
The Effect of the Tau Protein on D. melanogaster Lifespan Depends on GSK3 Expression and Sex

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
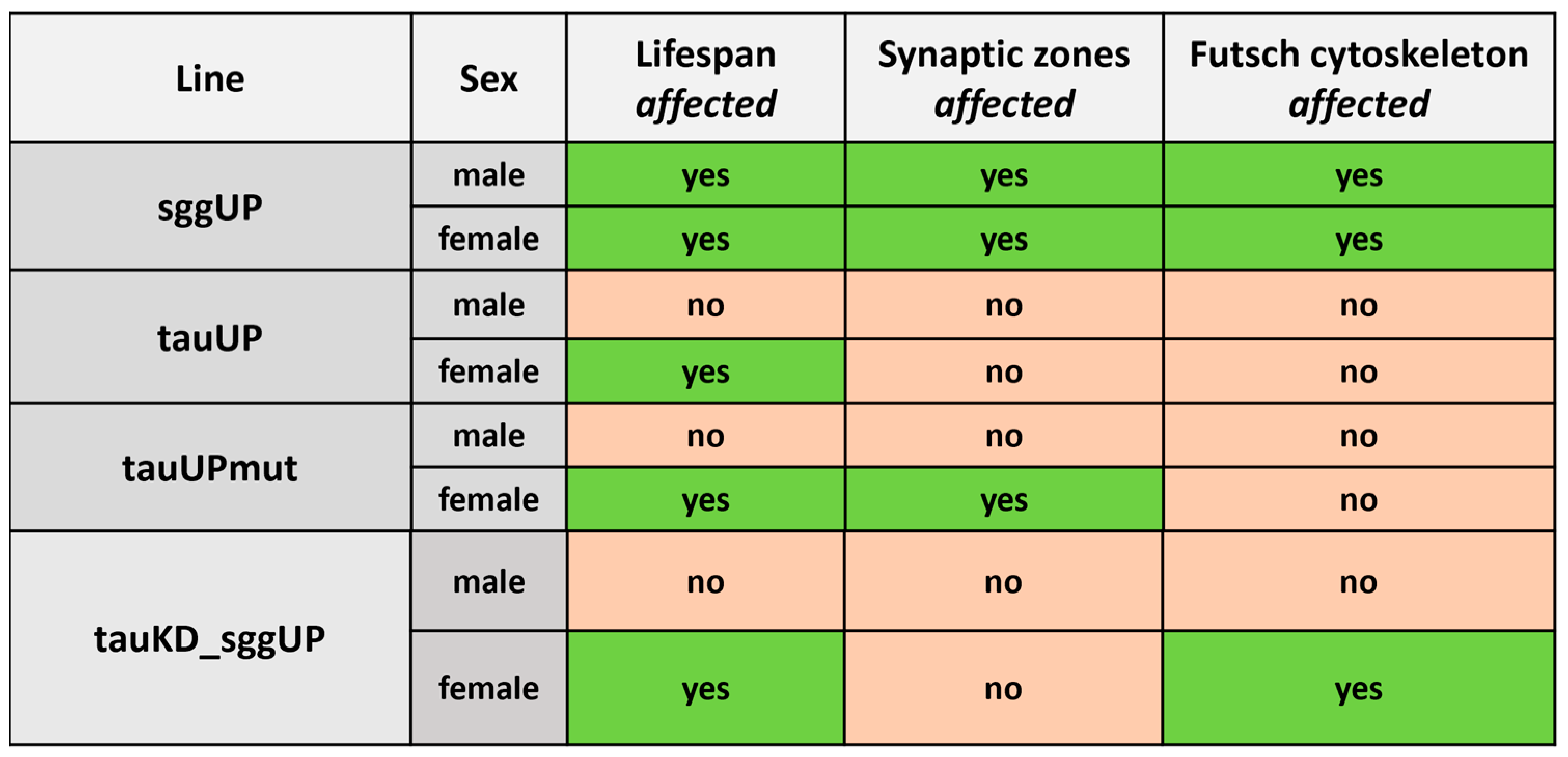
2.1. Tau Affects Drosophila Life Span
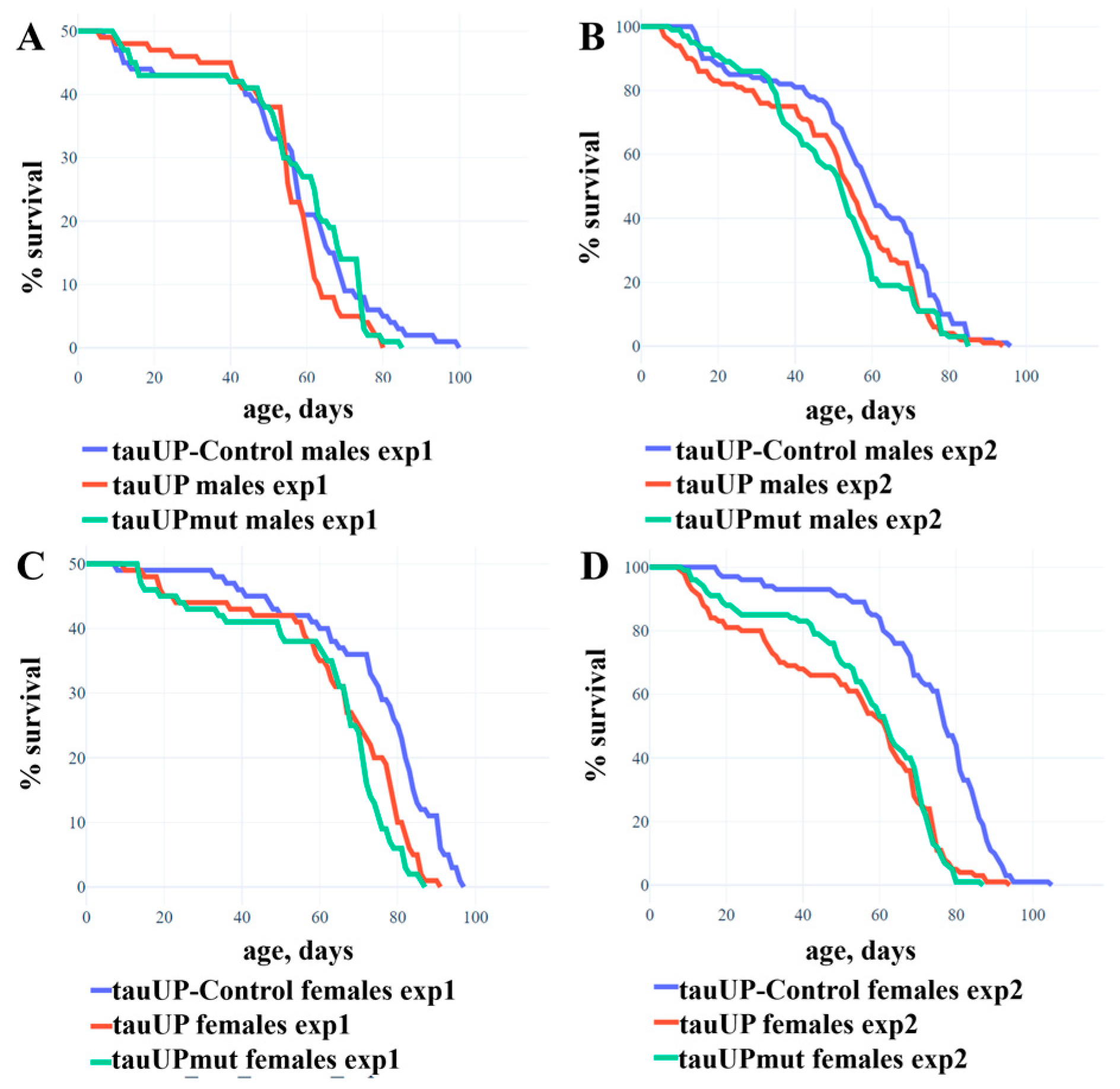
2.1.1. Overexpression of the RA Transcript of the Tau Gene Reduces the Lifespan of Females
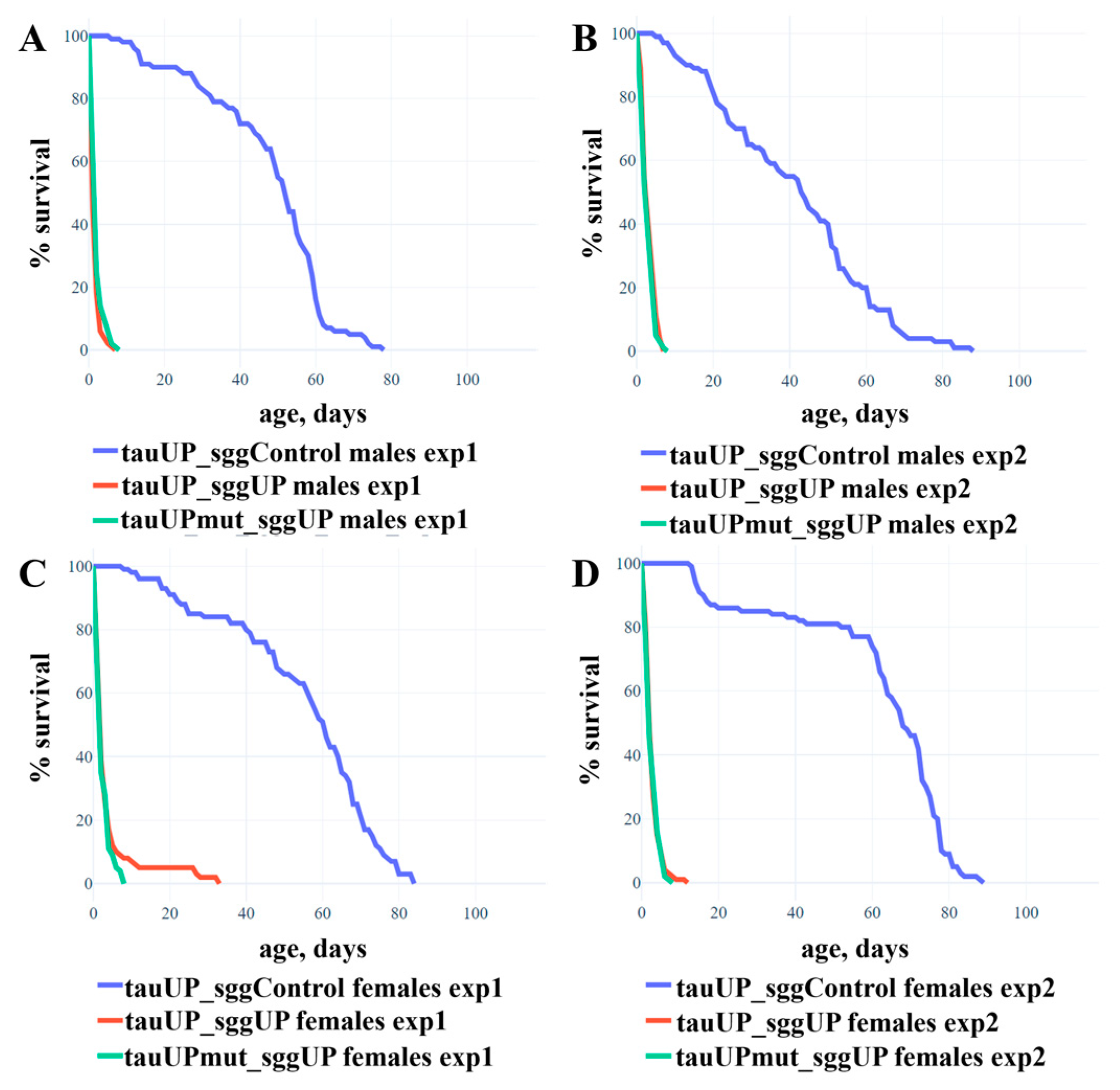
2.1.2. Overexpression of the RA Transcript of the Tau Gene Does Not Affect the Reduction in Lifespan Caused by the Overexpression of the RB Transcript of the Shaggy Gene
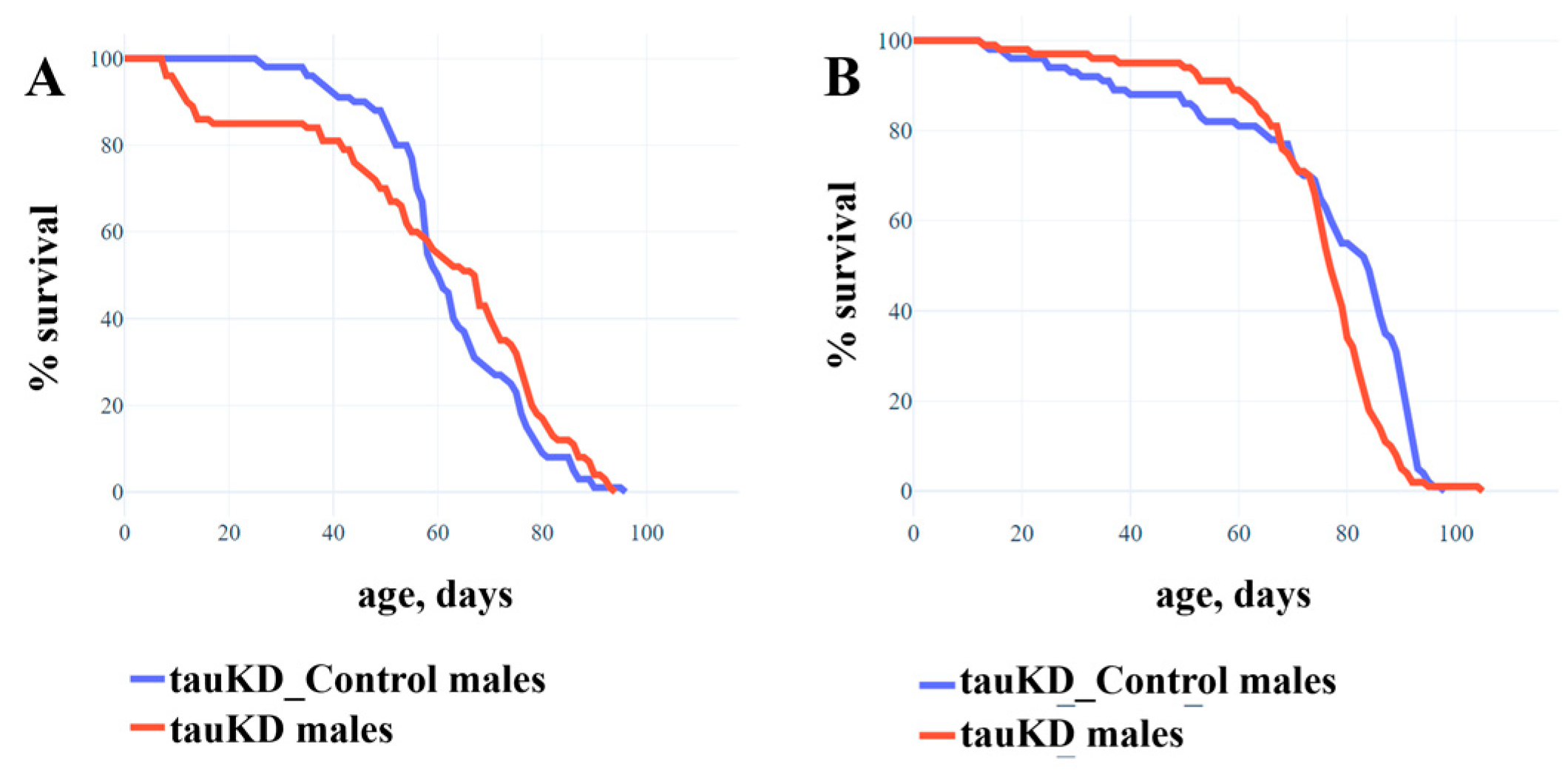
2.1.3. Knockdown of the Tau Gene Has No Effect on the Lifespan of Males and Females
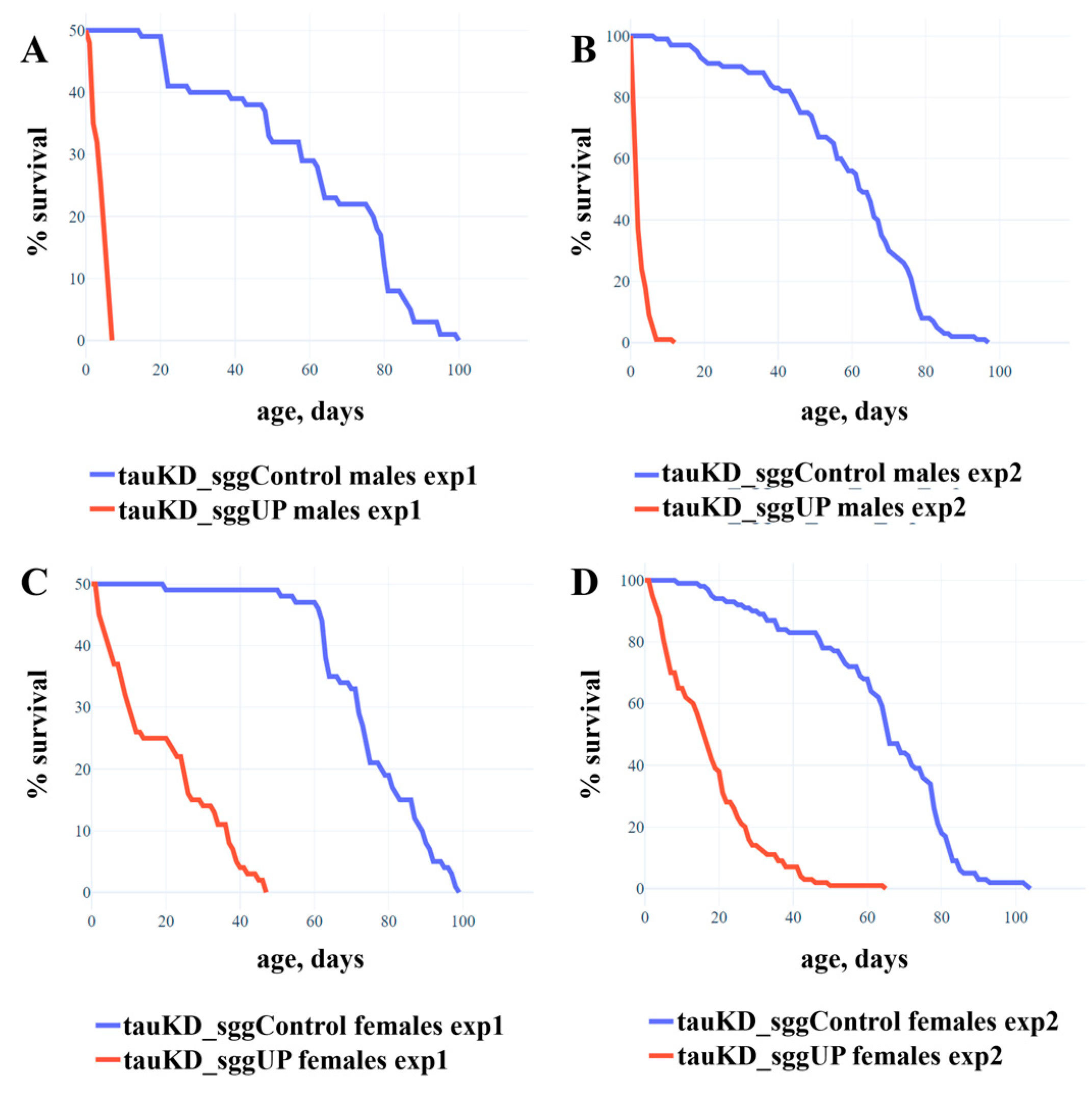
2.1.4. Knockdown of the Tau Gene Positively Affects the Lifespan of Females with the Overexpression of the RB Transcript of the shaggy Gene
2.2. Tau Affects the Properties of the Nervous System
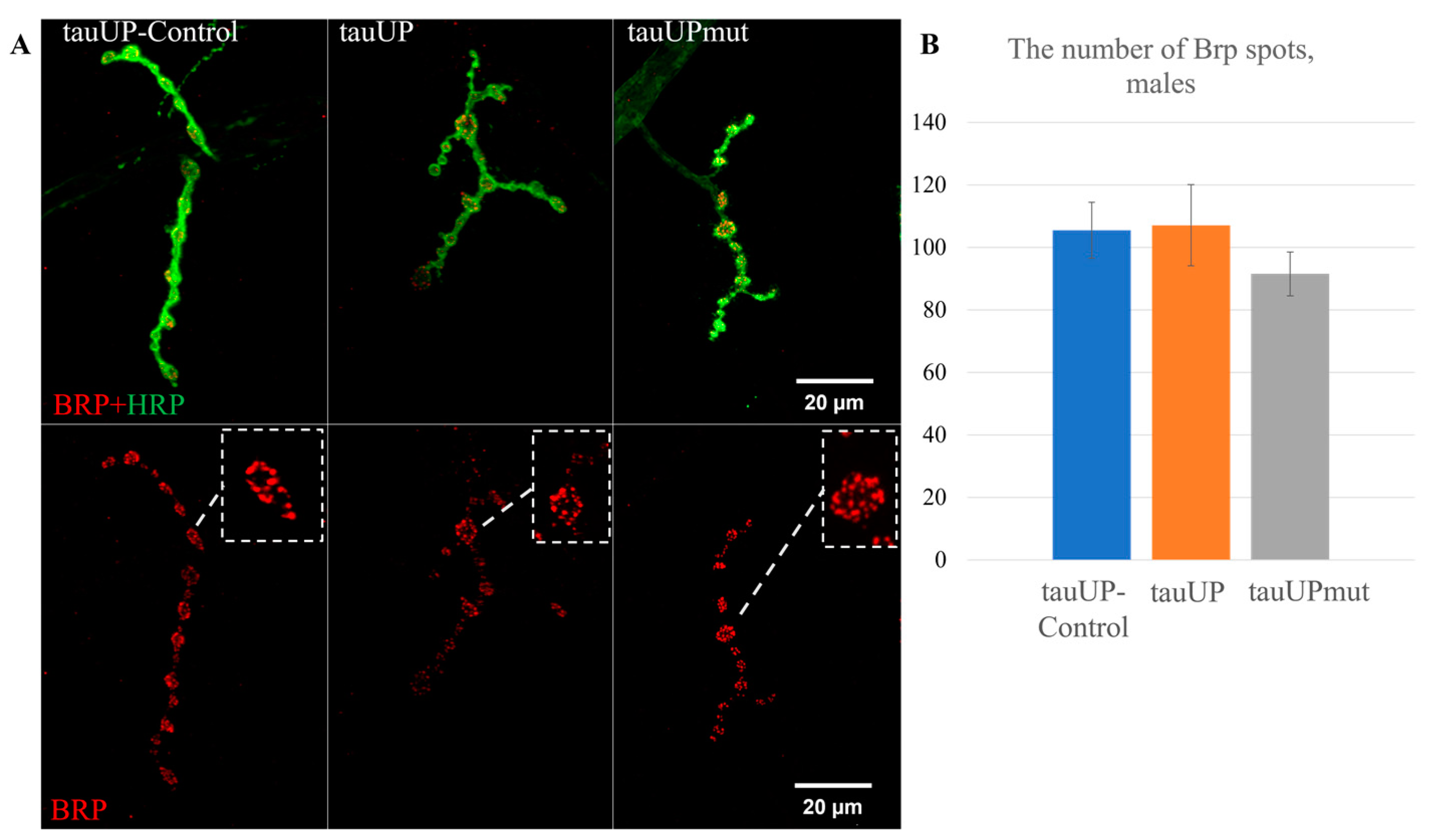
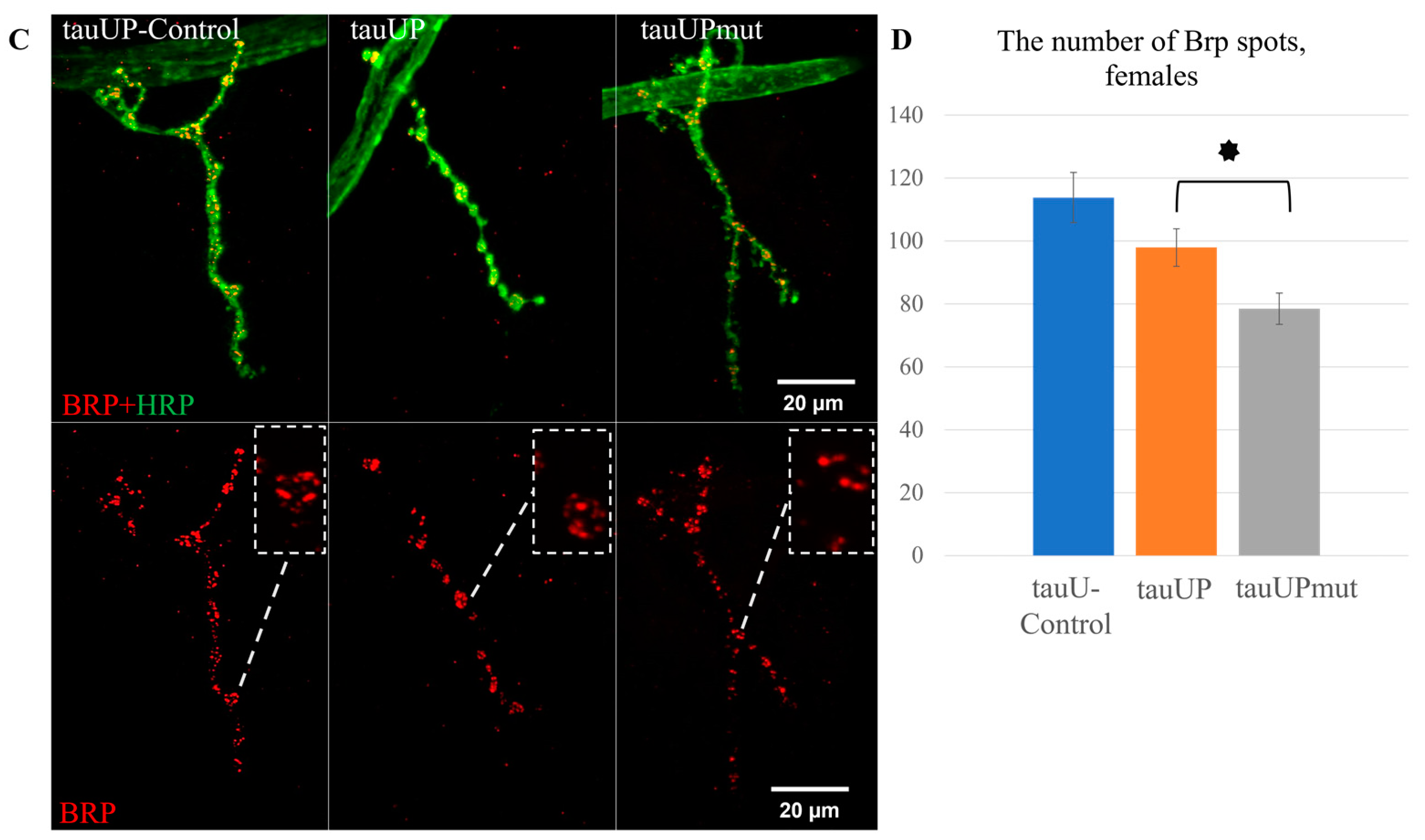
2.2.1. Overexpression of the RA Transcript of the Tau Gene Mutant for GSK3 Phosphorylation Sites Affects Synaptic Activity in Females
2.2.2. Overexpression of the RA Transcript of the Tau Gene Does Not Affect the Distribution of the Microtubule-Associated Futsch Protein
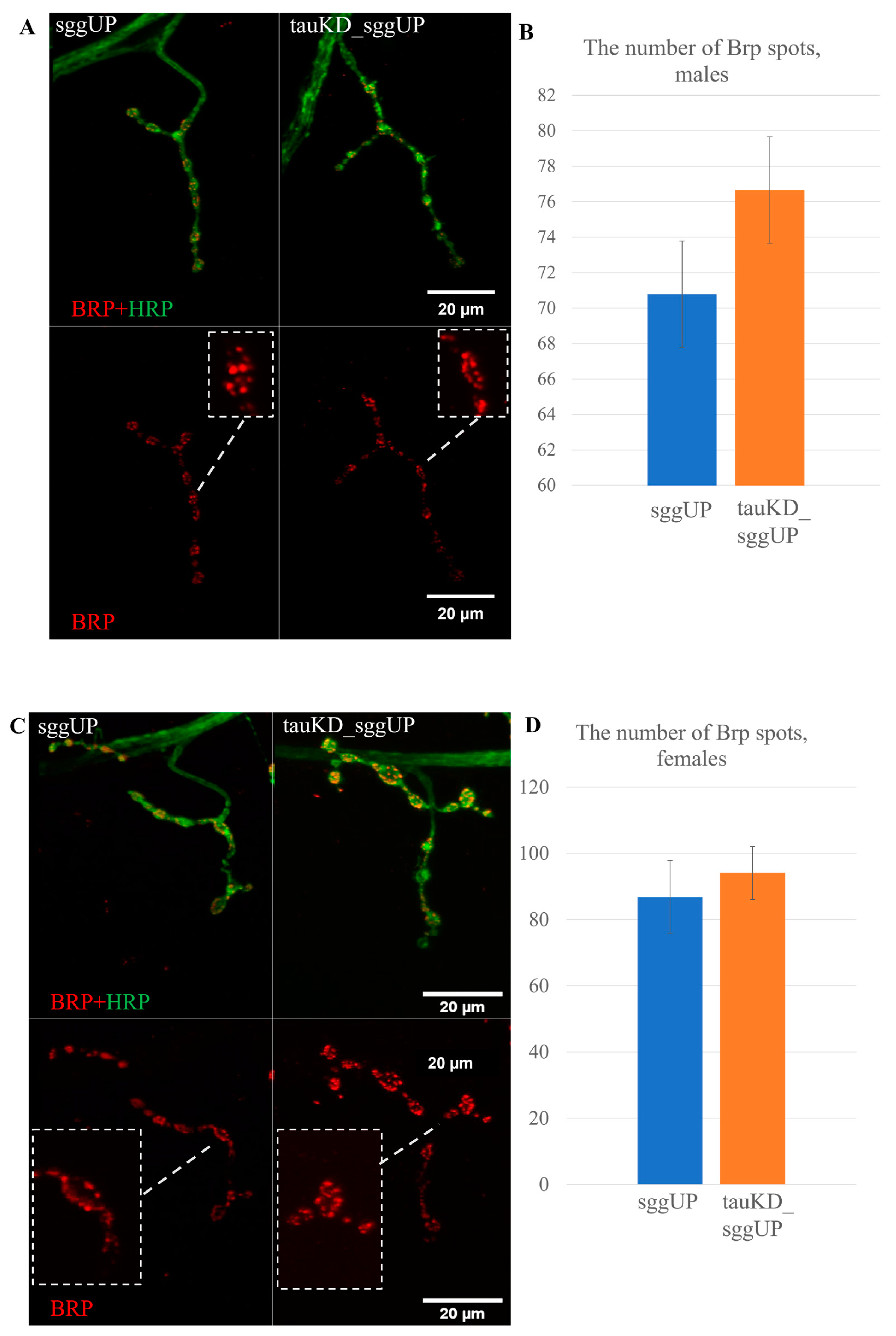
2.2.3. Knockdown of the Tau Gene Does Not Affect Synaptic Activity Reduced by the Overexpression of the RB Transcript of the Shaggy Gene
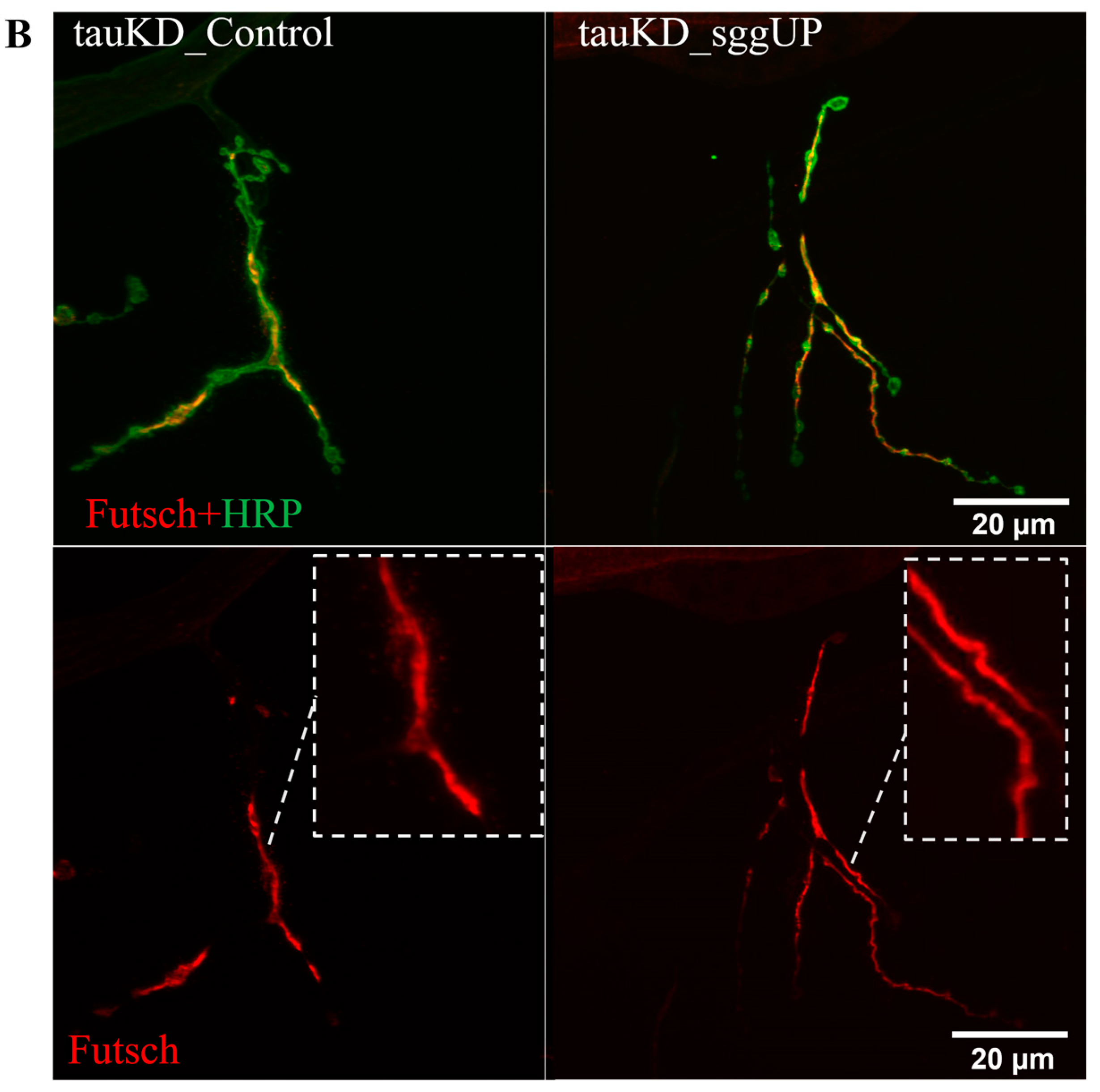
2.2.4. Knockdown of the Tau Gene Positively Affects the Distribution of the Microtubule-Associated Futsch Protein Impaired in Females with the Overexpression of the RB Transcript of the Shaggy Gene
3. Materials and Methods
3.1. Lines and Crossings
3.2. Tests for Wolbachia
3.3. Lifespan Assay
3.4. Immunostaining and Microscopy
3.5. Phosphorylation Sites and Mutagenesis
3.6. Statistical Analyses
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barbier, P.; Zejneli, O.; Martinho, M.; Lasorsa, A.; Belle, V.; Smet-Nocca, C.; Tsvetkov, P.O.; Devred, F.; Landrieu, I. Role of Tau as a Microtubule-Associated Protein: Structural and Functional Aspects. Front. Aging Neurosci. 2019, 11, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadavath, H.; Hofele, R.V.; Biernat, J.; Kumar, S.; Tepper, K.; Urlaub, H.; Mandelkow, E.; Zweckstetter, M. Tau Stabilizes Microtubules by Binding at the Interface between Tubulin Heterodimers. Proc. Natl. Acad. Sci. USA 2015, 112, 7501–7506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colonna, M.; Wang, Y. TREM2 Variants: New Keys to Decipher Alzheimer Disease Pathogenesis. Nat. Rev. Neurosci. 2016, 17, 201–207. [Google Scholar] [CrossRef]
- Kent, S.A.; Spires-Jones, T.L.; Durrant, C.S. The Physiological Roles of Tau and Aβ: Implications for Alzheimer’s Disease Pathology and Therapeutics. Acta Neuropathol. 2020, 140, 417–447. [Google Scholar] [CrossRef]
- Peng, C.; Trojanowski, J.Q.; Lee, V.M.-Y. Protein Transmission in Neurodegenerative Disease. Nat. Rev. Neurol. 2020, 16, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Holper, S.; Watson, R.; Yassi, N. Tau as a Biomarker of Neurodegeneration. Int. J. Mol. Sci. 2022, 23, 7307. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, K.-M.; Yang, L.; Dong, Q.; Yu, J.-T. Tauopathies: New Perspectives and Challenges. Mol. Neurodegener. 2022, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mandelkow, E. Tau in Physiology and Pathology. Nat. Rev. Neurosci. 2015, 17, 22–35. [Google Scholar] [CrossRef]
- Morris, M.; Knudsen, G.M.; Maeda, S.; Trinidad, J.C.; Ioanoviciu, A.; Burlingame, A.L.; Mucke, L. Tau Post-Translational Modifications in Wild-Type and Human Amyloid Precursor Protein Transgenic Mice. Nat. Neurosci. 2015, 18, 1183–1189. [Google Scholar] [CrossRef]
- Martin, L.; Latypova, X.; Terro, F. Post-Translational Modifications of Tau Protein: Implications for Alzheimer’s Disease. Neurochem. Int. 2011, 58, 458–471. [Google Scholar] [CrossRef]
- Hanger, D.P.; Byers, H.L.; Wray, S.; Leung, K.-Y.; Saxton, M.J.; Seereeram, A.; Reynolds, C.H.; Ward, M.A.; Anderton, B.H. Novel Phosphorylation Sites in Tau from Alzheimer Brain Support a Role for Casein Kinase 1 in Disease Pathogenesis. J. Biol. Chem. 2007, 282, 23645–23654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanger, D.P.; Anderton, B.H.; Noble, W. Tau Phosphorylation: The Therapeutic Challenge for Neurodegenerative Disease. Trends Mol. Med. 2009, 15, 112–119. [Google Scholar] [CrossRef]
- Kanaan, N.M.; Morfini, G.; Pigino, G.; LaPointe, N.E.; Andreadis, A.; Song, Y.; Leitman, E.; Binder, L.I.; Brady, S.T. Phosphorylation in the Amino Terminus of Tau Prevents Inhibition of Anterograde Axonal Transport. Neurobiol. Aging 2012, 33, 826.e15–826.e30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, D.; Mukrasch, M.D.; Biernat, J.; Bibow, S.; Blackledge, M.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. Conformational Changes Specific for Pseudophosphorylation at Serine 262 Selectively Impair Binding of Tau to Microtubules. Biochemistry 2009, 48, 10047–10055. [Google Scholar] [CrossRef] [PubMed]
- Kiris, E.; Ventimiglia, D.; Sargin, M.E.; Gaylord, M.R.; Altinok, A.; Rose, K.; Manjunath, B.S.; Jordan, M.A.; Wilson, L.; Feinstein, S.C. Combinatorial Tau Pseudophosphorylation: Markedly Different Regulatory Effects on Microtubule Assembly and Dynamic Instability than the Sum of the Individual Parts. J. Biol. Chem. 2011, 286, 14257–14270. [Google Scholar] [CrossRef] [Green Version]
- Biundo, F.; Del Prete, D.; Zhang, H.; Arancio, O.; D’Adamio, L. A Role for Tau in Learning, Memory and Synaptic Plasticity. Sci. Rep. 2018, 8, 3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sydow, A.; Van der Jeugd, A.; Zheng, F.; Ahmed, T.; Balschun, D.; Petrova, O.; Drexler, D.; Zhou, L.; Rune, G.; Mandelkow, E.-M.; et al. Tau-Induced Defects in Synaptic Plasticity, Learning, and Memory Are Reversible in Transgenic Mice after Switching Off the Toxic Tau Mutant. J. Neurosci. 2011, 31, 2511–2525. [Google Scholar] [CrossRef] [Green Version]
- Kosmidis, S.; Grammenoudi, S.; Papanikolopoulou, K.; Skoulakis, E.M.C. Differential Effects of Tau on the Integrity and Function of Neurons Essential for Learning in Drosophila. J. Neurosci. 2010, 30, 464–477. [Google Scholar] [CrossRef] [Green Version]
- Papanikolopoulou, K.; Kosmidis, S.; Grammenoudi, S.; Skoulakis, E.M.C. Phosphorylation Differentiates Tau-Dependent Neuronal Toxicity and Dysfunction. Biochem. Soc. Trans. 2010, 38, 981–987. [Google Scholar] [CrossRef] [Green Version]
- Papanikolopoulou, K.; Skoulakis, E.M.C. Temporally Distinct Phosphorylations Differentiate Tau-Dependent Learning Deficits and Premature Mortality in Drosophila. Hum. Mol. Genet. 2015, 24, 2065–2077. [Google Scholar] [CrossRef]
- Povellato, G.; Tuxworth, R.I.; Hanger, D.P.; Tear, G. Modification of the Drosophila Model of In Vivo Tau Toxicity Reveals Protective Phosphorylation by GSK3β. Biol. Open 2014, 3, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rankin, C.A.; Sun, Q.; Gamblin, T.C. Tau Phosphorylation by GSK-3β Promotes Tangle-like Filament Morphology. Mol. Neurodegener. 2007, 2, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avila, J.; León-Espinosa, G.; García, E.; García-Escudero, V.; Hernández, F.; DeFelipe, J. Tau Phosphorylation by GSK3 in Different Conditions. Int. J. Alzheimer’s Dis. 2012, 2012, 578373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, A.C.; Perez-Nievas, B.G.; Siao Tick Chong, M.; Gonzalez-Martinez, A.; Argente-Escrig, H.; Rubio-Guerra, S.; Commins, C.; Muftu, S.; Eftekharzadeh, B.; Hudry, E.; et al. Isoform-Selective Decrease of Glycogen Synthase Kinase-3-Beta (GSK-3β) Reduces synaptic Tau Phosphorylation, Transcellular Spreading, and Aggregation. Iscience 2021, 24, 102058. [Google Scholar] [CrossRef]
- Gómez de Barreda, E.; Pérez, M.; Gómez Ramos, P.; de Cristobal, J.; Martín-Maestro, P.; Morán, A.; Dawson, H.N.; Vitek, M.P.; Lucas, J.J.; Hernández, F.; et al. Tau-Knockout Mice Show Reduced GSK3-Induced Hippocampal Degeneration and Learning Deficits. Neurobiol. Dis. 2010, 37, 622–629. [Google Scholar] [CrossRef]
- Gistelinck, M.; Lambert, J.-C.; Callaerts, P.; Dermaut, B.; Dourlen, P. Drosophila Models of Tauopathies: What Have We Learned? Int. J. Alzheimer’s Dis. 2012, 2012, 970980. [Google Scholar] [CrossRef] [Green Version]
- Jackson, G.R.; Wiedau-Pazos, M.; Sang, T.-K.; Wagle, N.; Brown, C.A.; Massachi, S.; Geschwind, D.H. Human Wild-Type Tau Interacts with Wingless Pathway Components and Produces Neurofibrillary Pathology in Drosophila. Neuron 2002, 34, 509–519. [Google Scholar] [CrossRef]
- Chatterjee, S.; Sang, T.-K.; Lawless, G.M.; Jackson, G.R. Dissociation of Tau Toxicity and Phosphorylation: Role of GSK-3β, MARK and Cdk5 in a Drosophila Model. Hum. Mol. Genet. 2009, 18, 164–177. [Google Scholar] [CrossRef] [Green Version]
- Pereanu, W.; Shy, D.; Hartenstein, V. Morphogenesis and Proliferation of the Larval Brain Glia in Drosophila. Dev. Biol. 2005, 283, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Pandey, U.B.; Nichols, C.D. Human Disease Models in Drosophila Melanogaster and the Role of the Fly in Therapeutic Drug Discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef]
- Heidary, G.; Fortini, M.E. Identification and Characterization of the Drosophila Tau Homolog. Mech. Dev. 2001, 108, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Y.; Huang, J.; Cao, D.; Yang, G.; Liu, W.; Lu, H.; Guo, A. Study of Tauopathies by Comparing Drosophila and Human Tau in Drosophila. Cell Tissue Res. 2007, 329, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Law, A.D.; Cassar, M.; Long, D.M.; Chow, E.S.; Giebultowicz, J.M.; Venkataramanan, A.; Strauss, R.; Kretzschmar, D. FTD-Associated Mutations in Tau Result in a Combination of Dominant and Recessive Phenotypes. Neurobiol. Dis. 2022, 170, 105770. [Google Scholar] [CrossRef]
- Sealey, M.A.; Vourkou, E.; Cowan, C.M.; Bossing, T.; Quraishe, S.; Grammenoudi, S.; Skoulakis, E.M.C.; Mudher, A. Distinct Phenotypes of Three-Repeat and Four-Repeat Human Tau in a Transgenic Model of Tauopathy. Neurobiol. Dis. 2017, 105, 74–83. [Google Scholar] [CrossRef] [Green Version]
- Azam, S.; Haque, M.E.; Balakrishnan, R.; Kim, I.-S.; Choi, D.-K. The Ageing Brain: Molecular and Cellular Basis of Neurodegeneration. Front. Cell Dev. Biol. 2021, 9, 683459. [Google Scholar] [CrossRef]
- Therriault, J.; Pascoal, T.A.; Benedet, A.L.; Tissot, C.; Savard, M.; Chamoun, M.; Lussier, F.; Kang, M.S.; Berzgin, G.; Wang, T.; et al. Frequency of Biologically-Defined Alzheimer Disease in Relation to Age, Sex, APOEE4 and Cognitive Impairment. Neurology 2021, 96, e975–e985. [Google Scholar] [CrossRef]
- Pinares-Garcia, P.; Stratikopoulos, M.; Zagato, A.; Loke, H.; Lee, J. Sex: A Significant Risk Factor for Neurodevelopmental and Neurodegenerative Disorders. Brain Sci. 2018, 8, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; McNally, S.; Kilpatrick, J.I.; Jarvis, S.P.; O’Brien, C.J. Aging and Ocular Tissue Stiffness in Glaucoma. Surv. Ophthalmol. 2018, 63, 56–74. [Google Scholar] [CrossRef]
- Marques-Aleixo, I.; Beleza, J.; Sampaio, A.; Stevanović, J.; Coxito, P.; Gonçalves, I.; Ascensão, A.; Magalhães, J. Preventive and Therapeutic Potential of Physical Exercise in Neurodegenerative Diseases. Antioxidants Redox Signal. 2021, 34, 674–693. [Google Scholar] [CrossRef]
- Chowen, J.A.; Garcia-Segura, L.M. Role of Glial Cells in the Generation of Sex Differences in Neurodegenerative Diseases and Brain Aging. Mech. Ageing Dev. 2021, 196, 111473. [Google Scholar] [CrossRef] [PubMed]
- Lentini, E.; Kasahara, M.; Arver, S.; Savic, I. Sex Differences in the Human Brain and the Impact of Sex Chromosomes and Sex Hormones. Cereb. Cortex 2013, 23, 2322–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokouhi, S.; Taylor, W.D.; Albert, K.; Kang, H.; Newhouse, P.A.; Initiative, T.A.D.N. In Vivo Network Models Identify Sex Differences in the Spread of Tau Pathology across the Brain. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2020, 12, e12016. [Google Scholar] [CrossRef] [PubMed]
- Buckley, R.F.; Mormino, E.C.; Rabin, J.S.; Hohman, T.J.; Landau, S.; Hanseeuw, B.J.; Jacobs, H.I.L.; Papp, K.V.; Amariglio, R.E.; Properzi, M.J.; et al. Sex Differences in the Association of Global Amyloid and Regional Tau Deposition Measured by Positron Emission Tomography in Clinically Normal Older Adults. JAMA Neurol. 2019, 76, 542–551. [Google Scholar] [CrossRef] [Green Version]
- Shah, E.J.; Gurdziel, K.; Ruden, D.M. Sex-Differences in Traumatic Brain Injury in the Absence of Tau in Drosophila. Genes 2021, 12, 917. [Google Scholar] [CrossRef]
- Burnouf, S.; Grönke, S.; Augustin, H.; Dols, J.; Gorsky, M.K.; Werner, J.; Kerr, F.; Alic, N.; Martinez, P.; Partridge, L. Deletion of Endogenous Tau Proteins is not Detrimental in Drosophila. Sci. Rep. 2016, 6, 23102. [Google Scholar] [CrossRef] [Green Version]
- Glodzik, L.; de Santi, S.; Tsui, W.H.; Mosconi, L.; Zinkowski, R.; Pirraglia, E.; Wang, H.Y.; Li, Y.; Rich, K.E.; Zetterberg, H.; et al. Phosphorylated Tau 231, Memory Decline and Medial Temporal Atrophy in Normal Elders. Neurobiol. Aging 2011, 32, 2131–2141. [Google Scholar] [CrossRef] [Green Version]
- Alonso, A.D.; Cohen, L.S. Our Tau Tales from Normal to Pathological Behavior. J. Alzheimer’s Dis. 2018, 64, S507–S516. [Google Scholar] [CrossRef]
- Li, T.; Paudel, H.K. Glycogen Synthase Kinase 3β Phosphorylates Alzheimer’s Disease-Specific Ser396 of Microtubule-Associated Protein Tau by a Sequential Mechanism. Biochemistry 2006, 45, 3125–3133. [Google Scholar] [CrossRef]
- Vourkou, E.; Paspaliaris, V.; Bourouliti, A.; Zerva, M.-C.; Prifti, E.; Papanikolopoulou, K.; Skoulakis, E.M.C. Differential Effects of Human Tau Isoforms to Neuronal Dysfunction and Toxicity in the Drosophila CNS. Int. J. Mol. Sci. 2022, 23, 12985. [Google Scholar] [CrossRef]
- Krementsova, A.V.; Roshina, N.V.; Tsybul’Ko, E.A.; Rybina, O.Y.; Symonenko, A.V.; Pasyukova, E.G. Reproducible Effects of the Mitochondria-Targeted Plastoquinone Derivative SkQ1 on Drosophila melanogaster Lifespan under Different Experimental Scenarios. Biogerontology 2012, 13, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.C.; Banks, S.J.; Thompson, W.K.; Chen, C.-H.; McEvoy, L.K.; Tan, C.H.; Kukull, W.; Bennett, D.A.; Farrer, L.A.; Mayeux, R.; et al. Sex-Dependent Autosomal Effects on Clinical Progression of Alzheimer’s disease. Brain 2020, 143, 2272–2280. [Google Scholar] [CrossRef]
- Oveisgharan, S.; Arvanitakis, Z.; Yu, L.; Farfel, J.; Schneider, J.A.; Bennett, D.A. Sex Differences in Alzheimer’s Disease and Common Neuropathologies of Aging. Acta Neuropathol. 2018, 136, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Trostnikov, M.V.; Roshina, N.V.; Boldyrev, S.V.; Veselkina, E.R.; Zhuikov, A.A.; Krementsova, A.V.; Pasyukova, E.G. Disordered Expression of Shaggy, the Drosophila Gene Encoding a Serine-Threonine Protein Kinase GSK3, Affects the Lifespan in a Transcript-, Stage-, and Tissue-Specific Manner. Int. J. Mol. Sci. 2019, 20, 2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Symonenko, A.V.; Roshina, N.V.; Krementsova, A.V.; Pasyukova, E.G. Reduced Neuronal Transcription of Escargot, the Drosophila Gene Encoding a Snail-Type Transcription Factor, Promotes Longevity. Front. Genet. 2018, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Rybina, O.Y.; Schelkunov, M.I.; Veselkina, E.R.; Sarantseva, S.V.; Krementsova, A.V.; Vysokikh, M.Y.; Melentev, P.A.; Volodina, M.A.; Pasyukova, E.G. Knockdown of the Neuronal Gene Lim3 at the Early Stages of Development Affects Mitochondrial Function and Lifespan in Drosophila. Mech. Ageing Dev. 2019, 181, 29–41. [Google Scholar] [CrossRef] [PubMed]
- Patel, P.; Woodgett, J.R. Glycogen Synthase Kinase 3: A Kinase for All Pathways? Curr. Top. Dev. Biol. 2017, 123, 277–302. [Google Scholar] [CrossRef]
- Noble, W.; Hanger, D.P.; Miller, C.C.J.; Lovestone, S. The Importance of Tau Phosphorylation for Neurodegenerative Diseases. Front. Neurol. 2013, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.-L.; Wang, N.; Sun, F.-R.; Cao, X.-P.; Zhang, W.; Yu, J.-T. Tau in Neurodegenerative Disease. Ann. Transl. Med. 2018, 6, 175. [Google Scholar] [CrossRef]
- Wagh, D.A.; Rasse, T.M.; Asan, E.; Hofbauer, A.; Schwenkert, I.; Dürrbeck, H.; Buchner, S.; Dabauvalle, M.-C.; Schmidt, M.; Qin, G.; et al. Bruchpilot, a Protein with Homology to ELKS/CAST, Is Required for Structural Integrity and Function of Synaptic Active Zones in Drosophila. Neuron 2006, 49, 833–844. [Google Scholar] [CrossRef]
- Kittel, R.J.; Wichmann, C.; Rasse, T.M.; Fouquet, W.; Schmidt, M.; Schmid, A.; Wagh, D.A.; Pawlu, C.; Kellner, R.R.; Willig, K.I.; et al. Bruchpilot Promotes Active Zone Assembly, Ca2+ Channel Clustering, and Vesicle Release. Science 2006, 312, 1051–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chee, F.C.; Mudher, A.; Cuttle, M.F.; Newman, T.A.; MacKay, D.; Lovestone, S.; Shepherd, D. Over-Expression of Tau Results in Defective Synaptic Transmission in Drosophila Neuromuscular Junctions. Neurobiol. Dis. 2005, 20, 918–928. [Google Scholar] [CrossRef] [PubMed]
- Mondragón-Rodríguez, S.; Perry, G.; Luna-Muñoz, J.; Acevedo-Aquino, M.C.; Williams, S. Phosphorylation of Tau Protein at Sites Ser(396-404) is One of the Earliest Events in Alzheimer’s Disease and Down Syndrome. Neuropathol. Appl. Neurobiol. 2014, 40, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Maurage, C.-A.; Sergeant, N.; Ruchoux, M.-M.; Hauw, J.-J.; Delacourte, A. Phosphorylated Serine 199 of Microtubule-Associated Protein Tau is a Neuronal Epitope Abundantly Expressed in Youth and an Early Marker of Tau Pathology. Acta Neuropathol. 2003, 105, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Z.; Grundke-Iqbal, I.; Iqbal, K. Kinases and Phosphatases and Tau Sites Involved in Alzheimer Neurofibrillary Degeneration. Eur. J. Neurosci. 2007, 25, 59–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alonso, A.D.; Cohen, L.S.; Corbo, C.; Morozova, V.; ElIdrissi, A.; Phillips, G.; Kleiman, F.E. Hyperphosphorylation of Tau Associates With Changes in Its Function Beyond Microtubule Stability. Front. Cell. Neurosci. 2018, 12, 338. [Google Scholar] [CrossRef] [Green Version]
- Sferra, A.; Nicita, F.; Bertini, E. Microtubule Dysfunction: A Common Feature of Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 7354. [Google Scholar] [CrossRef]
- Dubey, J.; Ratnakaran, N.; Koushika, S.P. Neurodegeneration and Microtubule Dynamics: Death by a Thousand Cuts. Front. Cell. Neurosci. 2015, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Gögel, S.; Wakefield, S.; Tear, G.; Klämbt, C.; Gordon-Weeks, P.R. The Drosophila Microtubule Associated Protein Futsch is Phosphorylated by Shaggy/Zeste-White 3 at an Homologous GSK3β Phosphorylation Site in MAP1B. Mol. Cell. Neurosci. 2006, 33, 188–199. [Google Scholar] [CrossRef]
- da Cruz, A.B.; Schwärzel, M.; Schulze, S.; Niyyati, M.; Heisenberg, M.; Kretzschmar, D. Disruption of the MAP1B-related Protein FUTSCH Leads to Changes in the Neuronal Cytoskeleton, Axonal Transport Defects, and Progressive Neurodegeneration in Drosophila. Mol. Biol. Cell 2005, 16, 2433–2442. [Google Scholar] [CrossRef]
- Xiong, Y.; Zhao, K.; Wu, J.; Xu, Z.; Jin, S.; Zhang, Y.Q. HDAC6 Mutations Rescue Human Tau-Induced Microtubule Defects in Drosophila. Proc. Natl. Acad. Sci. USA 2013, 110, 4604–4609. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; An, P.; Li, H.; Zhou, Z.; Sun, Y.; Wang, J.; Ma, L.; Lu, B. Tau Accumulation via Reduced Autophagy Mediates GGGGCC Repeat Expansion-Induced Neurodegeneration in Drosophila Model of ALS. Neurosci. Bull. 2020, 36, 1414–1428. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-Q.; Wang, S.-H.; Liu, D.; Yin, Y.-Y.; Tian, Q.; Wang, X.-C.; Wang, Q.; Chen, J.-G.; Wang, J.-Z. Activation of Glycogen Synthase Kinase-3 Inhibits Long-Term Potentiation with Synapse-Associated Impairments. J. Neurosci. 2007, 27, 12211–12220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourouis, M. Targeted Increase in Shaggy Activity Levels Blocks Wingless Signaling. Genes. 2002, 34, 99–102. [Google Scholar] [CrossRef]
- Mockett, R.J.; Cockrell, J.C.; Puri, S.; Nguyen, M.; Nisa, M. Long-Lived Genotypes for Studies of Life Extension in Drosophila Melanogaster. Mech. Ageing Dev. 2012, 133, 359–367. [Google Scholar] [CrossRef] [PubMed]
- McGraw, E.A.; O’Neill, S.L. Wolbachia Pipientis: Intracellular Infection and Pathogenesis in Drosophila. Curr. Opin. Microbiol. 2004, 7, 67–70. [Google Scholar] [CrossRef]
- Holden, P.; Jones, P.; Brookfield, J. Evidence for a Wolbachia Symbiont in Drosophila melanogaster. Genet. Res. 1993, 62, 23–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshina, N.V.; Symonenko, A.V.; Krementsova, A.V.; Trostnikov, M.V.; Pasyukova, E.G. Embryonic Expression of Shuttle Craft, a Drosophila Gene Involved in Neuron Development, is Associated with Adult Lifespan. Aging 2014, 6, 1076–1093. [Google Scholar] [CrossRef] [Green Version]
- Roos, J.; Hummel, T.; Ng, N.; Klämbt, C.; Davis, G.W. Drosophila Futsch Regulates Synaptic Microtubule Organization and Is Necessary for Synaptic Growth. Neuron 2000, 26, 371–382. [Google Scholar] [CrossRef] [Green Version]
- Franco, B.; Bogdanik, L.; Bobinnec, Y.; Debec, A.; Bockaert, J.; Parmentier, M.-L.; Grau, Y. Shaggy, the Homolog of Glycogen Synthase Kinase 3, Controls Neuromuscular Junction Growth in Drosophila. J. Neurosci. 2004, 24, 6573–6577. [Google Scholar] [CrossRef] [Green Version]
- Bouleau, S.; Tricoire, H. Drosophila Models of Alzheimer’s Disease: Advances, Limits, and Perspectives. J. Alzheimer’s Dis. 2015, 45, 1015–1038. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Grundke-Iqbal, I.; Zaidi, T.; Merz, P.A.; Wen, G.Y.; Shaikh, S.S.; Wisniewski, H.M.; Alafuzoff, I.; Winblad, B. Defective Brain Microtubule Assembly in Alzheimer’s Disease. Lancet 1986, 328, 421–426. [Google Scholar] [CrossRef] [PubMed]
- Wilmoth, J.R.; Horiuchi, S. Rectangularization Revisited: Variability of Age at Death within Human Populations. Demography 1999, 36, 475–495. [Google Scholar] [CrossRef] [PubMed]
- Carey, J.R. Longevity: The Biology and Demography of Life Span; Princeton University Press: Princeton, NT, USA, 2003. [Google Scholar]
- Tan, Q.; Liang, N.; Zhang, X.; Li, J. Dynamic Aging: Channeled Through Microenvironment. Front. Physiol. 2021, 12, 702276. [Google Scholar] [CrossRef] [PubMed]
- Castelli, V.; Benedetti, E.; Antonosante, A.; Catanesi, M.; Pitari, G.; Ippoliti, R.; Cimini, A.; D’Angelo, M. Neuronal Cells Rearrangement During Aging and Neurodegenerative Disease: Metabolism, Oxidative Stress and Organelles Dynamic. Front. Mol. Neurosci. 2019, 12, 132. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Magnus, T. Ageing and Neuronal Vulnerability. Nat. Rev. Neurosci. 2006, 7, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Noble, W.; Hanger, D.P. Roles of Tau Protein in Health and Disease. Acta Neuropathol. 2017, 133, 665–704. [Google Scholar] [CrossRef] [Green Version]
- Violet, M.; Delattre, L.; Tardivel, M.; Sultan, A.; Chauderlier, A.; Caillierez, R.; Talahari, S.; Nesslany, F.; Lefebvre, B.; Bonnefoy, E.; et al. A Major Role for Tau in Neuronal DNA and RNA Protection In Vivo under Physiological and Hyperthermic Conditions. Front. Cell. Neurosci. 2014, 8, 84. [Google Scholar] [CrossRef] [Green Version]
- Cowan, C.M.; Mudher, A. Are Tau Aggregates Toxic or Protective in Tauopathies? Front. Neurol. 2013, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Maffioli, E.; Murtas, G.; Rabattoni, V.; Badone, B.; Tripodi, F.; Iannuzzi, F.; Licastro, D.; Nonnis, S.; Rinaldi, A.M.; Motta, Z.; et al. Insulin and Serine Metabolism as Sex-Specific Hallmarks of Alzheimer’s Disease in the Human Hippocampus. Cell Rep. 2022, 40, 111271. [Google Scholar] [CrossRef]
- Krishnankutty, A.; Kimura, T.; Saito, T.; Aoyagi, K.; Asada, A.; Takahashi, S.-I.; Ando, K.; Ohara-Imaizumi, M.; Ishiguro, K.; Hisanaga, S. In Vivo Regulation of Glycogen Synthase Kinase 3β Activity in Neurons and Brains. Sci. Rep. 2017, 7, 8602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trostnikov, M.V.; Veselkina, E.R.; Krementsova, A.V.; Boldyrev, S.V.; Roshina, N.V.; Pasyukova, E.G. Modulated Expression of the Protein Kinase GSK3 in Motor and Dopaminergic Neurons Increases Female Lifespan in Drosophila melanogaster. Front. Genet. 2020, 11, 668. [Google Scholar] [CrossRef] [PubMed]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veselkina, E.R.; Trostnikov, M.V.; Roshina, N.V.; Pasyukova, E.G. The Effect of the Tau Protein on D. melanogaster Lifespan Depends on GSK3 Expression and Sex. Int. J. Mol. Sci. 2023, 24, 2166. https://doi.org/10.3390/ijms24032166
Veselkina ER, Trostnikov MV, Roshina NV, Pasyukova EG. The Effect of the Tau Protein on D. melanogaster Lifespan Depends on GSK3 Expression and Sex. International Journal of Molecular Sciences. 2023; 24(3):2166. https://doi.org/10.3390/ijms24032166
Chicago/Turabian StyleVeselkina, Ekaterina R., Mikhail V. Trostnikov, Natalia V. Roshina, and Elena G. Pasyukova. 2023. "The Effect of the Tau Protein on D. melanogaster Lifespan Depends on GSK3 Expression and Sex" International Journal of Molecular Sciences 24, no. 3: 2166. https://doi.org/10.3390/ijms24032166