

Investigating the Resistance Mechanism of Wheat Varieties to Fusarium Head Blight Using Comparative Metabolomics
 and
and
Abstract
:1. Introduction
2. Results
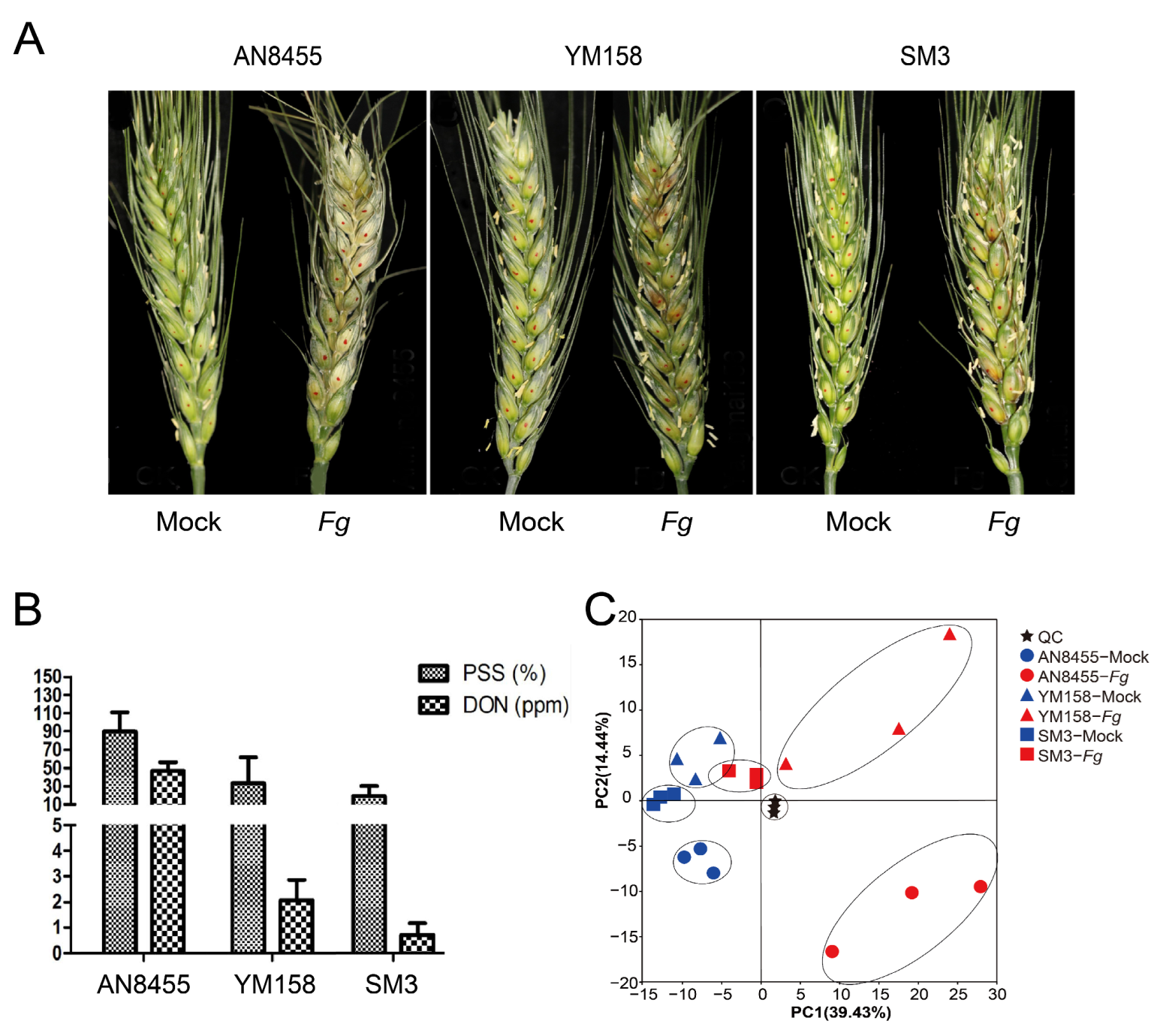
2.1. Evaluation of FHB Resistance among Wheat Varieties
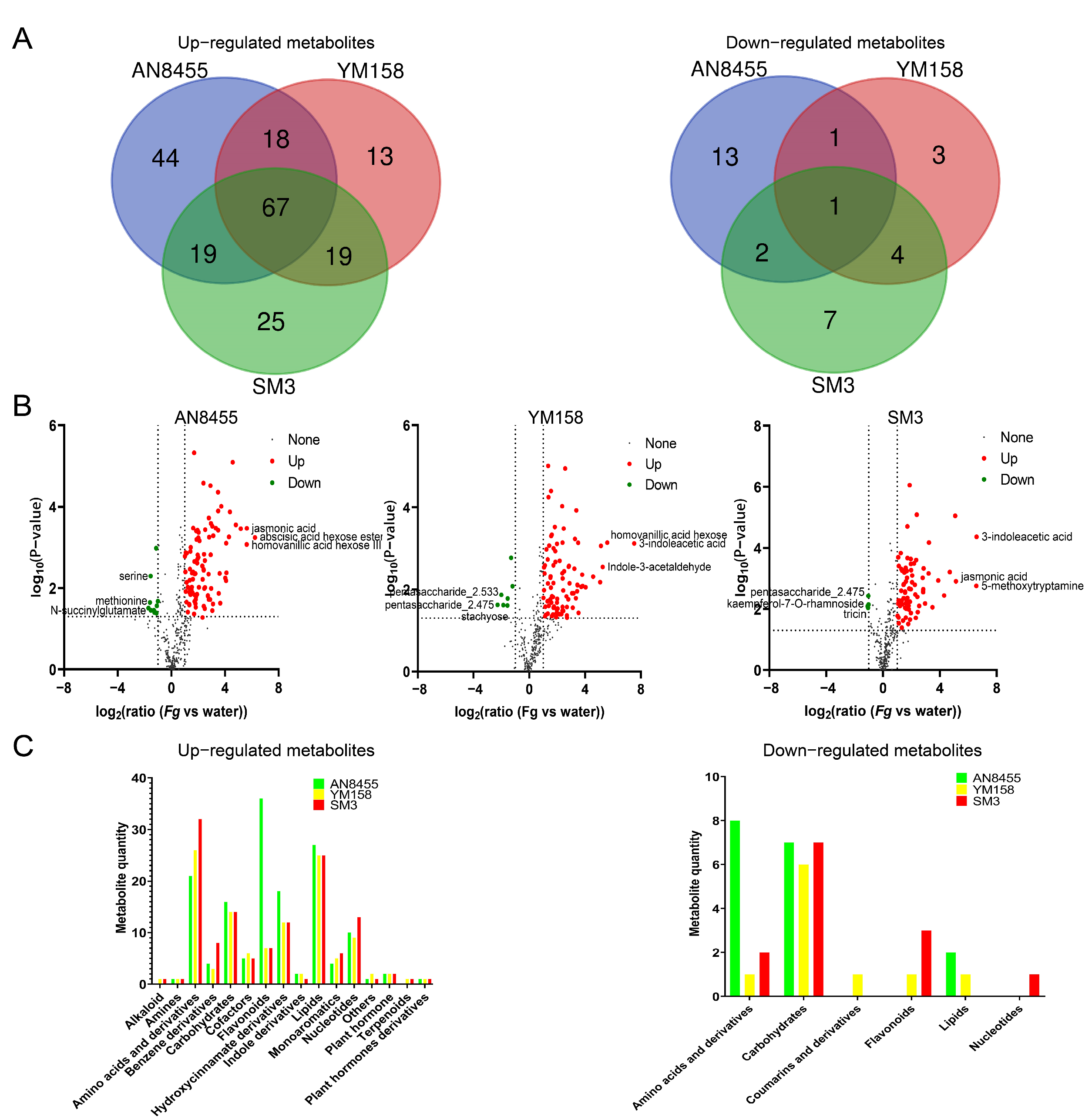
2.2. Dynamic Changes in Metabolites among Different Wheat Varieties during FHB Infection
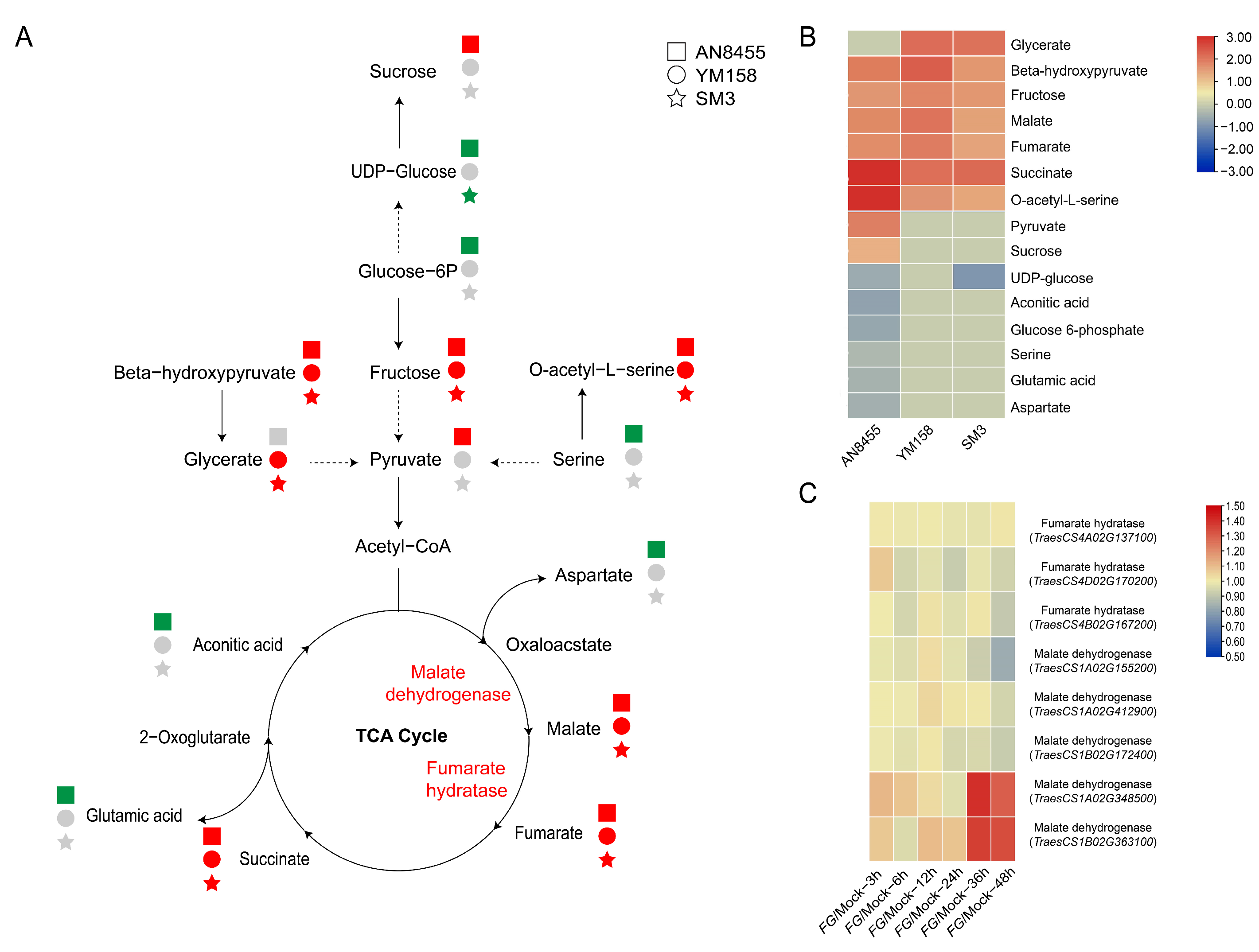
2.3. Response of Metabolites in Catabolic Processes
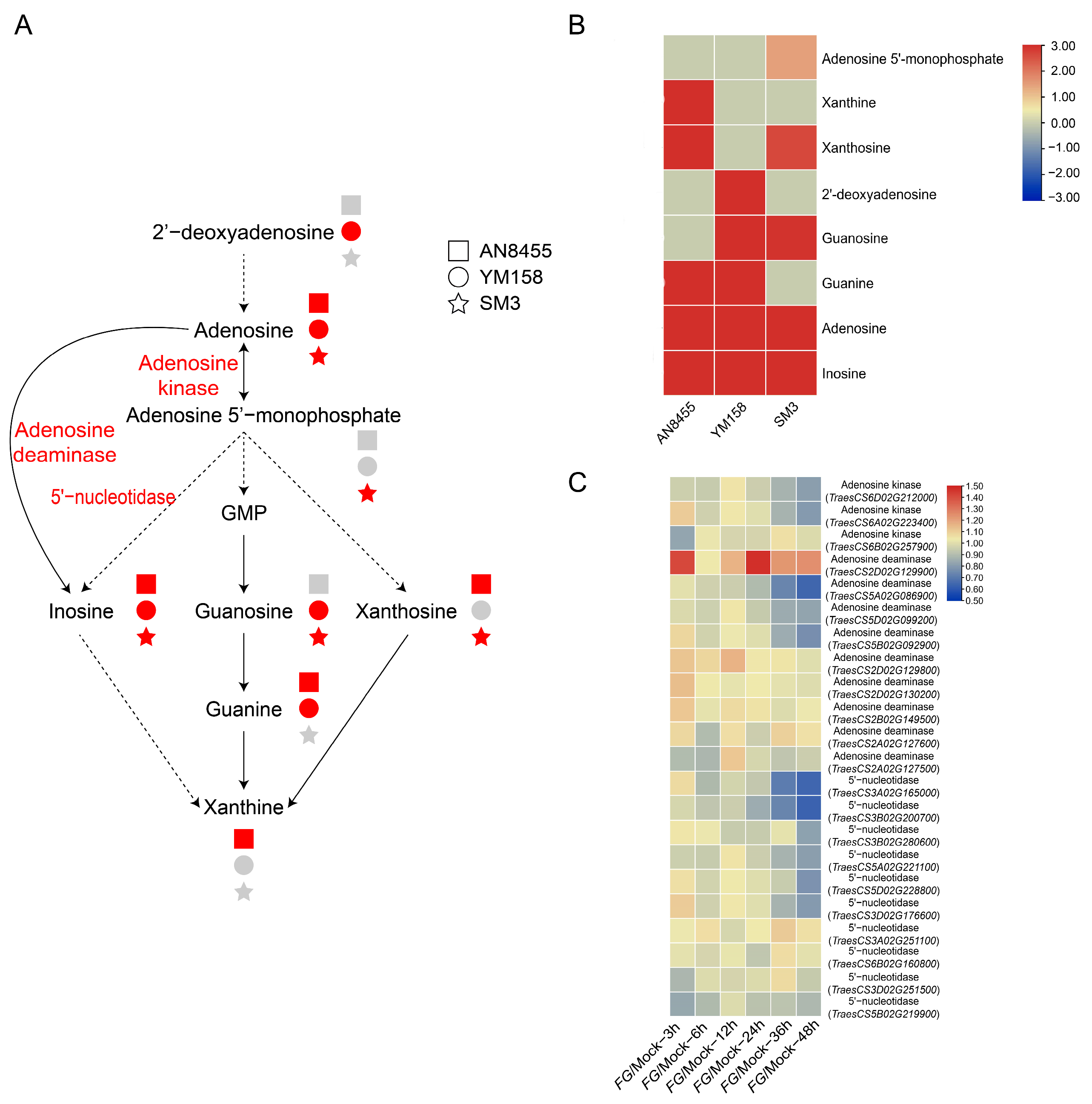
2.4. Response of Nucleotide Metabolism
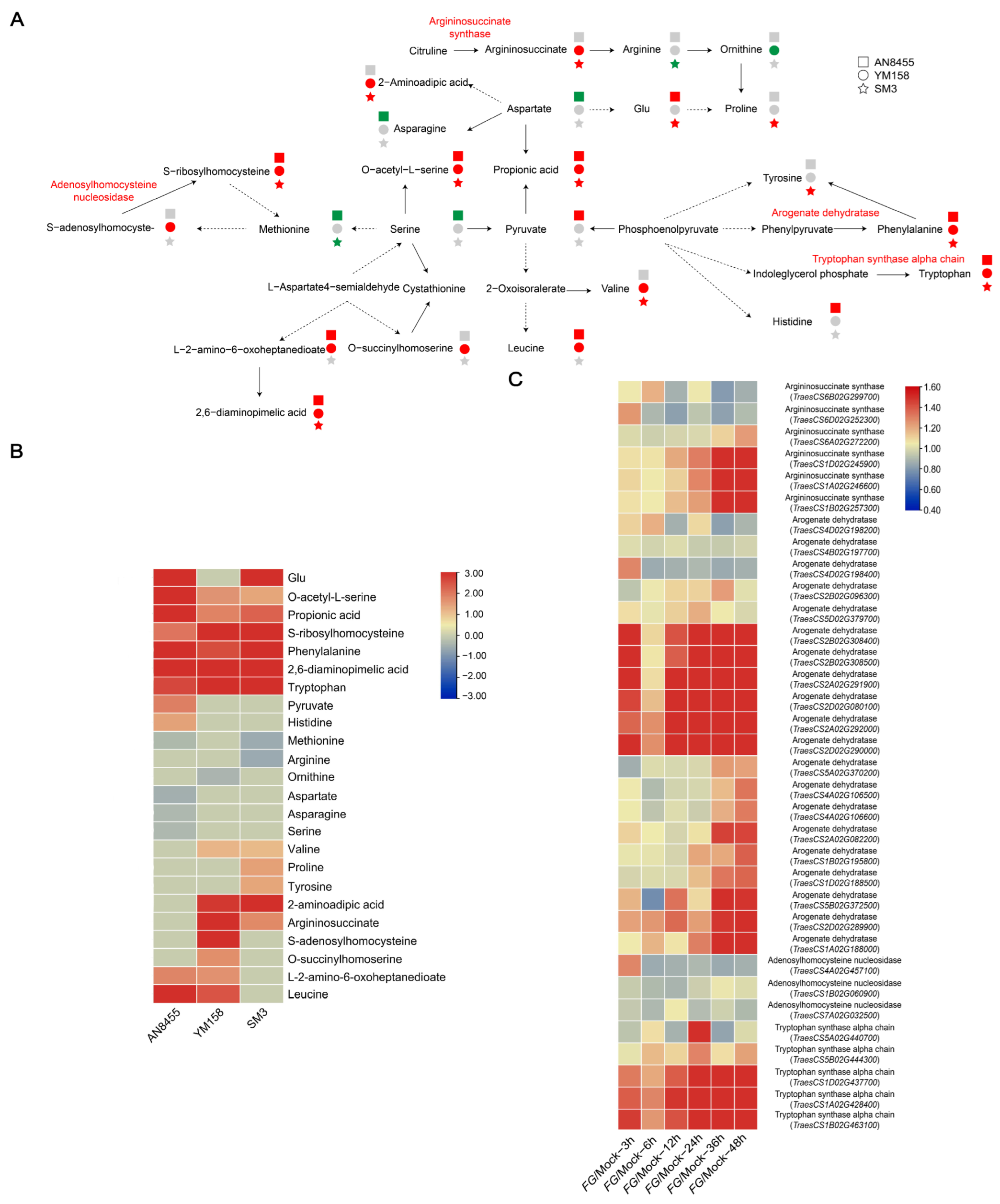
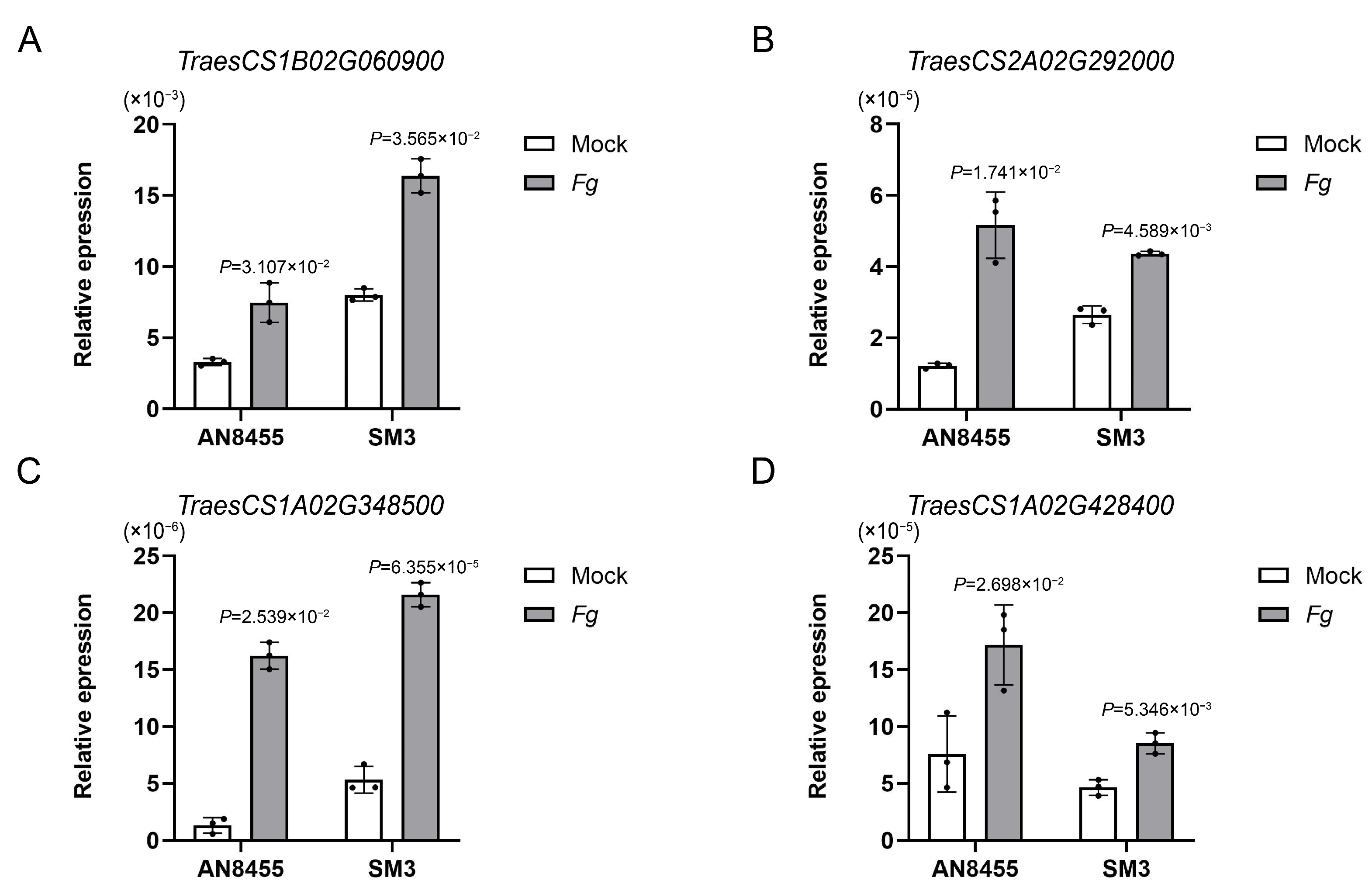
2.5. Response of Amino Acid and Derivatives
2.6. Response of Antifungal Metabolites
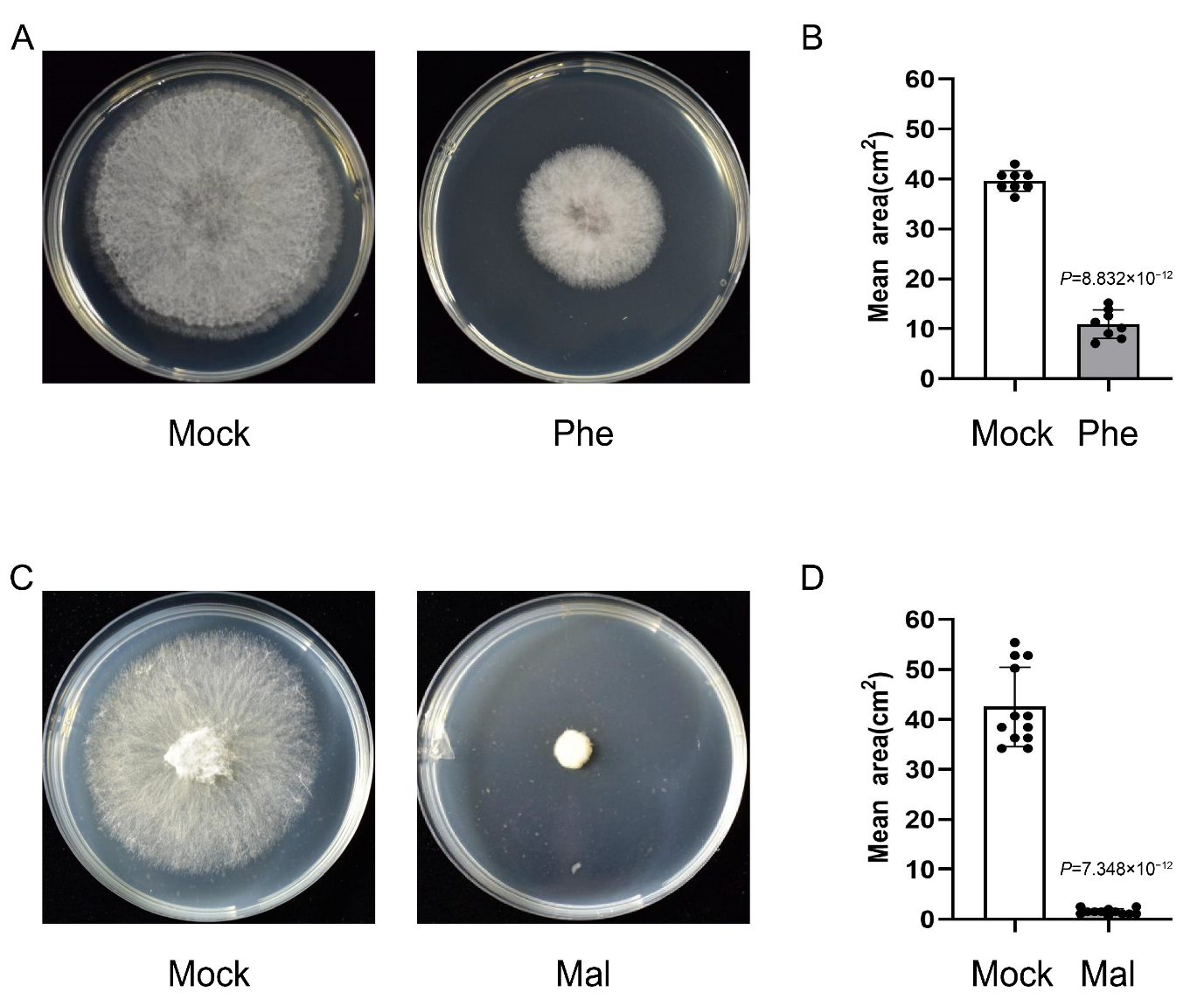
2.7. Phenylalanine and Malate Suppress the Growth of F. graminearum
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Compound Extraction
4.3. Chromatography and Mass Spectrometry
4.4. Identification of Compounds and Extraction of Peak Area
4.5. Data Analysis
4.6. Fungistasis Test
4.7. qRT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xiao, J.; Liu, B.; Yao, Y.; Guo, Z.; Jia, H.; Kong, L.; Zhang, A.; Ma, W.; Ni, Z.; Xu, S.; et al. Wheat genomic study for genetic improvement of traits in China. Sci. China Life Sci. 2022, 65, 1718–1775. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Nat. Ecol. Evol. 2019, 3, 430–439. [Google Scholar] [CrossRef] [PubMed]
- Boutigny, A.L.; Richard-Forget, F.; Barreau, C. Natural mechanisms for cereal resistance to Fusarium mycotoxins accumula-tion. Eur. J. Plant Pathol. 2008, 121, 411–423. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Management and resistance in wheat and barley to fusarium head blight. Annu. Rev. Phytopathol. 2004, 42, 135–161. [Google Scholar] [CrossRef]
- Zheng, T.; Hua, C.; Li, L.; Sun, Z.; Yuan, M.; Bai, G.; Humphreys, G.; Li, T. Integration of meta-QTL discovery with omics: Towards a molecular breeding platform for improving wheat resistance to Fusarium head blight. Crop. J. 2020, 9, 739–749. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Q.; Wang, G.; Zhang, X.; Liu, H.; Jiang, C. Combatting Fusarium head blight: Advances in molecular interactions between Fusarium graminearum and wheat. Phytopathol. Res. 2022, 4, 1–16. [Google Scholar] [CrossRef]
- Su, Z.; Bernardo, A.; Tian, B.; Chen, H.; Wang, S.; Ma, H.; Cai, S.; Liu, D.; Zhang, D.; Li, T.; et al. A deletion mutation in TaHRC confers Fhb1 resistance to Fusarium head blight in wheat. Nat. Genet. 2019, 51, 1099–1105. [Google Scholar] [CrossRef]
- Li, G.; Zhou, J.; Jia, H.; Gao, Z.; Fan, M.; Luo, Y.; Zhao, P.; Xue, S.; Li, N.; Yuan, Y.; et al. Mutation of a histidine-rich calcium-binding-protein gene in wheat confers resistance to Fusarium head blight. Nat. Genet. 2019, 51, 1106–1112. [Google Scholar] [CrossRef]
- Wang, H.W.; Sun, S.L.; Ge, W.Y.; Zhao, L.F.; Hou, B.Q.; Wang, K.; Lyu, Z.F.; Chen, L.Y.; Xu, S.S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, 844. [Google Scholar] [CrossRef]
- Kruger, W.M.; Pritsch, C.; Chao, S.; Muehlbauer, G.J. Functional and Comparative Bioinformatic Analysis of Expressed Genes from Wheat Spikes Infected with Fusarium graminearum. Mol. Plant-Microbe Interact. 2002, 15, 445–455. [Google Scholar] [CrossRef] [Green Version]
- Trail, F.; Xu, J.R.; San Miguel, P.; Halgren, R.G.; Kistler, H.C. Analysis of expressed sequence tags from Gibberella zeae (an-amorph Fusarium graminearum). Fungal Genet. Biol. 2003, 38, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, A.; Bai, G.; Guo, P.; Xiao, K.; Guenzi, A.C.; Ayoubi, P. Fusarium graminearum-induced changes in gene expression between Fusarium head blight-resistant and susceptible wheat cultivars. Funct. Integr. Genom. 2006, 7, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Jin, X.; Jia, X.; Wang, H.; Cao, A.; Zhao, W.; Pei, H.; Xue, Z.; He, L.; Chen, Q.; et al. Transcriptome-based discovery of pathways and genes related to resistance against Fusarium head blight in wheat landrace Wangshuibai. BMC Genom. 2013, 14, 197. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, A.; Manickavelu, A.; Kajihara, D.; Nakagawa, H.; Ban, T. Altered Gene Expression Profiles of Wheat Genotypes against Fusarium Head Blight. Toxins 2015, 7, 604–620. [Google Scholar] [CrossRef]
- Gao, T.; Chen, J.; Shi, Z. Fusarium graminearum pyruvate dehydrogenase kinase 1 (FgPDK1) Is Critical for Conidiation, Mycelium Growth, and Pathogenicity. PLoS ONE 2016, 11, e0158077. [Google Scholar] [CrossRef]
- Hofstad, A.N.; Nussbaumer, T.; Akhunov, E.; Shin, S.; Kugler, K.G.; Kistler, H.C.; Mayer, K.F.; Muehlbauer, G.J. Examining the transcriptional response in wheat near-isogenic lines to infection and deoxynivalenol treatment. Plant Genome 2016, 9, 1–15. [Google Scholar] [CrossRef]
- Chetouhi, C.; Bonhomme, L.; Zuber, P.L.; Cambon, F.; Pelletier, S.; Renou, J.-P.; Langin, T. Transcriptome dynamics of a susceptible wheat upon Fusarium head blight reveals that molecular responses to Fusarium graminearum infection fit over the grain development processes. Funct. Integr. Genom. 2016, 16, 183–201. [Google Scholar] [CrossRef]
- Gunnaiah, R.; Kushalappa, A.C. Metabolomics deciphers the host resistance mechanisms in wheat cultivar Sumai-3, against trichothecene producing and non-producing isolates of Fusarium graminearum. Plant Physiol. Biochem. 2014, 83, 40–50. [Google Scholar] [CrossRef]
- Gauthier, L.; Atanasova-Penichon, V.; Chéreau, S.; Richard-Forget, F. Metabolomics to Decipher the Chemical Defense of Cereals against Fusarium graminearum and Deoxynivalenol Accumulation. Int. J. Mol. Sci. 2015, 16, 24839–24872. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Transcriptomics of cereal-Fusarium graminearum interactions: What we have learned so far. Mol. Plant Pathol. 2018, 19, 764–778. [Google Scholar] [CrossRef] [Green Version]
- Paranidharan, V.P.; Abunada, A.N.; Hamzehzarghani, H.H.; Kushalappa, A.C.K.C.; Mamer, O.M.; Dion, Y.D.; Rioux, S.R.; Comeau, A.C.; Choiniere, L.C. Resistance-related metabolites in wheat against Fusarium graminearum. Botany 2008, 86, 1168–1179. [Google Scholar] [CrossRef]
- Kushalappa, A.C.; Gunnaiah, R. Metabolo-proteomics to discover plant biotic stress resistance genes. Trends Plant Sci. 2013, 18, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Schauer, N.; Fernie, A.R. Plant metabolomics: Towards biological function and mechanism. Trends Plant Sci. 2006, 11, 508–516. [Google Scholar] [CrossRef] [PubMed]
- Bollina, V.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S. Identification of metabolites related to mechanisms of resistance in barley against Fusarium graminearum, based on mass spectrometry. Plant Mol. Biol. 2011, 77, 355–370. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.H.; Chen, L.F.; Shaner, G. Breeding for resistance to Fusarium head blight of wheat in China. In Fusarium Head Blight of Wheat and Barley; Leonard, K.J., Ed.; APS Press: St. Paul, MN, USA, 2003; pp. 296–317. [Google Scholar]
- Owen, O.E.; Kalhan, S.C.; Hanson, R.W. The key role of anaplerosis and cataplerosis for citric acid cycle function. J. Biol. Chem. 2002, 277, 30409–30412. [Google Scholar] [CrossRef]
- Garcia, D.F.; Wang, Z.; Guan, J.; Yin, L.; Geng, S.; Li, A.; Mao, L. WheatGene: A genomics database for common wheat and its related species. Crop. J. 2021, 9, 1486–1491. [Google Scholar] [CrossRef]
- Zhao, L.; Su, P.; Hou, B.; Wu, H.; Fan, Y.; Li, W.; Zhao, J.; Ge, W.; Xu, S.; Wu, S.; et al. The Black Necrotic Lesion Enhanced Fusarium graminearum Resistance in Wheat. Front. Plant Sci. 2022, 13, 926621. [Google Scholar] [CrossRef]
- Zhao, P.; Gu, S.; Han, C.; Lu, Y.; Ma, C.; Tian, J.; Bi, J.; Deng, Z.; Wang, Q.; Xu, Q. Targeted and Untargeted Metabolomics Profiling of Wheat Reveals Amino Acids Increase Resistance to Fusarium Head Blight. Front. Plant Sci. 2021, 12, 762605. [Google Scholar] [CrossRef]
- Castro-Moretti, F.R.; Gentzel, I.N.; Mackey, D.; Alonso, A.P. Metabolomics as an Emerging Tool for the Study of Plant–Pathogen Interactions. Metabolites 2020, 10, 52. [Google Scholar] [CrossRef]
- Whitney, K.; Gracia−Gomez, G.; Anderson, J.A.; Simsek, S. Time Course Metabolite Profiling of Fusarium Head Blight−Infected Hard Red Spring Wheat Using Ultra−High−Performance Liquid Chromatography Coupled with Quadrupole Time of Flight/MS. J. Agric. Food Chem. 2022, 70, 4152–4163. [Google Scholar] [CrossRef]
- Browne, R.A.; Brindle, K.M. 1H NMR-based metabolite profiling as a potential selection tool for breeding passive resistance against Fusarium head blight (FHB) in wheat. Mol. Plant Pathol. 2007, 8, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Zhou, Y.; Shen, Y.; Sun, Z.; Li, L.; Li, T. Linking Multi-Omics to Wheat Resistance Types to Fusarium Head Blight to Reveal the Underlying Mechanisms. Int. J. Mol. Sci. 2022, 23, 2280. [Google Scholar] [CrossRef] [PubMed]
- Bollina, V.; Kumaraswamy, G.K.; Kushalappa, A.C.; Choo, T.M.; Dion, Y.; Rioux, S.; Faubert, D.; Hamzehzarghani, H. Mass spectrometry-based metabolomics application to identify quantitative resistance-related metabolites in barley against Fusarium head blight. Mol. Plant Pathol. 2010, 11, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Tauzin, A.S.; Giardina, T. Sucrose and invertases, a part of the plant defense response to the biotic stresses. Front. Plant Sci. 2014, 5, 293. [Google Scholar] [CrossRef]
- Qian, Y.; Tan, D.-X.; Reiter, R.J.; Shi, H. Comparative metabolomic analysis highlights the involvement of sugars and glycerol in melatonin-mediated innate immunity against bacterial pathogen in Arabidopsis. Sci. Rep. 2015, 5, 15815. [Google Scholar] [CrossRef]
- Arts, R.J.; Novakovic, B.; ter Horst, R.; Carvalho, A.; Bekkering, S.; Lachmandas, E.; Rodrigues, F.; Silvestre, R.; Cheng, S.-C.; Wang, S.-Y.; et al. Glutaminolysis and Fumarate Accumulation Integrate Immunometabolic and Epigenetic Programs in Trained Immunity. Cell Metab. 2016, 24, 807–819. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Lakshmanan, V.; Kobayashi, Y.; Asai, M.; Iuchi, S.; Kobayashi, M.; Bais, H.P.; Koyama, H. Overexpression of AtALMT1 in the Arabidopsis thaliana ecotype Columbia results in enhanced Al-activated malate excretion and beneficial bac-terium recruitment. Plant Signal Behav. 2013, 8, e25565. [Google Scholar] [CrossRef]
- Kumar, V.; Hatan, E.; Bar, E.; Davidovich-Rikanati, R.; Doron-Faigenboim, A.; Spitzer-Rimon, B.; Elad, Y.; Alkan, N.; Lewinsohn, E.; Oren-Shamir, M. Phenylalanine increases chrysanthemum flower immunity against Botrytis cinerea attack. Plant J. 2020, 104, 226–240. [Google Scholar] [CrossRef]
- Warth, B.; Parich, A.; Bueschl, C.; Schoefbeck, D.; Neumann, N.K.N.; Kluger, B.; Schuster, K.; Krska, R.; Adam, G.; Lemmens, M.; et al. GC–MS-based targeted metabolic profiling identifies changes in the wheat metabolome following de-oxynivalenol treatment. Metabolomics 2015, 11, 722–738. [Google Scholar] [CrossRef]
- Grayer, R.J.; Harborne, J.B. A survey of antifungal compounds from higher plants, 1982–1993. Phytochemistry 1994, 37, 19–42. [Google Scholar] [CrossRef]
- Reichling, J. Plant-microbe interactions and secondary metabolites with antibacterial, antifungal and antiviral properties. Annu. Plant Rev. 2010, 39, 214–347. [Google Scholar]
- Bednarek, P.; Pislewska-Bednarek, M.; Svatos, A.; Schneider, B.; Doubsky, J.; Mansurova, M.; Humphry, M.; Consonni, C.; Panstruga, R.; Sanchez-Vallet, A.; et al. A Glucosinolate Metabolism Pathway in Living Plant Cells Mediates Broad-Spectrum Antifungal Defense. Science 2009, 323, 101–106. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Zhang, X.; Zhang, Y.; Ahmad, D.; Wu, L.; Jiang, P.; Ma, H.X. Molecular characterization and expression of PFT, an FHB resistance gene at the Fhb1 QTL in wheat. Phytopathology 2018, 108, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Asselbergh, B.; De Vleesschauwer, D.; Höfte, M. Global switches and fine-tuning—ABA modulates plant pathogen defense. Mol. Plant Microbe Int. 2008, 21, 709–719. [Google Scholar] [CrossRef]
- Mauch-Mani, B.; Mauch, F. The role of abscisic acid in plant–pathogen interactions. Curr. Opin. Plant Biol. 2005, 8, 409–414. [Google Scholar] [CrossRef]
- Wang, L.; Li, Q.; Liu, Z.; Surendra, A.; Pan, Y.; Li, Y.; Zaharia, L.I.; Ouellet, T.; Fobert, P.R. Integrated transcriptome and hormone profiling highlight the role of multiple phytohormone pathways in wheat resistance against fusarium head blight. PLoS ONE 2018, 13, e0207036. [Google Scholar] [CrossRef]
- Makandar, R.; Nalam, V.J.; Lee, H.; Trick, H.N.; Dong, Y.; Shah, J. Salicylic Acid Regulates Basal Resistance to Fusarium Head Blight in Wheat. Mol. Plant-Microbe Interact. 2012, 25, 431–439. [Google Scholar] [CrossRef]
- Brauer, E.K.; Rocheleau, H.; Balcerzak, M.; Pan, Y.; Fauteux, F.; Liu, Z.; Wang, L.; Zheng, W.; Ouellet, T. Transcriptional and hormonal profiling of Fusarium graminearum-infected wheat reveals an association between auxin and susceptibility. Physiol. Mol. Plant Pathol. 2019, 107, 33–39. [Google Scholar] [CrossRef]
- Xia, X.; Zhang, X.; Zhang, Y.; Wang, L.; An, Q.; Tu, Q.; Wu, L.; Jiang, P.; Zhang, P.; Yu, L.; et al. Characterization of the WAK Gene Family Reveals Genes for FHB Resistance in Bread Wheat (Triticum aestivum L.). Int. J. Mol. Sci. 2022, 23, 7157. [Google Scholar] [CrossRef]
- Evans, A.M.; DeHaven, C.D.; Barrett, T.; Mitchell, M.; Milgram, E. Integrated, Nontargeted Ultrahigh Performance Liquid Chromatography/Electrospray Ionization Tandem Mass Spectrometry Platform for the Identification and Relative Quantification of the Small-Molecule Complement of Biological Systems. Anal. Chem. 2009, 81, 6656–6667. [Google Scholar] [CrossRef]
- Lisec, J.; Schauer, N.; Kopka, J.; Willmitzer, L.; Fernie, A.R. Gas chromatography mass spectrometry–based metabolite profiling in plants. Nat. Protoc. 2006, 1, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Rao, J.; Song, Y.; Chan, S.A.; Tohge, T.; Cui, B.; Lin, H.; Fernie, A.; Zhang, D.; Shi, J. Dissection of flag leaf metabolic shifts and their relationship with those occurring simultaneously in developing seed by application of non-targeted metabo-lomics. PloS ONE 2020, 15, e0227577. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Zhao, H.; Wang, W.; Xu, M.; Shi, J.; Nie, X.; Yang, G. Identification of Conserved and Diverse Metabolic Shift of the Stylar, Intermediate and Peduncular Segments of Cucumber Fruit during Development. Int. J. Mol. Sci. 2018, 19, 135. [Google Scholar] [CrossRef] [PubMed]
- Horai, H.; Arita, M.; Kanaya, S.; Nihei, Y.; Ikeda, T.; Suwa, K.; Ojima, Y.; Tanaka, K.; Tanaka, S.; Aoshima, K.; et al. MassBank: A public repository for sharing mass spectral data for life sciences. J. Mass Spectrom. 2010, 45, 703–714. [Google Scholar] [CrossRef]
- Guijas, C.; Montenegro-Burke, J.R.; Domingo-Almenara, X.; Palermo, A.; Warth, B.; Hermann, G.; Koellensperger, G.; Huan, T.; Uritboonthai, W.; Aisporna, A.E.; et al. METLIN: A Technology Platform for Identifying Knowns and Unknowns. Anal. Chem. 2018, 90, 3156–3164. [Google Scholar] [CrossRef]
- He, H.; Hao, X.; Zhou, W.; Shi, N.; Feng, J.; Han, L. Identification of antimicrobial metabolites produced by a potential bio-control Actinomycete strain A217. J. Appl. Microbiol. 2020, 128, 1143–1152. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Class | Ratio (Fg vs. Water) | ||
|---|---|---|---|---|
| AN8455 | YM158 | SM3 | ||
| agmatine | Amines | 3.22 | 5.21 | 4.53 |
| O-acetyl-L-serine | Amino acids and derivatives | 3.26 | 1.67 | 1.41 |
| S-ribosylhomocysteine | Amino acids and derivatives | 2.07 | 3.55 | 3.06 |
| cystine | Amino acids and derivatives | 1.74 | 3.35 | 3.18 |
| phenylalanine | Amino acids and derivatives | 3.34 | 2.60 | 3.28 |
| S-glutathionyl-cysteine | Amino acids and derivatives | 4.08 | 3.33 | 3.28 |
| N-acetylisoleucine | Amino acids and derivatives | 7.58 | 5.54 | 3.86 |
| spermidine | Amino acids and derivatives | 3.54 | 8.71 | 4.13 |
| N-acetylleucine | Amino acids and derivatives | 11.15 | 5.21 | 5.04 |
| homomethionine | Amino acids and derivatives | 5.49 | 9.42 | 5.19 |
| N-acetylmethionine | Amino acids and derivatives | 7.06 | 9.67 | 7.27 |
| 2,6-diaminopimelic acid | Amino acids and derivatives | 16.24 | 10.78 | 9.24 |
| tryptophan | Amino acids and derivatives | 2.68 | 3.96 | 9.42 |
| 5-methoxytryptamine | Amino acids and derivatives | 23.94 | 16.45 | 95.10 |
| hydroxybenzoic acid hexose II | Benzene derivatives | 3.05 | 2.55 | 1.65 |
| hydroxybenzoic acid hexose I | Benzene derivatives | 1.59 | 1.45 | 1.69 |
| kynurenate | Benzene derivatives | 6.85 | 13.87 | 3.73 |
| methyl-alpha-glucopyranoside | Carbohydrates | 0.41 | 0.34 | 0.55 |
| maleic acid | Carbohydrates | 1.73 | 1.87 | 1.38 |
| fumarate | Carbohydrates | 1.73 | 1.96 | 1.44 |
| malate | Carbohydrates | 1.80 | 2.08 | 1.46 |
| beta-hydroxypyruvate | Carbohydrates | 1.96 | 2.35 | 1.61 |
| fructose | Carbohydrates | 1.63 | 1.83 | 1.62 |
| succinate | Carbohydrates | 3.62 | 2.14 | 2.21 |
| propionic acid | Carbohydrates | 3.09 | 1.87 | 2.30 |
| threonate | Carbohydrates | 3.69 | 3.07 | 2.38 |
| pantothenic acid hexose | Cofactors | 2.73 | 2.52 | 2.55 |
| nicotinamide | Cofactors | 3.58 | 6.49 | 4.85 |
| isonicotinate | Cofactors | 2.61 | 5.12 | 5.36 |
| nicotinate | Cofactors | 12.20 | 14.19 | 5.45 |
| astilbin III | Flavonoids | 2.19 | 3.42 | 1.93 |
| naringenin | Flavonoids | 12.55 | 2.89 | 3.53 |
| feruloylquinic acid II | Hydroxycinnamate derivatives | 1.63 | 1.54 | 1.33 |
| caffeic acid hexose I | Hydroxycinnamate derivatives | 2.34 | 1.53 | 1.61 |
| cinnamic acid | Hydroxycinnamate derivatives | 2.88 | 2.17 | 2.84 |
| feruloylquinic acid III | Hydroxycinnamate derivatives | 5.70 | 3.27 | 2.93 |
| quinate | Hydroxycinnamate derivatives | 4.28 | 2.97 | 3.29 |
| o-coumaric acid | Hydroxycinnamate derivatives | 7.74 | 5.41 | 3.71 |
| coumaric acid hexoside I | Hydroxycinnamate derivatives | 17.02 | 10.38 | 4.72 |
| feruloylquinic acid-hexoside I | Hydroxycinnamate derivatives | 13.24 | 5.94 | 6.92 |
| Indole-3-acetaldehyde | Indole derivatives | 36.12 | 38.41 | 26.02 |
| 3-hydroxy-3-methyl-glutaric acid | Lipids | 2.22 | 1.52 | 1.42 |
| azelaic acid | Lipids | 5.25 | 2.69 | 1.60 |
| 1-LysoPE(16:0) | Lipids | 4.03 | 3.92 | 2.13 |
| 1-LysoPC(16:0) | Lipids | 3.77 | 3.09 | 2.14 |
| 1-LysoPE(18:1) | Lipids | 3.88 | 5.42 | 2.44 |
| 3-beta-D-Galactosyl-sn-glycerol | Lipids | 8.33 | 3.67 | 2.77 |
| 2-LysoPE(16:0) | Lipids | 5.41 | 6.36 | 3.06 |
| 1-LysoPE(18:3) | Lipids | 6.96 | 6.93 | 3.22 |
| 2-LysoPC(16:0) | Lipids | 5.62 | 6.52 | 3.88 |
| 2-palmitoylglycerol | Lipids | 4.96 | 4.47 | 3.95 |
| 1-LysoPC(18:3) | Lipids | 11.28 | 8.41 | 4.15 |
| hydroxyoctadecanedioic acid II | Lipids | 20.66 | 3.32 | 5.83 |
| LPG(16:0) | Lipids | 16.97 | 13.30 | 7.10 |
| homovanillic acid hexose II | Monoaromatics | 1.45 | 2.08 | 1.42 |
| guaiacol hexose-pentose II | Monoaromatics | 3.07 | 2.63 | 1.65 |
| guaiacol hexose-pentose I | Monoaromatics | 1.78 | 4.54 | 3.68 |
| homovanillic acid hexose III | Monoaromatics | 49.11 | 48.28 | 34.02 |
| 5′-deoxy-5′-(methylthio)adenosine | Nucleotides | 1.73 | 2.50 | 1.80 |
| adenosine | Nucleotides | 6.63 | 11.06 | 3.42 |
| succinyladenosine | Nucleotides | 8.74 | 7.38 | 7.10 |
| inosine | Nucleotides | 19.65 | 7.17 | 7.80 |
| thymidine | Nucleotides | 28.04 | 33.95 | 19.80 |
| adipic acid | Organic acids and derivatives | 9.97 | 5.24 | 2.50 |
| dehydrophaseic acid hexose | Others | 1.44 | 1.66 | 1.49 |
| 3-indoleacetic acid | Plant hormone | 17.24 | 184.78 | 96.02 |
| jasmonic acid | Plant hormones | 49.43 | 35.26 | 35.14 |
| abscisic acid hexose ester | Plant hormones derivatives | 75.40 | 23.96 | 14.82 |
| Compound Name | Class | Ratio (Fg vs. Water) | |||
|---|---|---|---|---|---|
| AN8455 | YM158 | SM3 | |||
| Antifungal metabolites | Arginine | Amino acids and derivatives | 0.61 | 0.52 | 0.58 |
| Phenylalanine | Amino acids and derivatives | 3.34 | 2.6 | 3.28 | |
| Spermidine | Amino acids and derivatives | 3.54 | 8.71 | 4.13 | |
| Tryptophan | Amino acids and derivatives | 2.68 | 3.96 | 9.42 | |
| Benzoic acid | Benzene derivatives | 1.25 | 1.05 | 1.22 | |
| Glucosamine | Carbohydrates | 0.81 | 1.06 | 1.22 | |
| Maleic acid | Carbohydrates | 1.73 | 1.87 | 1.38 | |
| Fumarate | Carbohydrates | 1.73 | 1.96 | 1.44 | |
| Fructose | Carbohydrates | 1.63 | 1.83 | 1.62 | |
| Nicotinamide | Cofactors | 3.58 | 6.49 | 4.85 | |
| Riboflavin | Cofactors | 8.64 | 4.94 | 2.36 | |
| Tricin | Flavonoids | 1.36 | 1.69 | 0.48 | |
| Kaempferol-3-O-glucoside II | Flavonoids | 1 | 1.08 | 1.29 | |
| Cinnamic acid | Hydroxycinnamate derivatives | 2.88 | 2.17 | 2.84 | |
| Phosphocholine | Lipids | 1.08 | 1.7 | 1.2 | |
| Uridine | Nucleotides | 3.85 | 4.24 | 3.25 | |
| Immunomodulatory metabolites | Agmatine | Amines | 3.22 | 5.21 | 4.53 |
| Valine | Amino acids and derivatives | 1.13 | 1.2 | 1.15 | |
| Proline | Amino acids and derivatives | 1.18 | 1.63 | 1.5 | |
| Argininosuccinate | Amino acids and derivatives | 1.47 | 3.09 | 1.78 | |
| Gamma-aminobutyrate | Amino acids and derivatives | 5.01 | 3.7 | 3.03 | |
| Glutamic acid | Amino acids and derivatives | 0.46 | 0.81 | 1.15 | |
| Glutamine | Amino acids and derivatives | 0.46 | 0.67 | 0.75 | |
| Amino oxononanoic acid | Amino acids and derivatives | 0.83 | 1.41 | 0.79 | |
| Aspartate | Amino acids and derivatives | 0.5 | 0.88 | 1.21 | |
| Leucine | Amino acids and derivatives | 5.04 | 2.51 | 1.22 | |
| Asparagine | Amino acids and derivatives | 0.35 | 0.75 | 1.08 | |
| Pipecolic acid I | Amino acids and derivatives | 0.58 | 2.76 | 0.92 | |
| M-salicylic acid | Benzene derivatives | 1.08 | 1.15 | 1.12 | |
| Succinate | Carbohydrates | 3.62 | 2.14 | 2.21 | |
| Raffinose | Carbohydrates | 0.57 | 0.43 | 0.75 | |
| Pyruvate | Carbohydrates | 1.92 | 1.09 | 1.18 | |
| Glucose 6-phosphate | Carbohydrates | 0.64 | 1.53 | 1.35 | |
| Guanosine | Nucleotides | 2.74 | 3.88 | 2.93 | |
| Guanine | Nucleotides | 6.62 | 10.89 | 3.03 | |
| Xanthine | Nucleotides | 3.8 | 1.38 | 1.1 | |
| Jasmonic acid | Plant hormones | 49.43 | 35.26 | 35.14 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, Y.; Xia, X.; Ahmad, D.; Wang, Y.; Zhang, X.; Wu, L.; Jiang, P.; Zhang, P.; Yang, X.; Li, G.; et al. Investigating the Resistance Mechanism of Wheat Varieties to Fusarium Head Blight Using Comparative Metabolomics. Int. J. Mol. Sci. 2023, 24, 3214. https://doi.org/10.3390/ijms24043214
Dong Y, Xia X, Ahmad D, Wang Y, Zhang X, Wu L, Jiang P, Zhang P, Yang X, Li G, et al. Investigating the Resistance Mechanism of Wheat Varieties to Fusarium Head Blight Using Comparative Metabolomics. International Journal of Molecular Sciences. 2023; 24(4):3214. https://doi.org/10.3390/ijms24043214
Chicago/Turabian StyleDong, Yifan, Xiaobo Xia, Dawood Ahmad, Yuhua Wang, Xu Zhang, Lei Wu, Peng Jiang, Peng Zhang, Xiujuan Yang, Gang Li, and et al. 2023. "Investigating the Resistance Mechanism of Wheat Varieties to Fusarium Head Blight Using Comparative Metabolomics" International Journal of Molecular Sciences 24, no. 4: 3214. https://doi.org/10.3390/ijms24043214