
Morphological and Molecular Functional Evidence of the Pharyngeal Sac in the Digestive Tract of Silver Pomfret, Pampus argenteus
 , ,
, ,
Abstract
:1. Introduction
2. Results
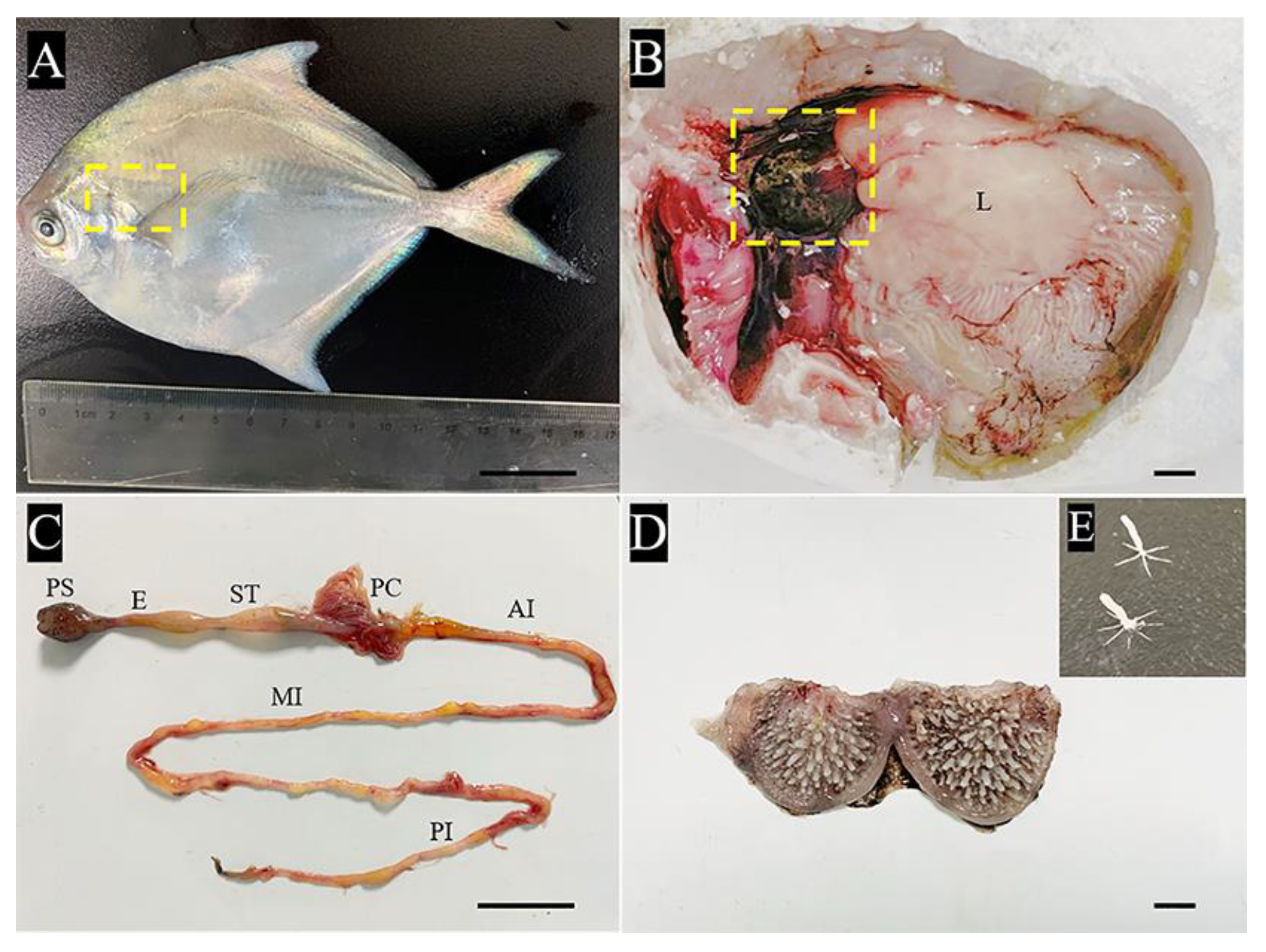
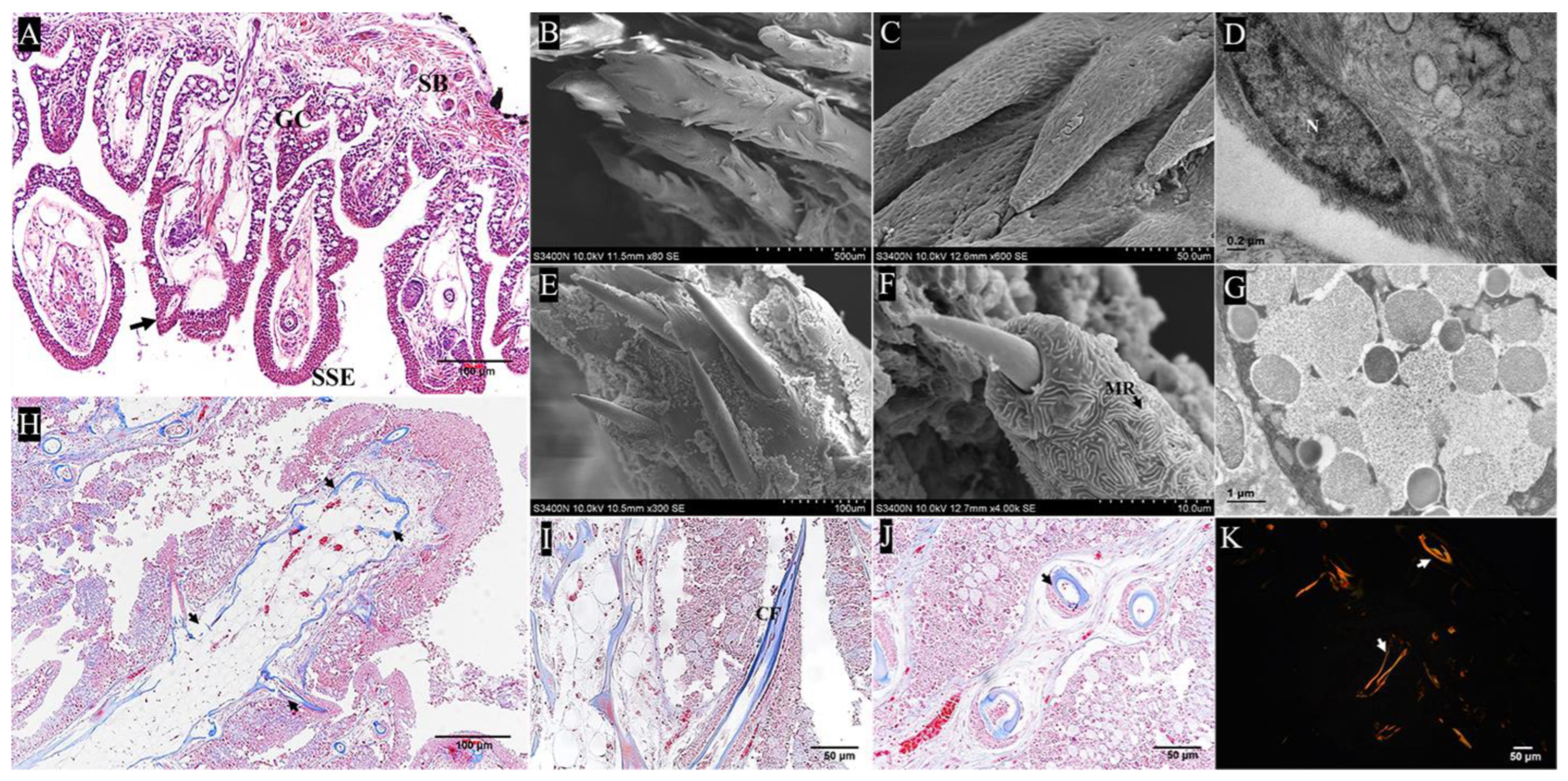
2.1. Gross Anatomy of the Pharyngeal Sac
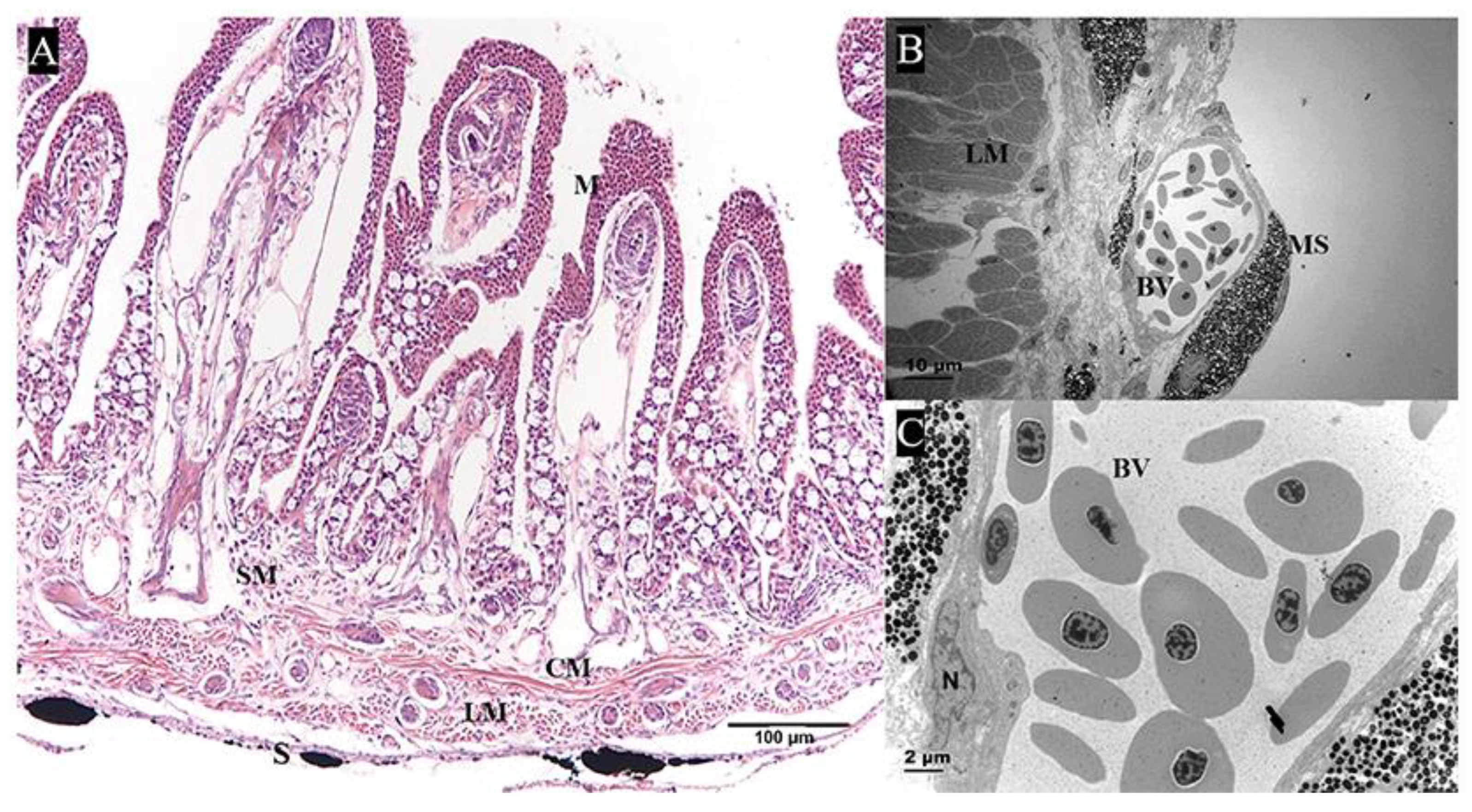
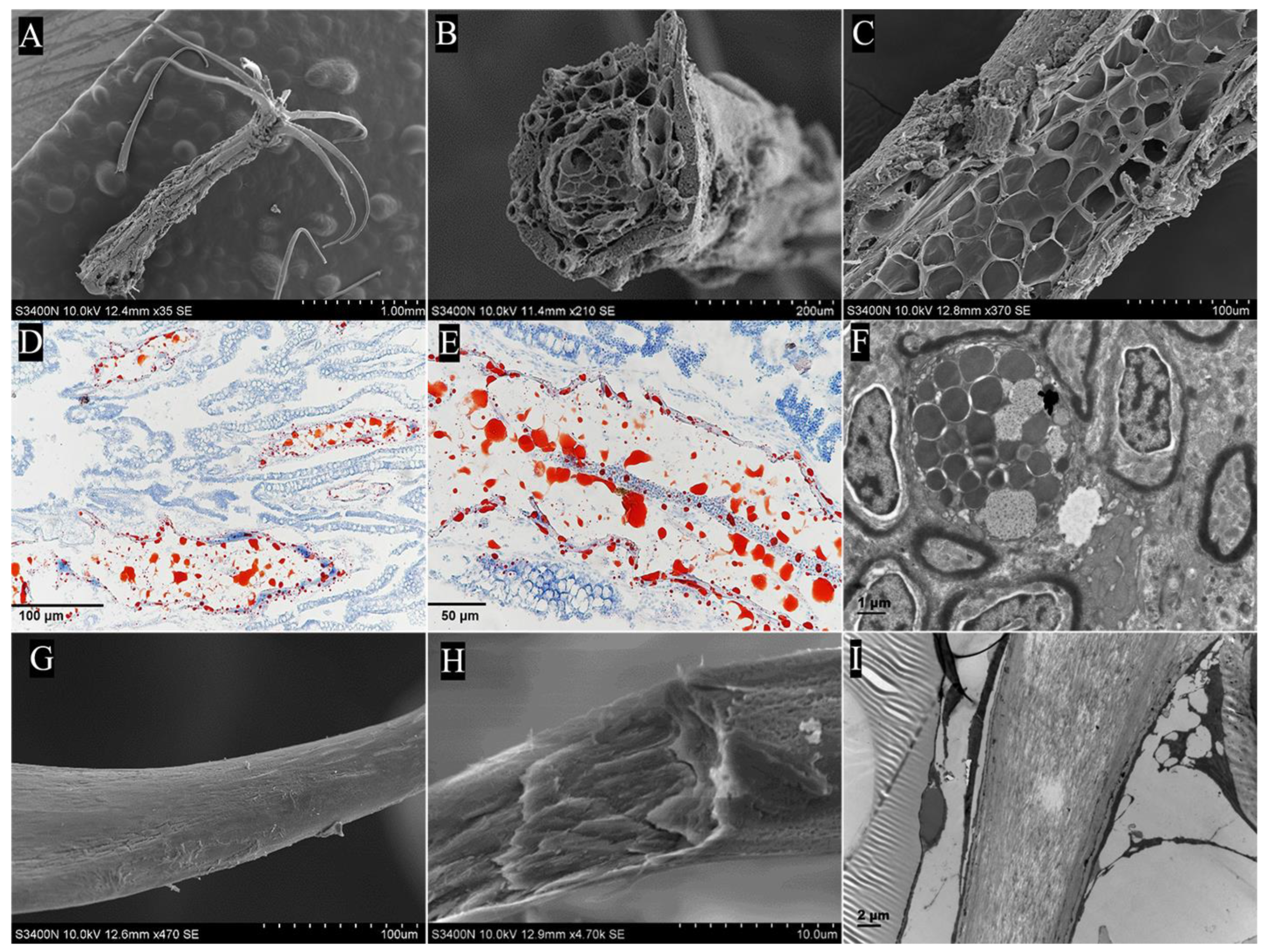
2.2. Analysis of Key Morphological Characteristics of the Pharyngeal Sac
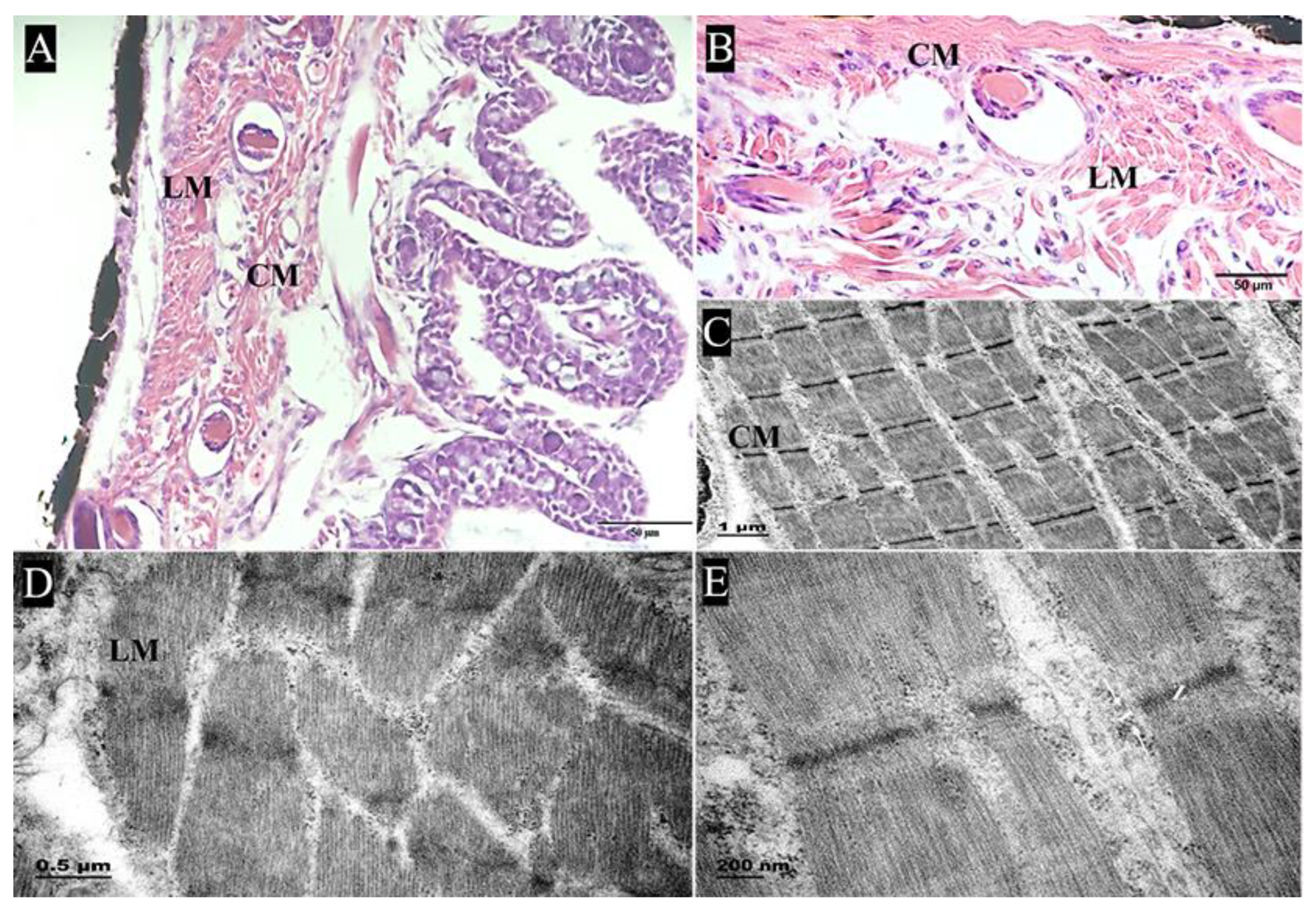
2.2.1. Muscularis
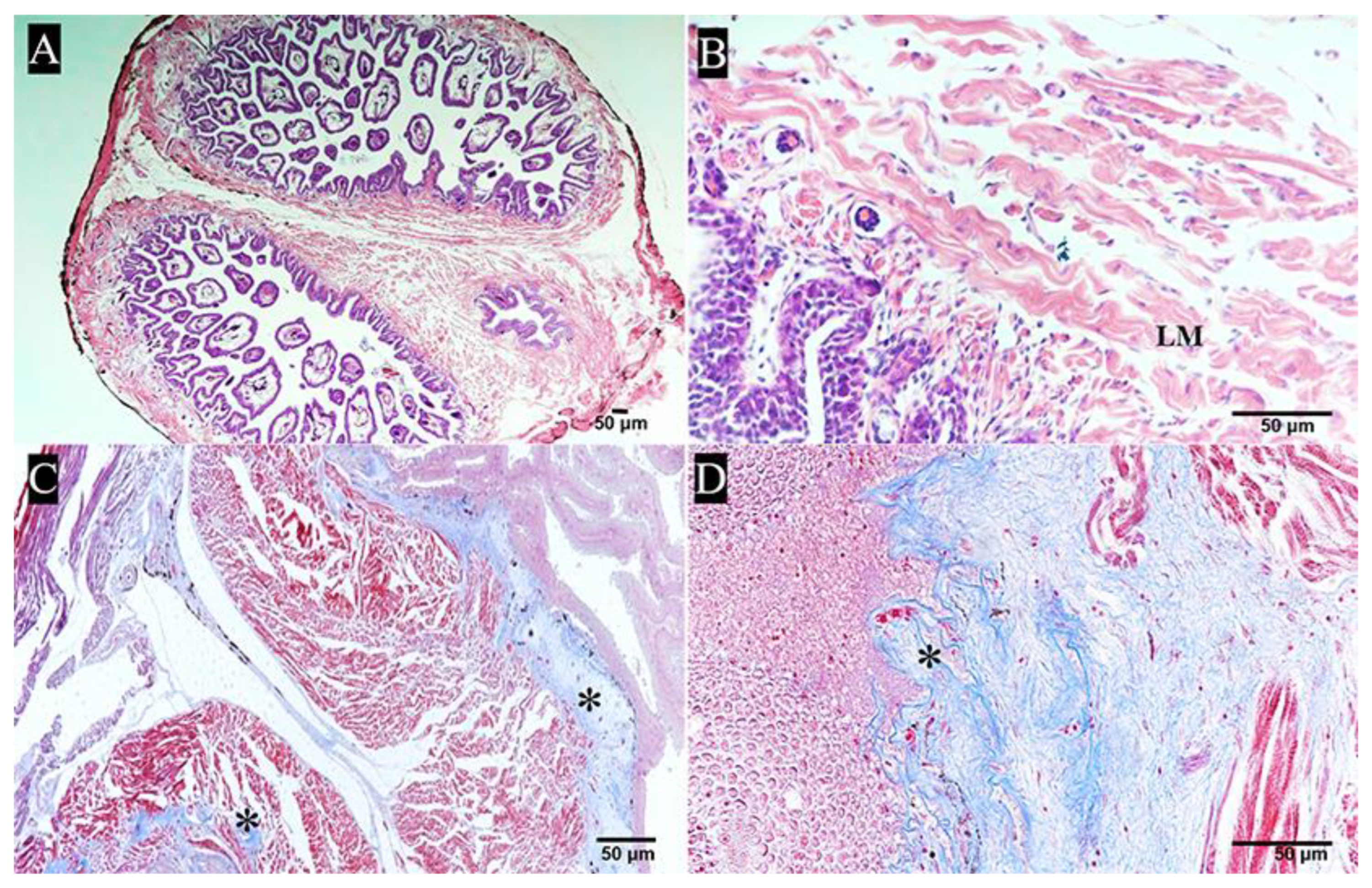
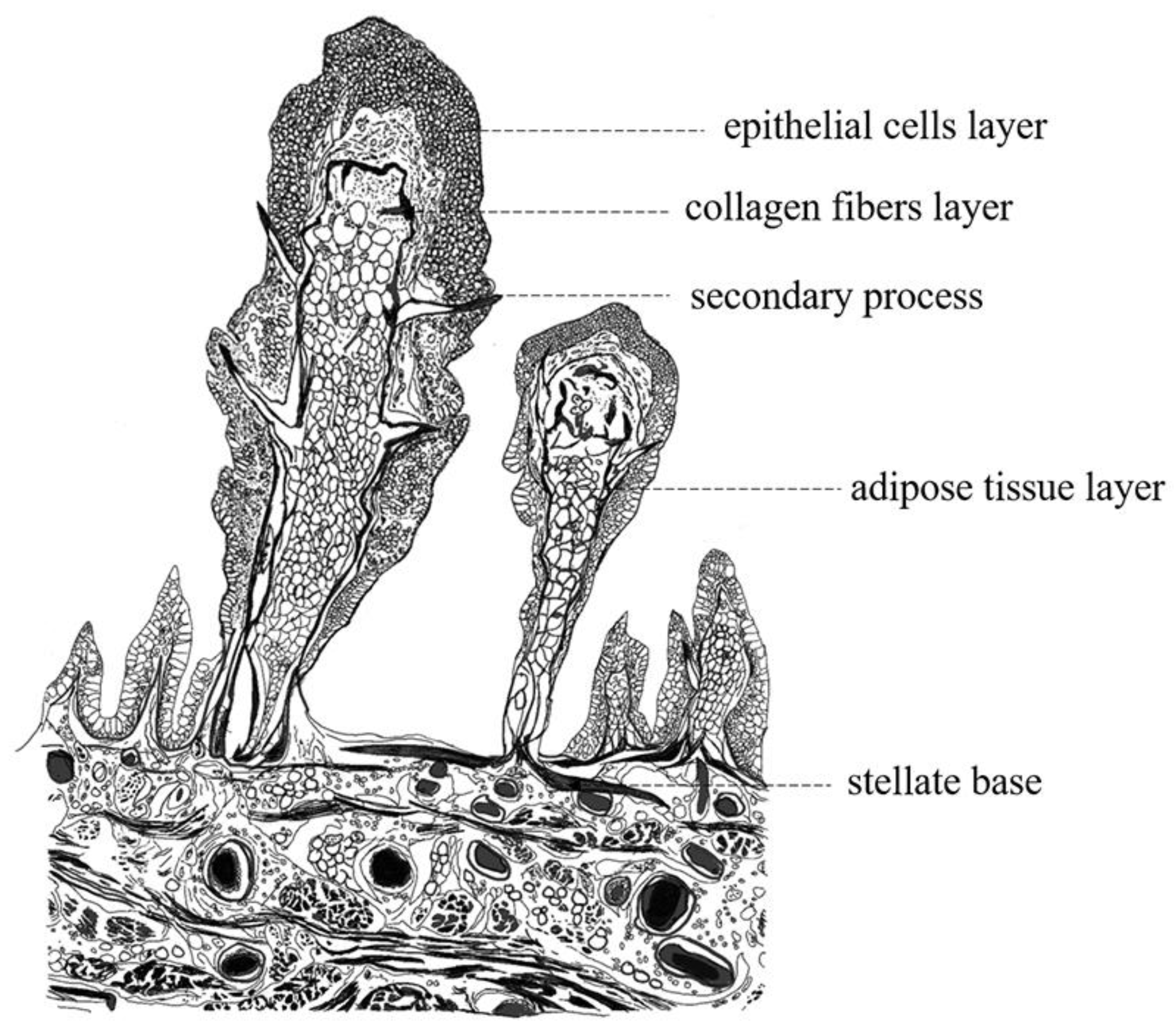
2.2.2. Papilla
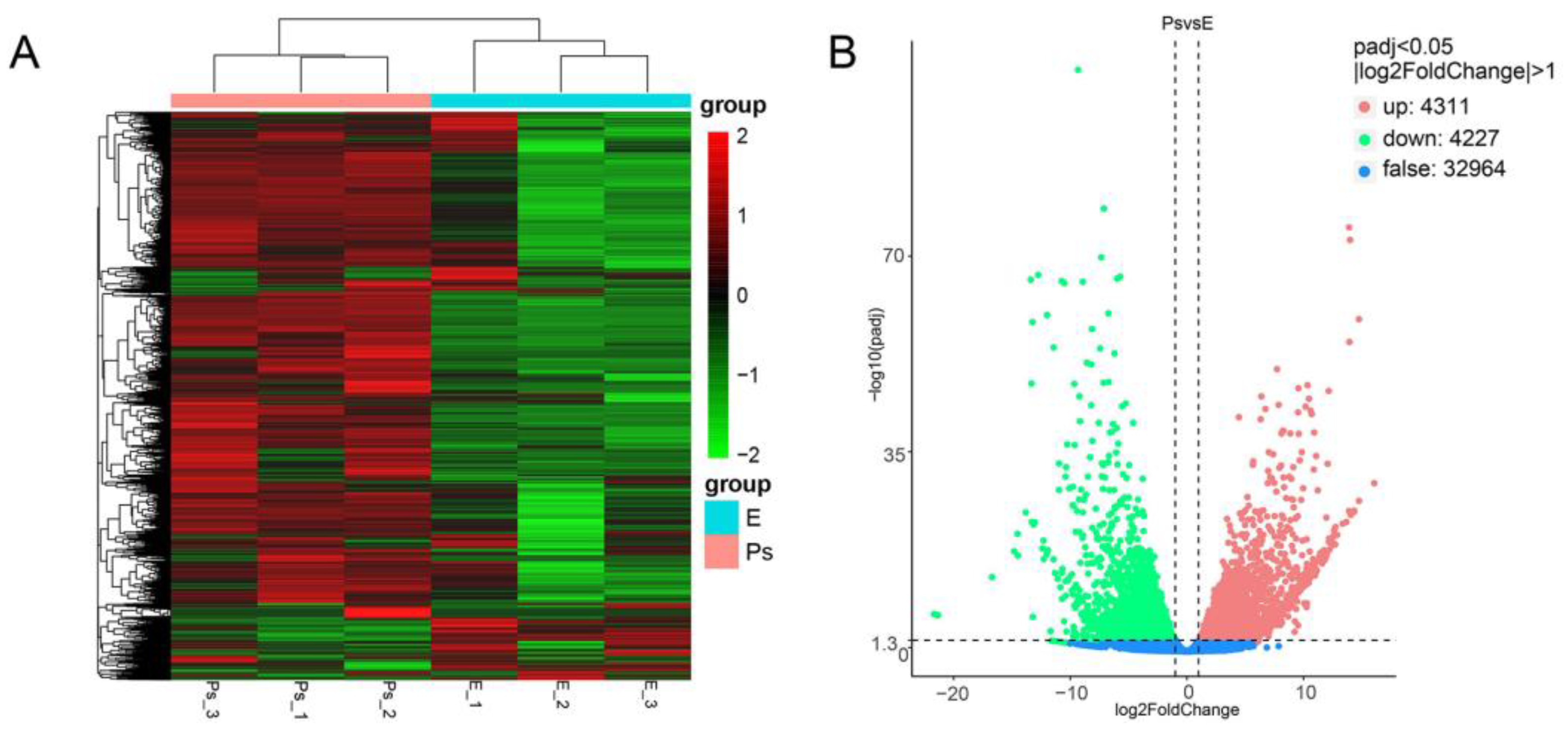
2.3. De Novo Assembly and Functional Annotation
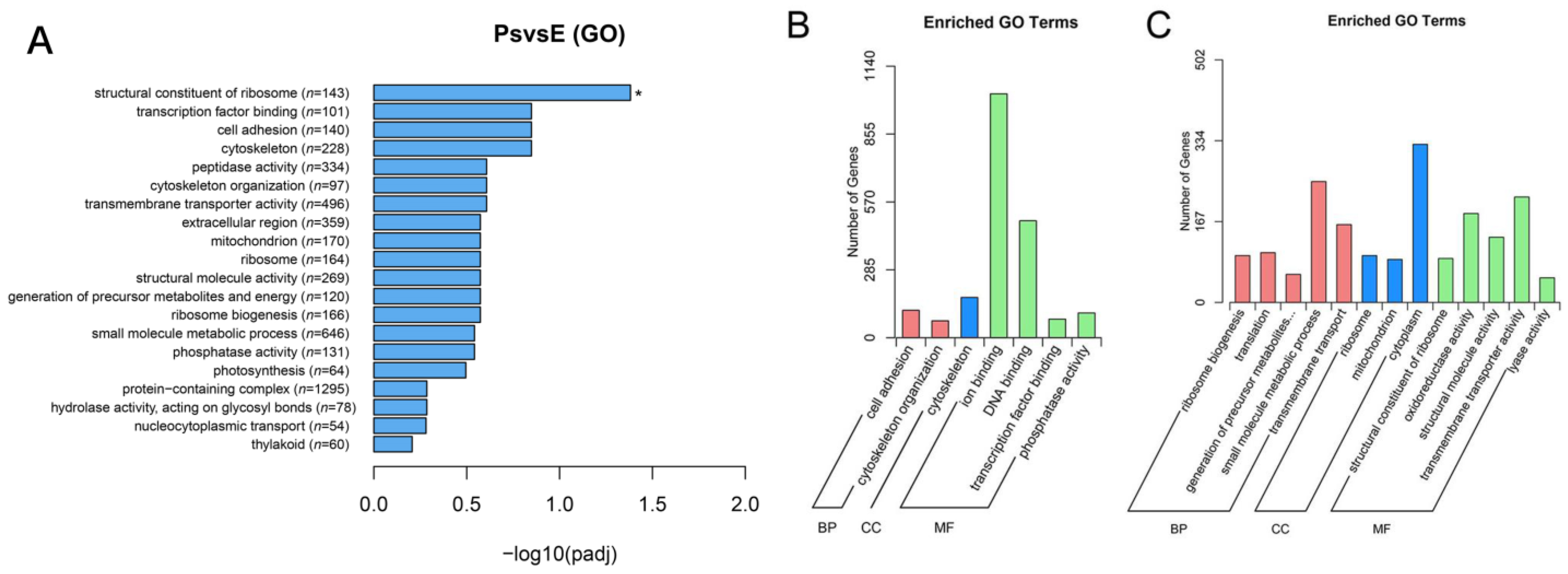
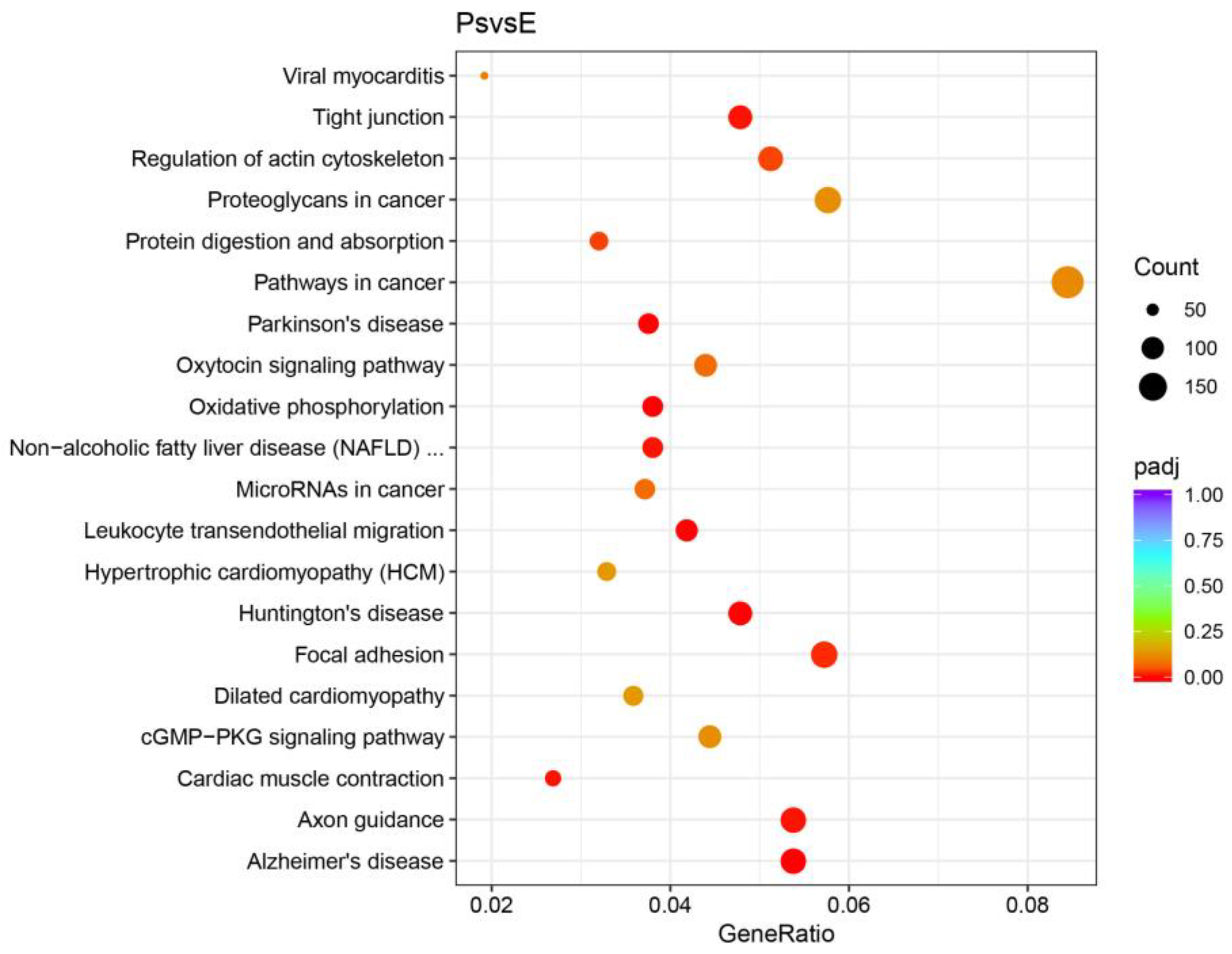
2.4. DEGs Potentially Correlated with Structure and Function of the Pharyngeal Sac
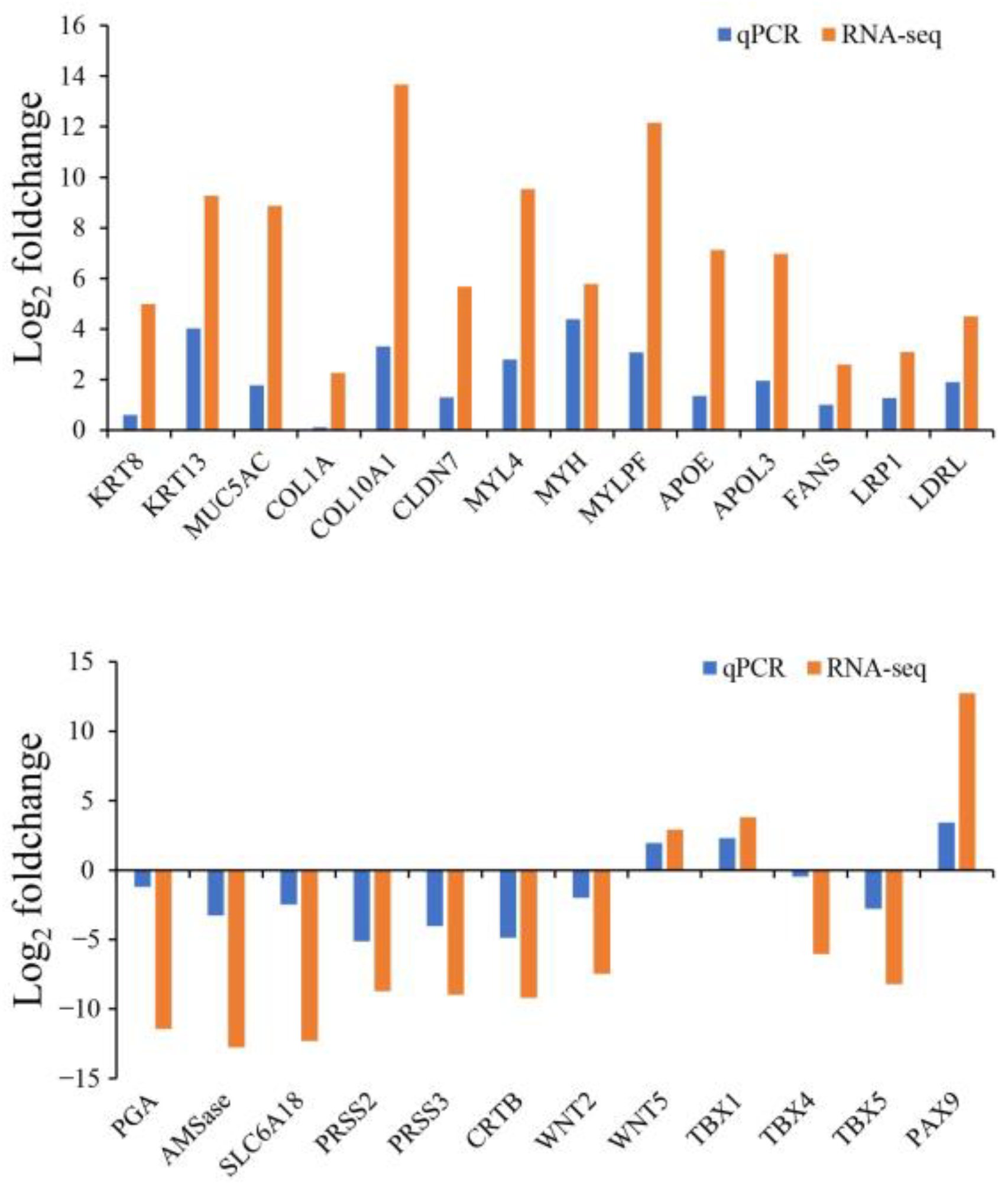
2.5. Validation of Gene Expression Levels
3. Discussion
3.1. Morphology of the Pharyngeal Sac
3.2. Candidate Genes Determined from Correlation between Structure and Function
4. Materials and Methods
4.1. Experimental Fish and Sampling
4.2. Light Miroscopy
4.3. Electron Microscopy
4.4. RNA Isolation, Library Preparation and Sequencing
4.5. De Novo Assembly, Unigene Annotation and Functional Classification
4.6. Validation of Transcriptome Data by RT-qPCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verma, C.R.; Gorule, P.A.; Kumkar, P.; Kharat, S.S.; Gosavi, S.M. Morpho-histochemical adaptations of the digestive tract in Gangetic mud-eel Ophichthys cuchia (Hamilton 1822) support utilization of mud-dwelling prey. Acta Histochem. 2020, 122, 151602. [Google Scholar] [CrossRef]
- Wilson, J.M.; Castro, L.F.C. Morphological diversity of the gastrointestinal tract in fishes. Fish Physiol. 2010, 30, 1–55. [Google Scholar] [CrossRef]
- Fagundes, K.R.C.; Rotundo, M.M.; Mari, R.B. Morphological and histochemical characterization of the digestive tract of the puffer fish Sphoeroides testudineus (Linnaeus 1758) (Tetraodontiformes: Tetraodontidae). An. Acad. Bras. Cienc. 2016, 88, 1615–1624. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.Q.; Cao, X.J.; Tian, X.C. Transcriptomic analysis of compromise between air-breathing and nutrient uptake of posterior intestine in loach (Misgurnus anguillicaudatus), an air-breathing fish. Mar. Biotechnol. 2016, 18, 521–533. [Google Scholar] [CrossRef]
- Gosavi, S.M.; Verma, C.R.; Kharat, S.S.; Pise, M.; Kumkar, P. Structural adequacy of the digestive tract supports dual feeding habit in catfish Pachypterus khavalchor (Siluriformes: Horabagridae). Acta Histochem. 2019, 121, 437–449. [Google Scholar] [CrossRef]
- Haedrich, R.L. The stromateoid fishes: Systematics and a classification. Bull. Mus. Comp. Zool. 1967, 135, 31–139. [Google Scholar]
- Hermann, B. Die verdauungsorgane der stromateidae. Zoomorphology 1930, 19, 59–115. [Google Scholar] [CrossRef]
- Gilchrist, J.D.F. XXVII—Note on the œsophageal teeth of the Stromateidæ. J. Nat. Hist. 1922, 9, 249–255. [Google Scholar] [CrossRef]
- Sminkey, T.R. A Morphological Study of the Pharyngeal Sac of Two Species of Stromateid Fishes: Perrilus triacanthus and P paru. Master’s Thesis, Virginia Institute of Marine Science, Gloucester Point, VA, USA. [CrossRef]
- Al-Qattan, E.; Dadzie, S.; Abou-Seedo, F. Feeding structures in the silver pomfret Pampus argenteus (Euphrasen) as observed by scanning electron microscopy. Kuwait. Int. J. Eng. Sci. 2000, 27, 329–341. [Google Scholar]
- Isokawa, S.; Kubota, K.; Kosakai, T.; Satomura, I.; Tsubouchi, M.; Sera, A. Some contributions to study of esophageal sacs and teeth of fishes. J Nihon Univ Sch Dent. 1965, 7, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Li, C.; Li, X. Phylogeny and biogeography of Chinese pomfret fishes (Pisces: Stromateidae). Studia. Marina. Sinica. 2002, 5, 235–239. (In Chinese) [Google Scholar]
- Doiuchi, R.; Nakabo, T. Molecular phylogeny of the stromateoid fishes (teleostei: Perciformes) inferred from mitochondrial DNA sequences and compared with morphology-based hypotheses. Mol. Phylogenet. Evol. 2006, 39, 111–123. [Google Scholar] [CrossRef]
- Pastana, M.; Johnson, G.D.; Datovo, A. Comprehensive phenotypic phylogenetic analysis supports the monophyly of stromateiform fishes (Teleostei: Percomorphacea). J. Linn. Soc. London, Zool. 2021, 3, 841–963. [Google Scholar] [CrossRef]
- Gao, L.J.; Shi, Z.H.; Yan, Y. Histological studies on development of digestive system in larval Pampus argenteus. J. Fish Sci. China 2007, 14, 540–546. (In Chinese) [Google Scholar] [CrossRef]
- Xu, S.L.; Qiu, C.G.; Gu, J.W.; Wang, D.L. The difference in ultrastructure changes of esophageal sac and liver in normal and hunger juvenile of Pampus argenteus. Oceanol. Limnol. Sin. 2013, 44, 1016–1023. (In Chinese) [Google Scholar]
- Zhou, J.K.; Xu, S.L.; Qiu, C.G.; Fang, Z.B.; Chen, P. Distribution and comparison of digestive enzyme activity of Pampus argenteus in different digestive organs. J. Ningbo Univ. Nat. Sci. Eng. Ed. 2014, 27, 0001–0006. (In Chinese) [Google Scholar]
- Mohamadi Yalsuyi, A.; Forouhar Vajargah, M. Recent advance on aspect of fisheries: A review. J. Coast. Life Med. 2017, 5, 141–148. [Google Scholar] [CrossRef]
- Dadzie, B.S.; Abou-Seedo, F.; Al-Qattan, E. The food and feeding habits of the silver pomfret, Pampus argenteus (Euphrasen), in kuwait waters. J. Appl. Ichthyol. 2000, 16, 61–67. [Google Scholar] [CrossRef]
- Peng, S.M.; Shi, Z.H.; Yin, F.; Sun, P.; Wang, J.G. Feeding habits of silver pomfret (Pampus argenteus) in East China Sea based on stable isotope techniques. Chin. J. Ecol. 2011, 30, 1565–1569. (In Chinese) [Google Scholar]
- Wei, X.J.; Zhang, B.; Shan, X.J.; Yang, T.; Jin, X.R.; Ren, Y.P. Trophic levels and feeding habits of silver pomfret Pampus argenteus in the bohai sea. J. Fish Sci. China 2019, 26, 904–913. (In Chinese) [Google Scholar]
- Zhang, C.; Jacques, K.J.; Zhang, S.; Xu, S.L.; Wang, Y.J.; Wang, D.L. Analyses of growth performance and realized heritability of Pampus argenteus in a breeding program in China. Front. Mar. Sci. 2022, 9, 935924. [Google Scholar] [CrossRef]
- Díaz, A.O.; García, A.M.; Devincenti, C.V.; Goldemberg, A.L. Morphological and histochemical characterization of the mucosa of the digestive tract in Engraulis anchoita. Anat. Histol. Embryol. 2003, 32, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.J.; Wang, W.M. Histology and mucin histochemistry of the digestive tract of yellow catfish, Pelteobagrus fulvidraco. Anat. Histol. Embryol. 2009, 38, 254–261. [Google Scholar] [CrossRef]
- Canan, B.; do Nascimento, W.S.; da Silva, N.B.; Chellappa, S. Morphohistology of the digestive tract of the damsel fish Stegastes fuscus (Osteichthyes: Pomacentridae). Sci. World J. 2012, 787316, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ikpegbu, E.; Nlebedum, U.C.; Ibe, C.S. The histology and mucin histochemistry of the farmed juvenile african catfish digestive tract (Clarias gariepinus b). Stud. Univ. Vasile Goldiş Ser. Ştiinţele Vieţii 2014, 24, 125–131. [Google Scholar]
- Qian, X.; Ba, Y.; Zhuang, Q.F.; Zhong, G.F. RNA-Seq technology and its application in fish transcriptomics. OMICS 2014, 18, 98–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.X.; Wang, S.N.; Hu, Z.W.; Yu, W.C.; Chou, X.M. Comparative analysis of intestine and stomach transcriptomes in cultured Takifugu rubripes. J. Zhejiang Ocean. Univ. Nat. Sci. 2021, 40, 384–391. [Google Scholar]
- Yasuike, M.; Iwasaki, Y.; Nishiki, I.; Nakamura, Y.; Matsuura, A.; Yoshida, K.; Noda, T.; Andoh, T.; Fujiwara, A. The yellowtail (Seriola quinqueradiata) genome and transcriptome atlas of the digestive tract. DNA Res. 2018, 25, 547–560. [Google Scholar] [CrossRef]
- Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Gene expression profiling reveals functional specialization along the intestinal tract of a carnivorous teleostean fish (Dicentrarchus labrax). Front. Physiol. 2016, 7, 359. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, M.L.; Arantes, F.P.; Santiago, K.B.; Ciências, J.E.D. Morphological characteristics of the digestive tract of Schizodon knerii (Steindachner, 1875), (characiformes: Anostomidae): An anatomical, histological and histochemical study. An. Acad. Bras. Cienc. 2015, 87, 867–878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matheus, V.A.; Faccioli, C.K.; Chedid, R.A.; Senhorini, J.A.; Franceschini-Vicentini, I.B.; Vicentini, C.A. Morphological and histochemical features of the digestive tract of Leiarius marmoratus (Gill, 1870). J. Fish Biol. 2021, 99, 1622–1631. [Google Scholar] [CrossRef] [PubMed]
- Bannikov, A.F. Fossil stromateoid fishes of the families Nomeidae and Ariommidae (Perciformes). Paleontol. J. 1993, 27, 108–119. [Google Scholar]
- Tang, D.J.; Liu, Y.; Zheng, X.B.; Ni, J.J.; Hou, C.C.; Wang, Y.J.; Xu, S.; Zhang, C.; Jin, S.; Xu, W.; et al. Morphology, histological structure characteristics and digestive enzyme activities of the digestive tract of Pampus argenteus. Acta Hydrobiol. Sin. 2022, 46, 643–653. (In Chinese) [Google Scholar] [CrossRef]
- Howe, J.C. A comparative analysis of the feeding apparatus in pomacanthids, with special emphasis of oesophageal papillae in Genicanthus personatus. J. Fish Biol. 1993, 43, 593–602. [Google Scholar] [CrossRef]
- Jeffrey, M.L. Larval development in the lutjanid subfamily lutjaninae (pisces): The Genus Macolor. Rec. Aust. Mus. 2007, 59, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Heiss, E.; Natchev, N.; Beisser, C.; Lemell, P.; Weisgram, J. The fish in the turtle: On the functionality of the oropharynx in the common musk turtle Sternotherus odoratus (chelonia, kinosternidae) concerning feeding and underwater respiration. Anat. Rec. 2010, 293, 1416–1424. [Google Scholar] [CrossRef]
- Pati, S. Food and feeding habits of silver pomfret Pampus argenteus (Euphrasen) from bay of bengal with a note on its significance in fishery. Indian J. Fish. 2011, 27, 244–256. [Google Scholar]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Du, J.G.; Lam, S.H.; Mathavan, S.; Matsudaira, P.; Gong, Z.Y. Morphological and molecular evidence for functional organization along the rostrocaudal axis of the adult zebrafish intestine. BMC Genom. 2010, 11, 392. [Google Scholar] [CrossRef] [Green Version]
- Bragulla, H.H.; Homberger, D.G. Structure and functions of keratin proteins in simple, stratified, keratinized and cornified epithelia. J. Anat. 2010, 214, 516–559. [Google Scholar] [CrossRef]
- Kopan, R.; Fuchs, E. A new look into an old problem: Keratins as tools to investigate determination, morphogenesis, and differentiation in skin. Genes Dev. 1989, 3, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Waseem, A.; Alam, Y.; Dogan, B.; White, K.N.; Leigh, I.M.; Waseem, N.H. Isolation, sequence and expression of the gene encoding human keratin 13. Gene 1998, 215, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Z.; Xu, Y.; Xie, H.Q.; Li, X.Q.; Wei, Y.Q.; Yang, Z.M. The possible role of myosin light chain in myoblast proliferation. Biol. Res. 2009, 42, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S. Muscle fiber type diversity revealed by anti-myosin heavy chain antibodies. FEBS J. 2018, 285, 3688–3694. [Google Scholar] [CrossRef]
- Deguara, S.; Jauncey, K.; Agius, C. Enzyme activities and pH variations in the digestive tract of gilthead sea bream. J. Fish Biol. 2010, 62, 1033–1043. [Google Scholar] [CrossRef] [Green Version]
- Dam, C.T.M.; Ventura, T.; Booth, M.; Pirozzi, I.; Salini, M.; Smullen, R.; Elizur, A. Intestinal transcriptome analysis highlights key differentially expressed genes involved in nutrient metabolism and digestion in yellowtail kingfish (Seriola lalandi) fed terrestrial animal and plant proteins. Genes 2020, 11, 621. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.X.; Xiao, Y.P.; Zhang, C.J.; Min, M.H.; Peng, S.M.; Shi, Z.H. Molecular characterization and expression analysis of toll-like receptor 2 in response to bacteria in silvery pomfret intestinal epithelial cells. Fish Shellfish Immunol. 2016, 58, 1–9. [Google Scholar] [CrossRef]
- Wu, C.X.; Deng, H.; Li, D.M.; Fan, L.H.; Yao, D.; Zhi, X.P.; Mao, H.; Hu, C. Ctenopharyngodon idella tollip regulates myd88-induced nf-κb activation. Dev. Comp. Immunol. 2021, 123, 104162. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, J.B.; Zhu, J.J.; Wang, Y.J.; Zhang, Y.Y.; Li, Y.Y.; Xu, S.; Yan, X.; Zhang, D. Transcriptome, antioxidant enzymes and histological analysis reveal molecular mechanisms responsive to long-term cold stress in silver pomfret (Pampus argenteus). Fish Shellfish Immunol. 2022, 121, 351–361. [Google Scholar] [CrossRef]
- Shi, P.; Meng, R.; Liao, K.; Li, S.; Hu, J.; Xu, J.; Zhang, L.; Cao, J.; Ran, Z.; Wang, D.; et al. Cadmium transcriptionally regulates Scd1 expression in silver pomfret. Environ. Toxicol. 2020, 35, 404–413. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Database | Number of Unigenes | Percentage (%) |
|---|---|---|
| NR | 23,111 | 55.44 |
| NT | 33,654 | 80.73 |
| KO | 14,746 | 35.37 |
| SwissProt | 20,189 | 48.43 |
| PFAM | 18,967 | 45.5 |
| GO | 18,960 | 45.48 |
| KOG | 10,032 | 24.06 |
| Annotated in all databases | 6862 | 16.46 |
| Annotated in at least one database | 34,762 | 83.39 |
| Total unigenes | 41,683 | 100 |
| Gene | Gene ID | Primer | Tm (°C) | Primer Efficiency (%) | Product (bp) | Melt Temp. |
|---|---|---|---|---|---|---|
| KRT8 | Cluster-5717.6135 | F: GATCTGCCGGCTGTAAATGG | 58.7 | 95.2 | 174 | 83.69 |
| R: GTCCTCAAGAATCAGCAGCG | 59.2 | |||||
| KRT13 | Cluster-5717.19474 | F: ACTTCCTGGAGAGCAAGACC | 58.3 | 99.4 | 110 | 85.17 |
| R: GGCAGCATTTCCACGAGTAG | 59.4 | |||||
| MUC5AC | Cluster-5717.4784 | F: TGTGAATGTGTGTGCAGTGG | 59.1 | 101.2 | 157 | 81.35 |
| R: AAGGATCACAGTCATGGGCA | 59.7 | |||||
| COL1A | Cluster-5717.14663 | F: GTCCTCTCTCACCAGGCTTT | 58.5 | 85.4 | 189 | 90.3 |
| R: CAAAGGGTATGACTGGCAGC | 58.8 | |||||
| COL10A1b | Cluster-5717.4734 | F: TGCAGCCCCTATTCAGTTCA | 59.6 | 89.7 | 152 | 83.44 |
| R: TGTAGAGAGCCACCAATGCA | 59.0 | |||||
| CLDN7 | Cluster-5717.4897 | F: GCATTCTTGGATCGTTGGCT | 58.4 | 104.6 | 163 | 86.48 |
| R: CAATACCAAGAGCAGGCGAC | 59.0 | |||||
| MYL4 | Cluster-5717.4933 | F: CACGGGAGATATGTTGCAGC | 58.5 | 96.3 | 196 | 85.41 |
| R: ACCTTGTTTGATCGCACACC | 59.6 | |||||
| MYH | Cluster-5717.4889 | F: GGCAGCTTTTGGTTTCTGGA | 59.7 | 95.7 | 196 | 84.64 |
| R: GGGGATTGAGTGGGAGTTCA | 59.1 | |||||
| MYLPF | Cluster-5717.4958 | F: AGGCGACCCACATGTTCTTA | 58.9 | 93.6 | 222 | 84.69 |
| R: CCATGATCAAGGAAGCCAGC | 59.0 | |||||
| APOE | Cluster-5717.9257 | F: ATGCGGAAGGGAGAACTCAA | 58.3 | 97.4 | 255 | 82.55 |
| R: TCAAGGAGACCACCACTGTC | 59.4 | |||||
| APOL3 | Cluster-5717.4775 | F: ACTGTCTCCGTTCACTCTGG | 59.1 | 87.5 | 170 | 87.83 |
| R: TGAATCTCAAGCAGGCCTGA | 58.6 | |||||
| FANS | Cluster-5717.20642 | F: TTGTTGACCGGCCTATTCCC | 58.7 | 102.1 | 154 | 85.99 |
| R: GGATGCGCCCTAGTCTTCTC | 58.9 | |||||
| LRP1 | Cluster-5717.15637 | F: CGGGCTGACTGTGTTTGAAA | 59.3 | 86.7 | 199 | 87.02 |
| R: CACCTCACATGCATGGCTAC | 59.7 | |||||
| LDRL | Cluster-5717.25245 | F: TGTGAATTCTTGTGCCTGGC | 59.0 | 86.4 | 171 | 85.87 |
| R: ATGAGGTGGGGTGGTAGTTG | 58.5 | |||||
| PGA | Cluster-5717.13230 | F: TGGAGGACTGCTTAGGGTTG | 58.9 | 79.4 | 250 | 86.32 |
| R: AAAGGATTGTGGGAGGCGTA | 59.3 | |||||
| AMSase | Cluster-5717.13038 | F: AACCGAGTCATCCACCCTTT | 58.6 | 98.6 | 158 | 83.54 |
| R: TGTGTCCCTTCTGCCTGATT | 59.2 | |||||
| SLC6A18 | Cluster-5717.11875 | F: GTGGGCAGTGCTCTTCTTTC | 58.3 | 97.5 | 152 | 85.29 |
| R: AGAGCCCAACGTGAAGATGA | 59.0 | |||||
| PRSS2 | Cluster-5717.10919 | F: GGTGGTCCTCTGGTGTGTAA | 58.8 | 93.5 | 226 | 86.61 |
| R: ACCTTCTCCTCCTGTCTCCT | 59.3 | |||||
| PRSS3 | Cluster-5717.13926 | F: GCAGCAACTATCCTGATCGC | 58.9 | 103.0 | 233 | 88.59 |
| R: GACTCCAGGCCTGTTCTTCT | 59.2 | |||||
| CRTB | Cluster-5717.13085 | F: GCACGGACAGCTTAATGAGG | 58.7 | 97.3 | 196 | 86.84 |
| R: ATCAACGAGAACTGGGTGGT | 59.0 | |||||
| WNT2 | Cluster-5717.15143 | F: TTTGTCCGAAAGTGCTCTGC | 58.6 | 87.5 | 159 | 83.9 |
| R: TCCCAGCAGTTCAGTCTCAG | 59.3 | |||||
| WNT5 | Cluster-5717.10855 | F: CCTTTGGCAGACTGTTGGAC | 58.9 | 96.4 | 239 | 75.08 |
| R: ACCTTCACCGATGTACTGCA | 59.2 | |||||
| TBX1 | Cluster-5717.8690 | F: CCCCACCACCTATGAACACT | 58.7 | 85.6 | 238 | 86.53 |
| R: CAGGACTGTGTTGCCTTTCC | 59.1 | |||||
| TBX4 | Cluster-5717.11891 | F: ACTATGCGAGGAGATGAGCC | 58.7 | 89.7 | 232 | 83.41 |
| R: TGGATCCAAGAACACTGCCT | 59.3 | |||||
| TBX5 | Cluster-5717.17018 | F: TCCGTCAGAGAGTGTGTTCC | 58.9 | 87.9 | 218 | 90.34 |
| R: CCTCTTGGTGCAGTGGTAGA | 59.0 | |||||
| PAX9 | Cluster-5717.24078 | F: GTCTACCCCTACAACCCCAC | 58.7 | 78.5 | 169 | 83.83 |
| R: TCATGTTGGTCCAGTCCTCC | 59.4 | |||||
| β-acting | [51] | F: ACCCTGAAGTACCCCATCGA | 59.1 | 98.5 | 157 | 83.58 |
| R: GGCCACTCTCAGCTCATTGT | 58.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, H.; Hu, J.; Xie, H.; Zhang, M.; Guo, C.; Zhang, Y.; Li, Y.; Zhang, C.; Xu, S.; Wang, D.; et al. Morphological and Molecular Functional Evidence of the Pharyngeal Sac in the Digestive Tract of Silver Pomfret, Pampus argenteus. Int. J. Mol. Sci. 2023, 24, 1663. https://doi.org/10.3390/ijms24021663
Jiang H, Hu J, Xie H, Zhang M, Guo C, Zhang Y, Li Y, Zhang C, Xu S, Wang D, et al. Morphological and Molecular Functional Evidence of the Pharyngeal Sac in the Digestive Tract of Silver Pomfret, Pampus argenteus. International Journal of Molecular Sciences. 2023; 24(2):1663. https://doi.org/10.3390/ijms24021663
Chicago/Turabian StyleJiang, Huan, Jiabao Hu, Huihui Xie, Man Zhang, Chunyang Guo, Youyi Zhang, Yaya Li, Cheng Zhang, Shanliang Xu, Danli Wang, and et al. 2023. "Morphological and Molecular Functional Evidence of the Pharyngeal Sac in the Digestive Tract of Silver Pomfret, Pampus argenteus" International Journal of Molecular Sciences 24, no. 2: 1663. https://doi.org/10.3390/ijms24021663