Ktsoyan, Z.; et al. Characterization of Salmonella Isolates from Various Geographical Regions of the Caucasus and Their Susceptibility to Bacteriophages. Viruses 2020, 12, 1418. [Google Scholar] [CrossRef] [PubMed]
- de Freitas Neto, O.; Penha Filho, R.a.C.; Barrow, P.; Berchieri Junior, A. Sources of human non-typhoid salmonellosis: A review. Braz. J. Poult. Sci. 2010, 12, 1–11. [Google Scholar] [CrossRef]
- Carstens, C.K.; Salazar, J.K.; Darkoh, C. Multistate Outbreaks of Foodborne Illness in the United States Associated With Fresh Produce from 2010 to 2017. Front. Microbiol. 2019, 10, 2667. [Google Scholar] [CrossRef]
- Ayukekbong, J.A.; Ntemgwa, M.; Atabe, A.N. The threat of antimicrobial resistance in developing countries: Causes and control strategies. Antimicrob. Resist. Infect. Control. 2017, 6, 47. [Google Scholar] [CrossRef]
- The Lancet. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis—The Lancet. 2022. Available online: https://www.thelancet.com/journals/lancet/article/PIIS0140-6736(21)02724-0/fulltext (accessed on 15 February 2022).
- Keen, E.C. A century of phage research: Bacteriophages and the shaping of modern biology. BioEssays News Rev. Mol. Cell Dev. Biol. 2014, 37, 6–9. [Google Scholar] [CrossRef]
- Ha, A.D.; Denver, D.R. Comparative Genomic Analysis of 130 Bacteriophages Infecting Bacteria in the Genus Pseudomonas. Front. Microbiol. 2018, 9, 1456. [Google Scholar] [CrossRef]
- Moye, Z.D.; Woolston, J.; Sulakvelidze, A. Bacteriophage Applications for Food Production and Processing. Viruses 2018, 10, E205. [Google Scholar] [CrossRef]
- Akhtar, M.; Viazis, S.; Diez-Gonzalez, F. Isolation, identification and characterization of lytic, wide host range bacteriophages from waste effluents against Salmonella enterica serovars. Food Control. 2014, 38, 67–74. [Google Scholar] [CrossRef]
- Bielke, L.; Higgins, S.; Donoghue, A.; Donoghue, D.; Hargis, B.M. Salmonella Host Range of Bacteriophages That Infect Multiple Genera. Poult. Sci. 2007, 86, 2536–2540. [Google Scholar] [CrossRef]
- Mahmoud, M.; Askora, A.; Barakat, A.B.; Rabie, O.E.-F.; Hassan, S.E. Isolation and characterization of polyvalent bacteriophages infecting multi drug resistant Salmonella serovars isolated from broilers in Egypt. Int. J. Food Microbiol. 2018, 266, 8–13. [Google Scholar] [CrossRef]
- Makalatia, K.; Kakabadze, E.; Bakuradze, N.; Grdzelishvili, N.; Stamp, B.; Herman, E.; Tapinos, A.; Coffey, A.; Lee, D.; Papadopoulos, N.; et al. Investigation of Salmonella Phage–Bacteria Infection Profiles: Network Structure Reveals a Gradient of Target-Range from Generalist to Specialist Phage Clones in Nested Subsets. Viruses 2021, 13, 1261. [Google Scholar] [CrossRef]
- Pelyuntha, W.; Ngasaman, R.; Yingkajorn, M.; Chukiatsiri, K.; Benjakul, S.; Vongkamjan, K. Isolation and Characterization of Potential Salmonella Phages Targeting Multidrug-Resistant and Major Serovars of Salmonella Derived From Broiler Production Chain in Thailand. Front. Microbiol. 2021, 12, 662461. [Google Scholar] [CrossRef]
- Yildirim, Z.; Sakin, T.; Çoban, F. Isolation of lytic bacteriophages infecting Salmonella Typhimurium and Salmonella Enteritidis. Acta Biol. Hung. 2018, 69, 350–369. [Google Scholar] [CrossRef]
- Dion, M.B.; Oechslin, F.; Moineau, S. Phage diversity, genomics and phylogeny. Nat. Rev. Microbiol. 2020, 18, 125–138. [Google Scholar] [CrossRef]
- Rashid, S.J.; Barylski, J.; Hargreaves, K.R.; Millard, A.A.; Vinner, G.K.; Clokie, M.R.J. Two Novel Myoviruses from the North of Iraq Reveal Insights into Clostridium difficile Phage Diversity and Biology. Viruses 2016, 8, 310. [Google Scholar] [CrossRef]
- Abedon, S.T. Phage Therapy: Eco-Physiological Pharmacology. Scientifica 2014, 2014, 581639. [Google Scholar] [CrossRef]
- Sinha, S.; Grewal, R.K.; Roy, S. Chapter Three—Modeling Bacteria–Phage Interactions and Its Implications for Phage Therapy. In Advances in Applied Microbiology; Sariaslani, S., Gadd, G.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 103–141. Available online: https://www.sciencedirect.com/science/article/pii/S0065216418300054 (accessed on 1 December 2022).
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S.; et al. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 50, D20–D26. [Google Scholar] [CrossRef]
- Turner, D.; Kropinski, A.; Adriaenssens, E. A Roadmap for Genome-Based Phage Taxonomy. Viruses 2021, 13, 506. [Google Scholar] [CrossRef]
- Jolenerr. New Tool: Linear Genome Plot. 2020. Available online: https://cpt.tamu.edu/new-tool-linear-genome-plot-2/ (accessed on 9 December 2022).
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Marçais, G.; Delcher, A.L.; Phillippy, A.; Coston, R.; Salzberg, S.; Zimin, A. MUMmer4: A fast and versatile genome alignment system. PLoS Comput. Biol. 2018, 14, e1005944. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modelling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y.; Willias, S.P.; Morsy, M.K.; Abdelnabby, H.E.; Liu, J.; Wang, X.; Li, J. Isolation, Characterization, and Application of Bacteriophage LPSE1 Against Salmonella enterica in Ready to Eat (RTE) Foods. Front. Microbiol. 2018, 9, 1046. [Google Scholar] [CrossRef]
- Uchiyama, J.; Takemura, I.; Satoh, M.; Kato, S.-I.; Ujihara, T.; Akechi, K.; Matsuzaki, S.; Daibata, M. Improved Adsorption of an Enterococcus faecalis Bacteriophage ΦEF24C with a Spontaneous Point Mutation. PLoS ONE 2011, 6, e26648. [Google Scholar] [CrossRef]
- Fortier, L.-C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Bao, H.; Zhang, H.; Wang, R. Isolation and characterization of bacteriophages of Salmonella enterica serovar Pullorum. Poult. Sci. 2011, 90, 2370–2377. [Google Scholar] [CrossRef] [PubMed]
- Ateba, C.N.; Akindolire, M.A. Isolation and Characterisation of Bacteriophages with Lytic Activity Against Virulent Escherichia coli O157:H7: Potential Bio-Control Agents. BIOLOGY. January 2019. Available online: http://www.preprints.org/manuscript/201901.0132/v1 (accessed on 5 February 2020).
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid Interface Sci. 2017, 249, 100–133. [Google Scholar] [CrossRef] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Jamal, M.; Hussain, T.; Rajanna Das, C.; Andleeb, S. Isolation and Characterization of a Myoviridae MJ1 Bacteriophage Against Multi-Drug Resistant Escherichia coli 3. Jundishapur J. Microbiol. 2015, 8, e25917. [Google Scholar] [CrossRef]
- Sofy, A.R.; El-Dougdoug, N.K.; Refaey, E.E.; Dawoud, R.A.; Hmed, A.A. Characterization and Full Genome Sequence of Novel KPP-5 Lytic Phage against Klebsiella pneumoniae Responsible for Recalcitrant Infection. Biomedicines 2021, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Yukgehnaish, K.; Rajandas, H.; Parimannan, S.; Manickam, R.; Marimuthu, K.; Petersen, B.; Clokie, M.R.J.; Millard, A.; Sicheritz-Pontén, T. PhageLeads: Rapid Assessment of Phage Therapeutic Suitability Using an Ensemble Machine Learning Approach. Viruses 2022, 14, 342. [Google Scholar] [CrossRef] [PubMed]
- D’Accolti, M.; Soffritti, I.; Mazzacane, S.; Caselli, E. Bacteriophages as a Potential 360-Degree Pathogen Control Strategy. Microorganisms 2021, 9, 261. [Google Scholar] [CrossRef]
- Cuervo, A.; Pulido-Cid, M.; Chagoyen, M.; Arranz, R.; González-García, V.A.; Garcia-Doval, C.; Caston, J.; Valpuesta, J.; van Raaij, M.J.; Martín-Benito, J.; et al. Structural Characterization of the Bacteriophage T7 Tail Machinery. J. Biol. Chem. 2013, 288, 26290–26299. [Google Scholar] [CrossRef]
- de Leeuw, M.; Baron, M.; Ben David, O.; Kushmaro, A. Molecular Insights into Bacteriophage Evolution toward Its Host. Viruses 2020, 12, 1132. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, R.; Xu, M.; Liu, Y.; Zhu, X.; Qiu, J.; Liu, Q.; He, P.; Li, Q. A Novel Polysaccharide Depolymerase Encoded by the Phage SH-KP152226 Confers Specific Activity Against Multidrug-Resistant Klebsiella pneumoniae via Biofilm Degradation. Front. Microbiol. 2019, 10, 2768. [Google Scholar] [CrossRef] [Green Version]
- Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil. In Bacteriophages; Clokie, M.R.J., Kropinski, A.M., Eds.; Methods in Molecular Biology book series; Humana Press: Totowa, NJ, USA, 2009; Volume 501, pp. 15–21. Available online: http://link.springer.com/10.1007/978-1-60327-164-6_2 (accessed on 27 April 2020).
- Bao, H.; Zhang, P.; Zhang, H.; Zhou, Y.; Zhang, L.; Wang, R. Bio-Control of Salmonella Enteritidis in Foods Using Bacteriophages. Viruses 2015, 7, 4836–4853. [Google Scholar] [CrossRef]
- Jakočiūnė, D.; Moodley, A. A Rapid Bacteriophage DNA Extraction Method. Methods Protoc. 2018, 1, 27. [Google Scholar] [CrossRef]
- Amarillas, L.; Rubí-Rangel, L.; Chaidez, C.; González-Robles, A.; Lightbourn-Rojas, L.; León-Félix, J. Isolation and Characterization of phiLLS, a Novel Phage with Potential Biocontrol Agent against Multidrug-Resistant Escherichia coli. Front Microbiol. 2017, 8, 1355. [Google Scholar] [CrossRef] [PubMed]
- Minas, K.; McEwan, N.R.; Newbold, C.J.; Scott, K.P. Optimization of a high-throughput CTAB-based protocol for the extraction of qPCR-grade DNA from rumen fluid, plant and bacterial pure cultures. FEMS Microbiol. Lett. 2011, 325, 162–169. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinforma. Oxf. Engl. 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.; Nikolenko, I.S.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Shen, A.; Millard, A. Phage Genome Annotation: Where to Begin and End. PHAGE 2021, 2, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinforma. Oxf. Engl. 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Kleinheinz, K.A.; Joensen, K.G.; Larsen, M.V. Applying the ResFinder and VirulenceFinder web-services for easy identification of acquired antibiotic resistance and E. coli virulence genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 2014, 4, e27943. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria/Phage | PRF-SP1 | PRF-SP2 | PRF-SP3 | PRF-SP4 | PRF-SP5 | PRF-SP6 | PRF-SP7 | PRF-SP8 | PRF-SP9 | PRF-SP10 | PRF-SP11 | PRF-SP12 | PRF-SP13 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. Paratyphi A CS/SPA18 | ++ | ++ | ++ | + | + | + | ++ | ++ | ++ | ++ | ++ | − | − |
| S. Paratyphi B CS/SPB18 | ++ | − | ++ | ++ | ++ | − | − | − | − | − | − | − | − |
| S. Paratyphi C CS/SPC18 | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | − | − |
| S. Typhi B37239/20 | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | − | − |
| S. Typhi B41529/20 | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | ++ | − | − |
| S. Typhimurium CS/STYM18 | ++ | − | ++ | ++ | + | − | − | − | − | − | − | − | − |
| S. Enteritidis CS/SEN18 | ++ | − | ++ | ++ | ++ | − | − | − | − | − | − | ++ | ++ |
| E. coli O145:H28 strain RM12581 | ++ | − | ++ | − | − | − | − | − | − | − | − | − | − |
| Pseudomonas aeruginosa | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Proteus mirabilis | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Klebsiella pneumoniae | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Vibrio cholerae El Tor | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Vibrio cholerae O139 Bengal | − | − | − | − | − | − | − | − | − | − | − | − | − |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mutusamy, P.; Banga Singh, K.K.; Su Yin, L.; Petersen, B.; Sicheritz-Ponten, T.; Clokie, M.R.J.; Loke, S.; Millard, A.; Parimannan, S.; Rajandas, H. Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest. Int. J. Mol. Sci. 2023, 24, 3678. https://doi.org/10.3390/ijms24043678
Mutusamy P, Banga Singh KK, Su Yin L, Petersen B, Sicheritz-Ponten T, Clokie MRJ, Loke S, Millard A, Parimannan S, Rajandas H. Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest. International Journal of Molecular Sciences. 2023; 24(4):3678. https://doi.org/10.3390/ijms24043678
Chicago/Turabian StyleMutusamy, Prasanna, Kirnpal Kaur Banga Singh, Lee Su Yin, Bent Petersen, Thomas Sicheritz-Ponten, Martha R. J. Clokie, Stella Loke, Andrew Millard, Sivachandran Parimannan, and Heera Rajandas. 2023. "Phenotypic Characterization and Comparative Genomic Analysis of Novel Salmonella Bacteriophages Isolated from a Tropical Rainforest" International Journal of Molecular Sciences 24, no. 4: 3678. https://doi.org/10.3390/ijms24043678